國立臺灣大學生物資源暨農學院植物病理與微生物學系 碩士論文
Department of Plant Pathology and Microbiology College of Bioresources and Agriculture
National Taiwan University Master Thesis
臺灣蟲草屬真菌之類緣關係及生物 活性物質之研究
Phylogeny of Cordyceps species and their bioactive compounds study in Taiwan
莊偉裕 Wei-Yu Chuang
指導教授:歐海仁 博士
Advisor: Hiran A. Ariyawansa, Ph.D.
中華民國 109 年 6 月
June 2020
ACKNOWLEDGEMENTS
First of all, I would like to express my gratitude towards my supervisor Dr. Hiran A. Ariyawansa for giving me a chance to explore the fascinating fungal kingdom.
Dr. Hiran allows me to study my interested topic and always does his best to support me. He encourages me when I am in depression and enlightens me when I am in confusion. Without his thoughtful guidance and meticulous attention, I might not be able to appreciate the beauty of fungi or complete my Master’s degree.
I would like to thank Dr. Chia-Lin Chung for generously allowing me to freely use Laboratory equipment to study my interested fungi when I was an undergraduate. Dr. Chung always supports students in need and is very welcome to any idea. Her valuable advices pinpoint my shortcomings and gave me courage to overcome. I also want to appreciate Dr. Tang-Long Shen for his kindly suggestion and insight in my study. Every talks with Dr. Shen broaden my horizon of knowledge and inspire my imagination towards the potential of fungi.
I would like to thank the committee members: Dr. Hiran A. Ariyawansa, Dr.
Shean-Shong Tzean, Dr. Tang-Long Shen, Dr. Ka-Lai Pang and Dr. Sung-Yuan Hsieh, who accepted our invitation. Dr. Tzean and Dr. Shen always support me with their extensive knowledge of fungal kingdom during the research study. Dr.
Pang and Dr. Hsieh have provided valuable suggestions and supports for the
I specifically thank the members of Taiwan Forest Research Institute and Forest Bureau for permitting my collection and their kindly support. I am also grateful for all the assistance and company from the members of Dr. Ariyawansa Laboratory, especially Mr. Xiao-Hua Yan, Mr. Kuei-Jr Liao and Ms. Ichen Tsai. I appreciate all the kind help of Mr. Shih-Cheng Wang and Ms. Dan-Rong Jhuang for helping the chemical analyses. Moreover, I want to thank all the kind help from Ms. Ya-Ni Tsai, Mr. Yung-Chin Cheng, and Ms. Mei-Hua Yan in our department office. Further, I want to thank all of my classmates, especially Mr.
Kuan-Chuan Tseng, Ms. Yue-Jie Chen and Ms. Chia-Min Hsieh for their company and assistance during these years.
In addition, I would like to thank following citizen scientists: Ms. Mei-Ling Lo, Ms. Yueh-Tseng Wei, Ms. Mei-Ju Chen, Mr. Li-jen Chu, Ms. Yueh-Yun Tang, Mr. Chia-Ming Hu, Mr. Chun-Yi Chen, Mr. Gaus Shang, Mr. Chun-Yu Lee for their kindly supply of studied materials. But for their assistance on materials collection, the study could not be completed thoroughly.
Finally, I want to express my gratitude to my dear family. It is their
understanding and support that I could fully involve in my research without any
worry. With their unconditional love and care, I am not afraid to dream a little
bigger, to explore the marvelous world and to conquer the unknown challenge
ahead.
摘要
蟲草屬類群(Cordyceps-like)真菌廣泛地作為藥用用途或食品添加劑,在中華歷史及西藏
中草藥文化中,作為珍貴藥材享譽盛名。一般而言,蟲草屬類群真菌可透過以下特點與 其他真菌進行區分:柄狀子囊座、子囊殼排列方式、子囊的形態特徵及寄主。臺灣有關蟲 草屬(Cordyceps)真菌之相關研究屈指可數並零星散佈於歷史文獻中。蟲草屬真菌能產生 各式生物活性物質,例如: 蟲草素(Cordycepin)、腺苷(Adenosine)和各種酵素,也因此蟲
草屬真菌常作為增能劑與民俗藥物,在傳統中醫學中更被認為有利於癌症與糖尿病的治 療。本研究目的有三: (一)依據形態與親緣演化評估台灣地區之蟲草屬真菌之自然分類地 位。(二)透過高效液相層析儀探討台灣蟲草屬真菌之菌絲溫水萃取液之蟲草素與腺苷含 量。(三)比較台灣地區新採集之蟲草屬真菌與市面商用菌株之蟲草素與腺苷含量。研究
期間,於臺灣各處採集共獲 59 株蟲草屬菌株,並於採後進行形態構造紀錄。利用單對
與合併的基因序列片段(nrLSU、ITS、tef1-α、rpb1、rpb2)進行親源演化分析並評估自然 分類地位。結合親源演化分析與形態鑑定,共發現七種蟲草屬新種(Cordyceps sp. nov. 1–
7)與四隻蟲草屬新紀錄種(C. blackwelliae、C. lepidopterorum、C. jakajanicola 與 C. rosea)。
同時,我們還描述了C. rosea 完整世代的型態。高效液相層析結果表示所有新採集之蟲
草屬菌株皆會產生腺苷。然而,只有三種蟲草屬真菌具有產生蟲草素的能力,分別為C.
militaris、Cordyceps sp. nov. 4 (NTUCC 18-144)與 Cordyceps sp. nov. 7 (NTUCC 18-145)。
此外,我們的結果指出新採集之C. militaris (NTUCC 18-120) 比市售菌株具有更強的蟲
草素生合成能力。
關鍵字:蟲草屬、新種、親緣關係、蟲草素、腺苷
ABSTRACT
Cordyceps-like taxa are widely used for medicinal purpose or food additives and have been described as precious herbal medicine in ancient Chinese history and Tibetan social community.
Generally, Cordyceps taxa are characterized by well-developed often stipitate stromata, arrangement of perithecia, ascospore morphology and host affiliation. Knowledge of the Cordyceps species in Taiwan is based on relatively few records that are scattered throughout
the literature. Cordyceps species produce various bioactive compounds, such as cordycepin, adenosine and enzymes, which have been widely used as energizers and folk medicines that can be conducive to cancer and diabetes treatments in traditional Chinese medicines. The main purposes of this study were 1) to evaluate the natural classification of Cordyceps species in Taiwan based on morphology and phylogeny, 2) to investigate cordycepin and adenosine production of Cordyceps species in Taiwan by high-performance liquid chromatography (HPLC) based on the hot-water extract from its mycelium, 3) to compare the level of cordycepin production of the newly collected Taiwan Cordyceps isolates with commercially available Cordyceps species. A total of 59 fresh Cordyceps strains were collected and their morphological
characteristics were recorded. Phylogenetic reconstruction using single and multi-loci (nrLSU+ITS+tef1-α+rpb1+rpb2) DNA sequences data were used to evaluate the natural classification of the new strains. Within this phylogenetic framework and considering the diagnostic morphological characters, seven new species (Cordyceps sp. nov. 1–7) were described. In addition, four new records, namely (C. blackwelliae, C. lepidopterorum, C.
rosea for the first time. The results of our HPLC analyses showed that all isolates used in this
study produced adenosine. However, only three Cordyceps species, namely C. militaris, Cordyceps sp. nov. 4 (NTUCC 18-144) and Cordyceps sp. nov.7 (NTUCC 18-145) proposed in
this study were able to produce cordycepin. Moreover, our results indicated that C. militaris (NTUCC 18-120) collected in this study, produced a higher level of cordycepin compared to commercially available Ophiocordyceps sinensis and C. militaris.
