科技部補助專題研究計畫報告
循環經濟之二氧化碳利用與高值化技術開發
報 告 類 別 : 成果報告 計 畫 類 別 : 個別型計畫
計 畫 編 號 : MOST 108-2621-M-006-015- 執 行 期 間 : 108年08月01日至109年07月31日 執 行 單 位 : 國立成功大學化學工程學系(所)
計 畫 主 持 人 : 吳意珣
計畫參與人員: 博士班研究生-兼任助理:譚世一 報 告 附 件 : 出席國際學術會議心得報告
本研究具有政策應用參考價值:□否 ■是,建議提供機關科技部
(勾選「是」者,請列舉建議可提供施政參考之業務主管機關)
本研究具影響公共利益之重大發現:■否 □是
中 華 民 國 109 年 09 月 09 日
中 文 摘 要 : 全球暖化已備受關注,且急需發展技術來封存利用二氧化碳或是減 緩二氧化碳濃度上升。在已發展的技術中,利用大腸桿菌來代謝二 氧化碳已成為近年來新興的方法來高值化轉換二氧化碳,然而目前 文獻中所報導之二氧化碳代謝能力卻不一致,其可能由於使用之反 應器有所不同。在此,我們基於質量平衡開發了一種簡單的二氧化 碳代謝評估方法,在考慮“綠色製程”下來評估大腸桿菌同化二氧 化碳的能力。接著,為了全面性評估反應器的影響,我們設計了三 種不同的設備:錐形瓶放置在二氧化碳培養箱(Flask-based device in the CO2 incubator, FIC)、雙層裝置(Two-layered device, TLD)和二氧化碳通氣裝置(CO2 bubbling device, CBD)。結果顯示,在CBD通以5%的二氧化碳賦予了二氧化碳對 RuBisCo的有效供給,二氧化碳同化能力從-5.03增加到了-2.63 g- CO2/g-DCW。此外,通過共表達GroELS分子伴侶可增強二氧化碳同化 能力,且二氧化碳釋放減少了71%。另一方面,通過DNA測序和串聯 質譜分析,確定了RuBisCo和PRK的毒性會干擾大腸桿菌的糖代謝系 統,從而影響能量代謝及細胞形態的改變。我們的研究為加強二氧 化碳同化提供了一個新的視角,以實現可持續的生態友好型綠色生 物製程。
中 文 關 鍵 詞 : 二氧化碳、大腸桿菌、RuBisCo、綠色製程、同化能力
英 文 摘 要 : Global warming is a surging issue that has provoked the demand of green process to mitigate CO2 concentration. In this context, CO2-assimilable E. coli has been reported, while possessed different CO2 assimilation capability probably due to the non-unified bioreactor. Herein, we first developed a simple assessment based on the mass balance to evaluate the CO2 assimilation capability with the consideration of “green process.” Next, we designed three different devices: Flask-based device in the CO2 incubator (FIC), two-layered device (TLD) and CO2 bubbling device (CBD) to exam device effect on the CO2 assimilation capability of RuBisCo-equipped E. coli systematically. With the forced diffusion of 5% CO2 in CBD, which confers an efficient attack of CO2 to RuBisCo, the CO2 assimilation capability increased from -5.03 to -2.63 g-CO2/g-DCW.
Furthermore, boosted CO2 assimilation ability was observed by co-expression of GroELS chaperone with 71% reduction on CO2 release. By DNA sequencing and tandem MS/MS analysis, the toxicity of RuBisCo and PRK was identified to interfere the sugar metabolism, energy producing and cell morphology in E. coli. Our study provides a new perspective into the enhancement of CO2 assimilation for sustainable to eco- friendly green bioprocess.
英 文 關 鍵 詞 : Carbon dioxide, Escherichia coli, RuBisCo, Green Bioprocess, Assimilation
科技部補助專題研究計畫 ■成果報告
□期中進度報告
循環經濟之二氧化碳利用與高值化技術開發
計畫類別:■個別型計畫 □整合型計畫
計畫編號:MOST 108-2621-M006-015
執行期間:108 年 8 月 1 日至 109 年 07 月 31 日
執行機構及系所:國立成功大學化工系 計畫主持人: 吳意珣 副教授
計畫參與人員: 譚世一 博士班研究生
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告 本計畫除繳交成果報告外,另須繳交以下出國心得報告:
□赴國外出差或研習心得報告 □赴大陸地區出差或研習心得報告
□出席國際學術會議心得報告 □國際合作研究計畫國外研究報告 處理方式:二年後公開
中 華 民 國 109 年 09 月 10 日
摘要
全球暖化已備受關注,且急需發展技術來封存利用二氧化碳或是減緩二氧 化碳濃度上升。在已發展的技術中,利用大腸桿菌來代謝二氧化碳已成為近年 來新興的方法來高值化轉換二氧化碳,然而目前文獻中所報導之二氧化碳代謝 能力卻不一致,其可能由於使用之反應器有所不同。在此,我們基於質量平衡 開發了一種簡單的二氧化碳代謝評估方法,在考慮“綠色製程”下來評估大腸 桿菌同化二氧化碳的能力。接著,為了全面性評估反應器的影響,我們設計了 三種不同的設備:錐形瓶放置在二氧化碳培養箱(Flask-based device in the CO2
incubator, FIC)、雙層裝置(Two-layered device, TLD)和二氧化碳通氣裝置
(CO2 bubbling device, CBD)。結果顯示,在 CBD 通以 5%的二氧化碳賦予了 二氧化碳對 RuBisCo 的有效供給,二氧化碳同化能力從-5.03 增加到了-2.63 g-
CO2/g-DCW。此外,通過共表達 GroELS 分子伴侶可增強二氧化碳同化能力,
且二氧化碳釋放減少了 71%。另一方面,通過 DNA 測序和串聯質譜分析,確 定了 RuBisCo 和 PRK 的毒性會干擾大腸桿菌的糖代謝系統,從而影響能量代 謝及細胞形態的改變。我們的研究為加強二氧化碳同化提供了一個新的視角,
以實現可持續的生態友好型綠色生物製程。
Abstract
Global warming is a surging issue that has provoked the demand of green process to mitigate CO2 concentration. In this context, CO2-assimilable E. coli has been reported, while possessed different CO2 assimilation capability probably due to the non-unified bioreactor. Herein, we first developed a simple assessment based on the mass balance to evaluate the CO2 assimilation capability with the consideration of
“green process.” Next, we designed three different devices: Flask-based device in the CO2 incubator (FIC), two-layered device (TLD) and CO2 bubbling device (CBD) to exam device effect on the CO2 assimilation capability of RuBisCo-equipped E. coli systematically. With the forced diffusion of 5% CO2 in CBD, which confers an efficient attack of CO2 to RuBisCo, the CO2 assimilation capability increased from - 5.03 to -2.63 g-CO2/g-DCW. Furthermore, boosted CO2 assimilation ability was observed by co-expression of GroELS chaperone with 71% reduction on CO2 release.
By DNA sequencing and tandem MS/MS analysis, the toxicity of RuBisCo and PRK was identified to interfere the sugar metabolism, energy producing and cell
morphology in E. coli. Our study provides a new perspective into the enhancement of CO2 assimilation for sustainable to eco-friendly green bioprocess.
