體表與內臟器官間神經聯繫徑路的探討
75
0
0
全文
(2)
(3) 中文摘要 神經系統依據支配的位置不同可粗分為本體與臟器兩大系統,又依功能上的不同可分 為感覺與運動兩種系統,因此產生了體感覺(somatic sensory)、體運動(somatic motor)、 內臟感覺(visceral sensory)、內臟運動(visceral motor)四種系統。在之前的文獻中已經 分別的定義出這四個系統的神經傳導路徑,體感覺可經由脊髓-丘腦徑(spinothalamic tract)或是後柱-內側蹄系徑(posterior column-medial lemniscal tract)傳入中樞神經系 統,體運動主要經由皮質脊髓徑(corticospinal tract)傳出,而本體神經系統的感覺與運 動之間還有著直接的聯系稱之為脊髓反射(spinal reflex)。內臟感覺則經由脊髓網狀組 織徑(spinoreticular tract)、脊髓-丘腦徑、後柱-內側蹄系徑、迷走及舌咽神經傳入中樞 神經系統,內臟運動則是由交感(sympathetic)及副交感(parasympathetic)系統所調控,內 臟感覺與運動之間的反射則是可由內臟感覺直接調控內臟的運動,也就是自律神經系 統(autonomic system)。然而目前的文獻對於本體與內臟系統之間的神經聯系卻沒有太 多的研究,西方醫學臨床上觀察到的轉移痛(referred pain)以及傳統中國醫學中的針灸 (acupuncture)都表明了本體與內臟之間應有神經上的聯系存在,但是這兩者間的交會 與切確的路徑都沒有詳細的了解。之前的研究利用神經性的血漿滲漏證實了子宮與腹 股溝之間應存在神經上的聯系,但卻沒有辦法確定兩者之間是怎麼樣的聯系。本論文 利用 fos 蛋白的表現以及親神經性病毒做為神經追蹤劑,分別標定腹股溝區及子宮之 間的神經支配,發現本體感覺與內臟運動之間在腦幹的孤立束核(nucleus of solitary tract)、迷走背運動神經核(dorsal motor nucleus of vagus)及下丘腦室旁核(paraventricular. I.
(4) hypothalamic nucleus)的同一顆神經細胞上有所交會,因此推導出兩條可能的體-內臟反 射路徑,一條是體-副交感路徑(somato-parasympathetic pathway),另一條是體-交感路徑 (somato-sympathetic pathway),這兩條反射路徑同樣是針灸可能的神經路徑。另外我們 發現本體感覺與內臟感覺交會在背側神經節(dorsal root ganglion)及孤立束核上,卻沒 有在脊髓背側角上,所以我們假設轉移痛可能是經由背側神經節直接傳入孤立束核再 向上傳遞到丘腦。本研究建立了三條本體與內臟神經系統的可能徑路,對神經系統有 進一步的了解。. 關鍵字:轉移痛;針灸;內臟體表反射. II.
(5) Abstract The nerve system based on different innervation can be divided into somatic and visceral system. The nerve system based different function also can be divided as sensory and motor system. Therefore, there are somatic sensory, somatic motor, visceral sensory and visceral motor systems. The major two pathways of somatic sensory system are spinothalamic tract and posterior column – medial lemniscal tract. The somatic motor innervation is from cortex to skeletal muscle via corticospinal tract. The somatic sensory and motor systems have direct neuronal connection called spinal reflex between each other. The visceral sensory system translates visceral afferent through spinothalamic tract, spinoreticular tract, vagus nerve and glossopharyngeal nerve. The visceral motor system regulates the function of visceral organs by activating sympathetic and parasympathetic systems. The autonomic system is the reflex from visceral sensory afferent directly activate visceral motor efferent. However, it is still not clear whether the somatic and visceral systems have neuronal connection. The previous study using neurogenic plasma extravasation method showed that uterine pain referred to the groin region through a neuronal mechanism (Wesselmann and Lai, 1997). But, the relay center of this pathway is still not clearly identified. Our study proposes to investigate the neuronal connection between specific somatic point and visceral organ. In the present study, bee venom was injected in the groin region to induce central Fos expression to map the sensory innervation. III.
(6) of groin region. Pseudorabies virus (PrV), a transneuronal tracer, was injected in the uterus to identify the innervation of the uterus. Immunohistochemistry staining revealed that the fos expression and PrV infected double labeled neurons in the nucleus of solitary tract (NTS), the dorsal motor nucleus of vagus (DMX), and the paraventricular hypothalamic nucleus (PVN). These results suggest a somato-parasympathetic neuronal connection (groin-spinal dorsal horn-NTS/DMX-uterus) and a somato-sympathetic neuronal connection (groin-spinal dorsal horn-NTS-PVN-uterus). These two neuronal connections could be the pre-requisites to the neuronal basis of the somatovisceral reflex and also the neuronal mechanism of acupuncture. On the other hand, Fos expression and PrV infected double labeled neurons were mainly found in the T11, T12, T13, L1, S1 dorsal root ganglia, the nucleus of solitary tract (NTS), the other nuclei of brainstem, but not in the spinal dorsal horn. These results suggest the peripheral processes of the dichotomizing DRG neurons receiving sensory afferents either from the uterus or the groin region and the central process projects to the NTS. This morphological evidence demonstrates that the neuronal substrate of referred pain is the dichotomizing DRG neurons (the first order sensory neurons) and the NTS (the second order sensory neurons). These connections between somatic and visceral systems provide more understanding about the interaction of the nerve system. Key words:referred pain; acupuncture; somatovisceral reflex. IV.
(7) 誌謝 漫長的博士生涯終於結束了,能夠走完這一段路要感謝許多的人,首先要感謝的 當然是我的指導教授簡基憲老師,除了研究要點的指導以外在許多地方也都十分的照 顧我,讓我能堅持下去把博士念完。同樣也要感謝我的口試委員所給予的張永賢老 師、黃勇三老師、蔡玉娟老師及許鍾瑜老師給予的意見及肯定。 在攻讀博士學位的過程中,解剖所上的許多老師也是我一定要感謝的人,所上的 黃步敏老師、郭余民老師、曾啟育老師、江美治老師、吳佳慶老師、許鍾瑜老師都給 了我很多的幫助及鼓勵。當然還有宏南學長、書秤學姐、楊萬生醫生、欣燕姐、麗鶯、 宇彥、純英、儷友、亮宇等眾多同學的支持,這些支持在我求學過程中幫助我堅持下 去沒有放棄。同樣還要感謝我大學時期的好朋友們,阿今、小西、阿良、阿龍、大頭、 建鋒,雖然他們不在同一個學校,但給予我最多關懷的人還是這些朋友。 最後要感謝我的父母,給我一個能夠專心攻讀博士學位的家庭環境,雖然花了不 少時間,我還是想跟父母說聲”謝謝,我拿到博士學位了!!”. V.
(8) Contents Chinese abstract ……………………………………………I English abstract ………………………………...……….III Contents ……………………………………………….VI List of tables……………………………………………VII List of figures……………………………………………VIII Abbreviations…………………………………………X Introduction……………………………………………1 Material and method………………………………14 Results ………………………………………………….18 Discussion …………………………………………….21 Figures and tables ……………………….................35 Reference ……………………………………………..53. VI.
(9) List of tables Table 1 Fos expression neurons in supraspinal area……….……41. Table 2 PrV infected neurons in supraspinal area…………………47. VII.
(10) List of figures Figure 1 Fos expression neurons in dorsal root ganglia ......................………………35. Figure 2 Fos expression neurons in the spinal cord…………………………………...37. Figure 3 Fos expression neurons in the supraspinal area……………………………………39. Figure 4 PrV infected neurons in dorsal root ganglia.......………………………………………43. Figure 5 PrV-infected neurons in the spinal cord...................................................................44. Figure 6 PrV-infected neurons in the supraspinal area................................................................45. Figure 7 Double labeled of Fos expression and PrV infected neurons in dorsal root ganglia.........48. Figure 8 Double labeled neurons of Fos expression and PrV infection in the supraspina area.........49. VIII.
(11) Figure 9 Schematic drawing the possible referred pain pathway..........................................51. Figure 10 Schematic drawing three neuronal pathway of somatovisceral reflex.......................52. IX.
(12) Abbreviations PrV. Pseudorabies virus. NTS. Nucleus of solitary tract. PVN. Paraventricular hypothalamic nucleus. DMX. Dorsal motor nucleus of vagus. PB. Parabrachial nucleus. LC. locus coeruleus. RPa. raphe pallidus nucleus. PVT. paraventricular thalamic nucleus. LH. lateral hypothalamic area. IRt. intermediate reticular nucleus. Amb. ambiguus nucleus. LRt. lateral reticular nucleus. A5. A5 noradrenaline cells group. Gi. gigantocellular reticular nucleus. MPA. medial preoptic area. CRF. corticitropin-releasing factor. IML. intermediolateral nucleus. SPN. sympathetic preganglionic neurons. X.