Keywords: Adenosine, Cordycepin, Cordyceps, Novel species, Phylogeny
TABLE OF CONTENTS
ACKNOWLEDGEMENTS ... I
摘要 ... III ABSTRACT ... IV
LIST OF TABLES ... VIII
LIST OF FIGURES ... IX
CHAPTER ONE: LITERATURE REVIEW... 1
Historical outline ... 1
Molecular studies and modern classification ... 8
Pharmacological implications of Cordyceps sensu lato ... 11
Adenosine and Cordycepin compounds ... 15
CHAPTER TWO: PHYLOGENY AND TAXONOMY ... 21
Introduction ... 21
Materials and methods ... 24
Fungal collecting, isolation and morphological examination... 24
DNA extraction and PCR amplification ... 26
Phylogenetic analyses ... 29
Results ... 39
Distribution and fungal isolations ... 39
Phylogeny ... 42
Taxonomy ... 43
Discussions ... 82
CHAPTER THREE: IDENTIFICATION OF CORDYCEPIN AND ADENOSINE FROM THE Cordyceps spp. IN TAIWAN ... 86
Introduction ... 86
Materials and methods ... 88
Fungal extract preparation ... 88
HPLC detection ... 89
Statistical analyses ... 90
Results ... 90
Detection and identification of concentrations of Adenosine and Cordycepin .... 90
Discussion ... 99
GENERAL CONCLUSION ... 101
REFERENCES ... 103
APPENDIX ... 124
APPENDIX A: Single-gene phylogenetic tree... 124
APPENDIX B: Chromatogram of 51 Cordyceps and 1 Ophiocordyceps sinensis ... 129
APPENDIX C: Media composition ... 133
APPENDIX D: Sequences of newly introduced Cordyceps species in this study. ... 136
LIST OF TABLES
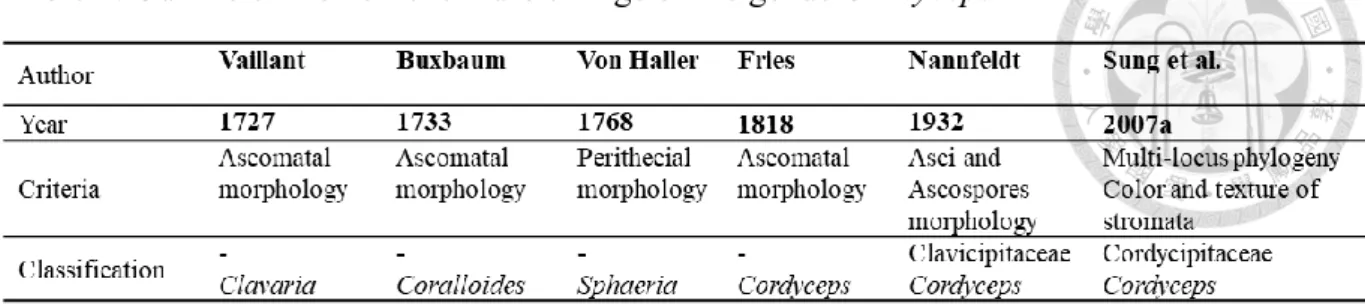
Table 1. Outline of the nomenclature change of the genus Cordyceps ... 2
Table 2. Primer sets and PCR condition used to amplify the five gene loci (nrLSU, ITS, tef1-α,
rpb1 and rpb2). ... 28
Table 3. Details of isolates included in the phylogenetic analyses. ... 31
Table 4. Evolutionary nucleotide substitution models used in the phylogenetic analyses by
MEGA v. 7.0.26. ... 39 Table 5. Cordycepin concentration of seven cordycepin producing isolates and
Ophiocordyceps sinensis. ... 94 Table 6. Adenosine concentration of 51 Cordyceps isolates and one Ophiocordyceps sinensis
strain... ... 95
LIST OF FIGURES
Figure 1. Chemical structure of Adenosine ... 16
Figure 2. Chemical structure of Cordycepin ... 17
Figure 3. Map of Taiwan showing the areas surveyed in this study. ... 26
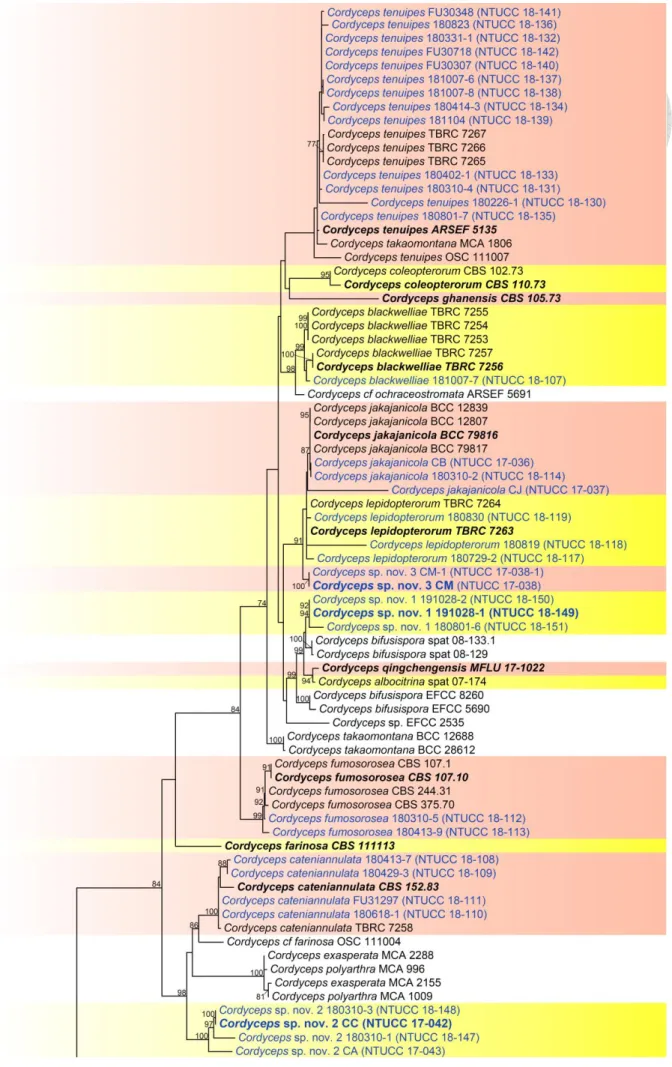
Figure 4. RAxML tree of Cordyceps based on a combined dataset of ITS, nrLSU, tef1, rpb1 and rpb2 gene regions. ... 40
Figure 5. Cordyceps tenuipes. ... 45
Figure 6. Cordyceps fumosorosea. ... 47
Figure 7. Cordyceps ninchukispora. ... 49
Figure 8. Cordyceps militaris. ... 51
Figure 9. Cordyceps javanica. ... 53
Figure 10. Cordyceps cateniannulata. ... 55
Figure 11. Cordyceps blackwelliae. ... 57
Figure 12. Cordyceps lepidopterorum. ... 59
Figure 13. Cordyceps jakajanicola. ... 61
Figure 14. Cordyceps rosea. ... 64
Figure 15. Cordyceps sp. nov. 1. ... 69
Figure 16. Cordyceps sp. nov. 2 ... 67
Figure 17. Cordyceps sp. nov. 3. ... 77
Figure 18. Cordyceps sp. nov. 4. ... 74
Figure 19. Cordyceps sp. nov. 5. ... 79
Figure 20. Cordyceps sp. nov. 6. ... 71
Figure 21. Cordyceps sp. nov. 7 ... 82
Figure 22. Chromatogram of Standard chemical-Adenosine and Cordycepin... 92
Figure 23. HPLC chromatogram of Cordycepin producing fungi and Ophiocordyceps sinensis. ... 93
Figure 24. Cordycepin concentration of seven cordycepin producing isolates and Ophiocordyceps sinensis presented in a bar chart based on Table 5.. ... 95
Figure 25. Adenosine content of 51 Cordyceps isolates and one Ophiocordyceps sinensis strain presented in a bar chart based on Table 6. ... 99
CHAPTER ONE: LITERATURE REVIEW
Historical outline
The genus Cordyceps (Ascomycota, Sordariomycetes, Hypocreales, Clavicipitaceae) was established by Fries (1818) and typified by C. militaris. However, Cordyceps militaris was initially introduced as Clavaria militaris by Vaillant (1727). The last International Botanical Congress (IBC) in Melbourne has brought about some drastic changes in fungal nomenclature (Hawksworth 2011, Norvell 2011). Out of those, especially the introduction of a unified nomenclature for pleomorphic fungi, has been discussed intensively. Thereafter Cordyceps species has been undergone numerous taxonomic and nomenclatural changes, leaving the natural classification and nomenclature of the Cordyceps and allied species in doubt (Table 1) (Rogerson 1970, Shrestha et al. 2014). The genus was initially characterized by clavate, cylindrical or filamentous stromata, cylindrical ascus with thickened ascus cap and filiform, multi-septate or aseptate ascospores (Kobayasi 1941, 1983). Traditionally, Cordyceps was comprised around 400 species that occurred in various hosts and habitats e.g. as parasites on invertebrate and few fungal genera such as Elaphomyces, can be found on the soil as saprobes or even as endophytes in various plant hosts (Bischoff and White 2003, Humber 2008, Kobayasi and Shimizu 1963, Shrestha et al. 2016, Sung et al. 2007a).
Table 1. Outline of the nomenclature change of the genus Cordyceps
Traditional classification of cordyceps-like taxa were primarily based on host affiliation, morphology of ascomata and microscopic structures such as asci, ascospore and ascus cap. For example, Cordyceps cicadae was characterized mainly based on its host (Cicada), while Cordyceps bifusispora was introduced as a new taxon considering its peculiar bifusiform
ascospores (Kobayasi 1941, Eriksson 1982). Besides, perithecia and ascospore disarticulation were also considered as important taxonomic characters during classification of closely-related species (Kobayasi 1941, 1982, 1983, Mains 1958, Shrestha et al. 2005, Sung et al. 2007a).
However, separation of Cordyceps species based entirely on host affiliation and morphology might be debatable due to various reasons such as 1) Same Cordyceps species could parasitize different hosts. For example, Cordyceps militaris on Tenebrionidae and Curculionidae (Bary 1867, Kryukov et al. 2011, Shrestha et al. 2016); 2) More than one species can parasitize the same host. For example, Cordyceps jakajanicola and Cordyceps cicadae on Cicada (Crous et al. 2019, Mains 1958); 3) Immature ascospores show septate without showing disarticulation such as Cordyceps militaris (Sung et al. 2007a).
On the other hand, several scientists noticed that the morphology of the asexual stage of Cordyceps species resembled to non-related asexual fungal genera such as Beauveria,
Evlachovaea, Isaria, Lecancillium, Metarhizium, Paecilomyces and so on (Zare and Gams 2001,
Huang et al. 2005, Liu et al. 2001). For example, Cordyceps bassiana, Cordyceps scarabaeicola and Cordyceps brongniartii all produced subglobose to flask-shaped conidiogenous cells in the culture with indeterminate, denticulate rachis on apex, which was morphologically similar to Beauveria species (Rehner and Buckley 2005). Additionally, Cordyceps taii was thought to be
the sexual morph of Metarhizium taii based on the microcycle conidiation features (Liang et al.