Content
摘要 ... 2
Abstract
... 3
Content
... 4
Figure Content
... 5
Table Content
... 6
Chapter 1 Introduction and Research Goal
... 7
Chapter 2 Literature Review
... 8
2.1 Biological approach to capture and store the CO2 ... 8
2.2 Manipulating mirobes to assimilate CO2 ... 8
2.3 Device effect on microbial cell factory ... 9
Chapter 3 Methods and Materials
... 11
3.1 Strains, plasmids, primers, and media ... 11
3.2 Plasmid Contruction ... 12
3.3 Plasmid Transformation into BL21(DE3) ... 12
3.4 Different bioreactor setup ... 13
3.5 Calibration of biomass by infrared moisture analyzer and element analysis ... 13
3.6 Protein analysis by SDS-PAGE and tandem MS/MS ... 14
3.7 HPLC analysis of metabolites ... 14
3.8 Scanning electron microscopy (SEM) analysis ... 15
Chapter 4 Results and Disccusion
... 16
4.1 Calculation of CO2 assimilation by mass balance ... 16
4.2 Effect of bioreactors on the CO2 assimilation ... 19
4.3 Optimization of CO2 utilization in RuBisCo-equipped E. coli ... 22
4.4 Enhance the CO2 assimilation by inducer and chaperone ... 25
4.5 New insight into the toxicity and evolution in the RuBisCo-equipped E. coli ... 29
4.6 Conclusion ... 33
Chapter 5 References
... 34
Figure Content
Figure 1 Schematic illustration of the calculation of CO
2assimilation. ... 17 Figure 2 HPLC spectrum of the medium as well as BL21(DE3), 2P and 3P
strain cultured in the CBD with 5% CO
2bubbling. ... 18 Figure 3 The effect of bioreactoron the capability of CO
2assimilation. ... 21 Figure 4 Effect of the CO
2concentration on the capability of CO
2assimilation in BD strain. ... 23 Figure 5 Effect of the CO
2concentration on the capability of CO
2assimilation in 2P strain. ... 24 Figure 6 Different strategies to enhance the performance of CO
2assimilation. ... 27 Figure 7 Device effect in 3P strain ... 28 Figure 8 Molecular level discovery of the toxicity of RuBisCo pathway in E.
coli... 30
Figure 9 Cell morphology of the CBD-cultured 2P strain by SEM analysis. ... 32
Table Content
Table 1 E. coli strains, plasmids and primers used in this study ... 11 Table 2 Calibration of OD
600to biomass of RuBisCO-equipped E. coli in
different bioreactors. ... 14 Table 3 Element analysis of the carbon content in E. coli. ... 19 Table 4 Summary of specific CO
2assimilation capability under different
conditions in this study ... 28
Table 5 Mascot protein analysis from LC–MS/MS. ... 32
Chapter 1 Introduction and Research Goal
Global warming is currently the surging issue since it strikes an imbalance in the ecological and economic environment. The statics from the Intergovernmental Panel on Climate Change (IPCC) argued that human activity strongly connected to
observable global warming (Change 2014). Therefore, it revokes the invention of strategies to mitigate the atmospheric CO2 concentration, including the CO2 capture, storage, and conversion in recent years (Bui et al., 2018; Goeppert et al., 2012). It has also been reported that CO2 is one of the potential carbon sources for bio-based chemical productions (Mikkelsen et al., 2010). Among all strategies, CO2 conversion is regarded as a prominent route to create alternative pathways of the CO2 into biomass and value-added chemicals. For example, electrical reduction of CO2 has applied to synthesize the molecules with higher energy density, such as formate, alcohol and hydrocarbons, but it still requires the luxurious catalysts (Liang et al.,
2020; Matafa et al., 2020). Alternatively, an auspicious approach is the assimilation of
CO2 by de novo metabolic pathway to produce different high value-added chemicals (Lin et al., 2019; Salehizadeh et al., 2020).For CO2 utilization via the biological way, there has been several publications in which the E. coli was engineered to assimilate CO2 to form the biomass or even other value-added chemical (Antonovsky et al., 2016; d Mattozi et al., 2013; Guadalupe-
Medina et al., 2013; von Borzyskowski et al., 2018), while the CO
2 assimilation capability was different probably due to the genetic design and devices used for cultivation. Thus, in the study, we first comprehensively considered devise effect on the CO2 assimilation capability in RuBisCo-equipped E. coli, which conferred a new insight into engineering E. coi with a better performance of CO2 assimilation.Chapter 2 Literature Review
2.1 Biological approach to capture and store the CO
2Since the industrial revolution, human being exceedingly relied on the fossil fuels as the main energy source. Statistics in 2006 showed that fossil fuels, such as coal mines and petroleum, accounted for 85% of global energy demand (Tubb, 2015).
Therefore, the usage of fossil fuels has led to serious carbon dioxide emissions which was mainly contributed by human economic activities and the use of fossil fuels (Stocker et al., 2013). In such context, increasing the proportion of green energy usage and lessening reliance on fossil fuels as well as the development of carbon capture and storage technology (CCS) was regarded as two major solutions to circumvent the green house effect caused by the emitted carbon dioxide.
Chemical carbon capture technology that uses adsorbents such as ethanolamine and limestone to capture carbon dioxide, although the adsorption efficiency is high, requires a plenty of energy to desorb, separate, transport and store the carbon dioxide as well as regenerate the adsorbent (Lin et al., 2012). At the same time, the adsorbent uasge also cause secondarily environmental pollution. On the other hand, biological carbon dioxide was a cleaner way as comparing to the chemical method. Besides, through the metabolic engineering, the CO2 could be further converted into the value- added chemicals.
2.2 Manipulating mirobes to assimilate CO
2Harnessing autotrophic microbes to assimilate the CO2 is the most straightforward concept to simultaneously fix the CO2 and produce different kinds of biofuel, high value-added chemicals even and functional proteins (Cheah et al., 2016; Levasseur
and Pozzobon, 2020; Thakur et al., 2018). Furthermore, the native pathways could be
enhanced, or non-native pathways could be incorporated for the production of various chemicals by genetic autotrophic microbes (Li et al., 2015a; Lin et al., 2019). Except for the autotrophic microbes, the heterologous organisms have also been engineered to equip the carbon fixation for CO2 utilization by enzymes (Antonovsky et al., 2016;d Mattozi et al., 2013; Guadalupe-Medina et al., 2013; von Borzyskowski et al., 2018).
E. coli is the most promising heterotrophic host to equip the RuBisCo-based
pathway, in which ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCo) and phosphoribulokinase (PRK) are introduced to incorporate the CO2 into metabolism due to highly available genetic tools, simple genetic procedure and high growth rate (Gong et al., 2015; Li et al., 2015b; Zhuang and Li, 2013). Recently, Gleizer and his colleagues transformed the heterotrophic E. coli as an autotrophic host by adaptive laboratory evolution (ALE) at the expense of one year (Gleizer et al., 2019).
Assimilated CO2 could be used to form the biomass and other metabolites. The biomass of E. coli with enzyme overexpression could be used as whole-cell
biocatalyst (He et al., 2018; Wang et al., 2020). Furthermore, the assimilated CO2 has also been demonstrated to synthesize 90% malate production in E. coli (Hu et al.,
2018), showing it is a high potential “green cell factory”.
2.3 Device effect on microbial cell factory
In a cell factory, the bioreactor is one of the determinative influences for targeted functions (Choi et al., 2019; Hu et al., 2012; Show et al., 2011). For example, Luo and his colleagues demonstrated that the production of methyl anthranilate, a widely used cosmetic grape flavoring, could be enhanced from 35.8 mg/L to 4.47 g/L as the
bioreactor changed from flask-based to the two-phased fed-batch bioreactor (Lou et
al., 2019). On the other hand, different bioreactors also affect the efficacy of ALE*.