(13) 1 Introduction The nerve system can be divided into sensory system and motor system according the functional difference. Furthermore, the sensory and motor systems can be divided into somatosensory, viscerosensory, somatomotor and visceromotor systems.. 1-1 Somatosensory system The somatosensory system transmits and analyzes information from the skin, joints and skeletal muscles that allow one to prepare and respond to afferent from the external environment. Different somatosensory pathways transmit different somatic sensations. There are two major somatosensory pathways, dorsal column-medial lemniscal pathway and spinothalamic pathway. The dorsal column-medial lemniscal pathway can transmit discriminative touch, flutter-vibration and proprioceptive information. The primary sensory fibers of dorsal column-medial lemniscal pathway enter the spinal cord via the medial division of the posterior root. They ascend cranially and contribute to the formation of the gracile and cuneate fascicule. These fiber bundles are termed the dorsal column, owing to their position in the spinal cord. The anterograde transport of HRP injected in dorsal root ganglia showed that the labeled central fibers from DRGs could be traced in the dorsal column and directly projected to the gracile and cuneate nuclei (Arvidsson and Pfaller,. 1.
(14) 1990). Other tracer and autoradiographic studies demonstrated the somatotopic organization of the the gracile and cuneate nuclear projections to the ventral posterolateral thalamic nucleus (Cliffer and Giesler, 1989, Gray, et al., 1981, Groenewegen, et al., 1975). In 1978 Brinkman’s study, monkeys were trained to investigate the somatosensory pathway. After complete bilateral section of the cuneate fascicule at C1-C2 spinal segment, the monkeys were still able to perform movement tasks normally. However, disturbance of discrimination ability was revealed if the animal was requied to detect differences in texture with only a small cutaneous area in contact with the object (e.g. using only the tip of the index finger)(Brinkman, et al., 1978). This study showed that the cuneate fascicule play an import role in the discriminative sensation afferent. The dorsal column-medial lemniscal pathway was adopted at last century and based on the pathological alterations see in certain disease states associated with dorsal column lesions (Al-Chaer, et al., 1998, Dmitriev, 1957). Another major somatosensory pathway is the spinothalamic pathway. The spinothalamic pathway can transmit nondiscriminative touch, innocuous thermal and pain sensations. In 1981, the electrophysiological percutaneous recording from C fibers suggested that nociceptive input via C fibers provoked by thermal or chemical stimulation correlates well with pain sensation (Van Hees and Gybels, 1981). The Phaseolus vulgaris leucoagglutnin (PHA-L), anterograde neuronal tracer, injection in dorsal root ganglia. 2.
(15) suggested that the central branchs of C-afferent fibers terminate in laminae I, II and V of the spinal cord (Sugiura, et al., 1989). The primary afferent axons with calcitonin gene-related peptide (CGRP), used as a marker for nociceptive primary afferents, immunoreaction terminate in lamina I and V (Traub, et al., 1990). Those studies prove that the central target of nociceptive primary afferent fibers (A-δand C fiber) of DRGs (first-order neurons) is the neurons in laminae I, II and V of the spinal dorsal horn. In 1991, Craig injected PHA-L in the spuerficial spinal dorsal horn to study the location of the ascending axons of spinal lamina I and showed that the axons of these second-order neurons in lamina I cross the midline and ascend in the contralateral anterolateral system(Craig, 1991). Neuronal tracer and electrophysiological study showed that thalamic (third-order) neurons of this pathway are located in the ventral posterolateral thalamic nucleus (Carstens and Trevino, 1978, Craig, et al., 1989).. 1-2 Somatic motor system The somatic motor system is the nerve system controlling the voluntary movements. The somatic motor system can simple be divided to lower motor neurons and upper motor neurons. The lower motor neurons are spinal anterior horn motor neurons whose axons innervate skeletal muscles. The upper motor neurons are cells in the brainstem and cerebral cortex projecting to lower motor neurons in the spinal cord. The major somatic motor. 3.
(16) system is the corticospinal pathway. The earlier studies about corticospinal pathway were based on the lesion of spinal or brain areas (Cardona, 1950, Lassek, 1948, Williamson, 1893). The studies of neuronal tracers make the better understanding about the corticospinal pathway. The upper motor neurons of the corticospinal pathway are located in deep portions of layer V of the primary motor cortex. The corticospinal fibers pass through the corona rediata and converge to enter the posterior limb of the internal capsule somatotopically. As the corticospinal fibers pass caudally from the internal capsule, they coalesce to form the middle third of the crus cerebri in the midbrain. Horseradish peroxidase (HRP) injections in various protions of the pontine nuclei resulted in retrograde labeling of neurons in layer V of the ipsilateral cerebral cortex (Kawamura and Chiba, 1979). This result show that the corticospinal fibers from layer V of the primary motor cortex continue into the basilar pons where they make their way longitudinally between the masses of neurons forming the basilar pontine nuclei. The corticospinal fibers aggregate on the anterior surface of medulla where they course within the medullary pyramids. The anterograde tracer injection in the motor cortex showed that the corticospinal fiber project to the contralateral spinal anterior horn (Ralston and Ralston, 1985). At the medullospinal junction, most of the corticospinal fibers cross the midline as the motor pyramidal decussation. The corticospinal fibers terminate primarily in the neurons of intermediate zone and anterior horn (lower motor neurons).. 4.
(17) 1-3 Visceral sensory system Visceral sensory system conveys information from receptors in visceral organ. This input allows the body to make appropriate responses to changes in its internal environment. The viscerosensory fibers are contained in the sympathetic and parasympathetic nerves as sympathetic afferent and parasympathetic afferent. Sympathetic afferent fibers conveying information from viscera travel via the cardiac and splanchnic nerves (Cervero, et al., 1984, Kuo, et al., 1981, Kuo, et al., 1984). The sympathetic primary sensory fibers join the cardiac and splanchnic nerves, enter the sympathetic trunk and pass through a white ramus to join the spinal nerve(Sato and Schmidt, 1971). The cell bodies of viscerosensory fibers are located in dorsal root ganglia and the central processes of these fibers terminate in laminae I and V. The neurons which receive these visceral inputs project mainly to the contralateral side as part of the anterolateral system. These fibers terminate in the ventral posterolateral nucleus of thalamus, which, in turn, projects to the inferolateral part of the postcentral gyrus and to the insular cortex. Sacral parasympathetic afferent fibers pass through the pelvic nerve, enter the spinal nerve and terminate in laminae I and V as sympathetic afferent fibers. These viscersensory inputs from neurons in laminae I and V project to the ventral posterolateral nucleus of thalamus via anterolateral system (Chernigovskii and Musiashchikova, 1969, Otake, et al., 1994). Visceral afferent fibers from chemoreceptors of the carotid body, baroreceptors of the carotid sinus wall and. 5.
(18) viscera in thoracic and abdominal cavities transmit a wide variety of physiologic information via the glossopharyngeal and vagus nerves(Calaresu and Henry, 1970). Cell bodies of these viscerosensory neurons are found within the inferior ganglia of the glossopharyngeal and vagus nerves. The central processes of these viscerosensory neurons enter the medulla, form the solitary tract and synapse with neurons of the adjacent solitary nucleus(Otake, et al., 1994). The solitary nucleus project to and influence a variety of neurons in the brainstem and hypothalamus.. 1-4 Visceral motor system The visceral motor system regulates cardiovascular, respiratory, digestive, urinary and reproductive organs. The visceral motor system provides neural control of smooth muscle, cardiac muscle, glandular secretory cells or combinations of these tissues. The visceral motor system has major sympathetic and parasympathetic subdivisions. The sympathetic and parasympathetic divisions have overlapping and generally antagonistic effects on those viscera. As the somatic motor system, the visceral motor system innervates viscera via motor neurons. However, the peripheral visceral motor pathway consists of two neurons, preganglionic neurons which have cell bodies in either the brainstem or the spinal intermediolateral nucleus, and postganglionic neurons which have cell bodies in the paravertebral, prevertebral or intramural ganglia(Niel, et al., 1980). The sympathetic. 6.
(19) preganglionic neurons are located in the intermediolateral nucleus of the spinal lateral horn and the axons exit the spinal cord in the ventral root to the sympathetic trunk via the white communicating ramus. These preganglionic fibers may synapse with sympathetic postganglionic neurons (sympathetic chain ganglioa) in sympathetic trunk or in prevertebral ganglia through splanchic nerve(Niel, et al., 1980). The postganglionic axons innervate blood vessels, smooth muscle and viscera. The cell bodies of parasympathetic preganglionic neurons are located in either sacral spinal lateral horn or the nuclei of cranial nerves III, VII, IX and X in brainstem. The cell bodies of parasympathetic neurons are scattered within nerve plexuses of the target organ(DeGroat and Saum, 1976).. 1-5 Patellar reflex and autonomic system The sensory and motor systems do not completely separate from each other. The connections between sensory and motor systems make many reflexes. Reflexes are involuntary responses to a particular bit of sensory input. Reflexes require afferent fibers, interneurons and motor neurons. Between somatic sensory and motor system, there are many reflexes including muscle stretch reflex and flexor reflex. In muscle stretch reflex, a tap on the patellar tendon stretches the primary somatic sensory endings in muscle spindles located in the quadriceps femoris muscle, sending an impulse toward the posterior root ganglion. The central processes of these afferent axons synapse on and excite motor. 7.
