中 國 醫 藥 大 學
醫學檢驗生物技術學系碩士班 碩士論文
Stenotrophomonas maltophilia 誘發 L1 及 L2 乙內醯胺酶基因表現之探討
Induction studies of L1 and L2 β-lactamases in Stenotrophomonas maltophilia
指導教授:楊翠青 副教授 Tsuey-Ching Yang 學生姓名:蕭盈如
Ying-Ju Hsiao 學生學號:9473005
中華民國 九十六 年 七 月 六 日
目 錄
致謝...i
中文摘要 ...ii
英文摘要 ... v
第一章 研究背景與研究動機 第一節 Stenotrophomonas maltophilia 介紹...1
第二節 Stenotrophomonas maltophilia 的抗藥性機轉...2
第三節 β-lactam 抗生素...3
第三節 β-lactamase 分類及命名...4
第四節 Stenotrophomonas maltophilia 之 L1 及 L2 基因...5
第五節 L1 及 L2 基因誘導分析...7
第六節 β-lactamase 抑制劑...7
第二章 研究目的...11
第三章 研究重要性...12
第四章 研究方法 第一節 研究材料 ...13
第二節 實驗方法 ...19
第五章 研究結果 ...29
第六章 結果討論...48
第七章 結論...60
參考文獻 ...61
圖 ...67
表 ...79
附錄一 ...88
附錄二 ...89
附錄三...90
附錄四...91
附錄五...92
致謝
論文完成了!在這兩年多的生活中,過的非常快樂及充實,雖然在求 知的過程中也有不愉快及沮喪的時候,但總覺得自己的決定是對的。首 先謝謝我敬愛的老師楊翠青博士,謝謝妳兩年多的來的細心指導,無論 是實驗上,報告或課業上,還是生活上的建議都令我收穫良多。感謝吳 禮字及胡若梅兩位老師抽空修改論文以及對實驗的寶貴建議。再謝謝郭 雅音老師對我的教誨與鼓勵。
謝謝實驗室的夥伴:善恩及冠勳學長、冠傑、詩雯、佳芳、素連、明 宏、婉麗、世娟及學弟妹們等。還有鼓勵我的好友:楹奇、小璦、羿謹、
永成等,在我難過沮喪時給我勇氣去面對挫折。真的很感謝我的阿嬤,
及父母親默默的為我付出。因為他們的鼓勵及容忍讓我曾想放棄的念頭 打消,然後繼續堅持下去。努力不見得可以成功,但不努力就更難成功 了。
每個人都有其優缺點,在人生過程中,讓我慢慢知道自己所擅長的與 不擅長的事物,謝謝自己能不斷給自己機會,我想,做自己擅長的事才 能有成就感及信心。我將帶著這些回憶,繼續人生的旅程。
中文摘要
Stenotrophomonas maltophilia 為多重抗藥性的細菌,其位於染色體上的 L1, L2 二基因可經由誘發(induction) 產生高活性 β-lactamase,可水解 β-lactam 類的抗生素。本實驗菌株來源為中國醫院大學附設醫院細菌檢 驗 室 所 提 供 臨 床 菌 株 ,S. maltophilia N 。 利 用 基 因 替 換 方 式 (Gene replacement mutagenesis )突變 N 菌的 L1, L2 二基因取得 NL1-及NL2- 之突變株,分別在L1,L2 二基因起始氨基酸 (Met)下游置入一個和 L1,
L2 二基因轉錄方向一致的報導基因, xlyE。因此可以觀察報導基因, xlyE 的表現來評估 L1,L2 二基因啟動子分別被誘發(induction)的能力。為了 評估L1 及 L2 基因是否會因為在培養基中加入 5%人類血清或因改變生長 環境中的碳源而誘發(induction)或影響二基因表現,我們在培養 NL1-及 NL2-突變株過程中加入5 %人類血清及在培養 NL1-及NL2-突變株過程中 分別加入 1%的碳源,不會對 L1 及 L2 基因產生誘發情形。為了評估 clavulanate 是否會誘發 S. maltophilia 的 Class B L1 β-lactamase, Class A
L2 β-lactamase 二基因表現而導致 clavulanate 和其他β-lactam 抗生素一 起治療時產生拮抗(antagonism)或協同(synergism)作用,因此分別測試 2、
8、10、16、50 μg/ml clavulanate 對 L1, L2 基因誘發的能力 ,分別得 clavulanate 對 L L2 二 基 因 表 現 的 誘 發 能 力 為 concentration
dependent。因此我們利用抗生素感受性的結果並分別測試在不同濃度的 clavulanate 及搭配不同β-lactams 抗生素而產生共同或拮抗誘發 L1, L2 二基因啟動子表現。得知NL1-在濃度為 Cab/CA(8/4) μg/ml ,對 L1 基因 有induction synergism 的情形。NL2-在濃度Azt /CA(32/16) μg/ml ,對 L2 基因有induction synergism 的情形發生。因 clavulanate/β-lactams 抗生素組 裝方式所引起的L1, L2 二基因 induction 表現,可提供在選擇治劑上會 影響療效的因素:1.選擇適當β-lactams 抗生素與 clavulanate 的組裝 2.選 擇治劑組裝的適當比例。
Abstract
Stenotrophomonas maltophilia has assumed an increasingly important role as a nosocomial pathogen in compromised patients.。S. maltophilia isolates are resistant to many clinically useful antibiotics,with β-lactam resistance mediated through two β-lactamases,L1 and L2,whose expression is induced when cells are exposed to β-lactam antibiotics。Little is known about the induction of L1 and L2 β-lactamases。A clinical S. maltophilia isolate, S.
maltophilia N, was obtained in China Medical University Hospital. The PCR amplicons containing L1 and L2 β-lactamase genes, respectively, were sequenced and analyzed. The L1- and L2- mutants of S. maltophilia N were constructed by gene replacement strategy, respectively. A reporter gene, xylE, is inserted into the L1 and L2 gene, down-strand the promoter of L1 and L2 gene and with the same orientation as that of L1 and L2 gene, respectively.
The induction of L1 and L2 gene promoters can be monitored by analyzing the catechol 2,3-dioxygenase (C23O) activities in different induced conditions.
The studies in the thesis were designed to elucidate the factors that affect the induction of L1 and L2 gene promoters. The induction potency of cefuroxime against NL1- and NL2- is independent on the type of carbon source in growth medium and the addition of 5% human serum. Clavulanic acid acts as an inducer toward the L1 and L2 gene promoters, and its induction potency against L1 and L2 genes is concentration dependent. The susceptibility test of S. maltophilia N against different combined regimens of β-lactams/clavulanate was evaluated.The factors affected the effect of combination regimens were elucidated using the isogeneic mutants, NL1- and
NL2-, as the assayed systems. To be a successful combined regimen, three important elements must be considered: (1) The hydrolytic ability of L1 toward the selected β-lactam. (2) The minimal concentration of clavulanate needed to inhibit the activity of induced L2 enzyme. (3) The occurrence of induction synergy by the combination of β-lactam and clavulanate.
