行政院國家科學委員會專題研究計畫 成果報告
陽離子性樹枝型高分子基因傳送系統與巨噬細胞間之交互 作用
研究成果報告(精簡版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 96-2221-E-041-024-
執 行 期 間 : 96 年 08 月 01 日至 97 年 07 月 31 日 執 行 單 位 : 嘉南藥理科技大學藥物科技研究所
計 畫 主 持 人 : 郭榮華
計畫參與人員: 碩士班研究生-兼任助理人員:林宜陵 碩士班研究生-兼任助理人員:曾若雯
處 理 方 式 : 本計畫可公開查詢
中 華 民 國 97 年 09 月 19 日
Journal of Biotechnology 129 (2007) 383–390
Remnant cationic dendrimers block RNA migration in electrophoresis after monophasic lysis
Jung-Hua Steven Kuo
∗, Yi-Lin Lin
Graduate Institute of Pharmaceutical Sciences, Chia Nan University of Pharmacy and Science, 60 Erh-Jen Road, Sec. 1, Jen-Te, Tainan 717, Taiwan Received 17 July 2006; received in revised form 3 November 2006; accepted 23 January 2007
Abstract
Cationic dendrimers such as poly(amidoamine) (PAMAM) and poly(propyleneimine) (PPI) have attractive characteristics for the delivery of nucleic acid and various biomedical applications. Most studies have focused on cationic dendrimer-based intracellular delivery, and very few studies have focused on the non-specific interaction of remnant cationic dendrimers with total RNA after isolation directly from cells in vitro.
We examined RNA isolation using the common method of monophasic lysis from human macrophage-like cells (U937) and mouse fibroblast cells (NIH/3T3) that had been exposed to dendrimers and DNA/dendrimer complexes using gel electrophoresis. We found that PAMAM and PPI dendrimers strongly altered the mobility of RNA in the gels. In addition, the extent of dendrimer-induced alteration in RNA mobility was directly dendrimer-generation-dependent: the alteration was greater with higher-generation dendrimers. We also found that DNA/dendrimer com- plexes at higher dendrimer to DNA ratios interacted with RNA after isolation while gene expression was maintained. The interactions between RNA and remnant dendrimers after isolation were caused by electrostatic bindings, and we recovered total RNA using high ionic strength solvents (2 M NaCl solution) to disrupt the electrostatic forces binding dendrimers to RNA. Because RNA isolation is routinely used for bio- logical applications, such dendrimer-induced alteration in RNA mobility should be accounted for in the further processing of RNA-related applications.
© 2007 Elsevier B.V. All rights reserved.
Keywords: Cationic dendrimer; RNA isolation; DNA/dendrimer complexes; Monophasic lysis; Gel electrophoresis
1. Introduction
Dendrimers are a family of highly branched polymers with radial symmetry characterized by their unique structures and properties, such as dense end groups at the surface, a spheri- cal shape in solution, biocompatibility, and the precise control of size (Boas and Heegaard, 2004; Esfand and Tomalia, 2001;
Svenson and Tomalia, 2005). These attractive nano-sized archi- tectures of dendrimers make them suitable for targeted delivery systems, for example, delivery of DNA and oligonucleotides into eukaryotic cells, solubilization of sparingly soluble drugs, nano-carriers for transepithelial transport, and platforms for can- cer therapeutics (Cloninger, 2002; Dennig and Duncan, 2002;
Kitchens et al., 2005; Kobayashi and Brechbiel, 2005). One of the most exciting research fields has been that of gene delivery
∗Corresponding author. Tel.: +886 6 266 4911x212; fax: +886 6 266 6411.
E-mail address:[email protected](J.-H.S. Kuo).
using cationic dendrimers such as poly(amidoamine) (PAMAM) and poly(propyleneimine) (PPI) (Dufes et al., 2005). Cationic dendrimers interact with anionic DNA to form highly con- densed complexes for increased binding to the cell surface, enter the cell via endocytosis, protect DNA from enzyme degra- dation, and lead to efficient gene expression (Ruponen et al., 2003).
Because most cationic dendrimers used are non- biodegradable, their non-specific binding to anionic molecules such as RNA may occur during or after intracellular trafficking.
Cationic dendrimers interact with anionic synthesized RNA as well as deliver RNA molecules into cells (Bielinska et al., 1996; Wu et al., 2005; Zhao et al., 2004). RNA has been regarded as a highly informative molecule involved in diverse cellular processes such as transcriptional regulation, post-transcriptional RNA processing, retroviral replication, and protein synthesis, among others (Bielinska et al., 1996).