(20) neurons that innervate the quadriceps femoris muscle. The result is a sudden contraction of these muscles and an extension of the leg at the knee(McCrea, 1986). Flexor reflex is initiated by cutaneous input, is frequently a response to nociceptive stimuli and represents an attempt to protect a body part by extricating it from the source of injury. Somatic primary sensory fibers conveying nociceptive input enter the spinal cord and form excitatory synaptic contacts with not only ascending tract cell but also interneurons. The excitatory interneurons synapse on flexor motor neurons, resulting in activation of the ipsilateral flexor muscles of the thigh, leg and foot, and withdrawal of the extremity(Danilov and Vein, 1996, McCrea, 1986). These reflexes are the connections between somatic sensory and motor system. In fact, the connections between visceral sensory and motor system are more common. The regulations of autonomic system in viscera are almost all involuntary and involve both visceral sensory and motor system. The autonomic reflexes can be divided to spinal autonomic reflex and supraspinal autonomic reflex. In spinal autonomic reflex, the DRG neurons receiving viscerosensory afferents activate the preganglionic neurons and the preganglionic axons project to the postganglionic neurons innervating the visceral functions (Kuo, et al., 1984, Sato and Schmidt, 1971). The supraspinal autonomic reflex transmit the viscerosensory input to the brainstem nuclei or hypothalamus which active the preganglionic neurons and regulate visceral activation (Calaresu and Henry, 1970). In the. 8.
(21) baroreceptor reflex, the primary visceral sensory fibers from vagus nerve project to the solitary nucleus in the medulla. The second order neurons in the solitary nucleus project to a variety neurons in the brainstem and hypothalamus. One of these targets is the dorsal motor nucleus of vagus which receives input from the solitary nucleus and project, via the vagus nerve, to parasympathetic ganglia of the heart. Activation of this pathway causes a decrease in hear rate and blood pressure, a vasodepressor response. Conversely, the neurons locating in rostral parts of the anterolateral medulla receive input from the solitary nucleus and project to the spinal cord where they influence the activity of preganglionic sympathetic motor neurons in the lateral horn. In doing so, the reflex causes an increase in blood pressure and serves a vasopressor function (Coote and Macleod, 1974, Spyer, 1975, Trzebski, et al., 1975).. 1-6 Acupuncture and referred pain The reflex pathways between somatic sensory and motor system have been known. The regulations of autonomic system between visceral sensory and motor system are also been studied for a long time. However, the connections between somatic and visceral system are still poorly understood. The somatovisceral reflex was mentioned by Sato in 1995 and suggested that somatic stimulation could evoke sympathetic reflex response and, thereby, modulate functioning of. 9.
(22) visceral organ (Sato, 1995). This phenomenon is in someway alike acupuncture that stimulate specific somatic points to relieve pain and treat many different diseases (World Health Organization. Regional Office for the Western Pacific., 1993). Many studies have shown that acupuncture can significantly modulate visceral function by stimulating specific acupoints (Ballegaard, et al., 1986, Bao, et al., 1982, Chao, et al., 1999, Ho, et al., 2009, Li, et al., 1998, Richter, et al., 1991). Previous research suggested that the activation of the somatosensry pathway played an important role in the physiological effects of acupuncture (Kim, et al., 2012). Li’s research showed that electroacupuncture-like stimulation diminishes regional myocardial ischemia triggered by sympathetically excitation (Li, et al., 1998). Other studies have shown that electroacupuncture-like stimulation can activate a sympathetic inhibitory system in the brain to regulate cardiovascular responses (Chao, et al., 1999, Peng, 2002, Sun and Yao, 1985). Both the somatovisceral reflex and acupuncture stimulation suggest the neuronal connection between somatic acupoint and its corresponding organ. However, the neuronal connection of the somatovisceral reflex or acupuncture is still not clear. Beside the somatovisceral reflex, the somatic and visceral sensory system may also have connection between each other. Referred pain indicates the deep pain, particularly from visceral organ, referred or transferred to a specific superficial area. The referred pain is one characteristic feature of inflamed visceral organ and also a clinical diagnostic tool. 10.
(23) (Cervero and Laird, 1999). The mechanism of referred pain has been studied for a long period of time and is sum up in several theories, including the convergence-facilitation theory, the convergence-projection theory, as well as, the dichotomizing afferent fibers. The convergence-projection theory mentioned by Ruch is one possible mechanism of referred pain which suggested afferent inputs from both somatic and visceral structures projected to the same second order spinal neuron (Cervero, 1995, Gebhart and Ness, 1991, Ness and Gebhart, 1990). Previous horseradish peroxidase (HRP) and electrophysiological studies suggested that visceral and somatic afferent fibers converge onto the lamina I and V in spinal dorsal horn (Cervero and Connell, 1984, Hobbs, et al., 1992, Sessle, et al., 1986). Another possible mechanism is the dichotomizing neurons, single neuron with dichotomizing fibers innervating two different tissues, in the dorsal root ganglion (DRG) mentioned by Sinclair in 1948 (Sinclair, et al., 1948). The dichotomizing DRG neurons were proved by injecting different neuronal tracers respectively to visceral organs, e.g. uterus and colon or bladder and colon (Chaban, et al., 2007, Christianson, et al., 2007). Other researches showed that the dichotomizing DRG neurons also receive both sensory inputs form both somatic and visceral afferents (Chen, et al., 2005, Laurberg and Sorensen, 1985, Pierau, et al., 1984, Sameda, et al., 2003, Wakai, et al., 2010). However, the morphological evidence is still not clear up to this date.. 11.
(24) 1-7 The neuronal connection between somatic area and visceral organ Pervious report demonstrated that gynecological pain induced by dysmenorrhea, ascending genital infection, or cystic or hemorrhagic ovarian pathology usually refer pain to the low back, thighs, and abdominal wall (Giamberardino, et al., 1997). Referred pain in the low back and abdominal wall were also reported by women in labor (Melzack and Belanger, 1989). These reports suggested that the groin region can account to the pain of uterine inflammation or diseases. According to traditional Chinese medicine, some acupoints, called A-shi points, do not have fixed specific locations and are usually pain-associated points (Liu, 2011, Xu and Li, 2005, Yang, et al., 2012). Therefore, the groin region could be the A-shi point related with the uterus. The groin region is not only the referred area of uterine pain but also the A-shi point related with the uterus. Although one is somatic region and another is visceral organ, the neuronal connection between the groin region and uterus should exist. In 1997, Wesselmann and Lai found that uterine inflammation in rats pretreated with Evans Blue Dye resulted in neurogenic plasma extravasation of dye in the skin over the abdomen, lower back, thighs and groin, after antidromic stimulation of peripheral nerves (Wesselmann and Lai, 1997). This result suggested the possibility of neural connection between the uterus and groin areas. Although these findings confirm the existence of a. 12.
(25) neural connection between the uterus and groin region, the exact location of this central neuronal connection remains unknown. The Fos protein is an immediate-early gene transcription factor induced by short-term signals and alter target gene expression causing long-term change in cellular phenotype (Curran and Morgan, 1995). It has been used to map the activated neural cells after different types of stimulation, and shows correlated anatomical neural pathways (de Medeiros, et al., 2003, Ghanima, et al., 2002, Luo, et al., 1998). Pseudorabies virus (PrV) is a swine neurotropic herpes virus that has been used for transneuronal tracing in many studies (Chien, et al., 1998, Lee and Erskine, 2000, Papka, et al., 1998, Rothermel, et al., 2007, Weiss and Chowdhury, 1998). The Pingtung (PT) strain of PrV has been demonstrated to label sympathetic pre- and postganglionic neurons after injection in the specific auricular kidney point (Chien, et al., 1998). The study showed that the PT strain of PrV was a useful transneuronal tracer in somatovisceral research. To establish the neural connection between the groin region and uterus, bee venom was injected in the groin region to induce c-Fos expression neurons innervating the groin region and PrV was injected in the uterus to infect the neurons innervating the uterus. Furthermore, to evaluates central doubled Fos-expression and PrV-infected neurons in order to identify the neuronal connection between the somatic point (groin region) and its related visceral organ (the uterus).. 13.
(26) 2 Materials and Methods The study protocol was approved by Animal Care and Use Committee, and all experiments were conducted in accordance with the animal care guidelines of the National Institutes of Health and the International Association for the Study of Pain.. 2-1 Animals Sprague-Dawley adult virgin female rats (250–350 g) were used. Animals were housed on a 12h–12-h light–dark cycle, and all animals had free access to standard food and water.. 2-2 Bee venom injection in the left groin region The rats were anesthetized with ketamine (95mg/kg) intraperitoneally. 50μl of 1% bee venom(Sigma) were dissolved in normal saline and administrated subcutaneously into the midpoint between genital pore and apex of the left groin region (n=6) according to Wesselmann’s research (Wesselmann and Lai, 1997). Saline were injected as the control. After 90 minutes, the rats were sacrificed and perfused with 250 ml of saline intracardially, followed by 1000 ml of 4% paraformaldehyde in 0.1 M phosphate buffer solution (PBS). T10-S1 segments of spinal cord, brainstem and brain were removed.. 2-3 Pseudorabies virus injection in left uterine horn 14.