第一章 研究背景與研究動機
第一節 Stenotrophomonas maltophilia 介紹
Stenotrophomonas maltophilia 早期稱 Pseudomonas maltophilia (Hugh &
Ryschenkow, 1961),後改名為 Xanthomonas maltophilia (Sutter, 1968),
最 近 這 幾 年 才 命 名 為 Stenotrophomonas maltophilia (Palleroni &
Bradbury,1993)。S. maltophilia 是屬於非發酵性、好氧的革蘭氏陰性桿 菌,嗜以麥芽糖為碳源,Stenotrophomonasmaltophilia 可以耐 silverlined catheters ( Ubeda et al., 1998) 且可生長於4-40℃,最適合的生長溫度為 35℃。S. maltophilia 在健康人的身上並非致病菌,但常在醫院內病人的 檢體中分離出來,和許多的不同的感染症狀都有相關性,但卻沒有明顯 臨床上的特性。S. maltophilia 常藉由許多媒介物,例如:醫療器材;治 療用的各種插管,或使用不當的隱形眼鏡,甚至不通氣的空調系統,皆 會引起免疫力缺陷及因器官移植而使用了免疫抑制藥物的病患感染 (Denton & Kerr, 1998),甚至造成院內感染。此菌與呼吸道的感染也有 關 , 最 後 可 能 會 導 致 菌 血 症 , 心 內 膜 炎 , 及 尿 道 感 染 (Maningo &
Watanakunakorn,1995),在一些觀察中認為此菌的胞外蛋白分解酵素可能 會導致感染的組織受損,是重要的一種致病因子(Windhorst et al., 2002)。目前有兩株 S. maltophilia 的菌株其基因體定序工作已完成。一
株 為 S. maltophilia R551-3 (http://www.ncbi.nlm.nih.gov/) , S. maltophilia R551-3 的 GC content平均為 66%,序列全長為 4,544,233 nt;另一株為 S. maltophilia K279a (http://www.sanger.ac.uk/)。S. maltophilia K279a 的 GC content平均為 66.32%,序列全長為 4,851,126 bp。
第二節Stenotrophomonas maltophilia 的抗藥機轉
革蘭氏陰性(Gram-negative) 菌的菌株有四種抗藥機制。一、細菌產生可 分解或抑制抗生素活性之酵素。如很多細菌會產生 β-lactamase 的酵 素,破壞β-lactam 類抗生素的環狀構造,而對 β-lactam 型的抗生素產 生抗藥性(Rkihmod etal., 1973)。二、變更抗生素所辨認的受器目標。
例如 β-lactam 型的抗生素其殺菌的方式主要是藉由抗生素與細菌之 penicillin binding proteins (PBPs) 結合,而抑制細菌細胞壁的合成,進 而達到殺菌的效果。菌可改變 PBPs (penicillin binding proteins),而使 得抗生素無法和 PBPs 結合,而產生抗藥性(Livermore et al., 1995)。
三、改變細胞膜的通透性,使抗生素難以進入到細胞膜內部,格蘭氏陰 性菌可改變細胞膜的 porin ,使抗生素無法順利進入細胞內部 (Krueger et al., 2001)。四、細菌細胞膜上有 efflux pump (multidrug pump) 的 結構,使抗生素一進入到細胞膜內部則馬上藉 pump 打到細胞膜外部
((Li et al., 2002; Sanchez et al., 2002; Zhang et al., 2001)。根據 研 究 報 導 指 出S. maltophilia 對 於 一 些 藥 物 , 如 ::aminoglycosides, quinolones,β-lactams, chloramphenicol, rifampin, tetracycline, trimethoprim等,皆會產生抗藥性(Avison et al., 2001)。目前已知 S. maltophilia 基因體上存有數個抗藥性基因,可產生對抗抗生素的酵素:
β-lactamase(Walsh et al., 1997) , aminoglycoside acetyltransferases (Lambert et al., 1999),及 erythromycin inactivating enzyme (Alonso et al., 2000)。其中L1、L2 β-lactamase 的表現及 efflux system (SmeABC and SmeDEF)的機制與S. maltophilia 的多重抗藥性有密切的關係(Gould et al., 2006)。
第三節 β-lactam 抗生素
β-lactam為全世界廣泛應用於醫療上的抗生素。β-lactam抗生素包括四大 類:penicillins、cephalosporins、carbapenem、monobactam Penicillins類 的抗生素包括有:benzylpenicillin (penicillin G),oxacillin,ampicillin , piperacillin , carbenicillin… 等 。 Cephalosporins 類 的 抗 生 素 包 括 有 cephaloridine , cefuroxime , cefoxitin , ceftriaxone , cefotaxime , cefoperazone,cefepime…等。Carbapenem類的抗生素包括有imipenem,
meropenem …等。而aztreonam 則屬於monobactam類的抗生素。此四類
β-lactam抗生素都有一共同的β-lactam ring結構。
第四節 β-lactamase的分類及命名
對於β-lactamase 分類的方式繁多,目前學者常以 Bush 所提出的分類方式 將β-lactamase 區分為四大類(Bush & Sykes, 1986)。主要依β-lactamase
的功能作為分類的標準:包括受質的種類及其活性是否可被抑制物抑制 而 分 為 四 大 類 。 第 一 類 (Group 1) 酵 素 受 質 為 cephaloridine 及 cephalothin ,不受 clavulanic acid 抑制,第二類 (Group 2) 酵素均可被 clavulanic acid 抑制,又依其受質種類不同而分為五個次分型,依序從 2a 到2e: Group 2a 是屬於 penicillinases 但會被 clavulanic acid 抑制,Group 2b 是 屬 於 broad-spectrum β-lactamases 其 受 質 為 penicillin 及 cephalosporins 類抗生素但會被 clavulanic acid 抑制,Group 2c 是屬於 carbenicillin-hydrolyzing β-lactamases 其受質為 cephalosporins,也會被 clavulanic acid 抑制,Group 2d 是屬於 cloxacillin-hydrolyzing β-lactamases 其 受 質 為 cloxacillin, 也會被 clavulanic acid 抑制,Group 2e 是 屬 cephalosporin-hydrolyzing β-lactamases 其受質為 cephalosporin 也會被 clavulanic acid 抑制。第三類 (Group 3) 酵素為含有鋅的酵素,又稱為 金屬酵素(metallo-enzyme),不受 clavulanic acid 抑制,受 EDTA 所抑 制其酵素受質為 imipenem。第四類 (Group 4) 是一群不受 clavulanic
acid 抑制的 penicillin-hydrolyzing β-lactamases ,其特性目前仍在研究中,