Formate utilization in E. coli has been explored by ALE recently, in which Kim and his coworkers demonstrated flask-based 150 serial subcultures, around 150 days, conferred an evolved E. coli strain with an optimized ability to utilize formate (Kim et
al., 2019b), while they trained the E. coli to equip a better and faster utilization of
formate within 13 serial subcultures around 80 days in tube-based culturing (Kim etal., 2020). Although the significance of the bioreactor has already been proven in
previous reports for chemical production, there is still limited literature regarding the effect of bioreactor design on the CO2 assimilation. Actually, in the previous reports, the bioreactors used for culturing RuBisCo-equipped E. coli differed from individual publication, which led to incomparable research outcomes.Chapter 3 Methods and Materials
3.1 Strains, plasmids, primers, and media
All the strains, plasmids and primers were listed in the Table 1. The E. coli DH5
was used in regular cloning while E. coli BL21(DE3) was used to express the targeted protein for CO2 assimilation. For the regular culture, Luria–Bertani (LB) medium (1.5% tryptone, 1.5% NaCl, and 0.5% yeast extract) was used, while the MM9XI (12.8 g/L Na2HPO4, 3 g/L KH2PO4, 1 g/L Tris, 0.5 g/L NaCl, 1 g/L NH4Cl, 0.24 g/L MgSO4, 0.011 g/L CaCl2, 5 g/L xylose and 0.1 mM IPTG) was used in the CO2
assimilation.
Table 1 E. coli strains, plasmids and primers used in this study
Name Description Remark
Strains
DH5 F-, Δ(argF-lac)169, φ80dlacZ58(M15), ΔphoA8,
glnX44(AS), λ-, deoR481, rfbC1, gyrA96(NalR), recA1, endA1, thiE1, hsdR17
For cloning
BL21(DE3) thrB1004 pro thi rpsL hsdS lacZΔM15 RP4-1360 Δ(araBAD)567 ΔdapA1341::[erm pir]
This study
2P BL21(DE3) harboring pET30a-rbcLS (6301) and pSB4*A3*-T7-B0034-SePRK
This study
3P BL21(DE3) harboring pET30a-rbcLS (6301), pSB4*A3*-T7-B0034-SePRK and pT-GroE
This study
Plasmids pET30a- rbcLS (6301)
pBR322 ori, KanR, T7 promoter, T7RBS, rbcLS from PCC6301)
Zhang and Li, 2013
pT-GroE p15a ori, CmR, T7 promoter, T7RBS, EcGroELS Yu et al., 2019
pSB4*A3- T7-B0034- sfGFP
pSC101* ori, AmpR and T7 promoter, B0034 RBS, sfGFP This study
pSB4*A3- T7-B0034- SePRK
pSC101* ori, AmpR and T7 promoter, B0034 RBS, PRK from Synechococcus elongatus PCC 7942
This study
Primers
HindIII-prk- F
GCAAGCTTATGAGTAAACCAGACCGCGTAGTGCT G
This study PstI-SpeI-
prk-R
CACTGCAGCGGCCGCTACTAGTTCAAACGGAGGC GGCAACTGGAGCA
This study P.S. Underline indicated the retriction site.
3.2 Plasmid Contruction
The amino acid sequence of phosphoribulokinase (PRK) was from the Synechococcus elongatus PCC 7942 (i.e., SePRK), and the DNA sequence was
designed by the codon-optimization tools and synthesized by the Integrated DNA Technology® (IDT, USA). For cloning the pSB4*A3-T7-B0034-SePRK, the SePRK was first amplified by the HindIII-PRK-F and PstI-SpeI-PRK first and then cloned into the pSB4*A3-T7-B0034-sfGFP by HindIII and PstI. The successful cloning was further analyzed by DNA sequencing.
3.3 Plasmid Transformation into BL21(DE3)
The successful-constructed plasmids were extracted from DH5 and transformed to BL21(DE3) by heat shock. The BL21(DE3) harboring pET30a-rbcLS (6301) and pSB4*A3*-T7-B0034-SePRK was designed as 2P strain, while 3P strain involved the addition of pT-GroE into 2P. For culturing 2P strain, 50 mg/L Ampicillin (Ap) and 25
mg/L Kanamycin (Km) were used, while 25 mg/L Ap, 12.5 mg/L Km, and 8.25 mg/L Cm were used for culturing 3P strain.
3.4 Different bioreactor setup
The RuBisCo-equipped E. coli was precultured in LB medium at 37oC for 12 h and sub-cultured to MM9X at 30oC for further 96 h. Three different bioreactors were set up, including flask-based device in CO2 incubator (FIC), two-layer device (TLD) containing the cell in upper layer as well as 2 M Na2CO3 and 2 M NaHCO3 in the lower layer. The third one is the CO2 bubbling device (CBD) with directly injected CO2 in different concentration. The cell growth was analyzed every 12 h.
3.5 Calibration of biomass by infrared moisture analyzer and element analysis
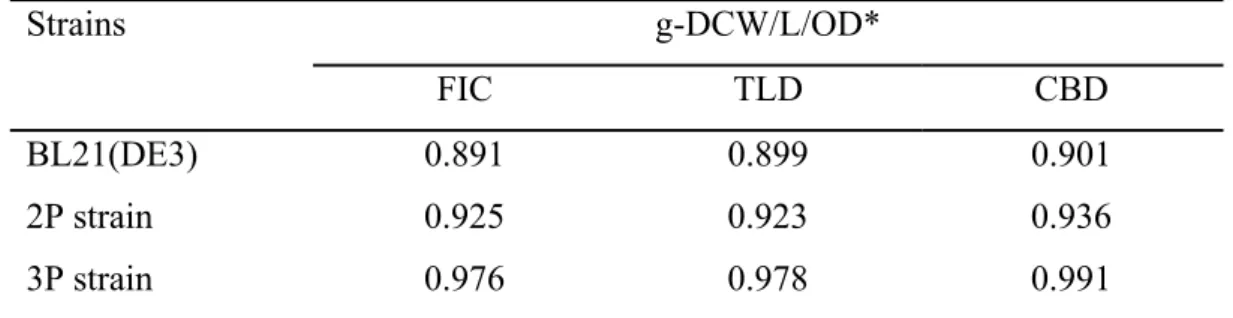
A 200 L of the broth was added into a transparent 96-well plate. Optical density at 600 nm wavelength was measured by the SpectraMax M2 (Molecular Devices, Sunnyvale, CA). For converting the OD600 to the dry cell weight (biomass), the cell was collected and washed for 3 times. Subsequently, the washed cell was suspended in the water and the cell suspension was adjusted to the OD600 at 10. Then, the cell suspension was subjected into the infrared moisture analyzer (Kett, Villa Park, CA) until the weight was constant, and the relationship between the OD600 and biomass was calculated as shown in Table 2. For the carbon content determination in biomass, the cell was washed, collected and dried and then direct subjected into the elemental analyzer (UNICUBE, Germany).
Table 2 Calibration of OD
600to biomass of RuBisCO-equipped E. coli.
Strains g-DCW/L/OD*
FIC TLD CBD
BL21(DE3) 0.891 0.899 0.901
2P strain 0.925 0.923 0.936
3P strain 0.976 0.978 0.991
*Collect the cell at 96 h and dry by infrared-radio dryer until the weight of biomass is kept constantly.
3.6 Protein analysis by SDS-PAGE and tandem MS/MS
The cells were harvested and then washed twice by deionized water. The cell density was adjusted to an OD600 of 4 and analyzed by SDS-PAGE, which contained 12% separating gel and 4% stacking gel. Proteins were visualized by staining with Coomassie blue R-250 and scanned by the Image scanner (Biolab2000, Taiwan). The targeted protein was analyzed by tandem MS/MS according to our previous reports (Lai et al., 2019).