The isolation and purification of total RNA are basic methods for a variety of biomedical applications and analyses, and
0168-1656/$ – see front matter © 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.jbiotec.2007.01.030
384 J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390
obtaining high quality intact RNA is important because this is the first and the most crucial step in performing fundamental molecular biology research. While most efforts have focused on cationic dendrimer-based intracellular delivery, few studies have focused on the interaction of remaining dendrimers with total RNA after isolation directly from cells in vitro. Among the methods of isolation and purification of total RNA from cells, monophasic lysis is the most frequently used method that allows the simultaneous recovery of RNA, DNA, and protein from cultured cells. The monophasic solution composed primarily of guanidine isothiocyanate, surfactant, and phenol has lysed cells and inhibited RNase. RNA in the aqueous supernatant was then separated from DNA and protein by adding chloroform, and recovered using precipitation with isopropyl alcohol (Sambrook et al., 2001).
We hypothesize that non-specific interactions between RNA and remnant dendrimers may interfere with the RNA purifica- tion process from cells treated with cationic dendrimers alone or nucleic acid-dendrimer complexes. In the present study, we tested this hypothesis by investigating the interaction of cationic dendrimers (PAMAM and PPI) with total RNA isolated from cells using monophasic solution. We evaluated RNA isolation from human macrophage-like cells (U937) and mouse fibrob- last cells (NIH/3T3) exposed to dendrimers and DNA/dendrimer complexes. We found that the strong interactions occurring between these remnant dendrimers and RNA were reflected in gel electrophoresis. These results provided direct experimental support for our hypothesis.
2. Materials and methods 2.1. Materials
Starburst PAMAM dendrimers (Generation 5 (PAMAM 5.0) and 2 (PAMAM 2.0)) and PPI dendrimers (DAB 3.0 and 2.0) were purchased from Sigma–Aldrich Chemicals (St. Louis, MO, USA). Starburst PAMAM dendrimers were supplied in methanol, which was removed using vacuum evaporation, and stored with phosphate buffered saline (PBS) at 4◦C.
All other chemicals were of the highest grade available. The pDNA (pSG5lacZ, 8 kb), which encodes the lacZ gene for - galactosidase, was driven by an SV40 promoter to assess gene expression. The pSG5lacZ was amplified in E. Coli and purified using a Pure Yield Plasmid Midiprep System (Promega Corp., Madison, WI, USA). The purity of pSG5lacZ was established using UV spectroscopy (an A260/A280ratio ranging from 1.80 to 1.89 was used).
2.2. Cell lines
A U937 (human macrophage-like) cell line was maintained in RPMI 1640 medium (Gibco, Grand Island, NY, USA) sup- plemented with 10% heat-inactivated fetal bovine serum (FBS) and 100 U/mL penicillin/100g/mL streptomycin (Sigma).
NIH/3T3 (mouse fibroblast) cells were grown in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with antibi- otics (penicillin and streptomycin) and 10% FBS.
2.3. Cationic dendrimer treatment of cells
Cells (1× 107) were subjected to cationic dendrimers in serum-free medium for 3 h of incubation at 37◦C, washed with PBS, and then analyzed for RNA isolation. For DNA/cationic dendrimer complexes, cells (1× 107) were treated to a stan- dard transfection protocol. Briefly, DNA/dendrimer complexes at various weight ratios were formed in serum-free medium for 10 min at room temperature before adding them to the medium of cells. Transfection was performed for 3 h in serum-free medium and then supplemented with 10% FBS and 1% antibiotics. Cells were harvested 48 h after transfection, washed with PBS, and then analyzed for gene expression of-galactosidase and RNA isolation. The transfection activity was quantified using a com- bined -Gal Assay kit (Invitrogen, Carlsbad, CA, USA) and BCA Protein Assay Reagent Kit (Pierce, Rockford, IL, USA) (Rosenthal, 1987).