尚未定論。除了Bush 的分類方式外,Ambler 提出依酵素胺基酸序列分類 的方式 (Ambler et al., 1980),大致可將 β-lactamase 分為 A,B,C,D 四類:
A 、 C 、 D 是 屬 於 以 serine 作 為 催 化 機 制 的 酵 素 (serine active-site enzyme),皆有 S-X-X-K motif 結構,而胺基酸 Ser 是 active site residue。
B 是屬於含鋅的酵素,受 EDTA 所抑制(Ambler et al.,1980)。整理於附錄 (一)
第五節 S. maltophilia 之 L1 及 L2 基因
位於S. maltophilia染色體上含有兩個可以 encode出β-lactamase的 L1 及 L2 基因。不論是 L1 或 L2 β-lactamases,其 N 端包含大約20個胺基酸 的訊號胜肽鍊(signal peptide) (Walsh et al., 1997),在細胞質產生的 β-lactamase是前趨物,最後成熟的β-lactamase會被送到細胞膜的外膜與 內膜間,在通過細胞內膜時signal peptide會被切除,signal peptide具有一 個hydrophobic core region 以 利 將 secreted polypeptide 插 入 內 膜 , 在 hydrophobic core region前N-terminal帶有正電荷胺基酸,會與細胞膜上帶 有負電荷的phospholipids產生吸引,而core region後的C-terminal則會有帶 極性的胺基酸,這時帶正電荷的胺基酸N端與hydrophobic core region會 形 成 具 有 功 能 性 的signal peptide 以 讓 signal peptidase 切 斷 (Izard &
Kendall, 1994) 。 根 據 報 導 知 道 L1 是 一 個 需 要 鋅 (Zn2+) 的 metallo-beta-lactamase,一個分子可以結合約2分子的Zn2+ ( Bush et al., 1995),依Bush的β-lactamases分類來說,L1是屬於Group 3,且會受EDTA的 抑制(Yush et al.,1982),在Ambler的β-lactamases分類是屬於Group B,而以 Bush-Jacoby-Medeiros分類是屬於Group 3a,(Bush et al., 1995)。L1可 水解的抗生素受質主要為penicillins 型的抗生素,部份cephalosporins 型 的抗生素及carbapenems型抗生素亦可被水解,但L1不能分解monobactam 型的抗生素。L1蛋白為homotetramer的結構,分子量約118 kDa,為neutral 或 acidic 的蛋白,S. maltophilia的L1的GC content平均68.4%左右,在臨 床分離菌株發現S. maltophilia的L1基因具有8~12%的變異性(Avison et al., 2001)。而L2是serine active-site cephalosporinase,在Ambler的β-lactamases 分類是屬於Group A,而以Bush-Jacoby-Medeiros分類是屬於Group 2e,會 受clavulanic acid,tazobactam,sulbactamn…等所抑制(Denton et al., 1998),
L2可水解的抗生素受質主要是cephalosporins類的抗生素、部分penicillins 型 的 抗 生 素 及monobactams , L2 不 能 水 解 cefoxitin 這 個 抗 生 素 及 carbapenem型的抗生素( Saino et al., 1984)。L2的蛋白為dimer的結構,分 子量約56 kDa,為basic的蛋白,GC content約為71.6%,具有S-T-D-K、
S-D-N、K-T-G的高度保留序列(Walsh, et al.,1997),在臨床分離菌株發 現S. maltophilia的L2基因具有9~25%的變異性(Avison, et al. 2001)。L1
與L2 蛋白整理於附錄四。
第六節 L1與L2基因誘導分析
在 Paton ,1994 報導研究中指出 S. maltophilia 的 L1 和 L2 基因是共 同被誘導產生β-lactamases。但是在 Avison 等人(Avison etal., 2002) 的 報 導 研 究 間 接 知 道 L1 和 L2 是 透 過 不 同 的 誘 導 機 制 產 生 β-lactamases。為了更進一步去了解 L1 與 L2 基因是被 β-lactam 抗生 素共同誘導(co-induction)或是個別被誘導(differential induction);在 本研究中,藉由不同的誘發條件,分別分析 L1 及 L2 基因被誘發的 程度,希望能對 L1 及 L2 基因的誘導機制有更清楚地瞭解。
第七節 ββ-lactamase 抑制劑
ββ-lactamase 抑 制 劑 有 clavulanic acid 、 penicillanic acid sulfones sulbactam 、EDTA、iodine、 YTR 830、7-acetylmethylenepenicillanic acid (AMPA)、或一些離子...等。這些 ββ-lactamase 抑制劑在抑制 ββ-lactamase的機制上不盡相同。如:EDTA可以和活化 ββ-lactamase 所 需之二價陽離子產生螯合作用,而使 β-lactamase 失活,如 Sβ . mlatophilia 的 L1 β-lactamase 即會受到 EDTA 而失去活性。文獻 指 出(Saino et al., 1982) 將 菌 株 S. maltophilia GN12873 的 L1
β-lactamase純化後,在50 mM phosphate buffer (pH 7.0)的環境下,將 L1 β-lactamase水解 100 μM penicillin G 的水解能力視為 100%;但 純化之L1 β-lactamase和 0.1 mM EDTA 作用後,其水解 100 μM penicillin G 的活性只剩約 3%,所以0.1 mM EDTA可抑制97% L1 β-lactamase的活性。文獻(Saino et al., 1984)報導中菌株 S. maltophilia GN12873 L2β-lactamase純化後, 在50 mM phosphate buffer (pH 7.0) 的環境下,將L2 β-lactamase水解100 μM cephaloridine 的活性視為
100%;L2 β-lactamase和 1 mM EDTA作用後,L2 β-lactamase 對100 μM cephaloridine 的水解活性約為原來的 96%,所以1 mM EDTA 可 抑制4% L2 β-lactamase的活性。此類型的抑制機制,EDTA 不會和
β-lactamase的受質產生競爭的效果。而另一種抑制 ββ β-lactamase的機 制如:clavulanic acid、sulbactam...等抑制劑,是在結構上相似於 β-lactam,因此會和ββ-lactamase的受質產生競爭的效果,而這類抑制 劑跟ββ-lactamase的親和性皆很好,因此容易結合在 β-lactamase的活 化位置上,進而使β-lactamase 酵素無法水解受質。在以往的文獻 (Saino et al., 1982),曾經評估 S. maltophilia GN12873 所純化之 L1 β-lactamase 受 clavulanic acid 及 sulbactam 抑制的情形。以 100 μM penicillin G 做為分析受質,clavulanic acid 及 sulbactam 對 L1 酵素之 Ki 值分別為 >100 μM 及 24.5 μM。所以對 L1 β-lactamase