1991). Moreover, Cordyceps brittlebankisoides was compared with fungal species in Metarhizium genus and initially considered as sexual morph of Metarhizium anisopliae var.
majus, but later linked to Metarhizium flavoviride and finally synonymized under Metarhizium brittlebankisoides (Huang et al. 2005, Liu et al. 2001, Kepler et al. 2014).
With molecular techniques becoming more popular, several scientists conducted phylogenetic studies to refine the classification of Cordyceps and allied taxa. Based on 18S and 28S ribosomal DNA loci phylogenetic analyses, Artjariyasripong et al. (2001) recognized four clades within Clavicipitaceae and concluded that strains classified under the genus Cordyceps were not monophyletic. Sung et al. (2001) conducted similar work but mainly emphasized on asexual-sexual relationships within the family Clavicipitaceae and also concluded that Cordyceps genus did not form a monophyletic clade within the family. Later, Stensrud et al.
(2005) used internal transcribed spacer (ITS) region of ribosome DNA loci to resolve the asexual-sexual connections of Cordyceps genus and found that Cordyceps taxa were
The most detailed and comprehensive study of cordyceps-like taxa based on both multi- gene phylogeny and morphology was done by Sung et al. (2007a). Sung et al. (2007a) studied 162 cordyceps-like taxa based on analyses comprising of five to seven loci, containing the nuclear ribosomal small and large subunits (nrSSU and nrLSU), the elongation factor 1α (tef1- α), the largest and the second largest subunits of RNA polymerase II (rpb1 and rpb2), β-tubulin,
and mitochondrial ATP6 (mtATP6). During the investigation, Sung et al. (2007a) discovered that both Clavicipitaceae and Cordyceps were not monophyletic. Therefore, two extra families, Cordycipitaceae and Ophiocordycipitaceae, were introduced and species previously classified in Cordyceps were maintained as affiliates of all three families. Clavicipitaceae and Ophiocordycipitaceae together formed a monophyletic group, however Cordycipitaceae, based on the phylogenetic placement of the type species of Cordyceps, C. militaris, shared a more recent common ancestor with Hypocreaceae (Kepler et al. 2017).
At the same time, four new genera were erected from the species which were formerly classified in Cordyceps, i.e., Ophiocordyceps and Tolypocladium were introduced to place Cordyceps blattae Petch (1924) and Cordyceps subsessilis Petch (1937) respectively and
classified under Ophiocordycipitaceae. Metacordyceps and Tyrannicordyceps were introduced to place Cordyceps taii Liang & Liu (1991) and Cordyceps fratricida Tanda & Kobayasi (1984) respectively and classified under Clavicipitaceae (Kepler et al. 2012, Sung et al. 2007).
In agreement with the reframing of Article 59 of the International Code of Nomenclature
reviewed by Kepler et al. (2017) to resolve competing names based on principles of priority, recognition of monophyletic clades, and the practical usage of affected taxa. Nine generic names were proposed to retain in the family, including Akanthomyces Lebert, Ascopolyporus Möller, Beauveria Vuill., Cordyceps Fr., Engyodontium de Hoog, Gibellula Cavara, Hyperdermium J.F. White et al., Parengyodontium C.C. Tsang et al. and Simplicillium W. Gams
& Zare. In contrast, Isaria, Evlachovaea, Microhilum and Phytocordyceps were proposed synonymized under Cordyceps because their type species were nested within the genus Cordyceps. Besides, two new genera Blackwellomyces Spatafora & Luangsa-ard and Hevansia
Luangsa-ard, Hywel-Jones & Spatafora were introduced to place Cordyceps cardinalis G.H.
Sung & Spatafora (2004) and Akanthomyces novoguineensis Samson & B.L. Brady (1982), respectively.
To date, the monophyly of Cordyceps sensu stricto is restricted to arthropods-pathogenic fungal taxa, classified under Cordycipitaceae and accommodates around 50 species (Shrestha et al. 2017, Spatafora et al. 2007, Sung et al. 2007a, b). The majority of taxa belongs to Cordyceps sensu stricto grown on larva and pupa of Lepidoptera and Coleoptera species, while
some species have the ability to colonize the nymph of Cicada (Mongkolsamrit et al. 2018, Sung et al. 2007a), whereas few species have been reported as pathogens on Hymenoptera (Mongkolsamrit et al. 2018, Sung et al. 2007a). The sexual morph of Cordyceps sensu stricto produce pallid to brightly colored, soft, fleshy stromata (Sung et al. 2007a, b). Perithecia
filiform, multi-septate or aseptate ascospores, disarticulating into part-spores or non- disarticulating (Kepler et al. 2017, Sung et al. 2007a, b). Asexual morph of Cordyceps sensu stricto is hyphomycetous and identified as conidiophores arising from the synnemata or hyphae,
single or consisting of verticillate phialides in whorls. Phialides possess swollen basal portion tapering into a distinct neck. The top of phialide bears powdery, unicellular, subglobous to cylindrical conidia (Kepler et al. 2017, Mongkolsamrit et al. 2018, Sung et al. 2007a, b).
Conidiogenesis pattern can be evlachovaea-like and isaria-like (Humber et al. 2013, Mongkolsamrit et al. 2018). The former refers to successive conidia produced in alternate orientations resulting in zipper-like chains, while the latter refers to successive conidia produced in end-to-end orientations resulting in straight chains (Humber et al. 2013, Mongkolsamrit et al. 2018). Some Cordyceps spp. were observed to have both types of conidiogenesis pattern, which is believed to be associated with different nutrition resources (Mongkolsamrit et al. 2018).
Taiwan is a unique island with a rich display of diverse flora and animal species.
Knowledge of the Cordyceps genus in Taiwan has a long history, which could date back to the study of Shimizu (1976). However, the information of Cordyceps was scattered all over the literatures. Tzean et al. (1997) systematically introduced the diversity of Cordyceps in Taiwan based on morphological characteristics. A total of 831 entomopathogenic fungi were collected in Taiwan during 1988 to 1996. In total, 24 genera consisting of 66 different species were
(Tzean et al. 1997). To date, based on current classification of Cordyceps sensu stricto, only eleven Cordyceps were reported in Taiwan, including C. takamontana, C. tuberculata, C.
fumosorosea, C. farinosus, C. javanica, C. tenuipes, C. ninchukispora, C. cicadae, C. militaris, C. cateniannulata and C. amoene-rosea (Chen 1978, Su and Wang 1986, Wang 1988, Tzean et al. 1997).
Molecular studies and modern classification
Cordyceps-like fungi are usually defined based on the morphological variation in relatively small number of characters. Problems can be encountered while defining the species limits, especially among closely related morpho-species within a fungal community, due to the lack of distinctive morphological variation (especially in the absence of a sexual stage), the interdependence of the morphological characters, and the environmental influences on morphology (Humber et al. 2013, Mains 1958, Mongkolsamrit 2018, Shrestha et al. 2016, Sung et al. 2007). For instance, species such as Beauveria brongniartii, Beauveria sungii, Blackwellomyces cardinalis and Blackwellomyces pseudomilitaris produce similar fruiting
body and sexual morphs to Cordyceps species (Kepler et al. 2017, Mongkolsamrit 2018).
However, this situation can be more complicated if the species are described only based on asexual morph because a wide range of fungal taxa produce similar sporulating structure similar to the asexual stage of Cordyceps (Gams et al. 1998, Humber 2013, Luangsa-Ard et al. 2005).
In recent decades, molecular taxonomy based on phylogenetic methods have extended our knowledge regarding the diversity and classification of Cordyceps and allied taxa.
Phylogenetic tree inferred from molecular data provides an explicit and reliable systematic framework to resolve the fungal species sharing similar morphology. The earliest study based on molecular data for cordyceps-like taxa was done by Spatafora and Blackwell (1993). During their study Spatafora and Blackwell (1993) performed cladistics analyses using partial region
of nrSSU to clarify the connection between Clavicipitales and Hypocreales, and proposed Clavicipitales as a monophyletic group which formed a sister clade to Hypocreales. However, this proposal was not supported by Gams et al. (1998) and Sung et al. (2001). Gams et al. (1998) indicated Hypocreales and Clavicipitales exhibited a paraphyletic relationship in phylogenetic tree inferred from the combined nrSSU and nrLSU gene regions. Sung et al. (2001) conducted a similar study and found neither Cordyceps nor Clavicipitaceae formed a monophyletic clade within Hypocreales.
Recent phylogenetic studies based on multi-gene phylogenetic analyses rejected monophyly of Clavicipitaceae but supported the separation of Clavicipitaceae into three lineages and presented as Cordycipitaceae, Ophiocordycipitaceae and Clavicipitaceae (Sung et al. 2007a, b, Spatafora et al. 2007). This classification was followed by the later researchers (Kepler et al. 2013, 2017, Quandt et al. 2014, Spatafora et al. 2007, Wei et al. 2019). Sung et al.
(2007b) conducted phylogenetic analyses including ninety-one taxa representing several families in Hypocreales based on seven different DNA loci: nrSSU, nrLSU, tef1-α, β-tubulin, rpb1 and rpb2, and mtATP6. All seven separate analyses rejected the monophyly of
Clavicipitaceae. The separate analyses of nrSSU, nrLSU and β-tubulin did not resolve the Clavicipitaceae family. The phylogenetic trees inferred from rpb1 and rpb2 and the combined- gene phylogeny of seven genes supported the separation of Clavicipitaceae into three monophyletic clades which were later designated as Clavicipitaceae, Cordycipitaceae and
Ophiocordycipitaceae. In contrast, mtATP6 single gene tree did not support for the monophyly of Ophiocordycipitaceae. Similar conclusion was made by Spatafora et al. (2007) using same genic regions without including mtATP6 locus.