3.7 HPLC analysis of metabolites
For the sample preparation, the medium was collected and then filtered by the 0.22
M filter. Afterward, the sample was subjected to the HPLC analysis (Hitachi, Japan) with the column of ICSep COREGEL-87H3 (Transgenomic, USA). The separation was performed at 70oC and mobile phase of 0.008 N sulfuric acid was flowed with the rate of 0.4 mL/min. The signal was detected by the reflected index.
3.8 Scanning electron microscopy (SEM) analysis
Cell was collected and washed by the 0.1 M phosphate buffer (PB). The washed cell was suspended in the 0.1 M PB with 2.5% glutaraldehyde and stored at 4oC for 4 hours. Afterward, the cell was dropped on the formvar-coated silicon chunk and then serially dehydrated by 100% ethanol. Eventually, the cell was immersed in the tert- butanol and then stored at 4oC for 1 hour. Subsequently, the cell was lyophilized for 12 hours. The cell morphology was observed by SEM (SU8100 Hitachi, Japan).
Chapter 4 Results and Disccusion
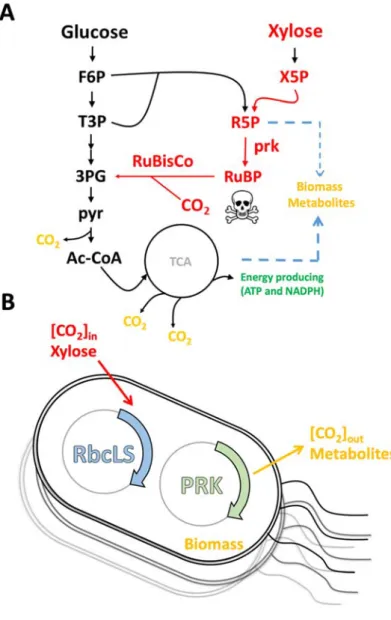
4.1 Calculation of CO
2assimilation by mass balance
As shown in Figure 1A, the CO2 assimilation capability was introduced into the E.
coli through the RuBisCo-based pathway. Due to the lack of substrate for ribulose-
1,5-bisphosphate carboxylase/oxygenase (RuBisCo) among the native metabolites in E. coli, the first step that ribose-5-phosphate (R5P) was phosphorized into ribose-1,5-
phosphate (RuBP) through the phosphoribulokinase (PRK) was introduced. Next, the RuBP and CO2 were fused to form two molecules of 3-phosphoglycerate (3PG) which would be directed into the downstream pathway (i.e., TCA cycle and its branches) to form the biomass, energy blocks and metabolites (Gong et al., 2015).
After introducing two enzymes, i.e., RuBisCo and PRK into the E. coli, the
capability of CO2 assimilation was calculated based on the mass balance (Gong et al.,
2015) in the E. coli cell as a controlled volume (Figure 1B). The input carbons include
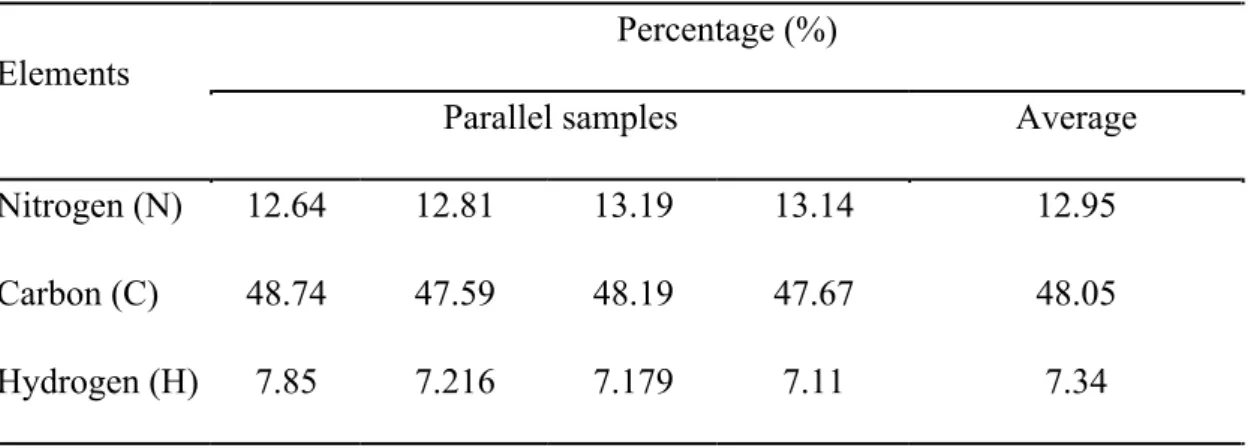
the CO2 and the sole organic carbon of xylose (Equation (1)), in which thecoefficients of 0.27 and 0.4 represent carbon content of the CO2 and xylose. The outputs are the biomass, CO2 and the metabolites (Equation (2)). The carbon content of biomass is 0.48, which was confirmed by the element analysis (Table 3) and consistent with previous report where the carbon content in E. coli was at the range of 45% to 48% (Folsom and Carlson, 2015). The steady-state assumption confers the
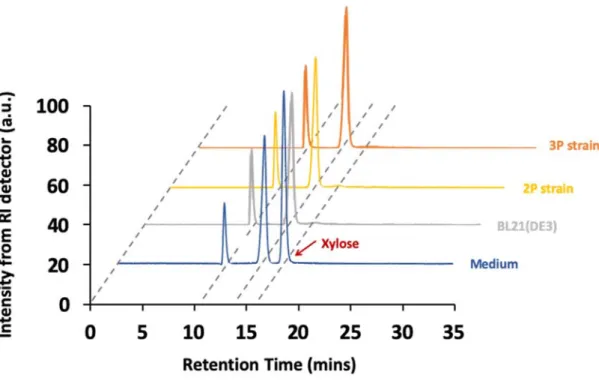
Equation (3) by the input carbons are equal to the output carbons. Next, we simplified
the equation by eliminating the metabolites term which determined by HPLC analysis because no significant organic acid can be detected in the stationary phase (Figure 2), thus, we denoted a parameter of [CO2]net* for evaluation of CO2 assimilationcapability as shown in Equation (4).
Input carbons: 0.27 𝐶𝑂 0.4 𝑋𝑦𝑙𝑜𝑠𝑒 ……… Equation (1) Output carbons: 0.27 𝐶𝑂 0.48 𝐵𝑖𝑜𝑚𝑎𝑠𝑠
∑ 𝐶 , ……… Equation (2)
𝐶𝑂 𝐶𝑂 𝐶𝑂 1.78 𝐵𝑖𝑜𝑚𝑎𝑠𝑠 ∑ ,
.
0.4 𝑋𝑦𝑙𝑜𝑠𝑒 ……… Equation (3) 𝐶𝑂 ∗ 1.78 𝐵𝑖𝑜𝑚𝑎𝑠𝑠 0.4 𝑋𝑦𝑙𝑜𝑠𝑒 ………….…Equation (4)
Figure 1 Schematic illustration of the calculation of CO
2assimilation.
(A) Pathway evolved in the RuBisCo and PRK protein. PRK protein catalyzes the conversion of R5P to RuBP. The RuBisCo performs the CO2 assimilation through
converting RuBP to 3PG. The carbon sources were finally used to form biomass and metabolite. The metabolites of F6P, T3P, 3PG, pyruvate (pyr), Ac-CoA, X5P, R5P and RuBP represent fructose 6-phosphate, propylphosphonic anhydride, 3-
phosphoglyceric acid, pyruvate acid, acetyl-CoA, xylulose 5-phosphate, ribose 5- phosphate and ribulose bisphosphate, respectively. Protein of PRK and RuBisCo was the abbreviation of phosphoribulokinase and ribulose-1,5-bisphosphate
carboxylase/oxygenase. (B) Mass balance model in our study. The input carbon sources contain the sole organic sugar of xylose and inorganic carbon from the CO2. The carbon will be converted to the biomass, metabolites and CO2.