而言,clavulanic acid 及 sulbactam 皆不是很好的抑制物。而在L2 β-lactamase 部 份 , (Saino et al., 1984) 曾 經 評 估 S. maltophilia
GN12873 所純化之L2 β-lactamase 受 clavulanic acid 及 sulbactam 抑制的情形。以100 μM cephaloridine做為分析受質,clavulanic acid 及 sulbactam 對 L2 酵素之 Ki 值分別為 0.58 μM 及1.87 μM。所以 對 L2 β-lactamase 而言,clavulanic acid 及 sulbactam 皆是好的抑
制物。臨床上有些 ββ-lactamase 抑制劑常會和倂 β-lactam型的抗生素 作為用藥,例如:ticarcillin/ clavulanic acid、amoxicillin/clavulanic acid、ampicillin/sulbactam、aztreonam/ clavulanic acid..等。這樣的合 併 使 用 常 用 於 會 產 生 β-lactamase 的 菌 株 。 如 : Pseudomonas β aeruginosa 、Bacteroides fragilis、 Citrobacter diversus...等。關於 Stenotrophomonas maltophilia對於合併藥物治療在文獻中有報導(Arpi et al., 1996) 作者收集124個臨床菌株,從agar dilution的方法發現124 個臨床菌株對藥物比例為 aztreonam/ clavulanic acid (1/1) 呈現85%感 受性;trimethoprim-sulfamethoxazole (1/19)呈100%藥物感受性;對 amoxicillin/clavulanic acid (2/1)及ampicillin/sulbactam( 2/1)皆呈現100%
抗性結果;(Mun˜oz Bellido et al., 1997)作者收集32個臨床菌株,從agar dilution的方法發現32個臨床菌株對aztreonam/ clavulanic acid (2/1)呈 現100%抑制、對amoxicillin/clavulanic acid (2/1)及ampicillin/sulbactam
(2/1)皆呈現100%抗性結果。文獻中(Smit etal.,1994) 作者收集55個臨 床 菌 株 , 從 agar dilution 的 方 法 發 現 , 55 個 臨 床 菌 株 對 trimethoprim/sulfamethoxazole (1/19) 藥 物 呈 94% 感 受 性 , 對 ticarcillin/clavulanic acid(clavulanic acid 固定濃度在 2 μg/ml)藥物呈 71%感受性;對Amoxicillin/clavulanic acid ( 2/1)藥物呈100%抗性。文獻 中(García-Rodríguez et al., 1991) 作者收集42個臨床菌株,以agar dilution的方法發現,S. maltophilia對trimethoprim/sulfamethoxazole (1/19 )藥物呈100%感受性,對aztreonam/clavulanic acid (1/1 )呈32.4%
感受性,對aztreonam/clavulanic acid (2:1)呈100%感受性。所以作者推 測在治療S. maltophilia當aztreonam/clavulanic acid(2:1)有 synergistic activity。文獻中報導(Wang et al.,2004)作者收集1999到2003年台灣 北部醫院的檢體,利用agar disk diffusion的方式發現72個檢體中有36 個檢體對ticarcillin/ clavulanic acid固定濃度在 2 μg/ml)藥物呈感受 性;60個檢體中有30個檢體對trimethoprim/sulfamethoxazole (1/19)藥 物呈感受性。相關β-lactams /clavulanic acid 整理於附錄五。
第二章 研究目的
將臨床所收集到的菌株 S. maltophilia N,前人利用 gene replacement 策 略分別得到突變株 NL1-及 NL2-,分別改變不同的誘發條件研究 L1, L2 二基因受誘發的情形。所研究的誘發條件包括: (1)在培養基中加入人類血 清(2)在培養基中加入不同濃度的 clavulanic acid (3)改變培養基中碳源的 成份(4)在培養基中中加入能抗癌的藥物 hesperetin 及能治療 benign prostatic hypertrophy (BPH)的疾病的β-sitostero。另一方面,評估不同組裝 的療方(combination regimen) 對菌株 N, NL1- 及 NL2- 抗生素感受性的 影響。
第三章 研究的重要性
目前,對於S. maltophilia 染色體中的兩個基因L1, L2 的誘發調控研 究甚少,在本篇研究中利用原菌分別將L1, L2 基因以插入突變法分別得 L1- 及 L2- 突變株。將所得的NL1-及NL2-突變株,設計不同的誘發條件 分別使L1, L2 基因被誘發(induction)後,評估L1, L2 基因promoter 分 別被誘發(induction)的情形。進而了解L1, L2 兩個基因在S. maltophilia 中 之誘發表現之機制。在感染S. maltophilia時,常使用β-lactamase的抑制劑 合倂其他β-lactam抗生素一起治療。本研究評估多種不同組裝 L2 抑制劑 /β-lactam 藥物對臨床 S. maltophilia 分離菌株的抗生素感受性。並以菌 株 S. maltophilia N 及其 isogenic mutants NL1- 及 NL2- 突變株作為 材料,研究藥物組合為拮抗(antagonism)或協同(synergism)作用的可能機 制。
第四章 研究方法
第一節 研究材料
一、 菌種及質體列於附錄一。
二、 藥品
本實驗中所使用的藥品及有機溶劑購自E. Merck, J. T. Backer Company, Sevrva Frvafeinrobiochemica, Boehringer mannhrim GmhH Biochemical, Pharmacia, Sigma Chemical Company, Biosolve 及日本和光藥物公司。
細 菌 培 養 基 使 用 的 藥 品 購 買 自 Difo Laboratories 和 Accumedia manufacturers, Inc.。抗生素藥品是向 Sigma Chemical Company,BBLTM 購買。限制酶以及其它酵素是購自TakaRa Shuzo Co. Ltd., New England Biolabs (NEB) 、promega company 和 Bethesda Research Laboratories 。 T4 DNA ligase 、DNA polymerase Ⅰ Klenow fragment 購買自 promega Co.。β-sitosterol 及 hesperetin 由林振文老師實驗室提供。結構列於圖十、
十一。
三、 引子
本實驗所使用的引子
(stock solution concentration 100 μM, working concentration 10μM)
5’-ACATTGCCTACTACACCTCC-3’
2. M13-R (20 mers)
5’-GGCTCGACTCTGTAAAGAGC-3’
3. Tc-F (18 mers)
5-CGTCAGGTGGCACTTTTC -3
4. Tc-R(19 mers)
5-GGCTTCCATTCAGGTCGAG-3