2.4. Isolation of RNA from cells
Total RNA from cells, untreated or treated with cationic den- drimers and DNA/cationic dendrimer complexes, was extracted using TRIZOL reagent consisting of phenol and guanidine isoth- iocyanate (Invitrogen, Gaithersburg, MD, USA). Briefly, the cell pellet was lysed and homogenized by adding 1 mL TRIZOL reagent. After 5 min of incubation at 25◦C, phase separation was achieved by adding 0.2 mL chloroform to the homogenized samples. After 10 min of centrifugation at 11,000× g at 4◦C, RNA from the aqueous phase was precipitated by mixing it with 0.5 mL of isopropyl alcohol. The RNA precipitate was washed three times with 1 mL of 75% ethanol in diethylpyrocarbonate (DEPC)-treated water and air-dried for 10 min. The RNA was dissolved in RNase-free water and stored at−80◦C for further analysis. The purity of extracted total RNA was confirmed using the A260/A280ratio.
2.5. Agarose gel electrophoresis
The integrity of the isolated RNA was assessed using agarose gel electrophoresis. One microgram of isolated RNA was elec- trophoresed at 100 V for 20 min on a 1% agarose gel containing formaldehyde and then stained with ethidium bromide.
2.6. Cytotoxicity assay
Before isolating the RNA, the activity of dehydrogenases (an indicator of cell viability) in the cells (untreated or treated with cationic dendrimers and DNA/dendrimer complexes) was simultaneously assessed. Negative control cells contained no cationic dendrimers. To measure cell viability, 10L of a cell- counting kit solution, a tetrazolium salt that produces a highly water-soluble formazan dye upon biochemical reduction in the presence of an electron carrier (1-methoxy PMS) (Cell-Counting Kit-8; Dojindo Laboratories, Tokyo, Japan), was added to a 100L culture medium and incubated for 1–4 h. The amount of yellow formazan dye generated by dehydrogenases in cells is directly proportional to the number of viable cells in a culture
J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390 385
medium. The absorbance at 450 nm was obtained using an ELISA reader with a reference wavelength of 595 nm. Results are reported as cell viability percentage (average OD/average negative control OD)± standard deviation (S.D.).
2.7. Recovering the RNA from the dendrimer–RNA pellets
After checking retarded RNA migration using agarose gel electrophoresis, the dendrimer–RNA pellets were precipitated by mixing them with 0.5 mL of isopropyl alcohol. The RNA precipitate was washed three times with 1 mL of 75% ethanol, air-dried for 10 min, and then dissolved in 1 mL of 2 M NaCl solution for 12 h. After it had been precipitated and washed, the RNA was restored in RNase-free water at−80◦C.
3. Results
3.1. Cell viability assays
To assess the cytotoxic effect of cationic dendrimers, cells before RNA isolation were incubated with cationic dendrimers and DNA/dendrimer complexes. Dehydrogenase activity was
then analyzed. An ideal carrier for biomedical applications should be non-toxic to cells. Thus, our study focused on the concentration ranges within which cationic dendrimers did not elicit toxic effects to cells. For the concentration ranges of the cationic dendrimers and their complexes with DNA used in the following experiments, cell viability was greater than 90%.
3.2. The interaction between cationic dendrimers and RNA molecules
We isolated total RNA from cells untreated or treated with cationic dendrimers using monophasic lysis. The integrity of the isolated RNA was assessed using agarose gel electrophore- sis and then ethidium bromide staining (Fig. 1). Untreated cells revealed sharp typical rRNA bands with more RNA in the 28S band than the 18S band. Cationic dendrimers exhib- ited dose- and generation-dependent blocks on the mobility of RNA in the gels. For U937 cells, high-generation dendrimers at high concentrations (PAMAM 5.0 (≥7.5 g/mL) and DAB 3.0 (≥20 g/mL)) completely prevented the mobility of RNA in the gels. PAMAM 5.0 (7.5g/mL) required less concentration than
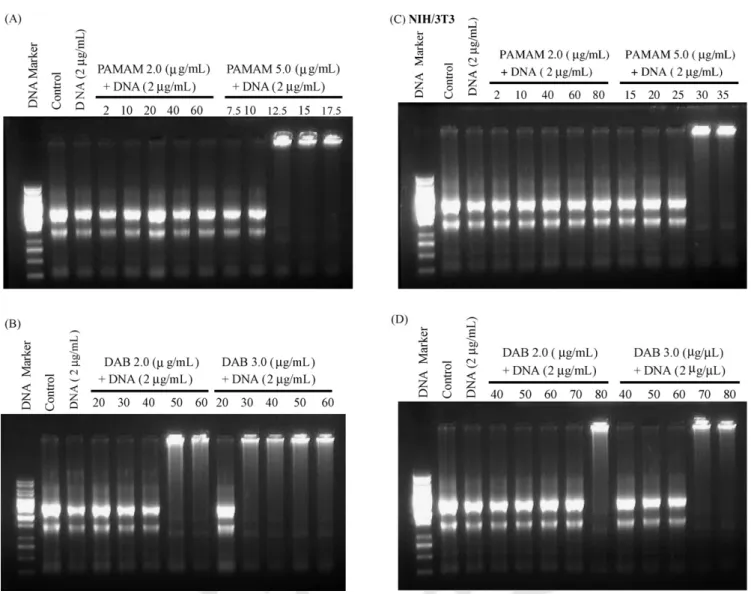
Fig. 1. Electrophoresis of isolated RNA from (A, B) U937 and (C, D) NIH/3T3 cells treated with PAMAM (PAMAM 2.0 and 5.0) and PPI (DAB 2.0 and 3.0) with an increasing concentration of dendrimers. Control samples were isolated RNA from cells without adding cationic dendrimers.