(27) The rats were anesthetized with ketamine (1ml/kg) intraperitoneally. A laparotomy was performed and 40 μl of Pingtung strain pseudorabies virus (Chien, et al., 1998) was injected into the left uterus horn (n=9). After the injections, the abdominal wall was sutured, the skin closed. The animals were sacrificed at 6 to 8 days after PrV injection (in the same way as described above). The spinal cord, dorsal root ganglion of T10-S2 segment, brainstem and brain were removed.. 2-4 Bee venom injection after PrV injection The rats were anesthetized and PrV was injected into the left uterus horn (in the same way as described above ) (n=9). After the injections, the abdominal wall was sutured, the skin closed and the animals allowed to survive for 6 to 8 days. Before the rats were sacrificed, 50μl of 1% bee venom were administrated subcutaneously into midpoint of left groin region and saline were injected as the control. After 90 minutes, the rats were sacrificed and perfused with 250 ml of saline intracardially, followed by 1000 ml of 4% paraformaldehyde in 0.1 M phosphate buffer solution (PBS). The spinal cord, dorsal root ganglion of T10-S1 segment, brainstem and brain were removed.. 2-5 Immunohistochemistry Tissues of groin region - bee venom injection group and uterine horn – pseudorabies. 15.
(28) virus injection group were postfixed up to 4 hr in paraformaldehyde PBS and then cryoprotected in 10, 20 and 30% sucrose in PB solution. Serial 30μm thick transverse section of all dorsal root ganglia, spinal cord, brainstem and brain were cut with a cryomicrotome. All sections from the ganglia and every five sections from other samples were collected in 0.01 M phosphate buffer saline (PBS). Floating sections were washed 30 min (10 min, 3 times) and incubated with blocking solution (5% normal goat serum, 0.05% Triton X-100, 3% BSA in 0.1M PB) 1 hr. The sections were washed and incubated with the primary antibody (IgG of rabbit anti-FOS in 1:2000 or IgG of swine anti-PrV in 1: 1000) in blocking solution for 72 hr at 4℃. After incubation, the sections were rinsed and incubated 1hr at 25℃ with secondary antibody (biotin-conjugated IgG of goat anti-rabbit in 1:500 or goat anti-swine in 1:200) in blocking solution. The sections were washed three times for 30 min and incubated using ABC kit (Vector) for 1hr. After rinsing, the sections were developed with GOD method followed by mounting on gelatin- coated slides and overslipped with mounting medium.. 2-6 Immunofluorescence Tissues of groin-uterus injection group were postfixed and sectioned in the same way described above. Floating sections were washed 30 min (10 min, 3 times) and incubated with blocking solution (5% normal goat serum, 0.05% Triton X-100, 3% BSA in 0.1M PB). 16.
(29) 1 h. The sections were washed and incubated with two kinds of primary antibody (IgG of rabbit anti-FOS in 1:2000 and IgG of swine anti-PrV in 1:1000) in blocking solution for 72 hr at 4℃. After incubation, the sections were rinsed and incubated 1hr at 25℃ with two secondary antibody(FITC-conjugated IgG of goat anti-swine in 1:200 and TRITC-conjugated IgG of goat anti-rabbit in 1:500) in blocking solution. The sections were washed for 30 min and mounted on gelatin- coated slides followed by coverslipping with mounting medium.. 2-7 Data and statistical analysis Fos and PrV immunoreactivity neurons developed with GOD method in dorsal root ganglia, spinal cord and brain were counted with bright field microscope. Fos and PrV double labeled neurons were observed with fluorescent microscope. Anatomical identification of brain structures was essentially based on the Rat Brain atlas of Paxinos and Watson (1998). All data were analysis by T-test.. 17.
(30) 3 Result 3-1 Fos expression neurons after bee venom stimulation in the groin region Injecting bee venom in the left somatic groin region induces central Fos expression and the contralateral side as the control. Fos expression neurons were distributed in the ipsilateral dorsal root ganglia of T10(8.7±3.4), T11(5.0±1.5), T12(6.3±1.5), T13(7.9±3.4), L1(13.1±4.4), L2(5.4±1.9), L3(0.9±0.5), L4(0.±0.1), L6(5.1±2.1) and S1(10.3±4.6) levels (fig. 1A, B). In the spinal cord, Fos protein is predominantly (70%)apparent in ipsilateral T13 (14.5±1.1), L1 (23.5±1.5) and L2 (26.6±2.5) spinal dorsal horn (Fig. 2A, B, C). Most of the c-Fos expression neurons are resided in lamina I (12.6±1.2) and II (10.9±1.4) of the dorsal horn (Fig. 2 D). In the supraspinal area, c-Fos expression neurons appeared in numerous nuclei of the thalamus, hypothalamus, pons, and medulla. The c-Fos expression nuclei include the nucleus of solitary tract (NTS) (Fig. 3A), parabrachial nucleus (PB), locus coeruleus (LC) (Fig. 3B), raphe pallidus nucleus (RPa) (Fig. 3C), paraventricular thalamic nucleus (PVT) (Fig. 3F), lateral hypothalamic area (LH) (Fig. 3E), and paraventricular hypothalamic nucleus (PVN) (Fig. 3D). Table 1 listed the fos expression neurons in suprasipnal areas of saline and bee venom groups. The NTS of bee venom group expressed significantly difference between fos expression neurons and saline group.. 18.
(31) 3-2 The appearance of PrV infection neurons after virus injection in the uterus After PrV was injected in the left uterine horn, PrV infected neurons were distributed in the dorsal root ganglia of T10(11.6±3.2), T11(6.7±1.4), T12(6.4±1.4), T13(8.8±2.9), L1(14.4±3.4), L2(3.7±1.4), L3(0.5±0.3), L4(0.1±0.1), L5(0.1±0.1), L6(5.8±2.6) and S1(16.3±6.1) spinal segment, mainly in T10-L1 (64.4%)and L6-S1(29.7%) levels (fig. 4A, B). PrV-infected neurons appeared in the central nuclei (Fig. 4, 5) 6-8 days after PrV injection in the left uterus horn. In the spinal cord, the most PrV-infected neurons were spotted in lamina IV and V of T11-L1 and L5-S2 spinal segments (Fig. 5 A, B, C), rarely in the superficial lamina (lamina I, II). In the supraspinal area, PrV-infected neurons were found in the hypothalamus, pons, and medulla, including the NTS, dorsal motor nucleus of vagus (DMX) (Fig. 6 A), intermediate reticular nucleus (IRt), ambiguus nucleus (Amb), lateral reticular nucleus (LRt), A5 noradrenaline cell group (A5) (Fig. 6 B), raphe pallidus nucleus (RPa) (Fig. 6 C), gigantocellular reticular nucleus (Gi) (Fig. 6 D), medial preoptic area (MPA), and PVN (Fig. 6 E). All PrV infected neurons in supraspinal area were listed in Table 2.. 3-3 Fos expression and PrV infected double labeled neurons in the NTS, DMX and PVN 19.
(32) After the uterine PrV injection and c-fos expression of the groin region bee venom stimulation, Fos expression and PrV infected double neurons are observed in the dorsal root ganglia (fig. 7) and supraspinal area(fig. 8), but not in spinal level. The Fos expression and PrV infected double neurons can be observed in T10-L1 and S1dorsal root ganglia, mainly in T11(3.0±1.6%), T13(2.8±1.4%), L1(2.2±1.0%) and S1(1.8±1.0%) levels (fig. 7). In supraspinal area, double labeled neurons appeared in the hypothalamus, and specifically in the PVN (Fig. 8 C1). Some other double labeled neurons are apparent in the NTS and DMX (Fig. 8 C2, C3, C4). Comparing saline and bee venom injection in PrV infected neurons, the percentage of the double labeled neurons in PVN of bee venom injection group were significantly predominant (p<0.1) (Fig. 8 D).. 20.
(33) 4. Discussion. 4-1 Sensory innervation of the left groin region The Fos protein is an immediate-early gene transcription factor induced by short-term signals and alter target gene expression causing long-term change in cellular phenotype (Curran and Morgan, 1995). Neuron cells including first, second or third order sensory neurons receive sensory inputs, like pain, and express Fos protein in 1 to 24 hr after stimulations. Neurons activated by stimulations can be detected by immunohistochemistry method. Detecting the Fos expression can clear and localize which neurons or nuclei in nerve system receive the input. It has been used to map the activated neuron cells after various stimulations and the demonstrations of the functional anatomical neural pathways (de Medeiros, et al., 2003, Ghanima, et al., 2002, Luo, et al., 1998). The stimulating acupoints elicits a composite of unique sensations called dechi. Dechi sensations including pressure, soreness, heaviness and dull pain are essential for clinical efficacy (Hui, et al., 2007). Pain, as one of the dechi sensations, is a relatively strong and easily induced sensory modality in animal study. Panic stimulation can induce neurons expressing the Fos protein which could be used to investigate either somatic (Pinto, et al., 2006, Roh, et al., 2006, Tavares, et al., 1993) or visceral (Martinez, et al., 2006, Monnikes, et al., 2003, Tavares, et al., 1993) noxious afferent pathways. Takahashi (Takahashi and Nakajima, 1996) intravenous injected in the Evans blue dye and observed extravasation in. 21.