四、 培養基
本實驗中所使用的培養基配方是參照Sambrook et al. (1989)。
1. Luria-Bertaoni broth medium (LB):其成分為每 1 公升水中含有 10g tryptone,5g 的 yeast extract 及 5g 的 NaCl。
2. Luria-Bertaoni agar (LA):於 LB broth medium 中加入 1.5% agar。
3. Mueller-Hinton agar(MHA): 其成分為每 1 公升水中含有 17.5g acid hydrolysate ofcasein ,2g 的 beef extract 及 17g 的 agar。
4. XOL medium:每公升的水中含有 0.7 克 K2HPO4 、0.2 克的 KH2PO4 、 1 克的(NH4)2SO4、 0.1 克的 MgCl26H2O、 0.1 克 FeSO47H2O 、及 0.001 克 MnCl2(pH7.0)。並外加適當濃度之醣類作為碳源。
5. XOLN medium:在 XOL medium 中,加入 0.0625% yeast extract、
0.0625%tryptone,並外加適當濃度之醣類作為碳源。
五、 血清
1. 依標準抽血程序抽取健康人血液約20cc 將血液置於無凝固劑 的無菌試管中。
2. 將試管放置於室溫30min 後,以 3000rpm,5min 離心取上清液。
3. 將剩餘血清檢體保存於-20℃。
六、 試劑與緩衝溶液
1. 抽取質體 DNA 試劑
質體DNA 之抽取是依照 Birnboin & Doly (1979) 所述之 alkaline method 修飾後進行。所需試劑如下:
(1) Solution Ⅰ:25 mM Tris HCl (pH 8.0),10mM EDTA (pH8.0) 及 50 mM glucose。
(2) Solution Ⅱ :0.2 N NaOH 及 1% SDS。
(3) Solution Ⅲ :3 M potassium acetate (pH4.8) 及 5 M glacial acetic acid。
2. DNA 電泳試劑
(1) 6× Loading dye:0.25% (w/v) bromophenol blue,0.25
%(w/v)xylene cyanol 及 30% (w/v) glycerol。
(2) 0.5× TAE running buffer:40 mM Tris-acetate (pH 8.0) 及 2 mM EDTA (pH 8.0)。
(3) Staining buffer:1mM EDTA (pH8.0),10mg/ml EtBr,
100μl EtBr,100μl RNase A 及 150 ml 去離子水。
3. catechol 2,3-dioxygenease 活性測試所需試劑
(1) Potassium phosphate buffer, 0.1M pH7.5
Solution A : 27.2g KH2PO4‧H2O per liter(0.2M) Solution B : 34.9g K2HPO4‧H2O per liter(0.2M)
再以solution A 16 ml 加 solution B 84 ml, 後加去離子 水至 200 ml
(2) Assay buffer
取80 ml 0.1M pH7.5 potassium phosphate buffer 及 20 ml 10% acetone, 後加去離子水至 200 ml
(3)0.1M catechol
1.1g catechol 粉末溶至 100ml Assay buffer,使用其完全 溶解後,再保存於 4℃。
4.β-lactamase 活性測試所需試劑
(1)Sodium phosphate buffer, 0.1M pH7.0
Solution A : 27.6g NaH2PO4‧H2O per liter(0.2M) Solution B : 53.65g Na2HPO4‧7H2O per liter(0.2M) 再以solution A 39ml 加 solution B 61 ml, 後加去離子水 至 200 ml
(2)Potassium phosphate buffer, 0.1M pH7.0
Solution A : 27.2g KH2PO4‧H2O per liter(0.2M) Solution B : 34.9g K2HPO4‧H2O per liter(0.2M)
再以solution A 39 ml 加 solution B 61 ml, 後加去離子 水至 200 ml
(3) Nitrocefin
1 mg Nitrocefin 粉 末 溶 至 100μl dimethylsulfoxide (DMSO),使用其完全溶解後,再加入 1.9 ml phosphate buffer 至 2ml,保存於-20℃。
(4) CENTA
25g CENTA粉末溶至46ml 50 mM Potassium phosphate
(pH 7.0),使用其完全溶解後,再分裝0.8ml於eppendrof 中,保存於-20℃。
(5) Protein determination
Bio-Rad 200μl,Cell extract 20μl,去離子水 780μl,再以 OD 595nm 測試。
5. EDTA ,0.5M pH8.0
146.125g EDTA 及 67.41g NaOH 粉末溶至去離子水中 調整 pH 值為 8.0,總體積為 1L。
第四章 研究方法
第二節 實驗方法
一、菌種的培養與保存 1. 短期保存
將S. maltophilia 菌株培養於不含抗生素的固態培養中,放置於 37℃培養 箱培養一晚後,可放置4℃短暫保存 5~7 天,予以備用。
2. 長期保存
將本實驗所用的菌株培養於含有合適抗生素濃度的液態培基中,37℃震 盪培養一晚,視菌種的生長速度,在對數期間(log phase)取 0.7ml 的 菌液加入0.3ml 87%無菌 glycerol,使細菌與甘油的比例為 3:1,混合均 勻存放於抗凍管中,並在管上貼上標籤註明編號、菌種名稱和製做日期,
存放於-80℃備用。
二、 質體之抽取
將含有質體之菌株培養於 3ml 液態培養基的小空瓶中,可視情況加入合 適濃度之抗生素,37℃震盪培養一晚,視菌種的生長速度,在對數期間
(log phase),分裝至 1.5ml eppendorf 中,以 8000rpm 5 分鐘離心,去 除上清液後加入 100ul solutionⅠ溶液,使菌體重新懸浮,置於室溫中 5
分鐘。再加入200 ul solutionⅡ 溫和上下倒置數次後,置於冰上 5 分鐘。
再加入 150ul solutionⅢ上下倒置數次後再置於冰上 10 分鐘。接著以 12000rpm 10 分鐘離心,取上清液至另一新的 eppendorf 中以 1 比 1 比例 加入phenol/chloroform 混合均勻以 12000rpm 5 分鐘離心後取上清液至另 一eppendorf 中,重複數次使界層無雜質後再加入等量 chloroform 混合均 勻,12000rpm 5 分鐘離心,取上清液至另一 eppendorf 中加入二倍體積 95%酒精,混合均勻置於冰上 10 分鐘後,再以 12000rpm 10 分鐘離心,
去除酒精得沉澱質體 DNA,等酒精揮發乾後回溶適當無菌水中儲存於 4
℃或-20℃備用。
三、 洋菜膠體電泳分析
取洋菜粉(agarose)加熱溶於 0.5 倍的 TAE 電泳緩衝液中,配置成 0.8
%或1.0%的膠體後,待膠體冷卻至 50-60℃時,倒入鑄膠槽中並插上齒 梳(comb)。待洋菜膠體冷卻凝固後,拔掉齒梳,將樣品 DNA 與 1/6 體 積的6 倍的 loading dye 混合均勻後,加入膠體之溝槽中,並於同一片膠 體的另ㄧ溝槽中注入適量之1kb DNA marker 作為對照,以 50 或 100 伏 特通電經適量時間後,取出膠體,將其浸泡於含 0.5μg/ml ethidium bromide(EtBr)之水溶液中經適當時間後再將膠體取出以去離子水退染 3-5 分鐘,置於 UV 燈下觀察並照相。可藉由與 DNA marker 已知的片