Figure 2 HPLC spectrum of the medium as well as BL21(DE3), 2P and 3P strain
cultured in the CBD with 5% CO
2bubbling.
Table 3 Element analysis of the carbon content in E. coli.
Elements
Percentage (%)
Parallel samples Average
Nitrogen (N) 12.64 12.81 13.19 13.14 12.95
Carbon (C) 48.74 47.59 48.19 47.67 48.05
Hydrogen (H) 7.85 7.216 7.179 7.11 7.34
4.2 Effect of bioreactors on the CO
2assimilation
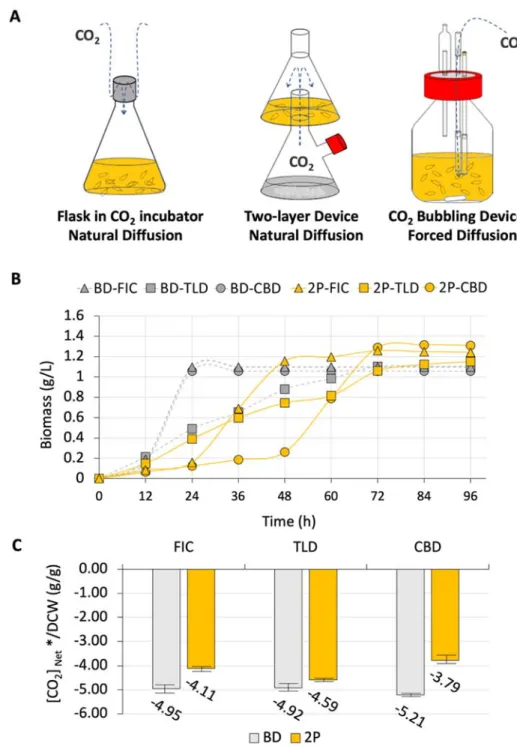
Device effect is one of the determinative factors for developing the cell factory.
Therefore, as shown in Figure 3A, we have designed three different bioreactors (i.e., FIC, TLD and CBD) to culture the RuBisCo-equipped E. coli. In FIC, the cell
obtained CO2 from the incubator by natural diffusion, while, CO2 was generated from the bicarbonate with approximately 1% CO2 and diffused to cell based on the natural diffusion in TLD (Lin et al., 2018). The third one is the CO2 bubbling device (CBD) with a direct injection of CO2 into the culture, indicating a forced diffusion of CO2. We reckoned that different approaches to provide the CO2 to the cell would lead to altered CO2 assimilation.
As shown in Figure 3B, the cell growth between three devices varied each other.
BL21(DE3) showed a constant growth rate without an obvious lag-phase in the TLD, while containing a 12 h-lag phase and faster growth rate in CBD and FIC. As
comparing the cell growth of strain 2P in three devices, CBD and FIC culturing conferred a more extended lag phase of 48 h and 24 h, respectively, while an
undetectable lag phase in the TLD culturing was observed. For the cell growth in the exponential phase, the strain 2P grew faster in CBD and FIC with a similar growth
rate, but possessed a slower growth rate in TLD which resembled to the results of BL21(DE3). Obviously, as the strain 2P was used, it grew slower than the BL21(DE3) but possessed higher biomass at 96 h, indicating a functional RuBisCo-based pathway was equipped in the E. coli. Therefore, the calculation described in the Equation (4) was performed. Considering a fair comparison, the [CO2]net* was normalized by the biomass, forming a new parameter of specific CO2 assimilation capability,
[CO2]net*/DCW. As shown in Figure 3C, the specific CO2 assimilation capability of BL21(DE3) was approximately -5 g-CO2/g-DCW and lower than that of stain 2P, indicating additional CO2 was assimilated into the metabolism. Furthermore, the order of specific CO2 assimilation capability in terms of “g-CO2/g-DCW” was followed by TLD < FIC < CBD, with the exact value of -4.59, -4.11 and -3.79 for TLD, FIC and CBD, respectively.
Figure 3 The effect of bioreactoron the capability of CO
2assimilation.
(A) Scheme of the selected device. FIC, TLD and CBD were abbreviation of flask in CO2 incubation, two-layered device and CO2 bubbling device, respectively. (B) Cell growth curve of the BL21(DE3) and strain 2P in three different devices. Circle, rectangle and triangle respectively represent the culturing in CBD, TLD and FIC. The gray and yellow symbol represent the BL21(DE3) and strain 2P. (C) Calculation
specific [CO2]net* value. The gray and orange color indicated the value calculated from the data from BL21(DE3) and strain 2P, respectively. The error bar represents the standard deviation from three parallel samples.
4.3 Optimization of CO
2utilization in RuBisCo-equipped E. coli
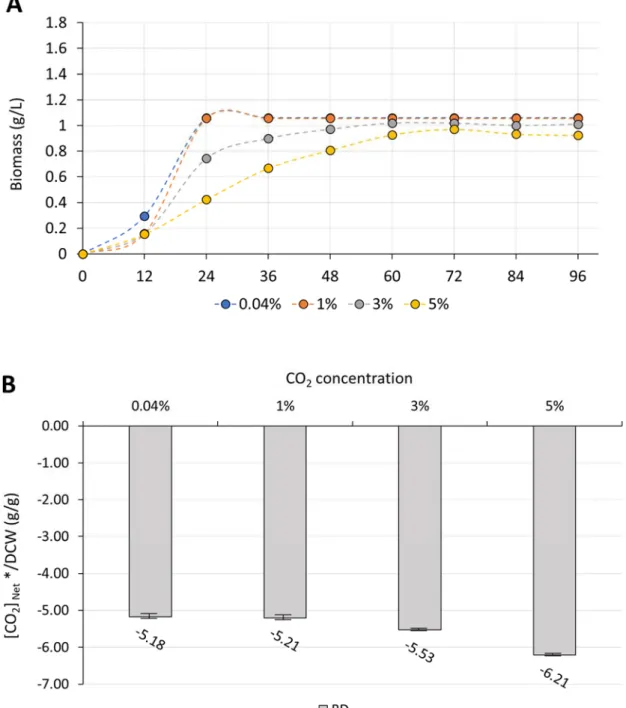
The better bioreactor is determined to be the CBD due to higher specific CO2assimilation capability. We wondered whether the CO2 concentration also affected the CO2 assimilation ability due to increased precursor supply. Therefore, we tested the CO2 concentration of 0.04% (i.e., air), 3% and 5% in CBD. The results showed that the trend of cell growth of BL21(DE3) under different CO2 concentration is similar, but increased CO2 concentration would inhibit the cell growth rate and final biomass.
The higher the CO2 concentration injected, the more serious in inhibition occurred (Figure 4A). Likewise, strain 2P possessed a lag phase of 36 h under the 0.04% CO2
bubbling, while grew with a longer lag phase of 48 h under 1%, 3% and 5% CO2
bubbling. However, the higher the CO2 concentration injected, the higher the final biomass formed in the stationary phase that 1.6 g/L in 5%, 1.5 g/L in 3%, 1.3 g/L in 1% CO2 (Figure 5A). The calculation of specific CO2 assimilation capability was shown in the Figure 4B and Figure 5B. Specific CO2 assimilation capability of BL21(DE3) decreased from -5.18 to -6.21 (Figure 4B), while that of strain 2P increased from -5.03 to -3.79, -2.92 and -2.63 (Figure 5B) as the CO2 concentration increased from 0.04%, 1%, 3% to 5%, respectively. As a result, it is consistence with that the RuBisCo would be activated by higher CO2 concentration (Bracher et al.,
2017). The phenomenon of the reduction in BL21(DE3) was attributed to that
exposure of CO2 to E. coli led to the cell stress and higher CO2 would damage the cell due to the inability to convert the excess CO2 (Debs-Louka et al., 1999).