Following previous literatures, Kepler et al. (2017) conducted a phylogenetic analyses based on five different gene loci (nrSSU, nrLSU, tef1-α, rpb1 and rpb2) to revise the classification of Cordycipitaceae mainly focusing on reconciliation of competing names.
Remarkably, the generic types of Evlachovaea (E. kintrischica), Microhilum (M. oncoperae) and Phytocordyceps (P. ninchukispora) were nested in Cordyceps genus. Therefore Kepler et al. (2017) proposed to synonymize Evlachovaea, Microhilum and Phytocordyceps under Cordyceps. Besides, Isaria was also proposed to suppress as well because the ex-epitype isolate
of I. farinosa (CBS 111113) was clustered within Cordyceps genus. Furthermore, Cordyceps cardinalis and Cordyceps pseudomilitaris were classified under new genus Blackwellomyces
(Kepler et al. 2017).
In recent studies, several new Cordyceps species were introduced from Thailand and China namely Cordyceps jakajanicola, Cordyceps kuiburiensis, Cordyceps blackwelliae, Cordyceps lepidopterorum and Cordyceps qingchengensis (Crous et al. 2019, Mongkolsamrit
et al. 2018, Zha et al. 2019). To date, the genus Cordyceps accommodates approximately 50 species which have been identified by both phylogeny and phenotypic characters (Crous et al.
2019, Mongkolsamrit et al. 2018, Shrestha et al. 2018, Zha et al. 2019).
Pharmacological implications of Cordyceps sensu lato
Cordyceps-like species have a long history of use as folk medicine in China and South-East Asia. Cordyceps-like species had its first pharmacological record in 5th century in Lei's Treatise on Preparation of Materia Medica and later mentioned in several literatures of Chinese medicinal books. Cordyceps species such as Cordyceps cicadae, Cordyceps militaris, Cordyceps ophioglossoides (Current name: Tolypocladium ophioglossoides), Cordyceps sobolifera (Current name: Ophiocordyceps sobolifera), Cordyceps sinensis (Current name:
Ophiocordyceps sinensis) have been identified to produce various chemical compounds
benefiting human health (Tuli et al 2014, Wang et al. 2015, Lee at al. 2019). One of the well- known caterpillar killer fungus Ophiocordyceps sinensis was also traditionally classified in Cordyceps and is known as Tibetan folk tonic “Yartsa Gunbu” or “Dong Chong Xia Cao” in
traditional Chinese medicine. Ophiocordyceps sinensis was officially reported in Ben Cao Bei Yao (Complete Essentials of the Materia Medica) written by Wang Yang in 1683 for its potential
performance to nourish kidney and revitalize lung. Native occurrence of the fungus is confined to the Tibetan Plateau and adjoining Himalayas including parts of India and western China at altitudes of over 3,000 meters (Chen et al. 2000). The larvae of Thitarodes spp. play a crucial role in the fungal ecology as Ophiocordyceps sinensis has been isolated from more than 40 species of Thitarodes (Wang and Yao 2011). The fungus and the dead body of caterpillar collectively considered as a valuable Chinese medicine (Wang and Yao 2011).
Ophiocordyceps sinensis. It has been reported that the compounds from the crude extracts of Ophiocordyceps sinensis can modulate immunity system of human (Koh et al. 2002, Zhang et
al. 2011), inhibit the growth of tumor cells of mice (Cai et al. 2018, Sheng et al. 2011), suppress the lipid biosynthesis (Guo et al. 2010), boost the anti-aging effect (Ji et al. 2009), improve the anti-oxidation activity and reduce the inflammation (Liu et al. 2011, Wang et al. 2012).
Main constituent of the extract derived from fungal species in Cordycipitaceae and Ophiocordycipitaceae comprise a biologically active metabolite cordycepin (Tuli et al 2014, Wang et al. 2015, Lee at al. 2019). Cordycepin is commonly known to be produced in Cordyceps militaris and Ophiocordyceps sinensis but the amount of cordycepin produced in O.
sinensis has been reported to be comparatively low (Tuli et al 2014, Wang et al. 2015, Lee at al.
2019). Other than cordycepin, these fungal species have ability to produce various amino acids, nucleotides, polyamines, cyclic dipeptides, saccharides, sugar derivatives, sterols and etc. (Liu et al. 2015). For examples, cordycepic acid (D-mannitol) shows great anti-oxidant ability;
ergosterol possesses antimicrobial activity; cordyglucans exhibits potent anti-tumor activity, which could be associated with its (1 → 3)-𝛽-D-glucan linkages. Cordycepin has anti-
inflammatory, anti-bacteria and anti-tumor effect with little toxicity towards normal cell of human (Tuli et al. 2013, Yalin et al. 2005). The amount of each chemical produced could vary due to different nutrition or culture methods such as the wild fruiting body, solid-state fermentation and submerged fermentation (Hsu et al. 2003, Wang et al. 2015).
regarding their various properties that can be used to improve the human health. The price of Ophiocordyceps sinensis can vary from US$18726 to US $30250 (Cunningham and Long 2019).
The wild fruiting body or powder of Ophiocordyceps sinensis and the related products are available in various forms such as, capsule, beverage and facial mask, which are mostly derived from submerged fermentation (Zhou et al. 2009). Among these products, the fruiting body of the fungus has the highest price because it is hard to induce artificially and also possesses more diverse chemical composition and higher content of cordycepin than submerged fermentation products (Lo et al. 2013). Due to limited distribution, high price, overexploitation, and difficult to induce in artificial culture, the well known Ophiocordyceps sinensis have become endangered (Caplins and Halvorson 2017, Wang and Yao 2011, Zhang et al. 2012). Therefore, the phylogenetically related species, Cordyceps militaris, has been identified as a substitution for Ophiocordyceps sinensis.
Even though Cordyceps militaris has similar pharmacological properties to Ophiocordyceps sinensis, however, it shows a very different morphology and life cycle.
Cordyceps militaris presents in diverse geographic regions and various ecological niches. It
mainly parasitizes lepidopteran larvae and pupae but a few records have been reported on species classified in orders such as Coleoptera and Hymenoptera (Shrestha et al. 2012a).
Cordyceps militaris is known as “Golden Chong Cha” because of its flesh cylindrical to clavate and yellow to light orange fruiting body. Cordyceps militaris can complete its life cycle on
conditions. (Kobayasi 1941, Lin et al. 2017, Shrestha et al. 2012b). Materials such as rice, bean powder, corn powder, wheat grain, cottonseed shell, silkworm pupae can be used to induce the fruiting body of Cordyceps militaris in-vitro (Hong et al. 2010, Huang et al. 2009, Lin et al.
2017, Shrestha et al. 2012).
With technological advancement in chemical analyses, a variety of compounds have been identified in Cordyceps militaris. Cordycepin was the first compound purified from Cordyceps militaris which was elucidated as 3'-deoxyadenosine, an analogue of adenosine
(Cunningham et al. 1950). Polysaccharides such as CPS1, CPS2, CPS3, P50-1, P70-1, P70-2 have also been isolated from water extract of Cordyceps militaris (Yu et al. 2004, 2007). Among these carbohydrate molecules, CPS1 shows the anti-inflammatory activity and P70-1 possesses the hydroxyl radical-scavenging activity (Yu et al. 2004, 2007).
High-performance liquid chromatography (HPLC) is one of the main resources in quality checking of food and medicine products. It has been commonly used in diversity determination (Huang et al. 2009). Until now, there are innumerable HPLC methods have been used in the determination of adenosine and cordycepin from cordycipitaceous taxa such as Ophiocordyceps formosana, O. sinensis, and Cordyceps militaris (Guo et al. 2006, Li et al.
2006, Huang et al. 2003, Wang et al. 2015).
Adenosine and Cordycepin compounds
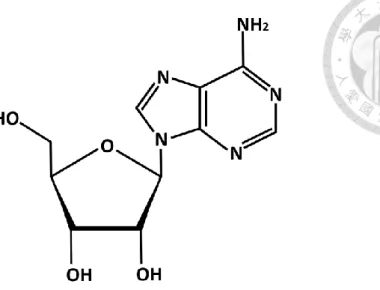
There have been a variety of bioactive compounds reported from Cordyceps species such as adenosine, cordycepin, D-mannitol, hypoxanthine, ergosterol, cordyheptapeptide, cordyxanthin and cordymin (Cunningham et al. 1950, Dong et al. 2013, Guo et al. 1998, Huang et al. 2003, Rukachaisirikul et al. 2006, Wonga et al. 2011, Yuan et al. 2007). Among these compounds, cordycepin and adenosine have received the great attention. Adenosine is a ribonucleotide, consisting of adenine bound to ribose, which is widely present in Cordyceps species (Figure 1) (Huang et al. 2009, Chu et al. 2015). Adenosine plays pleiotropic roles in the biological organism and can be existed in various kinds of forms in cells. For instances, phosphorylated forms of adenosine are involved in cellular energy transfer by adenosine triphosphate (ATP) and adenosine diphosphate (ADP) (Knowles 1980). Adenosine also plays a vital role in genetic material synthesis as a major component of DNA and RNA (Paton 1988). Besides, adenosine can exert signal transduction intracellularly or as cyclic adenosine monophosphate (cAMP) in intracellular signal transduction. In the human central nervous system, adenosine modulate several neurotransmitters and protecting neurons under ischemic and hypoxic stressful conditions (Haskó and Cronstein 2004, Ralevic and Burnstock 1998, Sebastião and Ribeiro 2000).