段的量與分子大小。
五、DNA 片段回收
DNA 片段回收使用 GeneMark DNA Clean/Extraction Kit,將要分離的 DNA 片段以洋菜膠體電泳分離,於 EtBr 染色 10 分鐘,置於紫外線箱上 觀查,使用刀片切取欲回收 DNA 片段的膠體,分裝至 eppendorf 中,加 入 500-700 ul Binding solution,置於乾浴機 65℃加熱 30 分鐘,時而上 下倒置搖晃,待膠體完全溶解後,再吸取溶液至Kit 提供的 Spin column 中,下接Collection column,以 12000rpm 3 分鐘,再加入 500ul Washing Solution 12000rpm 5 分鐘,重複二次,打開 Spin column 蓋子待酒精揮 發後,再視情況加入適量無菌水於Spin column 中,於室溫作用數分鐘後,
以12000rpm 離心 10 分鐘,即得回收產物,置於 4℃備用。
六、限制酶切割
利用限制酶可辨認特定序列的特性,將DNA 與所需要的一至兩種限制酶 及配合廠商建議的緩衝溶液混合,以及廠商所建議限制酶的反應溫度,
進行DNA 切割反應,反應時的 DNA 濃度約為 1ug/50ul,反應時間視選 擇限制酶的種類而調整,完成後再以洋菜膠體電泳分析切割情形。
七、DNA 片段之黏合反應
將所選擇的載體經限制酶切割後,再與需接黏的DNA 片段以適當比例混 合均勻,並加入1ul T4 黏接酶及廠商建議的緩衝溶液,最後用無菌水補 齊體積為20ul,混合均勻,置於 16℃下作用 12-16 小時,即完成連接作 用(Ligation)。
八、 E coli DH5α勝任細胞(competent cell)之備製
先將隔夜培養的E coli DH5α 菌液以合適比例接種至 40ml 液態培養基 中,37℃震盪培養至 OD600 0.5-1.0,將菌液以 4℃,8000rpm 離心 10 分 鐘,去除上清液,再加入20ml 100mM CaCl2 混合,置於冰上 30 分鐘,
再以4℃ 8000rpm 離心 10 分後,去除上清液,再以 100mM CaCl2 回溶 至適當的濃度,使菌種懸浮於其中就成為勝任細胞即可使用,未使用之 勝任細胞可至於4℃中備用。
九、E coli DH5α之轉形作用
取適量 DNA 或質體加入 100ul 勝任細胞,置於冰上作用 10 分鐘後,再 置於42℃水浴槽中進行熱休克(heat shock)反應 2 分鐘,再快速移回冰上 再作用 2 分鐘,再加入 500ul 液態培養基,37℃震盪培養約 2 小時,再 8000rpm 5 分鐘離心,最後再塗於含有篩選抗生素的固態培養基上。37
℃ 隔 夜 培 養 。 如 果 質 體 帶 有 β -galactosidase 基 因 , 則 視 所 需 加 入 IPTG(0.8mg/ml)及 X-gal(0.8mg/ml)以進行藍白篩選。
十、質體快速篩檢法
使用已滅菌的牙籤挑選各個單一的轉形菌株,分別接種於含有合適抗生 素1m 的液態培養基中,37℃震盪培養 6-8 小時,以 12000rpm 離心 5 分 鐘,去除上清液,加入含有20ul 1%SDS 的 loading dye,劇烈震盪使沉 澱物完全打散,再各加入等體積的 phenol/chloroform,混合均勻後,以 12000rpm 10 分鐘離心,吸取 10ul 上層液進行洋菜膠電泳分析,並將僅 含有載體的菌株以上述同樣方法注入同一洋菜膠體中,做為對照組。
十一、聚合酶連鎖反應(Polymerase Chain Reaction ,PCR)
取 5ul 的染色體或 DNA 片段當作模板,加入 5ul 的 1.25mM dNTPs (dATP、dTTP、dGTP、dCTP),加入 10X Taq DNA polymerase buffer 5ul,
再個別加入適當引子5ul,DMSO 5ul 和 1ul 的 polymerase,最後加無菌 水至 50ul。均勻混合後,再視條件設定反應所需的循環溫度,最後完成 PCR 產物置於 4℃備用。
十二、破菌法:
1. 超音波破菌法(sonocator)
將隔夜培養的菌液以OD550 0.2 接種至 20ml 液態培養基中,37℃震盪培 養至 OD550 0.7~0.8,加入抗生素(inducer)或不加抗生素(no inducer)誘 導,在誘導的時間後,將菌液裝於 50 ml 的離心管,將菌體離心 12000 rpm(5~15 分鐘),用 0.33 M NaOH 當做緩衝濃液洗菌三次,再將菌體離 心12000 rpm (5~15 分鐘)將菌體上清液倒去。將菌體以適當的緩衝溶液 (Sodium phosphate 或 Tris-HCl) 回溶 2 ml,將菌體劇烈搖晃倒菌體散開,
將菌液置於冰上並將破菌的探針插入菌液液面下破菌,破菌條件為 2.5 W,10 分鐘(打 10 秒停 10 秒),破完菌後,將菌液以 12000 rpm 離心 20 分鐘,收上清液。
2.外膜破菌收蛋白法(Lysozyme):
將隔夜培養的菌液以OD550 0.2 接種至 20ml 液態培養基中,37℃震盪培 養至OD550 0.7~0.8,並用抗生素給予誘導反應後,再以 8000rpm 5 分鐘 離心,取pellet,加入 2ml 0.33M NaCl 使菌體重新懸浮,以 8000rpm 5 分鐘離心,重複三次上步驟,加入 1.2ml(含 1mM EDTA,1mg/ml lysozyme,20% sucrose/30mM Tris-HCl(pH8.0)) 回溶,置冰上作用 1 小時。再以12000rpm 離心 30 分,取上清液分裝至 eppendorf 中,保存於 -80℃中備用。
十五、抗生素感受性試驗
1. 抗生素培養基之配製
本實驗抗藥性測試的方法主要是依照agar dilution method。
泡製2 倍數系列稀釋的抗生素濃度的 Mueller-Hinton 培養基。
所使用的抗生素包括:pencillin 類抗生素 (如 carbenicillin, piperacillin、
amoxicillin、ticarcillin、piperacillin)、clavulanate、aztreonam、trimethoprim、
sulfamethoxazole 泡製的濃度為 2-256μg/ml。
2. 將隔夜新鮮培養,濁度為 0.5 McFarland 的菌液稀釋 3 倍,以 multipoint inoculator 接種至含有不同濃度抗生素的 Mueller-Hinton 培養 基上,使得每一接種點有 104的菌落數(104/spot),以 E. coli DH5α菌 株做為為陰性對照組,培養於37℃培養箱中,24 小時後觀察並紀錄其生 長情形。
3. Synergy 及 antagonism 的定義
對一菌株而言,若該菌株進行抗藥性感受性試驗,對 β-lactam/inhibitor 所表現之 MIC 與對單一該 β-lactam 所表現之 MIC 的比值 (MIC β-lactam/inhibitor/MIC β-lactam) ≦2 時,定義為 Synergy;若此比值 2 時定義為antagonism。(Lecso-Bornet M & Bergogne-Bérézin E,1997)
十六、生長曲線測定
將待測之菌株培養於 LB 或 XOLN 液體培養基培養隔夜,取些許液體培 養基懸浮菌體以適當濃度在 OD450中測其吸光值,將菌液接種至新鮮 LB 或 XOLN 液體培養基中,調整菌液起始濃度使其 OD450 之吸光值為 0.3-O.35 之間,置於 37℃培養箱震盪培養,每隔 30min 取出菌液,每株 分析樣本皆取樣兩份,測其OD450吸光值。
十七、catechol-2,3-dioxygenase 酵素活性分析