Figure 4 Effect of the CO
2concentration on the capability of CO
2assimilation in BD strain.
(A) The cell growth of strain BD under 0.04% to 5% CO2 bubbling in CBD (B) The calculation of specific CO2 assimilation capability. The gray bar indicated the value calculated from the data strain 2P. The error bar represents the standard deviation from three parallel samples.
Figure 5 Effect of the CO
2concentration on the capability of CO
2assimilation in 2P strain.
(A) The cell growth of strain 2P under 0.04% to 5% CO2 bubbling in CBD (B) The calculation of specific CO2 assimilation capability. The orange bar indicated the value calculated from the data strain 2P. The error bar represents the standard deviation from three parallel samples.
4.4 Enhance the CO
2assimilation by inducer and chaperone
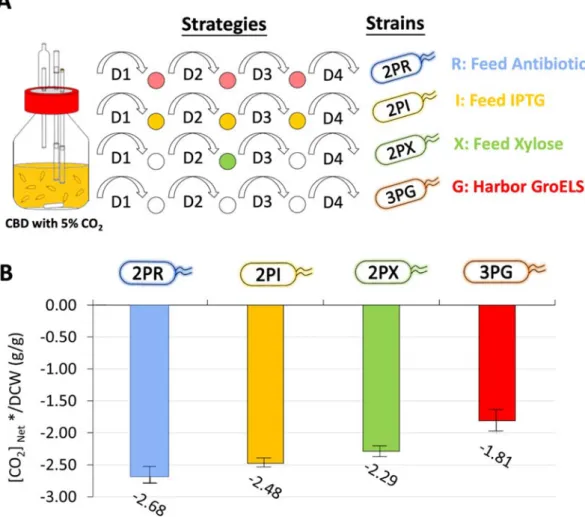
Since the CO2 assimilation performance may be altered by fermentation strategy or co-expressing chaperone to assist the folding of key proteins. Four strategies were designed as shown in Figure 6A. As our culturing maintained for four days, the resistance and IPTG was considered to decay, thus regular addition of appropriate antibiotic (i.e., R sample), or 0.05 mM IPTG (i.e., I sample) was performed.
Furthermore, the RuBisCo reaction depends on the availability of precursor; hence, we fed 2 g/L xylose at 2nd day culturing (i.e., X sample). The latest strategy involves the usage of strain 3P which overexpressed the GroELS chaperone (i.e., G sample) because RuBisCo has been reported requiring different auxiliary proteins and chaperones to perform the complete function (Bracher et al., 2017; Liu et al., 2010).
As shown in Figure 6B, the addition of resistance did not enhance the CO2
assimilation, indicating the plasmids were kept. The addition of IPTG slightly increased [CO2]net*/DCW from -2.63 to -2.48, implying that the IPTG would be decayed during the long-term culturing. In strain 2P, [CO2]net*/DCW could be more efficiently enhanced to -2.29 by feeding the xylose, due to enough supply of the precursor of the RuBisCo-based pathway, R5P. The final approach was introducing the GroELS chaperone (i.e., strain 3P) which was first tested in different bioreactors.
The results showed a similar tendency of cell growth but higher biomass as compared to strain 2P (Figure 7A). Besides, the highest [CO2]net*/DCW was calculated to be - 3.23 gCO2/gDCW in the CBD with 1% CO2 bubbling (Figure 7B), which is higher than -3.79 gCO2/gDCW obtained from strain 2P that cultured at the same condition (Figure 3C, bar of CBD-2P). The strain 3P demonstrated the specific CO2 assimilation capability up to -1.81 gCO2/gDCW in the CBD with 5% CO2 bubbling.
RuBisCo pathway has been incorporated into the E. coli to assimilate the CO2 and showed different efficacy of CO2 assimilation since the different bioreactors, genes and mediums were used (Gleizer et al., 2019; Gong et al., 2015; Li et al., 2015b; Hu
et al., 2018; Zhuang and Li, 2013;), which led to incomparable results. Here, we
considered the device effect on the CO2 assimilation capability in the RuBisCo-equipped E. coli. The quantification could base on the mass balance because a defined medium was used and the summary of specific CO2 assimilation capability in
different condition was listed in the Table 4. Most of the assessment on the CO2
assimilation in the literature was based on the C13 isotope analysis to calculate the amount of assimilated CO2, while it required the complicated procedure for sample preparation and expensive chemicals and instruments for quantification (Gleizer et al.,
2019; Gong et al., 2015). Alternatively, in silico analysis provided a simpler approach
to quantify the ability of CO2 assimilation by comparing the C2/C1 metabolite ratio, but it required the modification on the host (i.e. deletion of frd and ldh) to simplify the metabolites (Yang et al., 2016). On the other hand, the green process that requires the reduction on the CO2 footprint was considered (Kim et al., 2019a). Therefore, the net CO2 assimilation was applied to the assessment instead of the exact amount of assimilated CO2. The calculation results corresponded to the kinetic data that the efficient CO2 attacked to the RuBisCo could activate the activity of carboxylation (Bracher et al., 2017; Flamholz et al., 2019), where the CBD with forced injection and higher CO2 concentration accelerated the attach of CO2 to the RuBisCo, thus leading to higher calculated CO2 assimilation capability (Table 4). Furthermore, it has long been known that the folding of RuBisCo protein is highly dependent on the GroELS chaperone (Bracher et al., 2017; Liu et al., 2010). Hence, the strain 3P could further enhance the performance of CO2 assimilation (Table 4).Figure 6 Different strategies to enhance the performance of CO
2assimilation.
(A) Schematic illustration of different strategies. All the sample was cultured in the CBD with 5% CO2 bubbling. Strain 2P was used in R, I and X sample. Regular feeding of resistance and IPTG after the 1st, 2nd and 3rd -day culturing was performed for R and I sample, respectively. X sample was fed by the 2g/L after 2nd day culturing.
G sample utilized the strain 3P without any feeding. (B) The calculation of specific CO2 assimilation capability in different strategies.
Figure 7 Device effect in 3P strain
(A) cell growth curve, and (B) CO2 assimilation capability at 96 h of 3P strain in FIC, TLD and CBD bioreactor, respectively.
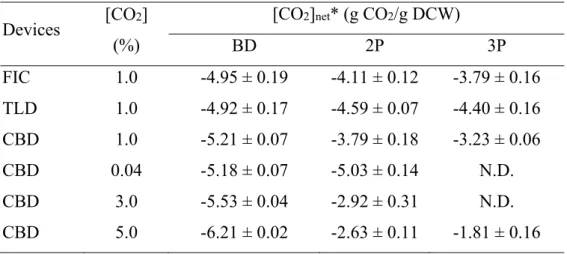
Table 4 Summary of specific CO
2assimilation capability in different conditions
Devices [CO2] (%)
[CO2]net* (g CO2/g DCW)
BD 2P 3P
FIC 1.0 -4.95 ± 0.19 -4.11 ± 0.12 -3.79 ± 0.16 TLD 1.0 -4.92 ± 0.17 -4.59 ± 0.07 -4.40 ± 0.16 CBD 1.0 -5.21 ± 0.07 -3.79 ± 0.18 -3.23 ± 0.06 CBD 0.04 -5.18 ± 0.07 -5.03 ± 0.14 N.D.