Figure 1. Chemical structure of adenosine
Adenosine also has various pharmacological properties. Adenosine has the ability to induce human transient heart block and slow down the heart rate (Pelleg and Belardinelli 1993).
This property makes adenosine to use as an antiarrhythmic drug for reentrant supraventricular tachycardia and in the anesthesiology clinical practices such as cardiothoracic anesthesia, critical care medicine (Eltzschig 2009, Pelleg and Belardinelli 1993, Stemp and Roy 1992, Trohman 2000). In addition to its antiarrhythmic property, adenosine also plays an important role in vasodilation and blood flow regulation (Eltzschig 2009). A recent study found that extracellular adenosine could suppress the growth of liver cancer and gastric cancer cells by induction of apoptosis (Ma et al. 2014, Saitoh et al. 2004). Similar results were observed in human colonic and epithelial cancer cells with the additions of low concentration of adenosine and adenosine deaminase inhibitors (Barry and Lind 2000, Saito et al. 2010).
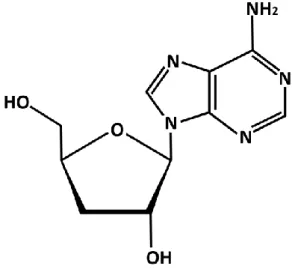
The bioactive compound cordycepin (Figure 2) was first isolated from Cordyceps militaris by Cunningham et al. (1950). Cordycepin or 3'-deoxyadenosine, 9-(3-Deoxy-β-D-
weight 251.24) and has a similar chemical structure with adenosine but differs from the latter by the absence of 3' hydroxyl group (Bentley et al. 1951, Radwan and Wilson 1980). Because of its similarity to adenosine, cordycepin has the ability to interfere synthesis of genetic materials. For example, during the process of transcription (RNA synthesis), cordycepin cannot be distinguished from adenosine by the enzymes and incorporated into the RNA sequences, leading to the premature termination of transcription (Holbein et al. 2009). Besides, cordycepin has been reported to interfere in purine biosynthesis by inhibiting activity of phosphoribosyl- pyrophosphate amidotransferase (Rottman and Guarino, 1964). Moreover, under low dosage cordycepin can inhibit the mRNA polyadenylation, leading to a decrease in cell proliferation.
In contrast, cordycepin in high dosage affected cell adhesion, and indirectly inhibited protein synthesis by affecting mTOR and AMPK signaling pathway (Wong et al. 2010).
Figure 2. Chemical structure of cordycepin
Cordycepin demonstrates multiple pharmacological properties including immunity system activation, anti-oxidation, anti-bacteria, anti-inflammatory and anti-cancer (Ramesh et
has received great attention due to its structural similarity to adenosine, as adenosine and adenosine receptors are correlated with tumor progression (Kazemi et al. 2018). Several studies indicated the anti-tumor property of cordycepin mainly depends on activation of apoptosis (Chaicharoenaudomrung et al. 2018, Hwang et al. 2017, Shao et al. 2016) and inhibition of metastatic activity (Noh et al. 2010). For example, Chaicharoenaudomrung et al. (2018) observed the induction of apoptosis after the treatment of cordycepin on the human brain cancer cell, which was attributed to upregulation of tumor suppressor gene and ROS generation. Noh et al. (2010) demonstrated cordycepin inhibited the cancer cell invasion by interfering with the MAPK signaling pathway, which was associated with the regulation of cell proliferation, apoptosis and MMPs production.
Cordycepin can be obtained either by living organisms or chemical synthesis. Cordyceps militaris is the first species to report the existence of cordycepin. Later, several species
classified in Cordyceps sensu lato were also reported to produce cordycepin such as Ophiocordyceps sinensis (≡Cordyceps sinensis) (Hsu et al. 2002), Cordyceps cicadae (≡
Paecilomyces cicadae) (Wang et al. 2012), Cordyceps pruinosa (Meng et al. 2014), Ophiocordyceps formosana (Cordyceps formosana) (Wang et al. 2015), Ophiocordyceps xuefengensis (Jin et al. 2017), Cordyceps kyusyuensis (Xia et al. 2017). However, the identifications of these fungal strains are debatable because for most studies identification were based on either morphology only or coupled with low-informative gene loci. For example,
single-gene phylogenetic tree. Apart from cordyceps-like fungi, a few studies have been reported to show the ability to produce cordycepin such as Aspergillus nidulans. For example, Kaczka et al. (1964) identified cordycepin from the broth of Aspergillus nidulans via Infrared (IR) and Nuclear Magnetic Resonance (NMR) spectra. Similarly, Kodama et al. (1979) also found Aspergillus nidulans produced cordycepin when cultured on wheat bran. Zhang et al.
(2006) compared the cordycepin production of fermented broth of Cordyceps militaris and Aspergillus nidulans, and concluded that Aspergillus nidulans can be used as a better resource
for cordycepin compared to Cordyceps militaris. However, recent studies have shown that among the cordycepin producing fungi, Cordyceps militaris has higher cordycepin yield rate in most of the cases (Yang and Dong 2011).
In addition to extraction from living organisms, several synthetic and semisynthetic pathways have been proposed for cordycepin production. Synthetic pathway reported by Todd and Ulbricht (1960), 3’-O-p-Nitrobenzenesulphonyladenosine served as a starting material, reacting with sodium iodide in hot acetonylacetone to form 3’-deoxy-3’-iodoadenosin. Later, 3’-deoxy-3’-iodoadenosin was converted to cordycepin by hydrogenation. All materials used in the pathway of Todd and Ulbricht (1960) were available in the market and the synthetic procedure was short but the yield rate was around 17% only. Aman et al. (2000) proposed that the synthesis started with adenosine and was transferred to 3’-bromo intermediate by using 2- acetoxyisobutyryl bromide. The 3’-bromo intermediate was then subjected to debromination,
al. (2000) received 20% cordycepin yield rate. However, this proposal has several drawbacks such as the shortage of acetoxyisobutyryl bromide and issues with the purification process.
Moreau et al. (2013) proposed another synthetic pathway. According to Moreau et al. (2013) proposal, Adenosine was first cyclized by trimethylorthoacetate and brominated with acetyl bromide and cordycepin was finally generated via debromination and deprotection reactions.
Even though the procedure showed a short synthetic route but the final yield is only 20% due to the low conversion of the bromination step. Li et al. (2013) developed a novel synthetic method for cordycepin production. The synthetic pathway started with D-glucose or D-xylose and followed by deoxygenation of 3’-OH via Barton–McCombie reaction to obtain the 3’- deoxyribose derivative. These 3’-deoxyribose derivative was then subjected to a series of steps including removal of protecting group and Vorbrüggen glycosylation to yield cordycepin. The reaction provided a high yield rate of 37–40% but the procedure was more complicated.
Overall, the synthetic pathways mentioned above had yield rate around 20–40% (Huang et al. 2017). However, the synthetic process is not cost-effective and produce environmental pollutions due to the usage of toxic catalyst and organic compounds. Furthermore, the chemical used are corrosive and toxic to the organism and the purification of the target compound is time- consuming and expensive. Synthetic pathways with low cost and high efficiency remain further study.
CHAPTER TWO: PHYLOGENY AND TAXONOMY
Introduction
The genus Cordyceps was established by Fries (1818) and typified by Cordyceps militaris.
Fungal species parasitizing mainly on invertebrate with unitunicate ascus containing filiform ascospores were initially classified in Cordyceps (Kobayasi 1941, 1983). Considering these morphological features and host affiliation, 400 sexually reproducing fungal species were initially classified in Cordyceps (Bischoff and White, 2003, Humber 2008, Kobayasi and Shimizu 1963, Shrestha et al. 2016, Sung et al. 2007a). Furthermore, perithecia arrangement and ascospore disarticulation were also taken into consideration during delimitations of closely- related species of Cordyceps (Kobayasi 1941, 1982, 1983, Mains 1958, Shrestha et al. 2005, Sung et al. 2007). Traditionally several asexual fungi morphologically similar to genera such as Beauveria, Evlachovaea, Isaria, Lecancillium, Metarhizium, Paecilomyces were also linked to Cordyceps (Gams and Zare 2001, Huang et al. 2005, Liu et al. 2001).
With the development of molecular techniques over the last two decades, new knowledge has become available thus several scientists conducted phylogenetic studies to refine the classification of cordyceps-like taxa (Cordyceps sensu lato) (Artjariyasripong et al.
2001, Sung et al. 2001, Stensrud et al. 2005, Spatafora et al. 2007, Sung et al. 2007a, b). Early publications based on phylogenetic analysis of ITS alone or combined with nrSSU and nrLSU regions were suggested that cordyceps-like taxa (Cordyceps sensu lato) are paraphyletic and
al. 2001, Stensrud et al. 2005, Sung et al. 2001). However, these studies did not fully resolve the natural classification of species in Cordyceps sensu lato because of the insufficient sampling and low informative DNA barcode regions (Stensrud et al. 2005, Sung et al. 2001). Spatafora et al. (2007) and Sung et al. (2007a, b) conducted comprehensive studies focusing on the classification of Clavicipitaceae family mainly concentrating on taxa classified under genus Cordyceps. These studies proposed to reject the monophyly of Clavicipitaceae as well as Cordyceps and divided Clavicipitaceae into three families (Cordycipitaceae, Clavicipitaceae
and Ophiocordycipitaceae) based on multi-gene phylogenetic tree generated from seven different DNA loci namely nrSSU, nrLSU, tef1-α, β-tubulin, rpb1 and rpb2, and mtATP6.