386 J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390
Fig. 2. Electrophoresis of mixing purified RNA from (A, B) U937 and (C, D) NIH/3T3 cells with PAMAM (PAMAM 2.0 and 5.0) and PPI (DAB 2.0 and 3.0) with an increasing concentration of dendrimers. Control samples were isolated RNA from cells without adding cationic dendrimers.
DAB 3.0 (20g/mL) to block the mobility of RNA in the gels.
No retarded RNA migration was observed with low-generation PAMAM 2.0, even at higher concentrations (up to 40g/mL).
However, low-generation PPI dendrimers (DAB 2.0) at a higher concentration (≥40 g/mL) retarded RNA migration in the gels.
Also, fibroblasts (NIH/3T3) required a greater concentration of dendrimers than did macrophages (U937) to retard RNA migration in the gels. To further confirm electrostatic interac- tions between RNA and cationic dendrimers, electrophoresis by mixing purified RNA with cationic dendrimers was performed.
All cationic dendrimers strongly bonded to RNA molecules in both U937 and NIH/3T3 cells (Fig. 2). These findings provided direct evidence that electrostatic interactions occurred between cationic dendrimers and RNA molecules. As the generation and concentration of cationic dendrimers became higher, stronger bonding occurred between RNA and cationic dendrimers. These findings correlated well with previous results obtained using synthesized RNA (Wu et al., 2005; Zhao et al., 2004).
3.3. The interaction between DNA/dendrimer complexes and RNA molecules
Because cationic dendrimers bind with DNA and mediate gene transfer into eukaryotic cells, the effect of complexing DNA with cationic dendrimers on the isolation of total RNA
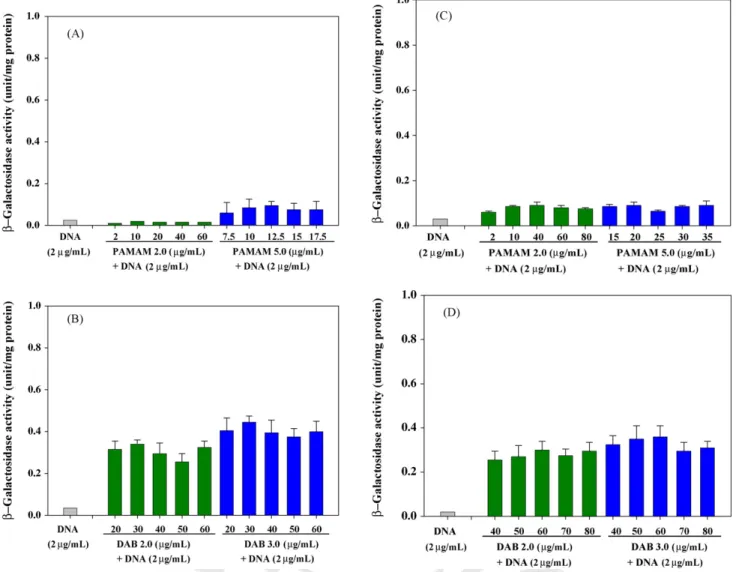
from cells was also tested. Complexes between DNA and high- generation dendrimers (PAMAM 5.0 and DAB 3.0) blocked RNA migration in the gels at higher ratios of dendrimer to DNA (Fig. 3). No significantly preventive effects on RNA migration were observed for complexes between DNA and low-generation PAMAM dendrimers (PAMAM 2.0), but DNA/DAB 2.0 com- plexes at higher ratios of dendrimer to DNA also blocked RNA migration. Due to attenuated cationic charges by anionic DNA, complexes between DNA and dendrimers required a greater con- centration of dendrimers than did dendrimers alone to block the mobility of RNA in the gels. Also, fibroblasts (NIH/3T3) required a greater concentration of DNA/dendrimer complexes than did macrophages (U937) to retard RNA migration in the gels. As nitrogen/phosphorus (N/P) ratios of complexes became higher (except for PAMAM 2.0), stronger bonding occurred between RNA and cationic dendrimers. These results further confirmed that electrostatic interactions between RNA and cationic DNA/dendrimer complexes occurred during or after intracellular trafficking. We also simultaneously examined the in vitro gene expression of DNA/dendrimer complexes (Fig. 4) and found significantly higher gene expression for DAB 3.0 and 2.0 than for PAMAM 5.0 and 2.0. These findings revealed that the transfection activity of DNA/dendrimer complexes was not altered by the interaction between dendrimers and intracellular RNA.