(34) the groin after electrical stimulation of the spinal nerves. This result proves that the sensory innervation of groin region comes from T13, L1, and L2 spinal nerves. Our data show that bee venom injection in the left groin region induces c-fos expression in the ipsilateral DRG, spinal dorsal horn of T13, L1 and L2 segments, specifically on superficial laminae (lamina I and II). This suggests that the spinal Fos expression neurons in our study were specifically activated by noxious stimulation of the groin region. The Fos expression neurons were also observed in many brainstem nuclei which were noxious respond related nuclei after bee venom stimulation. Fos expression neurons appeared in the NTS, gigantocellular reticular nucleus (Gi), raphe pallidus nucleus (RPa), and PVN after bee venom stimulated groin region. The NTS can be not only activated by somatic noxious stimulation but also triggered by vagal afferent activation as a physiological adaptation to pain (Ter Horst, et al., 2001). Previous studies showed that Gi can be activated by noxious stimulation related to the activation of the descending anti-nociceptive pathway (Burton, 1968, Goldman, et al., 1972, Zhuo and Gebhart, 1990, Zhuo and Gebhart, 1992). Electrically stimulating the raphe nucleus could induce analgesia, proving that the raphe nucleus plays a crucial role in pain inhibition response (Lee and Beitz, 1993, Llewelyn, et al., 1986, Oliveras, et al., 1975). The thermal stimualtion from hind feet inducing Fos expression in the PVN showed that the PVN can receive the noxious input (Pan, et al., 1999). The activation of PVN initiates the. 22.
(35) hypothalamus-pituitary-adrenal hormone cascade by releasing corticotropin-releasing factor (CRF) and arginine vasopressin from its parvocellular cells (Whitnall, 1988). Fos expression neurons in those nuclei suggest that injecting bee venom in the groin region activating not only pain transmission pathway but also the nuclei regulating physiological responses and inhibiting pain in the central nervous system.. 4-2 Hierarchical innervation of the uterus The PrV transneuronal tracing method is widely used to detect the hierarchically central innervation of urethra, clitoris, penis, urinary bladder, and uterus (Lee and Erskine, 2000, Marson, 1995, Marson, et al., 1993, Moore, et al., 1995, Nadelhaft, et al., 1992, Papka, et al., 1998, Park, et al., 1997, Vizzard, et al., 1995, Wiesel, et al., 2004). In our previous study, the Pingtung strain of PrV applied to the auricular kidney point transneuronally and specifically infected sympathetic pre- and postganglionic neurons (Chien, et al., 1998). In order to investigate the highest central control of uterus, the survival time was proportionally extended to 6-8 days in this study. All PrV infected nuclei in our study (Table 2) could be found in these nuclei reported by other strain of PrV transneuronal studies (Lee and Erskine, 2000, Papka, et al., 1998). This result confirms the Pingtung strain of PrV was a sustainable strain as a transneuronal tracer in hierarchical innervation studying.. 23.
(36) Lee and Papka discovered PrV-infected sympathetic and parasympathetic preganglionic neurons at T11-13 and L6-S1 spinal segments, after PrV injection in the uterine cervix (Lee and Erskine, 2000, Papka, et al., 1998). The results indicate the visceral efferent to the uterus are mainly from T11-T13 and L6-S1 segments. In our study, PrV-infected preganglionic neurons are mainly in the intermediolateral nucleus (IML) and sacral parasympathetic nucleus of T10-L2 and L6-S1 spinal segments (Fig. 5). Our result is accord with previous studies (Lee and Erskine, 2000, Papka, et al., 1998, Wiesel, et al., 2004). Supraspinal PrV-infected high hierarchical uterine neurons are located in the NTS, dorsal motor nucleus of vagus (DMX), A5 noradrenaline cell group (A5), raphe pallidus nucleus (RPa), gigantocellular reticular nucleus (Gi), and PVN (Fig. 4). Retrograde tracer studies have shown that the DMX innervates the cecum, uterus, and colon directly or indirectly (Altschuler, et al., 1991, Collins, et al., 1999, Ortega-Villalobos, et al., 1990, Vizzard, et al., 2000, Wiesel, et al., 2004). The neurons in the A5 cell group, RPa, and Gi innervating the uterus were also confirmed by previous PrV tracing reasearches (Lee and Erskine, 2000, Papka, et al., 1998, Wiesel, et al., 2004). Therefore, the PVN function is not only a uterus-related hormone regulation center (Patisaul, et al., 2002), but also a direct neuronal connect to the uterus (Lee and Erskine, 2000, Papka, et al., 1998). After PHA-L injection in the PVN, terminal varicosities appeared in the IML of the thoracic spinal cord (Hosoya, et al., 1991). Retrograde tracer studies also showed a neuronal connection. 24.
(37) between the PVN and the uterus (Lee and Erskine, 2000, Papka, et al., 1998, Wiesel, et al., 2004). In summary, all these results suggest that the PVN has either a direct or indirect neuronal connection and it regulates the uterus through both hormonal and neuronal innervation.. 4-3 Sensory innervation of the uterus PrV transneuronal tracing method is widely used to detect hierarchically central innervation of urethra, clitoris, penis, urinary bladder and uterus (Kim, et al., 1999, Lee and Erskine, 2000, Marson, 1995, Marson, et al., 1993, Moore, et al., 1995, Nadelhaft, et al., 1992, Papka, et al., 1998, Park, et al., 1997, Vizzard, et al., 1995, Wiesel, et al., 2004). However, PrV is not only a retrograde tracer but also infect DRG and spinal dorsal sensory neurons. In 1998, Weiss use PrV as anterograde tracer to investigate the renal afferent pathway and showed the PrV infected DRG and spinal dorsal horn (Weiss and Chowdhury, 1998). Anterograde neuronal tracer injection and electrophysiological studies showed that the sensory innervation to the uterine horns is from the dorsal root ganglia of T13, L1, L2 and L6, S1 levels (Berkley, et al., 1988, Nance, et al., 1988, Peters, et al., 1987). Lee and Papka proved PrV infected sensory uterine cervix neurons were observed at L6 and S1 spinal segments (Lee and Erskine, 2000, Papka, et al., 1998). In summary, those data are in accordance with our finding that the first order sensory neurons innervating uterus dwell. 25.
(38) mainly in T10-L2 and L6-S1 dorsal root ganglia, respectively (fig. 4). PrV infected neurons were also found in several supraspinal sensory nuclei (fig. 6). Nucleus of solitary tract (NTS) is the major brainstem structure that receives general and special visceral sensory inputs including visceral pain (Altschuler, et al., 1991, Appia, et al., 1986). NTS has also been proven having neuronal connection with uterus in either electrophysiologically or HRP studies (Hubscher and Berkley, 1994, Ortega-Villalobos, et al., 1990). Vagotomy or hypogastric nerve section after uterine cervical stimulation confirmed that the NTS receive the visceral afferents information from the uterus (Guevara-Guzman, et al., 2001). Our viral tract tracing method specifically labelled the central afferent innervation of the uterus.. 4-4 Somatic sensory and visceral sensory convergence 4-4-1 Double labeled neurons in first order sensory neuron “ dorsal root ganglion” In 1948, Sinclair hypothesized that the DRG neurons with dichotomizing fibers (i.e. a single sensory neuron innervating two different tissues) is the morphological basis of visceral referred pain (Sinclair, et al., 1948). In fact, the dichotomizing DRG neurons was also believed to be the mechanism of visceral organ cross-sensitization (Brumovsky and Gebhart, 2010). To prove the dichotomizing DRG neuron, different fluorescent dye were. 26.
(39) applied to each organs respectively and double tracers labeled DRG neurons were observed, i.e. colon and bladder, colon and uterus (Brumovsky and Gebhart, 2010, Chaban, et al., 2007, Christianson, et al., 2007, Keast and De Groat, 1992, Li, et al., 2008). Other tracers tracing studies also showed sensory innervations between skin and visceral organ convergences in the same DRG neurons (Pierau, et al., 1984), for example : shoulder skin and diaphragm (Laurberg and Sorensen, 1985), lumbar disc and groin skin (Sameda, et al., 2003), intervertebral disc and psoas muscle (Takahashi, et al., 2010), prostate gland and pelvic floor (Chen, et al., 2005). All of those morphologic studies suggest that the dichotomizing DRG neurons play an important role in visceral refereed pain and visceral organ cross-sensitization. Previous study suggested the averaged pre-spinal somato-visceral convergence was 2% (Dawson, et al., 1992). The c-fos expression and transneuronal viral tracing double labeled method in our study specifically identified the sensory innervation of both groin and uterus. The Fos expression and PrV infected double neurons can be found mainly in T11, T12, T13, L1 and S1 dorsal root ganglia about 3% (fig. 7 B). Our result strongly suggests the existent of dichotomizing sensory neurons in dorsal root ganglia innervating both the uterus and the groin region.. 4-4-2 Double labeled neurons in the nucleus of solitary tract NTS receives either general or special visceral sensory inputs, including visceral pain. 27.