The family Cordycipitaceae was introduced to accommodate the clade containing the generic type of Cordyceps, C. militaris (Fries 1818). Most of the species classified in this group are pathogenic on arthropods (Sung et al. 2007a). According to the current classification, the sexual morph of Cordyceps sensu stricto is characterized by pallid to brightly colored, soft, fleshy stromata. Perithecia erect completely immersed to superficial with unitunicate cylindrical asci plus thickened ascus cap reside within perithecia. Ascospores are filiform, multi-septate or aseptate, disarticulating into part-spores or nondisarticulating (Kepler et al.
2017, Sung et al. 2007a, b). The asexual morph of Cordyceps sensu stricto is defined as hyphomycetes and characterized by conidiophores arising from the synnemata or hyphae.
Conidiophore could be single or consist of verticillate phialides, which are usually swollen at
(Kepler et al. 2017, Mongkolsamrit et al. 2018, Sung et al. 2007a, b).
Taiwan is an island with rich biodiversity. However, this rich treasure of diversity has been gradually destroyed due to recent overexploitation of natural resources and environmental pollution. A few studies have provided the groundwork for the Cordyceps diversity and classification in Taiwan with limited molecular data (Chen 1978, Kobayasi and Shimizu 1981, Su and Wang 1986, Tzean et al. 1997). According to recent classification, only nine Cordyceps species can be recognized in Taiwan and they are namely, C. takamontana, C. tuberculata, C.
fumosorosea, C. farinosus, C. javanica, C. tenuipes, C. ninchukispora, C. cicadae and C.
militaris (Chen 1978, Su and Wang 1986, Wang 1988, Tzean et al. 1997). After 1997, only two
new records Cordyceps spp. (Cordyceps cateniannulata and Cordyceps amoene-rosea) were available in Bioresource Collection and Research Centre (BCRC).
However, the rich diversity of organisms was threatened due to global warming. For example, the rising temperature resulted in the changes of population and geographical distribution of organisms such as insect and fungi and either decrease or increase their diversity.
(Steidinger et al. 2020, Wallisdevries and Van Swaay 2006). The increasing temperature allows the expansion of highly competitive organism which may result in the extinction of other organisms before they noticed (Selbmann et al. 2012).
During the literature review, we realized that Cordyceps diversity in Taiwan gains very few attentions. Studies related to Cordyceps in Taiwan are limited and stagnated after 1997.
the application of molecular data and One fungus=One name. We think it’s necessary to review the diversity of Cordyceps in Taiwan. As a result, the objective of this study was designed to discover the cryptic Cordyceps species and exploring the diversity of the genus Cordyceps in Taiwan.
Materials and methods
Fungal collecting, isolation and morphological examination
During 2017–2019, natural forests in Taiwan were examined for cordyceps-like fungal fruiting bodies colonizing various insects. The investigation was carried out in mountain areas ranging from 10 to 3952 meters above sea level in Taiwan (Figure 3). The specimens were placed in plastic containers to keep the moisture. Preliminary observation of fruiting bodies was conducted under the dissecting microscope (Hamlet® SEM-HB and Leica® S9D). Hand sections of fruiting body were made by Gillette blades and suspended in the water. Single spore isolation technique was used to obtain the pure culture from both sexual and asexual structures of the samples, following the method of Ariyawansa et al. (2016). In brief, spores were mounted in a drop of sterile distilled water on a flame-sterilized cavity slide, tweaked and mixed to uniform spore suspension. Spore suspension was then diluted to proper concentration, spread on 2%
water agar (WA, BioShop®) and incubated at 25°C in the dark for 12 hours or until spore germination observed. Germinated spores were separately transferred to Malt Extract Agar (MEA; HIMEDIA®) by using sterile syringe needle and cultured at 25°C for 7–14 days. For
minute, rinsed with autoclaved ddH2O and wiped with autoclaved towel paper. After the surface sterilization, the small amount of fruiting body was pinched by flame-sterilized forceps under the compound microscope (Olympus® BX51) and placed on 2% water agar (WA, BioShop®) at 25°C for 7–14 days to allow the growth of fungi. The growing tips of fungal mycelium was transferred to Malt Extract Agar (MEA; HIMEDIA®) to obtain the pure culture (Yadav et al.
2014).
To describe the morphological features of collected fungi, samples were first observed and captured using Canon PowerShot SX730 HS camera. Hand section of perithecia were mounted in distilled water and photoed under compound microscope (Olympus® BX51) through a differential interference contrast (DIC) illumination, equipped with a microscope digital camera (Olympus® DP27). To observe the asexual morphologies, pure culture of the fungus was grown on 2% PDA at 25°C for 7-21 days. Sporulation structures were located under dissecting microscope (Hamlet® SEM-HB) and placed on the slide to measures the sizes of conidiophores, phialides and conidia. For each structure, 30 measurements were taken and recorded by using cellSense Standard software (Olympus®).
Voucher specimens of taxonomic novelties and new records were deposited at the herbarium of Department of Plant Pathology and Microbiology, National Taiwan University (NTUH) and National Museum of Natural Science. Ex-type living cultures were deposited in Department of Plant Pathology and Microbiology, National Taiwan University Culture
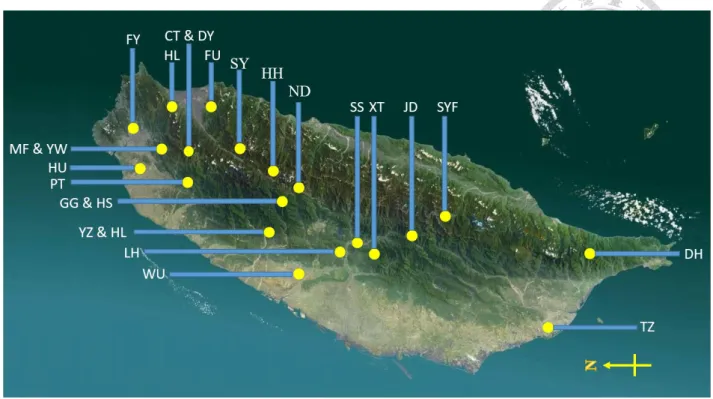
Figure 3. Map of Taiwan showing the areas surveyed in this study.
(MF & YW: Manyueyuan Forest Recreation & Yunsen Waterfall, New Taipei City; HL:
Honludi, Yilan County; CT & DY: Chatianshan Nature Reserve, Taoyuan City & Mt. Dongyan, Nantou County; GG & HS: Gugang, Taichung City & Huisun Experimental Forest, Nantou County; YZ & HL: Mt. Yuanzui & Mt. Hengling Trail, Taichung City; FU: Fushan Botanical Garden, Yilan County; FY: Fuyang Eco Park, Taipei City; HU: Mt. Hutou, Taoyuan City; PT:
Mt. Ptlaman, Hsinchu County; SY: Sihyuan Yakou, Yilan County; WU: Wufong, Taichung City;
LH: Lianhuachi Research Center, Nantou County; HH: Mt. Hehuan, Nantou County; ND: Mt.
Nandongyan, Nantou County; SS: Mt. Shuishe, Nantou County; XT: Xitou Forest Recreation Area, Nantou County; JD: Mt. Jade, Chiayi County; SYF: Siangyang Forest Recreation Area, Taitung County; DH: Dahan Forest Road, Pingtung County; TZ: Mt. Tzai, Kaohsiung City) DNA extraction and PCR amplification
1.5ml tube and homogenized with the glass beads by using 2010 Geno homogenizer (Grinder®
SPEX SamplePrep). Genomic DNA was extracted by the homogenized mycelium via using EasyPure Genomic DNA Spin Kit (Bioman®) (BIOMAN SCIENTIFIC CO., LTD), following the manufacturer’s protocol. The PCR amplification was carried out in a 25μl reaction containing 12.5 μl of 2X Taq PCR Mix-RED (Bioman®), 9.5 μl of dH2O, 1 μl of each forward and reverse primer, and 1 μl of fungal DNA template.
Five genic regions (ITS, nrLSU, tef1-α, rpb1 and rpb2) were selected following recent studies of Kepler et al. (2017), Mongkolsamrit et al. (2018) and Tasanathai et al. (2016).
Corresponding primers pairs and PCR conditions are listed in Table 2. The PCR products were visualized on 1.5% agarose electrophoresis gels integrated with BioGreenTM Safe DNA Gel buffer (Bioman®). Purification and sequencing of PCR products were carried out at Genomics company, New Taipei, Taiwan. All the sequences obtained in this study were deposited to NCBI GenBank.