J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390 387
Fig. 3. Electrophoresis of isolated RNA from (A, B) U937 and (C, D) NIH/3T3 cells treated with complexes between dendrimers (PAMAM (PAMAM 2.0 and 5.0) and PPI (DAB 2.0 and 3.0)) and DNA (2g/mL) with an increasing concentration of dendrimers. Control samples were isolated RNA from cells without adding cationic dendrimers. Lane 3 in each panel presents isolated RNA from cells after adding 2g/mL of DNA.
3.4. Recovering the RNA from the dendrimer–RNA pellets by using NaCl solution
The electrostatic forces binding cationic molecules to nucleic acid can be disrupted by high ionic strength solvents (Dahle and Macfarlane, 1993; Tang and Szoka, 1997). We recovered RNA from the dendrimer–RNA pellets using 2 M NaCl solution for 12 h to reduce the electrostatic bonding between the isolated RNA and cationic dendrimers (PAMAM 5.0, DAB 3.0, and DAB 2.0). The integrity of the recovered RNA was then assessed using agarose gel electrophoresis and then ethidium bromide staining (Fig. 5). We found that the isolated RNA was totally recovered for cells treated with PAMAM 5.0, DAB 3.0, and DAB 2.0. Total RNA was also recovered from cells treated with DNA/dendrimer (PAMAM 5.0, DAB 3.0, and DAB 2.0) complexes.
4. Discussion
The successful isolation of total RNA from cells is funda- mentally important for molecular biology research. Cationic
molecules interact with anionic RNA in cells. Cationic surfactant (Catrimox-14) has been successfully used to isolate total RNA from cells in culture and clinical samples because it lyses cells and protects RNA from degradation (Macfarlane and Dahle, 1997; Dahle and Macfarlane, 1993). PAMAM dendrimers bind trans-acting responsive element (TAR) synthesized RNA and inhibit the catalytic activity of Candida ribozymes by forming RNA/dendrimer complexes (Wu et al., 2005; Zhao et al., 2004).
In addition to the delivery of nucleotides and small interfering RNA (siRNA), PPI dendrimers alter the expression of many endogenous genes (Omidi et al., 2005). However, no study has indicated that RNA electrophoresis after the isolation of total RNA from cells using monophasic lysis is blocked by remnant cationic dendrimers.
Dendrimer-induced alteration in RNA mobility after isola- tion is difficult to detect solely by checking the A260/A280ratio (typically greater than 1.9) using UV spectroscopy, because PAMAM and PPI dendrimers also contribute to the intensity of the absorbance band (data not shown). However, we found, using agarose gel electrophoresis, that PAMAM and PPI den-
388 J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390
Fig. 4. In vitro gene expression after transfection into (A, B) U937 and (C, D) NIH/3T3 cells. Cells (1× 107) were transfected with 2g/mL of plasmid DNA alone or complexed with increasing concentrations of PAMAM (PAMAM 2.0 and 5.0) and PPI (DAB 2.0 and 3.0) dendrimers. The data are given as the mean± S.D.