(40) (Altschuler, et al., 1991, Appia, et al., 1986). Previous research also showed Fos protein expression in the NTS after noxious trigeminal stimulation with capsaicin (Ter Horst, et al., 2001). In our study, Fos expression and PrV infected double neurons in the NTS suggest noxious somatic stimulation from the groin region which activates uterus-related neurons in the NTS. The NTS may play an important role in the referred uterine pain to the groin region.. 4-4-3 No double labeled neurons in spinal dorsal horn The convergence-projection theory suggests that neurons in spinal cord receive both visceral and somatic inputs and project through nociceptive pathway to the thalamus (Cervero, 1985). Electrophysiological researches and HRP tracer studies showed the lamina I and V of spinal cord receiving stimulations from both the somatic and visceral tissues, respectively (Bolser, et al., 1991, Cervero, 1983, Cervero, 1985, Cervero and Connell, 1984, Ellrich, et al., 1999, Foreman, et al., 1984). These studies propose the possibilities of the convergence of the visceral and cutaneous afferents onto the neurons found in the spinal dorsal horn. Our study however, showed no Fos expression and PrV infected double neurons in spinal dorsal horn. The reasons which cause the difference could be the various methodologies. Previous studies used electrical stimulation or neuronal tracer injection in the visceral (splanchnic) and somatic (intercostal) nerve,. 28.
(41) respectively. Instead, our study used bee venom and PrV injection specifically and, in turn, identified the sensory innervation to a visceral organ (uterus) and its corresponding somatic area (the groin region). Therefore, we confirmed the dichotomizing DRG neurons as the recipient of the afferents both from the uterus and groin region and proved to be the morphological evidence of referred pain (Chen, et al., 2005, Laurberg and Sorensen, 1985, Pierau, et al., 1984, Sameda, et al., 2003, Takahashi, et al., 2010). Another reason of the difference could be the number of dichotomizing neurons in the dorsal root ganglia. The number of dichotomizing sensory neurons varies in different studies from 0.1% to 21% (Dawson, et al., 1992, Taylor, et al., 1983). Our date also showed that the number of dichotomizing neurons are less than 5% (fig. 7 B). The scarcity of the number of dichotomizing neurons may result in the previous studies which did not acknowledge the fact that the DRG may play an important role in the referred pain.. 4-4-4 Somatic and visceral sensory convergence – the referred pain pathway of the uterus and groin region The dichotomizing DRG neuron is the first order sensory neuron of the referred pain pathway. Furthermore in our study, Fos expression and PrV infected double neurons were observed in the NTS rather than in the spinal level. NTS receives not only visceral pain but was also activated by somatic noxious stimulation (Altschuler, et al., 1991, Appia, et al.,. 29.
(42) 1986, Ter Horst, et al., 2001). This morphological evidence showed that the first order sensory neurons of the referred pain pathway should be the dichotomizing DRG neurons and the second order sensory neuron is in the NTS (fig. 9). In summary, our study provides morphological evidence that the sensory innervation between the uterus and groin region comes from the dichotomizing DRG neuron and project to the NTS. It is the morphological substrate of uterine referred pain.. 4-5 Somatic sensory and visceral motor convergence 4-5-1 Double labeled neurons in the NTS, DMX, and PVN Retrograde tracer injection in the NTS suggests that the spinal superficial dorsal horn neurons (lamina I) directly project to the NTS (Esteves, et al., 1993). Our result (Fig 2A) and previous study (Ter Horst, et al., 2001) also showed that the NTS can be activated by somatic noxious stimulation. Several anatomical and electrophysiological studies have shown the neuronal connection between the uterus and NTS, DMX through the vagus nerve (Hubscher and Berkley, 1994, Lee and Erskine, 2000, Ortega-Villalobos, et al., 1990, Papka, et al., 1998, Wiesel, et al., 2004). The PrV infected neurons in the NTS and DMX (Fig 8) also confirmed the efferent innervation of the NTS and DMX to the uterus as previous study (Ortega-Villalobos, et al., 1990). Neurons with cell bodies in the DMX send their dendrites into the gelatinosus solitary nucleus, where they receive synaptic inputs. 30.
(43) from gastric sensory afferents (Rinaman, et al., 1989). These researches suggested that the NTS and DMX play important roles in visceral innervation, including the digestive functions of the stomach and baroreceptor reflex (Chang, et al., 2003, Herman, et al., 2008, Rinaman, et al., 1989, Spary, et al., 2008, Streefland, et al., 1998, Travagli, et al., 2006). The double labeled NTS and DMX neurons in our study suggest that the nuclei may be one of the relay centers of the somatovisceral reflex of the groin A-shi point and uterus. Retrograde labeling study shows that the NTS receives spinal projections from the superficial dorsal horn (Esteves, et al., 1993) and projects to the PVN (Pan, et al., 1999). Fos expression studies also suggested that the PVN could be activated by somatic noxiuos stimulations (Pan, et al., 1994, Pan, et al., 1999). Swanson proved that the PVN, a higher hormonal regulation center, projects neuronal fibers to the pituitary gland, medulla, and spinal cord (Swanson and Kuypers, 1980). The neuronal connection between the PVN and the uterus was also proved by previous indicators (Lee and Erskine, 2000, Papka, et al., 1998, Wiesel, et al., 2004) and our studies (Fig 8). Noxious stimulation and retrograde tracer injection in different visceral organs showed that the PVN was not only receive noxious afferent but also innervate visceral organs as well (Gerendai, et al., 2001, Papka, et al., 1998, Sinniger, et al., 2004, Traub, et al., 1996, Wang, et al., 2009). The reflex inhibition of the heart rate elicited by acupuncture-like stimulation likely occurs through the activation of the hypothalamic nucleus, which inhibits the activity of premotor. 31.
(44) sympathetic neurons in the rostral ventrolateral medulla (rVLM) (Li and Longhurst, 2010, Uchida, et al., 2008). Although bee venom stimulation in groin A-shi point induced less neuronal activity in the PVN, comparing to saline group (Table 1) in our study, but there was higher percentage of double labeled neurons in the PVN than saline injection group (Fig. 8D). Those previous researches and our result suggest that the PVN may be another relay center of the somatovisceral reflex between the groin and uterus. Although the electrophysiological studies show lamina I and V of the spinal dorsal horn receive afferent information from both somatic and visceral tissues (Bolser, et al., 1991, Ellrich, et al., 1999, Foreman, et al., 1984, Hobbs, et al., 1992, Takahashi and Yokota, 1983), however, no double labeled spinal dorsal neurons can be detected in our study. This solid evidence shows the neuronal connection between the groin region and the uterus is not admitted through the spinal cord.. 4-5-2 The morphologic evidence of somatovisceral reflex – a possible neuronal pathway of acupuncture Previous study suggested that acupuncture may influence visceral function via the activation of the somatosensory neurons (Kim, et al., 2012). However, most researches focused on the physiological responses induced by acupuncture (Chao, et al., 1999, Peng, 2002, Sun and Yao, 1985). The purpose of our study is to investigate and provide the. 32.
(45) morphological evidence of somatovisceral reflex and possible neuronal pathway of acupuncture. Our result suggests that the PVN, NTS, and DMX could be the relay center of the somatovisceral reflex. The visceral organs usually receive sympathetic and parasympathetic dual interacted and the interaction is antagonistic. Therefore, our study proved morphological evidence of both sympathetic and parasympathetic pathways of somatovisceral reflex between the groin A-shi point and the uterus (Fig. 10). The somato-parasympathetic pathway starts from the stimulation of the groin A-shi point, which activates neurons in the spinal dorsal horn. The signal in turn elicits noxious input to the NTS (Esteves, et al., 1993). Neurons in the NTS relay the information and project to the DMX (Norgren, 1978), which innervates the uterus through the vagus nerve (Ortega-Villalobos, et al., 1990) (Fig. 10 A). The somato-sympathetic pathways from the neurons in the spinal dorsal horn project to the NTS (Esteves, et al., 1993) than direct connection to the PVN (Ter Horst, et al., 1989), it innervates the visceral organ through the sympathetic pre- and postganglionic neurons (Hosoya, et al., 1991) (Fig. 10 B). These complementary somato-sympathetic and -parasympathetic systems coincidentally match the concept of yin-yang theory in traditional Chinese medicine (Paton, et al., 2005, World Health Organization. Regional Office for the Western Pacific., 1993). The present study provides the morphological evidences of the neuronal connection between somatic groin A-shi point and its corresponding visceral organ-uterus. Therefore,. 33.
(46) we come up to the conclusion that the somato-sympathetic/-parasympathetic pathways are the morphological basis of somatovisceral reflex and also the neuronal substrate of acupuncture pathways.. In conclusion, the study helps us better understanding the interaction between somatic and visceral system. The mechanism of somatovisceral reflex, acupuncture and referred pain are all involved the connection between somatic and visceral system. This study provides the morphological evidences of the pathways between somatic and visceral system.. 34.
(47) 35.
(48) Figure 1 Fos expression neurons in dorsal root ganglia A:Fos expression neuron (arrow) in T13 DRG (scale bar : 100um) B:Fos expression neurons in T10-S1 DRG, T10(8.7±3.4), T11(5.0±1.5), T12(6.3±1.5), T13(7.9±3.4), L1(13.1±4.4), L2(5.4±1.9), L3(0.9±0.5), L4(0.±0.1), L6(5.1±2.1) and S1(10.3±4.6) (±SEM). 36.
(49) 37.