CBD 3.0 -5.53 ± 0.04 -2.92 ± 0.31 N.D.
CBD 5.0 -6.21 ± 0.02 -2.63 ± 0.11 -1.81 ± 0.16 N.D. not-determined.
4.5 New insight into the toxicity and evolution in the RuBisCo- equipped E. coli
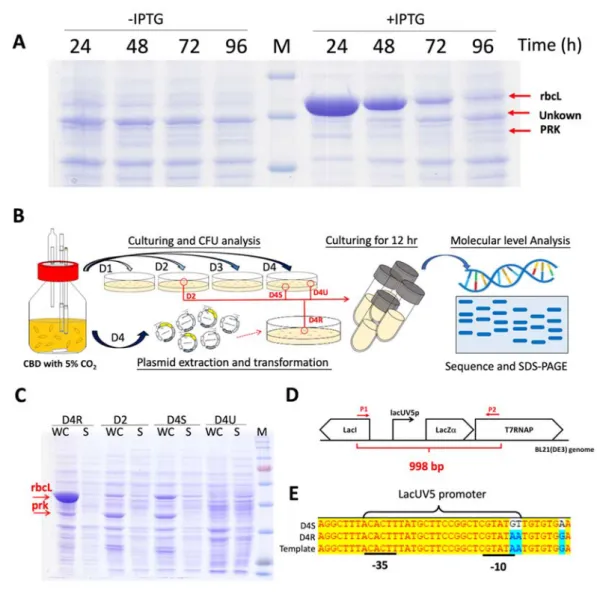
As a serious growth retardation was shown in Figure 3B, indicating the toxicity presented in the RuBisCo-equipped E. coli. We further performed the protein analysis by SDS-PAGE and coupled with tandem MS/MS for protein identification. The results showed that the RuBisCo and PRK protein expression decreased along with the culturing and, meanwhile, an unknown protein band which existed constantly in the non-induction condition recovered (Figure 8A). In order to ensure the reason why the RuBisCo and PRK protein expression decreased and induced the stress, two strategies were performed (Figure 8B). First approach was CFU analysis and 3 colonies were selected, including one from 2nd day culturing (i.e., D2) and two from 4th day culturing (i.e., D4S and D4U). The other approach was extracting the plasmids from the cell of 4th day culturing and then retransformed into BL21(DE3) (i.e., D4R).
The results showed that retransformed one (i.e. D4R) was successfully overexpressed RuBisCo and PRK protein in LB medium, while strains D2 and D4S decreased the expression level of both proteins. Even worst, strain D4U could not express both enzymes (Figure 8C). Due to the only concern of the T7-based expression as shown in
Figure 8D, we sequenced the promoter region of T7 RNAP and found 3 bp mutation,
where an AA to GT mutation occurred on the -35 region of promoter lacUV5 (Figure8E). Actually, the long-term culturing has been reported to trigger the plasmid loss
and inherent mutation due to the toxicity from the plasmid replication or geneexpression (Kang et al., 2018; Tan and Ng; 2020; Tyo et al., 2009; Zhang et al, 2019), while, in our system, there is no significant plasmid loss and inherent mutation (Fig.
6B and 8C). Instead, it seems that the mutation was distributed in the genome
sequence, which was consistent to recent work on detailed genome sequence of ALE-
evolved autotrophic E. coli (Antonovsky et al., 2016; Gleizer et al., 2019; Herz et al.,
2017;).
Figure 8 Molecular level discovery of the toxicity of RuBisCo pathway in E. coli.
(A) SDS-PAGE analysis of CBD-cultured 2P strain with and without IPTG induction.
(B) Schematic illustration of the flowchart to discover the reason of the toxicity observed in the RuBisCo-equipped E. coli. (C) SDS-PAGE analysis of the protein expression from the selected colonies. (D) Schematic illustration of the sequencing region in the genome (E) Sequencing result of D4S and D4R colony.
Next, tandem MS/MS analysis was performed to identify the unknown band and the predicted identities were listed in the Table 5. The selected proteins have been identified as the elongation Tu 2, xylose isomerase and NADH dehydrogenase. The xylose isomerase and NADH dehydrogenase would interfere the xylose metabolism and the energy generation (Berrisford, et al., 2016; Kobayashi et al., 2019), which was attributed to the reason of longer lag phase when two heterologous proteins in the RuBisCo-based pathway were overexpressed. Elongation Tu promotes the ribosome elongation on the mRNA and maintained the translational accuracy (Ogle et al.,
2003), correspondingly one of the reasons for the longer lag-phase. Besides,
elongation Tu has been reported to maintain the rod-shape morphology by co-localizing with the cell-shape determining factor MreB on the cell membrane (Soufo
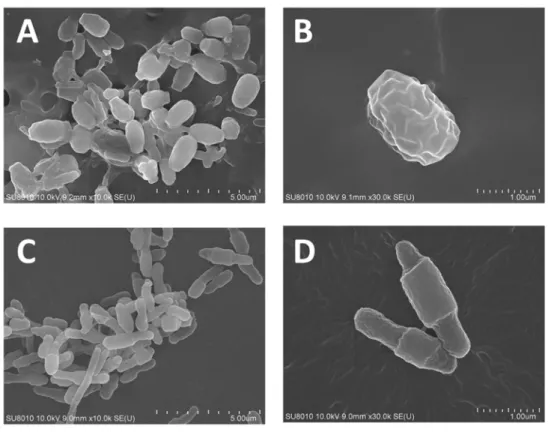
et al., 2010). Therefore, SEM was performed to observe the cell morphology at the 1
st day culturing and 4th day culturing. As shown in Figure 9, the cell became a spherical shape with an average length of 2.5 m and diameter of 1.5 m at the 1st dayculturing (Figure 9A and 9B), corresponding to the results of tandem MS/MS. After further 3-day culturing, the cell recovered the morphology to the rod-shape with an average length of 2 m and diameter of 0.75 m. Intriguing, the cell inherited the cell morphology of 1st day culturing with a thicker circle on the middle of body (Figure
9C and 9D).
Table 5 Mascot protein analysis from LC–MS/MS.
Protein name Accession No. Score Mw
(kDa) pI Coverage
(%)
Elongation factor Tu 2 EFTU2_ECO24 6344 44.3 5.36 89.6 Xylose isomerase XYLA_ECOHS 4721 49.7 5.7 85.9NADH dehydrogenase DHNA_ECOLI 1575 47.3 8.96 75.3
Figure 9 Cell morphology of the CBD-cultured 2P strain by SEM analysis.
The cell was collected at the (A) and (B) at the 1st day, while (C) and (D) at the 4th day culturing.
The phenomenon of the alteration from protein analysis and cell morphology after expression of the RuBisCo and PRK has ascribed to the RNA binding ability from the RuBisCo. In the previous report, the large subunit of RuBisCo has been verified to equip with the RNA binding domain by in vitro and in vivo assay and the domain was used to regulate the RuBisCo expression in Chlamydomonas reinhardtii (Yosef et al.,
2004). Our first molecular level discovery and strain 3P not only revealed the
incidental event that the heterologous protein of RuBisCo interfered the translational process, but also provided a rational way to solve the toxicity of RuBisCo system in E. coli, which paves the way to a robust “green E. coli” by co-expressed chaperone as a cell factory.
4.6 Conclusion
Reengineering the fast-growing E. coli to utilize CO2 is already feasible and has been regarded as a prominent route to the next generation of bioenergy. Our study investigated the first insight into the effect of bioreactor on the CO2 assimilation capability from the aspect of an engineer. Besides, we provide the new mindfulness to evaluate the system in a “green” manner by calculating the net CO2 instead of the exact assimilated CO2. Furthermore, an original perspective of the toxicity was elucidated and paved the way to further enhance RuBisCo-equipped E. coli from the aspect of a scientist.