(n = 3).
drimers strongly block the mobility of RNA after it has been isolated from cells. Therefore, RNA inhibition by dendrimers during isolation might hinder other RNA applications such as Northern blotting, RT-PCR, and microarray target labeling. In the present study, we demonstrated that dendrimer-induced alter- ation in RNA mobility can be totally recovered using high ionic strength solvents (2 M NaCl solution) to disrupt the electrostatic forces binding dendrimers to RNA. We suggest that washing RNA with 2 M NaCl solution after its isolation from cells treated with cationic dendrimers is necessary to obtain high quality and pure RNA for further biological applications.
Although other RNA isolation methods are available, monophasic lysis is the most frequently used method that allows the simultaneous recovery of RNA, DNA, and protein from cul- tured cells. No matter of what kind of RNA isolation methods are used, cell lysis using various lysis reagents is the common first step for further purification. After this stage, cytoplasm rRNA is separated from protein in aqueous medium and forms RNA/remnant dendrimer complexes via electrostatic attractions.
Because the cationic dendrimers we used were highly water sol- uble and nonspecifically binded to anionic molecules, we expect that dendrimer-induced alteration in RNA mobility still occurs when using other methods of isolation.
The fate of the dendrimer–nucleotide complexes within cells and the mechanism of dendrimer-mediated cellular uptake of nucleotides have not been fully clarified. Recently, PAMAM 5.0 dendrimers were shown to interact with cell membranes by forming holes (Hong et al., 2004). Internalizing DNA/dendrimer complexes was previously shown to be mediated by membrane rafts and escape into the cytosol before trafficking to the nucleus (Manunta et al., 2004). Also, dendrimers may cause endosomal rupture and release the complex or DNA into the cytoplasm by acting as a “proton sponge” (Tang et al., 1996). DNA/dendrimer complexes at higher ratios of dendrimers to DNA have been shown to markedly inhibit transcriptional activity in vitro and not affect elongation of the RNA transcript and translation (Bielinska et al., 1997). We found that DNA/dendrimer com- plexes at higher ratios of dendrimers to DNA also interacted
J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390 389
Fig. 5. Using 2 M NaCl solution to recover the isolated RNA from (A) U937 and (B) NIH/3T3 cells treated with cationic dendrimers and DNA/dendrimer complexes.
with RNA during isolation while gene expression was main- tained (Figs. 3 and 4). Although various dendrimers (generation or PAMAM/PPI) interacted differentially on a weight-basis, a similarity existed on dendrimer-induced alteration in RNA mobility.
We also isolated RNA from U937 and NIH/3T3 cells using cationic lipid and lipoplexes (50–100g/mL, TransFast (Promega)) and found no RNA block in the gels (data not shown). This indicated that cationic dendrimers produced a stronger RNA block than cationic lipids did. We also found that another non-viral vector that is similar to a cationic den- drimer, polyethyleneimine 25 K (Sigma–Aldrich), blocked the mobility of RNA in the gels isolated from U937 and NIH/3T3 cells (data not shown). The preventative effect of dendrimers on the mobility of RNA in other cell types remains to be established.
In conclusion, remnant cationic dendrimers (PAMAM 5.0, DAB 2.0, and DAB 3.0) strongly blocked RNA electrophoresis after the isolation of total RNA from cells using monophasic lysis. These remnant dendrimers strongly blocked the mobility of RNA via electrostatic interactions. The extent of dendrimer- induced alteration in RNA mobility is dependent on the dendrimer generation: alteration in RNA mobility is greater with higher-generation dendrimers. At higher dendrimers-to-DNA ratios, RNA electrophoresis was blocked by DNA/dendrimer
complexes, but gene expression of these complexes was not affected. We also found that dendrimer-induced alteration in RNA mobility was totally recovered using high ionic strength solvents (2 M NaCl solution) to disrupt the electrostatic forces binding dendrimers to RNA. Because RNA isolation is routinely used for biological applications, such dendrimer-induced alter- ation in RNA mobility should be accounted for when further processing RNA for related applications.
Acknowledgment
This work was supported by grant NSC 95-2216-E-041-003 from the National Science Council, Taiwan.
References
Bielinska, A., Kukowska-Latallo, J.F., Johnson, J., Tomalia, D.A., Baker, J.R., 1996. Regulation of in vitro gene expression using antisense oligonucleotides or antisense expression plasmids transfected using starburst PAMAM den- drimers. Nucleic Acids Res. 24, 2176–2182.
Bielinska, A.U., Kukowska-Latallo, J.F., Baker, J.R., 1997. The interaction of plasmid DNA with polyamidoamine dendrimers: mechanism of complex formation and analysis of alterations induced in nuclease sensitivity and tran- scriptional activity of the complexed DNA. Biochim. Biophys. Acta 1353, 180–190.