(50) Figure 2 Fos expression neurons in the spinal cord after bee venom injection in the left groin region (n =9) A: Neurons express Fos protein (arrow) in ipsilateral L2 spinal dorsal horn. B: Higher magnification of Fos expression neurons in A (scale bar : 100um). C: Mean number of Fos expression neurons in T10 to S3 spinal segments (±SEM). *P<0.05 D: Mean number of Fos expression neurons in lamina I to V of L2 spinal segment.. 38.
(51) 39.
(52) Figure 3 Fos expression neurons (arrow) in the supraspinal area after bee venom injection in the left groin region (n=9). A: nucleus of solitary tract (NTS). B: locus coeruleus (LC), parabrachial nucleus (PB). C: raphe pallidus nucleus (RPa). D: paraventricular. hypothalamic nucleus (PVN). E: lateral hypothalamic area. F: paraventricular thalamic nucleus (PV). 3 V: third ventricle; 4 V: fourth ventricle; D3 V: dorsal third ventricle; d: dorsal; f: fornix; ic: internal capsule; l: lateral; m: medial; opt: optic tract; scp: superior cerebellar peduncle; v: ventral (scale bar : 100um).. 40.
(53) 41.
(54) Table 1 Fos expression neurons in supraspinal area between saline and bee venom injection group. (*:significant difference between saline group, p<0.1). 42.
(55) Fig. 4 PrV infected neurons in dorsal root ganglia 7~8 days after PrV injection in left uterine horn A:PrV infected neuron (arrow) in L1 dorsal root ganglion B:Mean of PrV infected neurons in T10~S1 dorsal root ganglia (±SEM). 43.
(56) Figure 5 PrV-infected neurons in spinal cord after 6 to 8 d PrV-injection in the left uterine horn (n=8). A: PrV-infected neuron (arrow) in T12 spinal segment. B: Higher magnification of PrV-infected neuron in A (arrow) (scale bar : 100um) C: Mean number of PrV-infected neurons inT10 to S2 spinal segments (±SEM).. 44.
(57) 45.
(58) Figure 6 PrV-infected neurons (arrow) in the supraspinal area after 7-8 d PrV injection in the left uterus horn (n=8). A: PrV-infected neurons in the NTS and DMX. B: PrV-infected neurons in the A5 noradrenaline cell group. C: PrV-infected neurons in the RPa and RMg. D: PrV-infected neurons in the Gi. E: PrV-infected neurons in the PVN. 3 V: third ventricle; AP: area postrema; DMX: motor nucleus of vagus; Gi: gigantocellular reticular nucleus; Gr: gracile nucleus; NTS: nucleus of solitary tract; PVN: paraventricular hypothalamic nucleus; RPa: raphe pallidus nucleus; RMg: raphe magnus nucleus (scale bar : 100um).. 46.
(59) Table 2 PrV infected neurons in supraspinal area. (+:1-3 infected neurons, ++:4-8 infected neurons, +++:>9 infected neurons). 47.
(60) Figure 7 Double labeled of Fos expression and PrV infected neurons in dorsal root ganglia Fos expression neurons (A, red), PrV infected neurons (B, green) and merged double labeled Fos expression and PrV infected neuron (C, yellow) in T11 DRG. D:Percentage of Fos expression and PrV infected double dorsal root ganglion neurons (double labeled / PrV infected neurons) through T10-S1 spinal segment.. 48.
(61) 49.
(62) Figure 8 Double-labeled neurons of Fos expression and PrV infection (n=8) Fos expression neurons (A, red, arrow) in the PVN (A1), NTS (A2, A3) and DMX (A2, A4). PrV-infected neurons (B, green, arrow) in the PVN (B1), NTS (B2, B3) and DMX (B2, B4). Merged double labeling Fos expression and PrV-infected neurons (C, yellow, arrow) in the PVN (C1), NTS (C2, C3), and DMX (C2, C4) (scale bar : 1000um). D : The percentage of double labeled neurons in PVN, DMX and NTS between saline and bee venom injection groups (*:p<0.1). 50.
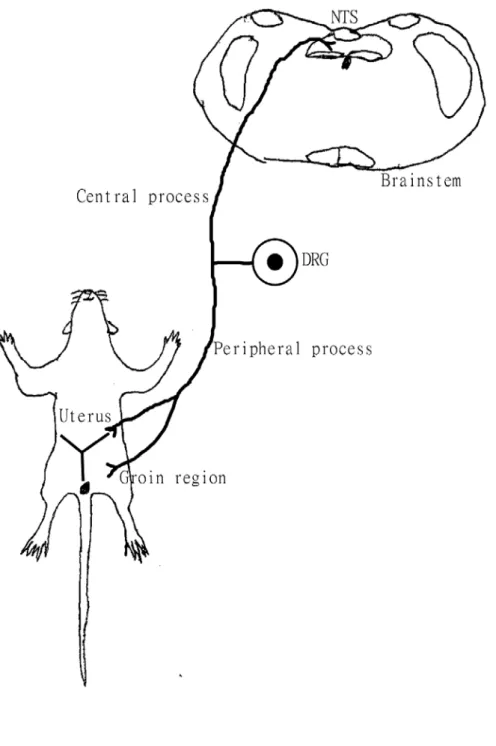
(63) Figure 9 Diagram showing the peripheral process of dichotomizing DRG neurons receive afferents from both the uterus and groin. The central process of the dichotomizing DRG neurons projects to the NTS.. 51.
(64) Figure 10 Schematic drawing three neuronal pathway of somatovisceral reflex. A:Somato-parasympathetic reflex pathway through the vagus nerve B:Somato-sympathetic reflex pathways through the NTS and PVN DMX:dorsal motor nucleus of vagus. DRG:dorsal root ganglia. NTS:nucleus of. solitary tract SPN:sympathetic preganglionic neurons. PVN:paraventricular hypothalamic nucleus. Blue:somatic afferent Red:visceral efferent. 52.
(65) Reference 1.. Al-Chaer, E. D., Feng, Y., and Willis, W. D., 1998. A role for the dorsal column in. 2.. nociceptive visceral input into the thalamus of primates. J Neurophysiol 79, 3143-3150. Altschuler, S. M., Ferenci, D. A., Lynn, R. B., and Miselis, R. R., 1991. Representation of the cecum in the lateral dorsal motor nucleus of the vagus nerve and commissural subnucleus of the nucleus tractus solitarii in rat. J Comp Neurol. 3.. 4.. 304, 261-274. Appia, F., Ewart, W. R., Pittam, B. S., and Wingate, D. L., 1986. Convergence of sensory information from abdominal viscera in the rat brain stem. Am J Physiol 251, G169-175. Arvidsson, J., and Pfaller, K., 1990. Central projections of C4-C8 dorsal root ganglia in the rat studied by anterograde transport of WGA-HRP. J Comp Neurol. 5.. 292, 349-362. Ballegaard, S., Jensen, G., Pedersen, F., and Nissen, V. H., 1986. Acupuncture in. 6.. severe, stable angina pectoris: a randomized trial. Acta Med Scand 220, 307-313. Bao, Y. X., Yu, G. R., Lu, H. H., Zhen, D. S., Cheng, B. H., and Pan, C. Q., 1982.. 7.. Acupuncture in acute myocardial infarction. Chin Med J (Engl) 95, 824-828. Berkley, K. J., Robbins, A., and Sato, Y., 1988. Afferent fibers supplying the uterus. 8.. 9.. in the rat. J Neurophysiol 59, 142-163. Bolser, D. C., Hobbs, S. F., Chandler, M. J., Ammons, W. S., Brennan, T. J., and Foreman, R. D., 1991. Convergence of phrenic and cardiopulmonary spinal afferent information on cervical and thoracic spinothalamic tract neurons in the monkey: implications for referred pain from the diaphragm and heart. J Neurophysiol 65, 1042-1054. Brinkman, J., Bush, B. M., and Porter, R., 1978. Deficient influence of peripheral stimuli on precentral neurones in monkeys with dorsal column lesions. J Physiol. 10.. 276, 27-48. Brumovsky, P. R., and Gebhart, G. F., 2010. Visceral organ cross-sensitization —. 11.. An integrated perspective. Autonomic Neuroscience 153, 106-115. Burton, H., 1968. Somatic sensory properties of caudal bulbar reticular neurons in. 12.. 13.. the cat (Felis domestica). Brain Res 11, 357-372. Calaresu, F. R., and Henry, J. L., 1970. The mechanism of the cardio-acceleration elicited by electrical stimulation of the parahypoglossal area in the cat. J Physiol 210, 107-120. Cardona, F., 1950. [Study of the medullary pyramidal tracts in cerebral hemiplegia. 53.