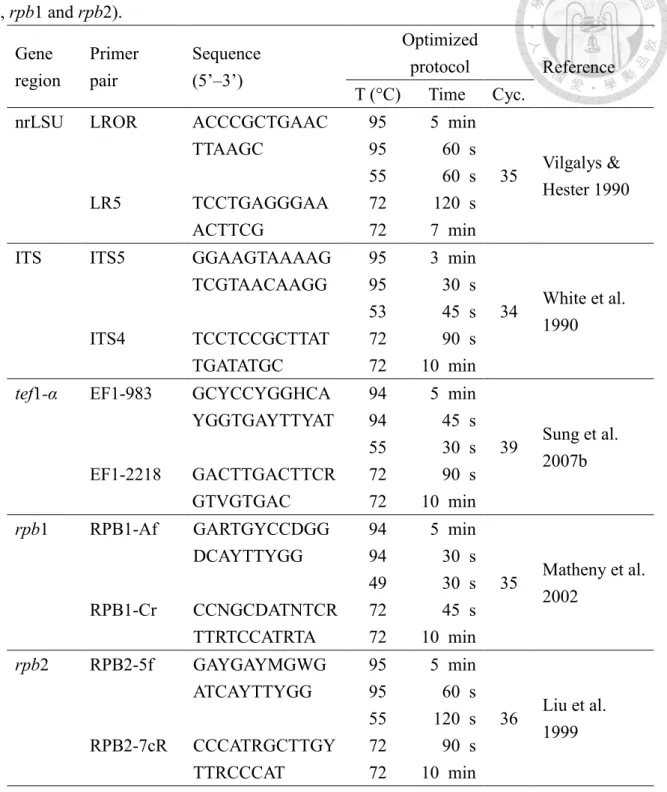
Table 2. Primer sets and PCR condition used to amplify the five gene loci (nrLSU, ITS, tef1- α, rpb1 and rpb2).
Gene region
Primer pair
Sequence (5’–3’)
Optimized
protocol Reference T (°C) Time Cyc.
nrLSU LROR
LR5
ACCCGCTGAAC TTAAGC
TCCTGAGGGAA ACTTCG
95 5 min
Vilgalys &
Hester 1990 95 60 s
35 55 60 s 72 120 s 72 7 min ITS ITS5
ITS4
GGAAGTAAAAG TCGTAACAAGG TCCTCCGCTTAT TGATATGC
95 3 min
White et al.
1990 95 30 s
34 53 45 s 72 90 s 72 10 min tef1-α EF1-983
EF1-2218
GCYCCYGGHCA YGGTGAYTTYAT GACTTGACTTCR GTVGTGAC
94 5 min
Sung et al.
2007b 94 45 s
39 55 30 s 72 90 s 72 10 min rpb1 RPB1-Af
RPB1-Cr
GARTGYCCDGG DCAYTTYGG CCNGCDATNTCR TTRTCCATRTA
94 5 min
Matheny et al.
2002 94 30 s
35 49 30 s 72 45 s 72 10 min rpb2 RPB2-5f
RPB2-7cR
GAYGAYMGWG ATCAYTTYGG CCCATRGCTTGY TTRCCCAT
95 5 min
Liu et al.
1999 95 60 s
36 55 120 s 72 90 s 72 10 min
Notes: nrLSU: Large subunit ribosomal DNA. ITS: Internal transcribed spacer regions. tef1-α:
translation elongation factor 1-alpha gene. rpb1: RNA polymerase II largest subunit gene. rpb2:
RNA polymerase II second largest subunit gene.
Phylogenetic analyses
Newly generated sequences were observed via BioEdit 7.2.5 software to check the quality of the sequences. All DNA sequences generated in this study were used to carry out BLAST search on NCBI website for preliminary identification. Sequences of related species and outgroup were selected based on previous studies (Sung et al. 2007, Chen et al. 2017, Kepler et al. 2017, Mongkolsamrit et al. 2018, Zhou et al. 2018) and downloaded from GenBank to construct the multiple sequence alignments (Table 3). Multiple sequence alignments were performed via MAFFT v. 7.409 (http://mafft.cbrc.jp/alignment/server/index.html) with the default settings.
Alignments were manually improved by BioEdit 7.2.5 software.
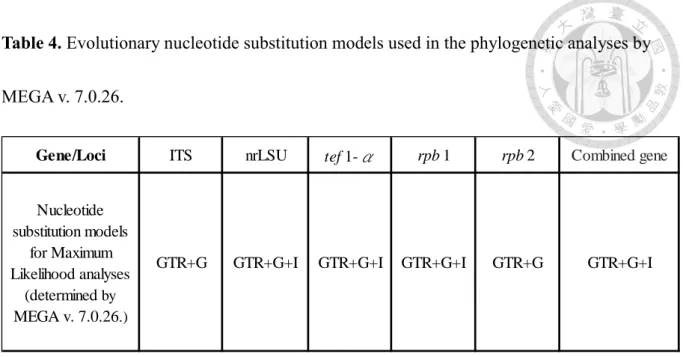
A maximum likelihood analyses (ML) was executed by raxmlGUI v. 1.5b (Silvestro and Michalak, 2012) with bootstrapping in 1000 replicates. The evolution models for each single- gene and concatenated dataset were determined by MEGA v. 7.0.26 (Table 4). To identify closely related taxa, single-loci phylogenies were inferred for LSU, ITS, tef1-α, rpb1 and rpb2 and finally a multi-gene combined analysis (LSU+ITS+tef1-α+rpb1+rpb2) was conducted. For each locus, the tree topologies were compared manually in order to confirm that the topologies of single-gene alignments were similar to each other, and that those of single- and concatenated gene alignments were comparable.
The resulting phylogenetic trees and data files were viewed in FigTree v. 1.4 (Rambaut and Drummond 2008: Retrieved from: http://tree.bio.ed.ac.uk/software/figtree/ on 2020/04/28).
support lower than 70% were considered unresolved. The concept of Genealogical Concordance Phylogenetic Species Recognition (GCPSR) (Taylor et al. 2000) was applied to delineate phylogenetic species boundaries. In brief, a clade was recognized as an independent taxon if it satisfied two criteria: Genealogical concordance and Genealogical non-discordance (Dettman et al. 2003). A taxon is genealogical concordance if it was present in the majority of phylogenetic trees (single-and multi-gene trees). A taxon is genealogical non-discordance if it is well supported by ML bootstrap value in at least one single-gene tree with no contradict in any other single-gene tree at same level of support.
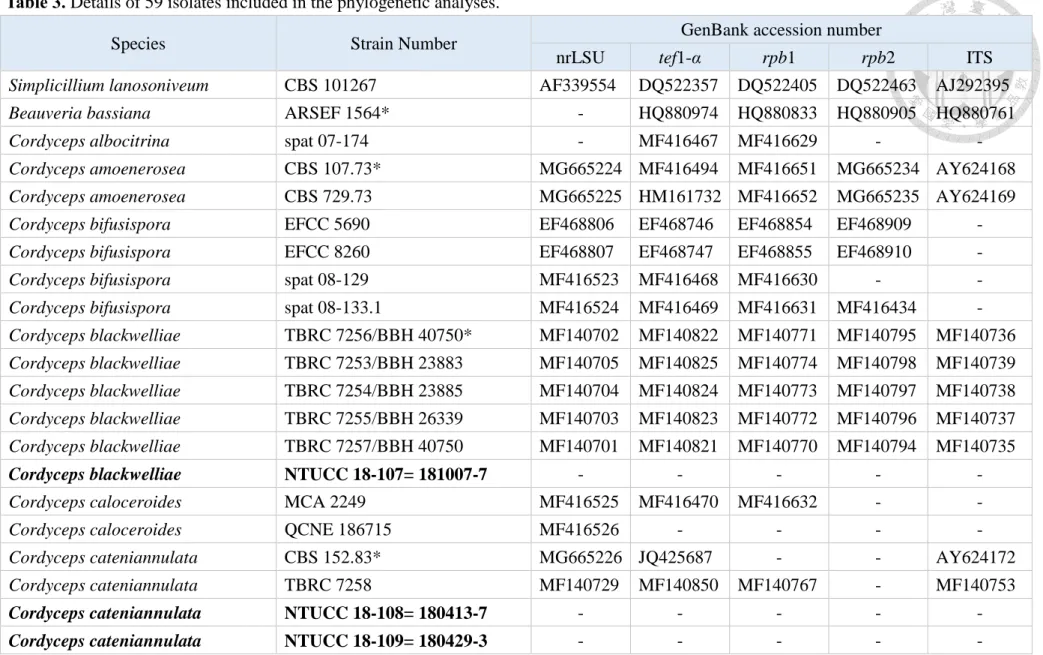
Table 3. Details of 59 isolates included in the phylogenetic analyses.