Boas, U., Heegaard, P.M., 2004. Dendrimers in drug research. Chem. Soc. Rev.
33, 43–63.
Cloninger, M.J., 2002. Biological applications of dendrimers. Curr. Opin. Chem.
Biol. 6, 742–748.
Dahle, C.E., Macfarlane, D.E., 1993. Isolation of RNA from cells in culture using Catrimox-14 cationic surfactant. Biotechniques 15, 1102–1105.
Dennig, J., Duncan, E., 2002. Gene transfer into eukaryotic cells using activated polyamidoamine dendrimers. J. Biotechnol. 90, 339–347.
Dufes, C., Uchegbu, I.F., Schatzlein, A.G., 2005. Dendrimers in gene delivery.
Adv. Drug Deliv. Rev. 57, 2177–2202.
Esfand, R., Tomalia, D.A., 2001. Poly(amidoamine) (PAMAM) dendrimers:
from biomimicry to drug delivery and biomedical applications. Drug Discov.
Today 6, 427–436.
Hong, S., Bielinska, A.U., Mecke, A., Keszler, B., Beals, J.L., Shi, X., Balogh, L., Orr, B.G., Baker, J.R., Banaszak Holl, M.N., 2004. Interaction of poly(amidoamine) dendrimers with supported lipid bilayers and cells: hole formation and the relation to transport. Bioconjug. Chem. 15, 774–782.
Kitchens, K.M., El-Sayed, M.E., Ghandehari, H., 2005. Transepithelial and endothelial transport of poly(amidoamine) dendrimers. Adv. Drug Deliv.
Rev. 57, 2163–2176.
Kobayashi, H., Brechbiel, M.W., 2005. Nano-sized MRI contrast agents with dendrimer cores. Adv. Drug Deliv. Rev. 57, 2271–2286.
Macfarlane, D.E., Dahle, C.E., 1997. Isolating RNA from clinical samples with Catrimox-14 and lithium chloride. J. Clin. Lab. Anal. 11, 132–139.
Manunta, M., Tan, P.H., Sagoo, P., Kashefi, K., George, A.J., 2004. Gene delivery by dendrimers operates via a cholesterol dependent pathway. Nucleic Acids Res. 32, 2730–2739.
Omidi, Y., Hollins, A.J., Drayton, R.M., Akhtar, S., 2005. Polypropylenimine dendrimer-induced gene expression changes: the effect of complexation with DNA, dendrimer generation and cell type. J. Drug Target. 13, 431–443.
Rosenthal, N., 1987. Identification of regulatory elements of cloned genes with functional assays. Methods Enzymol. 152, 704–720.
Ruponen, M., Honkakoski, P., Ronkko, S., Pelkonen, J., Tammi, M., Urtti, A., 2003. Extracellular and intracellular barriers in non-viral gene delivery. J.
Control. Release 93, 213–217.
Sambrook, J., MacCallum, P., Russell, D. (Eds.), 2001. Molecular Cloning: A Laboratory Manual, 3rd ed. Cold Spring Harbor Laboratory Press, NY, USA (Chapter 7).
Svenson, S., Tomalia, D.A., 2005. Dendrimers in biomedical applications—
reflections on the field. Adv. Drug Deliv. Rev. 57, 2106–2129.
390 J.-H.S. Kuo, Y.-L. Lin / Journal of Biotechnology 129 (2007) 383–390
Tang, M.X., Redemann, C.T., Szoka, F.C., 1996. In vitro gene delivery by degraded polyamidoamine dendrimers. Bioconjug. Chem. 7, 703–714.
Tang, M.X., Szoka, F.C., 1997. The influence of polymer structure on the inter- actions of cationic polymers with DNA and morphology of the resulting complexes. Gene Ther. 4, 823–832.
Wu, J., Zhou, J., Qu, F., Bao, P., Zhang, Y., Peng, L., 2005. Polycationic den- drimers interact with RNA molecules: polyamine dendrimers inhibit the catalytic activity of Candida ribozymes. Chem. Commun. 3, 313–315.
Zhao, H., Li, J., Xi, F., Jiang, L., 2004. Polyamidoamine dendrimers inhibit binding of Tat peptide to TAR RNA. FEBS Lett. 563, 241–245.