(66) 14.. 15. 16.. due to capsular lesions]. Riv Patol Nerv Ment 71, 216-224. Carstens, E., and Trevino, D. L., 1978. Anatomical and physiological properties of ipsilaterally projecting spinothalamic neurons in the second cervical segment of the cat's spinal cord. J Comp Neurol 182, 167-184. Cervero, F., 1983. Somatic and visceral inputs to the thoracic spinal cord of the cat: effects of noxious stimulation of the biliary system. J Physiol 337, 51-67. Cervero, F., 1985. Visceral nociception: peripheral and central aspects of visceral nociceptive systems. Philosophical transactions of the Royal Society of London.. 17.. Series B, Biological sciences 308, 325-337. Cervero, F., 1995. Visceral pain: mechanisms of peripheral and central sensitization.. 18.. Ann Med 27, 235-239. Cervero, F., and Connell, L. A., 1984. Distribution of somatic and visceral primary. 19.. afferent fibres within the thoracic spinal cord of the cat. J Comp Neurol 230, 88-98. Cervero, F., Connell, L. A., and Lawson, S. N., 1984. Somatic and visceral primary afferents in the lower thoracic dorsal root ganglia of the cat. J Comp Neurol 228, 422-431.. 20. 21.. 22.. 23.. Cervero, F., and Laird, J. M., 1999. Visceral pain. Lancet 353, 2145-2148. Chaban, V., Christensen, A., Wakamatsu, M., McDonald, M., Rapkin, A., McDonald, J., and Micevych, P., 2007. The same dorsal root ganglion neurons innervate uterus and colon in the rat. Neuroreport 18, 209-212. Chang, H. Y., Mashimo, H., and Goyal, R. K., 2003. Musings on the wanderer: what's new in our understanding of vago-vagal reflex? IV. Current concepts of vagal efferent projections to the gut. Am J Physiol Gastrointest Liver Physiol 284, G357-366. Chao, D. M., Shen, L. L., Tjen, A. L. S., Pitsillides, K. F., Li, P., and Longhurst, J. C., 1999. Naloxone reverses inhibitory effect of electroacupuncture on sympathetic. 24.. cardiovascular reflex responses. Am J Physiol 276, H2127-2134. Chen, Y., Song, B., Jin, X. Y., Xiong, E. Q., and Zhang, J. H., 2005. Possible mechanism of referred pain in the perineum and pelvis associated with the prostate. 25.. in rats. J Urol 174, 2405-2408. Chernigovskii, V. N., and Musiashchikova, S. S., 1969. [Participation of the ventral posterior-lateral nucleus of the cat's thalamus in the transmission of afferent. 26.. visceral signals]. Izv Akad Nauk SSSR Biol 1, 5-19. Chien, C. H., Shieh, J. Y., Liao, M. H., Ling, E. A., and Wen, C. Y., 1998. Neuronal connections between the auricular skin and the sympathetic pre- and postganglionic neurons of the dog as studied by using pseudorabies virus. Neurosci Res 30, 169-175.. 54.
(67) 27.. 28. 29.. 30.. Christianson, J. A., Liang, R., Ustinova, E. E., Davis, B. M., Fraser, M. O., and Pezzone, M. A., 2007. Convergence of bladder and colon sensory innervation occurs at the primary afferent level. Pain 128, 235-243. Cliffer, K. D., and Giesler, G. J., Jr., 1989. Postsynaptic dorsal column pathway of the rat. III. Distribution of ascending afferent fibers. J Neurosci 9, 3146-3168. Collins, J. J., Lin, C. E., Berthoud, H. R., and Papka, R. E., 1999. Vagal afferents from the uterus and cervix provide direct connections to the brainstem. Cell Tissue Res 295, 43-54. Coote, J. H., and Macleod, V. H., 1974. Evidence for the involvement in the. 31.. baroreceptor reflex of a descending inhibitory pathway. J Physiol 241, 477-496. Craig, A. D., 1991. Spinal distribution of ascending lamina I axons anterogradely labeled with Phaseolus vulgaris leucoagglutinin (PHA-L) in the cat. J Comp Neurol. 32.. 313, 377-393. Craig, A. D., Jr., Linington, A. J., and Kniffki, K. D., 1989. Cells of origin of spinothalamic tract projections to the medial and lateral thalamus in the cat. J Comp. 33. 34.. 35. 36.. 37.. Neurol 289, 568-585. Curran, T., and Morgan, J. I., 1995. Fos: an immediate-early transcription factor in neurons. J Neurobiol 26, 403-412. Danilov, A. B., and Vein, A. M., 1996. [The nociceptive flexor reflex: a method for studying the brain mechanisms of pain control]. Zh Nevrol Psikhiatr Im S S Korsakova 96, 107-111. Dawson, N. J., Schmid, H., and Pierau, F. K., 1992. Pre-spinal convergence between thoracic and visceral nerves of the rat. Neurosci Lett 138, 149-152. de Medeiros, M. A., Canteras, N. S., Suchecki, D., and Mello, L. E., 2003. Analgesia and c-Fos expression in the periaqueductal gray induced by electroacupuncture at the Zusanli point in rats. Brain Res 973, 196-204. DeGroat, W. C., and Saum, W. R., 1976. Synaptic transmission in parasympathetic. 38.. ganglia in the urinary bladder of the cat. J Physiol 256, 137-158. Dmitriev, V. D., 1957. [Morphological and physiological analysis of compensatory mechanisms of sensory and motor functions following injuries of the spinal cord].. 39.. Arkh Patol 19, 50-57. Ellrich, J., Andersen, O. K., Messlinger, K., and Arendt-Nielsen, L., 1999. Convergence of meningeal and facial afferents onto trigeminal brainstem neurons:. 40.. an electrophysiological study in rat and man. Pain 82, 229-237. Esteves, F., Lima, D., and Coimbra, A., 1993. Structural types of spinal cord marginal (lamina I) neurons projecting to the nucleus of the tractus solitarius in the rat. Somatosens Mot Res 10, 203-216.. 55.
(68) 41.. 42.. Foreman, R. D., Blair, R. W., and Weber, R. N., 1984. Viscerosomatic convergence onto T2-T4 spinoreticular, spinoreticular-spinothalamic, and spinothalamic tract neurons in the cat. Exp Neurol 85, 597-619. Gebhart, G. F., and Ness, T. J., 1991. Central mechanisms of visceral pain. Can J. 43.. Physiol Pharmacol 69, 627-634. Gerendai, I., Toth, I. E., Kocsis, K., Boldogkoi, Z., Medveczky, I., and Halasz, B., 2001. Transneuronal labelling of nerve cells in the CNS of female rat from the. 44.. mammary gland by viral tracing technique. Neuroscience 108, 103-118. Ghanima, A., Bennis, M., and Rampin, O., 2002. c-Fos expression as endogenous marker of lumbosacral spinal neuron activity in response to. 45.. 46.. 47.. 48.. 49.. 50.. 51.. vaginocervical-stimulation. Brain Res Brain Res Protoc 9, 1-8. Giamberardino, M. A., Berkley, K. J., Iezzi, S., deBigontina, P., and Vecchiet, L., 1997. Pain threshold variations in somatic wall tissues as a function of menstrual cycle, segmental site and tissue depth in non-dysmenorrheic women, dysmenorrheic women and men. Pain 71, 187-197. Goldman, P. L., Collins, W. F., Taub, A., and Fitzmartin, J., 1972. Evoked bulbar reticular unit activity following delta fiber stimulation of peripheral somatosensory nerve in cat. Exp Neurol 37, 597-606. Gray, T. S., Hazlett, J. C., and Martin, G. F., 1981. Organization of projections from the gracile, medial cuneate and lateral nuclei in the North American opossum. Horseradish peroxidase study of the cells projecting to the cerebellum, thalamus and spinal cord. Brain Behav Evol 18, 140-156. Groenewegen, H. J., Boesten, A. J., and Voogd, J., 1975. The dorsal column nuclear projections to the nucleus ventralis posterior lateralis thalami and the inferior olive in the cat: an autoradiographic study. J Comp Neurol 162, 505-517. Guevara-Guzman, R., Buzo, E., Larrazolo, A., de la Riva, C., Da Costa, A. P., and Kendrick, K. M., 2001. Vaginocervical stimulation-induced release of classical neurotransmitters and nitric oxide in the nucleus of the solitary tract varies as a function of the oestrus cycle. Brain Res 898, 303-313. Herman, M. A., Niedringhaus, M., Alayan, A., Verbalis, J. G., Sahibzada, N., and Gillis, R. A., 2008. Characterization of noradrenergic transmission at the dorsal motor nucleus of the vagus involved in reflex control of fundus tone. Am J Physiol Regul Integr Comp Physiol 294, R720-729. Ho, M., Huang, L. C., Chang, Y. Y., Chen, H. Y., Chang, W. C., Yang, T. C., Tsai, H. D. C. I. N. T. J. O. G. S., and author reply, P., 2009. Electroacupuncture reduces uterine artery blood flow impedance in infertile women. Taiwanese journal of obstetrics & gynecology 48, 148-151.. 56.
數據
相關文件
Robinson Crusoe is an Englishman from the 1) t_______ of York in the seventeenth century, the youngest son of a merchant of German origin. This trip is financially successful,
fostering independent application of reading strategies Strategy 7: Provide opportunities for students to track, reflect on, and share their learning progress (destination). •
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
volume suppressed mass: (TeV) 2 /M P ∼ 10 −4 eV → mm range can be experimentally tested for any number of extra dimensions - Light U(1) gauge bosons: no derivative couplings. =>
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
• Formation of massive primordial stars as origin of objects in the early universe. • Supernova explosions might be visible to the most
Monopolies in synchronous distributed systems (Peleg 1998; Peleg
Corollary 13.3. For, if C is simple and lies in D, the function f is analytic at each point interior to and on C; so we apply the Cauchy-Goursat theorem directly. On the other hand,