Species Strain Number GenBank accession number
nrLSU tef1-α rpb1 rpb2 ITS
Simplicillium lanosoniveum CBS 101267 AF339554 DQ522357 DQ522405 DQ522463 AJ292395
Beauveria bassiana ARSEF 1564* - HQ880974 HQ880833 HQ880905 HQ880761
Cordyceps albocitrina spat 07-174 - MF416467 MF416629 - -
Cordyceps amoenerosea CBS 107.73* MG665224 MF416494 MF416651 MG665234 AY624168
Cordyceps amoenerosea CBS 729.73 MG665225 HM161732 MF416652 MG665235 AY624169
Cordyceps bifusispora EFCC 5690 EF468806 EF468746 EF468854 EF468909 -
Cordyceps bifusispora EFCC 8260 EF468807 EF468747 EF468855 EF468910 -
Cordyceps bifusispora spat 08-129 MF416523 MF416468 MF416630 - -
Cordyceps bifusispora spat 08-133.1 MF416524 MF416469 MF416631 MF416434 -
Cordyceps blackwelliae TBRC 7256/BBH 40750* MF140702 MF140822 MF140771 MF140795 MF140736 Cordyceps blackwelliae TBRC 7253/BBH 23883 MF140705 MF140825 MF140774 MF140798 MF140739 Cordyceps blackwelliae TBRC 7254/BBH 23885 MF140704 MF140824 MF140773 MF140797 MF140738 Cordyceps blackwelliae TBRC 7255/BBH 26339 MF140703 MF140823 MF140772 MF140796 MF140737 Cordyceps blackwelliae TBRC 7257/BBH 40750 MF140701 MF140821 MF140770 MF140794 MF140735
Cordyceps blackwelliae NTUCC 18-107= 181007-7 - - - - -
Cordyceps caloceroides MCA 2249 MF416525 MF416470 MF416632 - -
Cordyceps caloceroides QCNE 186715 MF416526 - - - -
Cordyceps cateniannulata CBS 152.83* MG665226 JQ425687 - - AY624172
Cordyceps cateniannulata TBRC 7258 MF140729 MF140850 MF140767 - MF140753
Cordyceps cateniannulata NTUCC 18-108= 180413-7 - - - - -
Cordyceps cateniannulata NTUCC 18-109= 180429-3 - - - - -
Cordyceps cateniannulata NTUCC 18-110= 180618-1 - - - - - Cordyceps cateniannulata NTUCC 18-111= BCRC
FU31297 - - - - -
Cordyceps cateniobliqua CBS 153.83* - JQ425688 - MG665236 AY624173
Cordyceps chiangdaoensis TBRC 7274* MF140732 KT261403 - - KT261393
Cordyceps chiangdaoensis BCC75756 - KT261406 - - KT261396
Cordyceps chiangdaoensis BCC75733 - KT261407 - - KT261397
Cordyceps coleopterorum CBS 110.73* JF415988 JF416028 JN049903 JF416006 AY624177
Cordyceps coleopterorum CBS 102.73 - - - - AY624176
Cordyceps exasperata MCA 2155 MF416542 MF416486 MF416643 - -
Cordyceps exasperata MCA 2288 MF416538 MF416482 MF416639 - -
Cordyceps farinosa CBS 111113* FJ765253 GQ250022 MF416656 GU979973 AY624181
Cordyceps fumosorosea CBS 107.10* MG665227 HM161735 - MG665237 AY624184
Cordyceps fumosorosea CBS 337.52 MG665228 MG665233 MF416657 MF416451 EF411219
Cordyceps fumosorosea CBS 375.70 MG665229 HM161736 - MG665238 AY624183
Cordyceps fumosorosea CBS 244.31 MG665230 JQ425690 MF416660 MF416454 AY624182
Cordyceps fumosorosea CBS 107.1 MF416556 MF416502 MF416659 MF416453 -
Cordyceps fumosorosea NTUCC 18-112= 180310-5 - - - - -
Cordyceps fumosorosea NTUCC 18-113= 180413-9 - - - - -
Cordyceps ghanensis CBS 105.73* MH872340 - - - AY624185
Cordyceps jakajanicola BCC 79816* MN275696 MN338479 MN338484 MN338489 -
Cordyceps jakajanicola BCC 79817 MN275697 MN338480 MN338485 MN338490 -
Cordyceps jakajanicola BCC 12807 MN275694 MN338477 - - -
Cordyceps jakajanicola BCC 12839 MN275695 MN338478 - - -
Cordyceps jakajanicola NTUCC 17-036= CB - - - - -
Cordyceps jakajanicola NTUCC 17-037= CJ - - - - -
Cordyceps jakajanicola NTUCC 18-114= 180310-2 - - - - -
Cordyceps javanica CBS 134.22* MF416558 MF416504 MF416661 MF416455 NR_111172
Cordyceps javanica TBRC 7259/BBH 40411 MF140711 MF140831 MF140780 MF140804 MF140745 Cordyceps javanica TBRC 7260/BBH 40412 MF140710 MF140830 MF140779 MF140803 MF140744 Cordyceps javanica TBRC 7261/BBH 42149 MF140709 MF140829 MF140778 MF140802 MF140743
Cordyceps javanica TBRC 7262/BBH 41986 MF140712 MF140832 - MF140733 MF140746
Cordyceps javanica NTUCC 18-115= 180820-2 - - - - -
Cordyceps javanica NTUCC 18-116= BCRC
FU30878 - - - - -
Cordyceps kintrischica ARSEF 7218* - GU734751 - - EU553278
Cordyceps kintrischica ARSEF 8058 - GU734750 - - GU734764
Cordyceps kuiburiensis BCC 90322* MK968816 MK988032 MK988030 - MN099707
Cordyceps kuiburiensis BBC 90323 MK968817 MK988033 MK988031 - MN099708
Cordyceps kyusyuensis EFCC 5886 EF468813 EF468754 EF468863 EF468917 -
Cordyceps lepidopterorum TBRC 7263/BBH 40735* MF140699 MF140819 MF140768 MF140792 MF140765 Cordyceps lepidopterorum TBRC 7264/BBH 40737 MF140700 MF140820 MF140769 MF140793 MF140766
Cordyceps lepidopterorum NTUCC 18-117= 180729-2 - - - - -
Cordyceps lepidopterorum NTUCC 18-118= 180819 - - - - -
Cordyceps lepidopterorum NTUCC 18-119= 180830 - - - - -
Cordyceps militaris OSC 93623 AY184966 DQ522332 DQ522377 AY545732 JN049825
Cordyceps militaris NBRC30377 JN941385 AB968605 JN992490 AB968566 JN943300
Cordyceps militaris NBRC100741 JN941386 AB968606 JN992489 AB968567 JN943437
Cordyceps militaris NTUCC 17-039= CH - - - - -
Cordyceps militaris NTUCC 18-120= 180505-2 - - - - -
Cordyceps militaris NTUCC 18-121= BCRC 34266 - - - - -
Cordyceps militaris NTUCC 18-122= BCRC
FU30317 - - - - -
Cordyceps militaris NTUCC 18-123= C. mili A - - - - -
Cordyceps militaris NTUCC 18-124= C. mili B - - - - -
Cordyceps militaris NTUCC 18-125= C. mili Shen - - - - -
Cordyceps morakotii BCC 68398* MF140730 KT261398 - - KT261388
Cordyceps morakotii BCC 55830 MF140731 KT261400 - - KT261390
Cordyceps morakotii BCC 55820 - KT261399 - - KT261389
Cordyceps morakotii BCC 55831 - KT261401 - - KT261391
Cordyceps morakotii BCC 68403 - KT261402 - - KT261392
Cordyceps ninchukispora EFCC 5197 EF468820 EF468760 EF468868 - -
Cordyceps ninchukispora EFCC 5693 EF468821 EF468762 EF468869 - -
Cordyceps ninchukispora EGS 38.165 EF468846 EF468795 EF468900 - -
Cordyceps ninchukispora NHJ 10627 EF468822 EF468763 EF468870 - -
Cordyceps ninchukispora NHJ 10684 EF468823 EF468761 EF468871 - -
Cordyceps ninchukispora EGS 38.166 EF468847 EF468794 EF468901 - -
Cordyceps ninchukispora NTUCC 17-040= CP - - - - -
Cordyceps ninchukispora NTUCC 18-126= 180910-1 - - - - -
Cordyceps ninchukispora NTUCC 18-127= 180930-1 - - - - -
Cordyceps ninchukispora NTUCC 18-128= 180930-2 - - - - -
Cordyceps ninchukispora NTUCC 18-129= 180930-3 - - - - -
Cordyceps ningxiaensis HMJAU 25074* KF309671 - - - KF309668
Cordyceps ningxiaensis HMJAU 25075 KF309672 - - - KF309669
Cordyceps ningxiaensis HMJAU 25076 KF309673 - - - KF309670
Cordyceps oncoperae AFSEF 4358 AF339532 EF468785 EF468891 EF468936 -
Cordyceps polyarthra MCA 996 MF416543 MF416487 MF416644 - -
Cordyceps polyarthra MCA 1009 MF416544 MF416488 MF416645 - -
Cordyceps pruinosa ARSEF 5413 AY184968 DQ522351 DQ522397 DQ522451 JN049826
Cordyceps qingchengensis MFLU 17-1022* MK761211 MK770630 - - -
Cordyceps rosea spat 09-053 MF416536 MF416480 MF416637 MF416442 -
Cordyceps rosea NTUCC 17-041= CE - - - - -
Cordyceps rosea NTUCC 17-041-1= CE-1 - - - - -
Cordyceps roseostromata ARSEF 4871 AF339523 - - - -
Cordyceps roseostromata ARSEF 4870 - - - - EF368022
Cordyceps takaomontana BCC 12688 MF416545 MF416489 MF416646 - EU807996
Cordyceps takaomontana BCC 28612 FJ765252 FJ765268 - - FJ765285
Cordyceps takaomontana MCA 1806 MF416541 MF416485 MF416642 - -
Cordyceps tenuipes ARSEF 5135* JF415980 JF416020 JN049896 JF416000 AY624196
Cordyceps tenuipes OSC 111007 DQ518773 DQ522349 DQ522395 DQ522449 -
Cordyceps tenuipes TBRC 7265 MF140707 MF140827 MF140776 MF140800 MF140741
Cordyceps tenuipes TBRC 7266 MF140708 MF140828 MF140777 MF140801 MF140742
Cordyceps tenuipes TBRC 7267 MF140706 MF140826 MF140775 MF140799 MF140740
Cordyceps tenuipes NTUCC 18-130= 180226-1 - - - - -
Cordyceps tenuipes NTUCC 18-131= 180310-4 - - - - -
Cordyceps tenuipes NTUCC 18-132= 180331-1 - - - - -