將菌液的起始濃度OD450調整為0.3-O.35 之間,加入依實驗需求不同之 條件變化,置於37℃培養箱震盪培養。約培養 40-60 min 後加入不同濃 度的抗生素或藥品試劑後,繼續培養依不同時間收菌液,每次所收取的 菌液皆為 1 ml,以 8000 rpm,5 min 離心去除上清液,回溶於 1 ml 的 C230 assay buffer 中,調適當反應濃度後 置於石英管中以 OD375先Blank 後 , 在 加 入 受 質 (0.1 M catechol) 於 375 nm 測 其 吸 光 值 。 catechol-2,3-dioxygenase 酵素活性分析皆維持低溫環境中。酵素活性定 義一單位(U)為在室溫下,50 mM potassium phosphate (pH7.5)環境下,
一分鐘內水解1 nmole catechol 所需的 catechol-2,3-dioxygenase 量。
十八、β-lactamase 酵素活性分析
1.取待測液 450μl 加入 50μl nitrocefin 混合均勻至比色計石英管中利用 比色計測定在486 nm 波長時吸光值的變化,毎 10 秒紀錄一次,總測量 時間為 3 分鐘。所得之吸光值變化以 extinction coefficient 常數 (εΔ486
=16,000 M-1cm-1) 換算在每一分鐘nitrocefin 被分解的量。蛋白質定量利 用 Bio-Rad protein assay kit 測得。最後 β-lactamase 酵素活性 specific activity (U/mg) 以菌液蛋白量每一 mg 中含有多少酵素活性 (U) 表示 之。酵素活性定義一單位(U)為在室溫下, sodium phosphate (pH7.0) 環境下,一分鐘內水解1 nmole nitrocefin 所需要的β-lactamase 量。
2. 取待測液 450μl 加入 50μl CENTA 混合均勻至比色計石英管中利用比 色計測定在405nm 波長時吸光值的變化,毎 10 秒紀錄一次,總測量時間 為3 分鐘。所得之吸光值變化以 extinction coefficient 常數 (εΔ405 = 6400 M-1cm-1) 換算在每一分鐘CENTA 被分解的量。蛋白質定量利用 Bio-Rad protein assay kit 測得。最後β-lactamase 酵素活性 specific activity (U/mg) 以菌液蛋白量每一 mg 中含有多少酵素活性 (U) 表示之。酵素活性定 義一單位(U)為在室溫下, sodium phosphate (pH7.0)環境下,一分鐘 內水解1 nmole CENTA 所需要的β-lactamase 量。(Bebrone et al., 2001)
十九、抑制劑分析
1. 取 200μl 的 EDTA (0.5M ,pH8.0)加入已稀釋好的 Cell extract 800 μl,混合均勻後使之 EDTA 濃度為 100 mM 置室溫 10 分鐘後,以 CENTA 為受質,取490μl 至比色管中加入 10μl CENTA 利用分光光度計在 405 nm 波長下測定吸光值的變化,測試 EDTA 濃度依序為 100、150,200,
250 mM。
2. 取 0.4μl/ml 的 clavulanic acid 加入已稀釋好的 Cell extract 999 μ l,混合均勻後使之 clavulanic acid 濃度為 4μg/ml 置室溫 10 分鐘後,以 ENTA 為受質,取 490μl 至比色管中加入 10μl CENTA 利用分光光度 計在405 nm 波長下測定吸光值的變化,測試 clavulanic acid 濃度依序為 8,10,100 μg/ml。
十九、蛋白質濃度測定
取 200μl Bio-Rad protein assay 染劑加入 dd H2O 780μl,加入已 稀釋好的Cell extract 20 μl,混合均勻後至比色管中利用分光光度計在 595 nm 波長下測定吸光值的變化,並分析蛋白質濃度。
第五章 研究結果
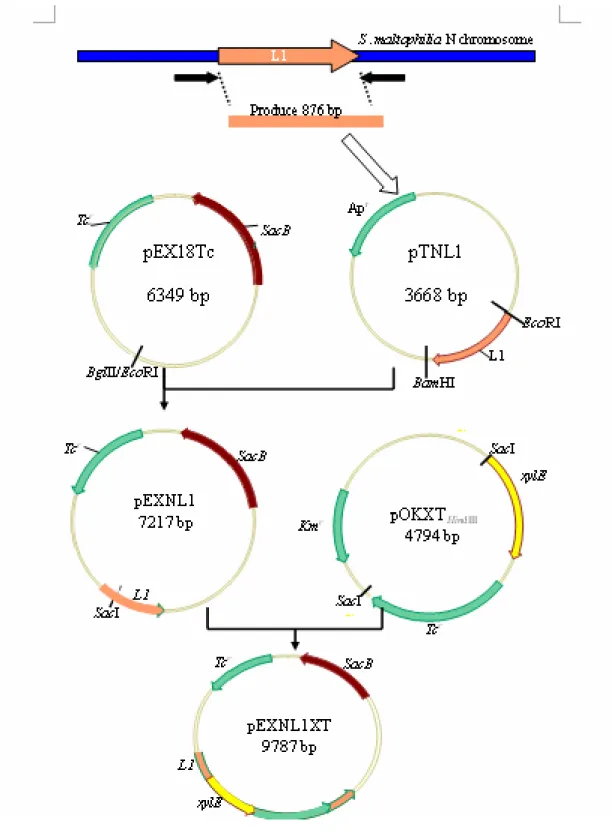
第一節 重組質體之構築
質體pOKXTHindIII之構築( xylE-Tc 基因套組)
以質體pEX18Tc 當作模板,利用 Tc-F (5-CGTCAGGTGGCA CTTTTC -3) 及Tc-R (5-GGCTTCCATTCAGGTCGAG-3)當引子進行 PCR。條件為 94℃
10 分鐘;30 個循環的 94℃ 1 分鐘,50℃ 1 分鐘,72℃ 1 分鐘;最後 72℃
5 分鐘。 PCR 產物以 0.8% 洋菜膠體電泳分離確定,所得產物片段大小 為1408 bp,回收 1408 kb 的 DNA 片段後,以 T-Vector 承接後經轉形作 用送入E.coli DH5α 勝任細胞內。以含有 ampicillin 100 μg/ml 及 tetracyclin 10 μg/ml 的 LB 平板培養基進行篩選,得到具有抗 ampicillin 及 tetracyclin
的轉殖株。抽取其質體DNA,以限制酶切割檢查確定無誤後,將構築的
質體命名為pTTc。
將質體pTTc 以限制酶 BamHΙ 與 XbaΙ 處理後,回收約 1480 kb 的 DNA 片段,將此片段與已經被 BglⅡ與 XbaΙ 處理後的載體 pTXylERI(前人所 構築,具 xylE 基因,可表現 C23O 活性) 承接後,經轉形作用送入 E. coli DH5α 勝任細胞內。以含有 ampicillin 100 μg/ml 及 tetracyclin 10 μg/ml 的 LB 平板培養基進行篩選,得到具有抗 ampicillin 及 tetracyclin 的轉殖株。
抽取其質體DNA,以限制酶切割檢查後,將構築的質體命名為 pTXylETc。
將質體pTXylETc 轉殖於載體 pOK12,經轉形作用送入 E. coli DH5α 勝任細胞內。以含有kanamycin 100 μg/ml 及 tetracyclin 10 μg/ml 的 LB 平 板培養基進行篩選,得到具有抗kanamycin 及 tetracyclin 的轉殖株。將構 築的質體命名為pOKXylETcRI。
將質體pOKXylETcRI 轉殖於載體 pX1918GT,經轉形作用送入 E. coli DH5α 勝任細胞內。以含有 ampicillin 100 μg/ml 及 tetracyclin 10 μg/ml 的 LB 平板培養基進行篩選,得到具有抗 ampicillin 及 tetracyclin 的轉殖株。
可得在xylE-Tc 基因套組兩側有一系列對稱的多重限制酶切位,將此構築 的質體命名為pXXT。
將具 xylE-Tc 基因套組的質體 pXXT 轉殖於載體 pOK12,經轉形作 用送入 E. coli DH5α 勝任細胞內。以含有 kanamycin 100 μg/ml 及 tetracyclin 10 μg/ml 的 LB 平板培養基進行篩選,得到具有抗 kanamycin 及tetracyclin 的轉殖株。將構築的質體命名為 pOKXTHindIII。如圖一
第二節 S. maltophilia N isogenic mutant NL1- 及 NL2- 獲得
菌株 S. maltophilia N 的來源來自中國醫藥大學附屬醫院臨床細菌室,
前人利用 gene replacement 策略分別將 L1, L2 基因以 xylE-Tc 基因套 組插入,得到突變株NL1-及NL2-突變株。如圖二及圖三所示。
第三節 誘發 (induction) Stenotrophomonas maltophilia 菌株 L1 及 L2 基因啟動子表現因素之研究
一、 人類血清對Stenotrophomonas maltophilia 菌株 L1 及 L2 基因啟動子 誘發 (induction) 的影響
將 NL1- 及 NL2- 突變株,分別培養在兩種培養基中,一種為 LB 培養 基,另一種為加入5 %的健康人血清之 LB 培養基。在這兩種培養基中又 分有加入抗生素(cefuroxime,50 μg/ml)及不加抗生素的對照組。將這兩個
突變株培養於上述四種培養基後於一定時間後收菌,以 OD450 測得 S.
maltophilia 的生長情形,並以 C230 assay buffer 破菌測得 catechol-2,3- dioxygenase 的活性。由實驗結果得知 N strain 的 NL1-及 NL2-突變株在 有加入5 %的健康人血清的 LB 培養基培養後,對於菌株的生長上是沒有 差異的。所以,將NL1-及NL2-突變株培養於有加入5 %的健康人血清的
LB 培養基中,不會影響到菌株的生長。加入抗生素(cefuroxime,50 μg/ml)
後對於 NL1-及 NL2-這兩個突變株的生長情形也是沒有差異。所以,將 NL1-及 NL2-突變株培養於加抗生素(cefuroxime,50 μg/ml)的 LB 培養基 中,不會影響到菌株的生長。從實驗中也發現NL1-及NL2-這兩個突變株 的catechol-2,3-dioxygenase 的活性,在 5 %的健康人血清培養基中沒有明 顯可測得的活性,顯示只有 5 % 健康人血清並不能誘發 L1 或 L2 基因 啟動子的表現。加入抗生素(cefuroxime,50 μg/ml)後, NL1-及 NL2-這兩 個突變株的 catechol-2,3-dioxygenase 的活性有增加,但其活性的表現量 與加有5%的健康人血清及抗生素(cefuroxime,50 μg/ml)的實驗組相比,
並沒有明顯的差異,所以,5 % 的健康人血清對 cefuroxime 誘發 L1、
L2 基因啟動子表現的能力沒有明顯的影響。此外,在受到 cefuroxime 誘 發表現的 NL1- 及 NL2- 菌株,catechol-2,3- dioxygenase 活性的表現量 都是 NL2-大於 NL1-。所以,由實驗中可知 cefuroxime 對 L2 基因啟動 子的誘發能力大於對 L1 基因啟動子的作用。NL1-及 NL2-突變株在 5%
人類血清的影響下其生長情形及 catechol-2,3-dioxygenase 的活性列於表 一。
二、 不同濃度的clavulanic acid 對 Stenotrophomonas maltophilia 菌株 L1 及L2 基因啟動子誘發 (induction) 的影響
1. 分別將 NL1-及 NL2-突變株培養在含有三種不同濃度 clavulanic acid (10,50,100 μg/ml) 的 LB 培養基中,另一以不加 clavulanic acid 的 LB 培養基為對照組,預養到 2.5 小時後收菌,以 OD450 測得 S.
maltophilia 的生長情形,並以 C230 assay buffer 破菌測得 catechol-2,3- dioxygenase 的活性。實驗結果發現與對照組相比,NL1-及 NL2-突變 株培養在含有濃度為100 μg/ml clavulanic acid 的培養基中生長緩慢,
在含有50 μg/ml 及 10 μg/ml clavulanic acid 的培養基中生長情形和 對照組生長情形一致。所以,高濃度clavulanic acid (100 μg/ml) 對 S. maltophilia 的生長有明顯的抑制效果。從實驗中也發現 clavulanate 會 誘發 NL1- 及 NL2- 這兩個突變株 catechol-2,3-dioxygenase 活性的表 現,顯示 clavulanate 對 L1 及 L2 基因的表現是一有效的誘發子 (inducer)。分別比較 NL1- 及 NL2-在不同濃度clavulanate 作用下,其 catechol-2,3-dioxygenase 活性被誘發程度。結果顯示,不論對 L1 或 L2 基因啟動子而言,50 μg/ml clavulanic acid 的誘發力較 10 μg/ml clavulanic acid 強。NL1- 及 NL2-突變株在不同濃度 clavulanate 影響 下,其生長情形及誘發catechol-2,3-dioxygenase 的活性列於表二。
2. 分別將 NL1-及 NL2-突變株培養在含有五種不同濃度 clavulanic acid (2,8,10,16,50 μg/ml) 的 LB 培養基中,以及培養於加抗生素 (cefuroxime,50 μg/ml)的 LB 培養基中,另一以不加 clavulanic acid
及cefuroxime 的 LB 培養基為對照組,預養到 2.5 小時後收菌,以 OD450 測得 S. maltophilia 的生長情形,並以 C230 assay buffer 破菌測得 catechol-2,3-dioxygenase 的活性。實驗結果發現與對照組相比,NL1- 及 NL2-突變株培養在含有五種不同濃度 clavulanic acid,及培養於加 抗生素(cefuroxime,50 μg/ml)的 LB 培養基中生長情況一致。
從實驗中發現不同濃度的 clavulanate 誘發 NL1- 及 NL2- 這兩個突變 株 catechol-2,3-dioxygenase 活 性 的 表 現 不 同 , NL1-在 分 別 含 有 clavulanic acid 2,8,10 μg/ml 的 LB 培養基中,catechol-2,3-dioxygenase 活性的表現隨clavulanic acid 濃度增加而增加活性表現,但在分別含 有 clavulanic acid 16 , 50 μg/ml 的 LB 培 養 基 中 , catechol-2,3-dioxygenase 活性表現不隨 clavulanic acid 濃度增加而增
加活性表現。NL2-在分別含有clavulanic acid 2,8,10,16,50 μg/ml 的 LB 培養基中,catechol-2,3-dioxygenase 活性的表現隨 clavulanic acid 濃度增加而增加活性表現。顯示不同濃度(2,8,10,16,50 μg/ml) clavulanate 對 L2 基因的誘發能力大於 L1 基因的誘發能力。分別比較 NL1- 及 NL2-在 不 同 濃 度 clavulanate 作用下,及培養於抗生素
(cefuroxime,50 μg/ml)的 LB 培養基中其 catechol-2,3-dioxygenase 活 性被誘發程度。結果顯示,不論對 L1 或 L2 基因啟動子而言,50 μg/ml cefuroxime 的 誘 發 力 較 這 五 種 濃 度 (2 , 8 , 10 , 16 , 50 μg/ml)clavulanate 強。NL1- 及 NL2-突變株在不同濃度 clavulanate 及
抗生素的影響下,其誘發catechol-2,3-dioxygenase 的活性整理於圖十 二。
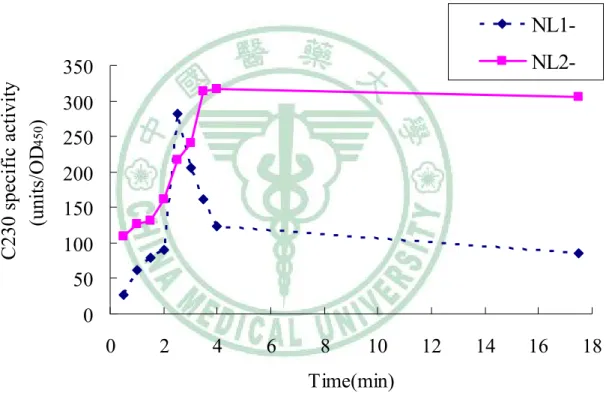
三、 Clavulanic acid (50 μg/ml) 對 Stenotrophomonas maltophilia NL1- 及 NL2- 菌株誘發 (induction)動力學的研究
分別將NL1-及NL2-突變株培養在含有clavulanic acid ( 50 μg/ml) 的 LB 培養基中,每30 min 收菌連續監測四小時後,將其隔夜培養後再收一次 菌。每次分析樣本以OD450測得S. maltophilia的生長情形,並以C230 assay buffer 破菌測得 catechol-2,3-dioxygenase 的活性。實驗結果發現培養在含 有clavulanic acid ( 50 μg/ml) 的 LB 培養基中,所測得的 NL1-及NL2-突 變株生長情形相近,且經隔夜培養後,NL1-及NL2-突變株並沒有死亡。
實驗結果發現NL1-突變株及NL2-突變株的catechol-2,3-dioxygenase 的活 性皆在含有clavulanic acid ( 50 μg/ml)的培養基中培養兩小時後出現活性 高峰,隨後,NL1-突變株的 catechol-2,3-dioxygenase 的活性會隨培養時