Chapter 5 References
[1]
Antonovsky, N., Gleizer, S., Noor, E., Zohar, Y., Herz, E., Barenholz, U., Zelcbuch, L., Amram, S., Wides, A., Tepper, N., Davidi, D., Bar-On, Y., Bareia, T., Wernick, D. G., Shani, I., Malitsky, S., Jona, G., Bar-Even, A., Milo, R., 2016. Sugar synthesis from CO
2in Escherichia coli. Cell. 166(1), 115-125.
[2] Berrisford, J. M., Baradaran, R., Sazanov, L. A., 2016. Structure of bacterial respiratory complex I. Biochimi. Biophys. Acta -Bioenerg. 1857(7), 892-901.
[3]
Bracher, A., Whitney, S. M., Hartl, F. U., Hayer-Hartl, M., 2017. Biogenesis and metabolic maintenance of Rubisco. Annu. Rev. Plant Biol. 68, 29-60.
[4]
Bui, M., Adjiman, C. S., Bardow, A., Anthony, E. J., Boston, A., Brown, S., Fennell, P. S., Fuss, S., Galindo, A., Hackett, L. A., Hallett, J. P., Herzog, H. J., Jackson, G., Kemper, J., Krevor, S., Maitland, G. C., Matuszewski, M., Metcalfe, I. S., Petit, C., Puxty, G., Reimer, J., Reiner, D. M., Rubin, E. S., Scott, S. A., Shah, N., Smit, B., Trusler, J. P. M., Webley, P., Wilcoxx, J., Dowell, N. M., 2018. Carbon capture and storage (CCS): the way forward. Energy Environ. Sci.
11(5), 1062-1176.
[5]
Change, I. C., 2014. Synthesis Report. Contribution of working groups I. II and
III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, 151(10.1017).[6] Cheah, W. Y., Ling, T. C., Juan, J. C., Lee, D. J., Chang, J. S., Show, P. L., 2016. Biorefineries of carbon dioxide: from carbon capture and storage (CCS) to bioenergies production. Bioresour. Technol. 215, 346-356.
[7]
Choi, K. R., Jang, W. D., Yang, D., Cho, J. S., Park, D., Lee, S. Y., 2019.
Systems metabolic engineering strategies: integrating systems and synthetic
biology with metabolic engineering. Trends Biotechnol. 37(8), 817-837.
[8]
d Mattozzi, M., Ziesack, M., Voges, M. J., Silver, P. A., Way, J. C., 2013.
Expression of the sub-pathways of the Chloroflexus aurantiacus 3-
hydroxypropionate carbon fixation bicycle in E. coli: Toward horizontal transfer of autotrophic growth. Metab. Eng. 16, 130-139.
[9] Debs-Louka, E., Louka, N., Abraham, G., Chabot, V., Allaf, K., 1999. Effect of compressed carbon dioxide on microbial cell viability. Appl. Environ. Microbiol.
65(2), 626-631.
[10] Flamholz, A. I., Prywes, N., Moran, U., Davidi, D., Bar-On, Y. M., Oltrogge, L.
M., Alves, R., Savage, D., Milo, R., 2019. Revisiting trade-offs between Rubisco kinetic parameters. Biochem. 58(31), 3365-3376.
[11] Folsom, J. P., Carlson, R. P., 2015. Physiological, biomass elemental composition and proteomic analyses of Escherichia coli ammonium-limited chemostat growth, and comparison with iron-and glucose-limited chemostat growth. Microbiol. 161(Pt 8), 1659.
[12]
Gleizer, S., Ben-Nissan, R., Bar-On, Y. M., Antonovsky, N., Noor, E., Zohar, Y., Jona, G., Krieger, E., Shamshoum, M., Bar-Even, A., Milo, R., 2019. Conversion of Escherichia coli to generate all biomass carbon from CO
2. Cell. 179(6), 1255- 1263.
[13]
Goeppert, A., Czaun, M., Prakash, G. S., Olah, G. A., 2012. Air as the renewable carbon source of the future: an overview of CO
2capture from the atmosphere.
Energy Environ. Sci. 5(7), 7833-7853.
[14]
Gong, F., Liu, G., Zhai, X., Zhou, J., Cai, Z., Li, Y., 2015. Quantitative analysis of an engineered CO
2-fixing Escherichia coli reveals great potential of
heterotrophic CO
2fixation. Biotechnol. Biofuels. 8(1), 86.
[15] Guadalupe-Medina, V., Wisselink, H. W., Luttik, M. A., de Hulster, E., Daran, J.
M., Pronk, J. T., van Maris, A. J., 2013. Carbon dioxide fixation by Calvin-Cycle enzymes improves ethanol yield in yeast. Biotechnol. Biofuels. 6(1), 125.
[16]
He, Y. C., Jiang, C. X., Chong, G. G., Di, J. H., Ma, C. L., 2018. Biological synthesis of 2, 5-bis (hydroxymethyl) furan from biomass-derived 5-
hydroxymethylfurfural by E. coli CCZU-K14 whole cells. Bioresour. Technol.
247, 1215-1220.
[17]
Herz, E., Antonovsky, N., Bar-On, Y., Davidi, D., Gleizer, S., Prywes, N., Noda-Garcia, L., Lyn Frisch, K., Zohar, Y., Wernick, D. G., Savidor, A., Barenholz, U., & Milo, R., 2017. The genetic basis for the adaptation of E. coli to sugar synthesis from CO
2. Nat. Commun. 8(1), 1-10.
[18]
Hu, D., Li, M., Zhou, R., Sun, Y., 2012. Design and optimization of photo bioreactor for O
2regulation and control by system dynamics and computer simulation. Bioresour. Technol. 104, 608-615.
[19]
Hu, G., Zhou, J., Chen, X., Qian, Y., Gao, C., Guo, L., Xu, P., Chen, W., Chen, J., Li, Y., Liu, L., 2018. Engineering synergetic CO
2-fixing pathways for malate production. Metab. Eng. 47, 496-504.
[20]
Kang, C. W., Lim, H. G., Yang, J., Noh, M. H., Seo, S. W., Jung, G. Y., 2018.
Synthetic auxotrophs for stable and tunable maintenance of plasmid copy number. Metab. Eng. 48, 121-128.
[21] Kim, G. Y., Roh, K., Han, J. I., 2019a. The use of bicarbonate for microalgae cultivation and its carbon footprint analysis. Green Chem. 21(18), 5053-5062.
[22]
Kim, S. J., Yoon, J., Im, D. K., Kim, Y. H., Oh, M. K., 2019b. Adaptively
evolved Escherichia coli for improved ability of formate utilization as a carbon
source in sugar-free conditions. Biotechnol. Biofuels. 12(1), 207.
[23]
Kim, S., Lindner, S. N., Aslan, S., Yishai, O., Wenk, S., Schann, K., Bar-Even, A., 2020. Growth of E. coli on formate and methanol via the reductive glycine pathway. Nat. Chem. Biol. 16(5), 538-545.
[24]
Kobayashi, J., Sasaki, D., Bamba, T., Hasunuma, T., Kondo, A., 2019.
Sustainable production of glutathione from lignocellulose-derived sugars using engineered Saccharomyces cerevisiae. Appl. Microb Biotechnol. 103(3), 1243- 1254.
[25]
Lai, Y. C., Chang, C. H., Chen, C. Y., Chang, J. S., Ng, I. S., 2019. Towards protein production and application by using Chlorella species as circular economy. Bioresour. Technol. 289, 121625.
[26]
Levasseur, W., Perré, P., Pozzobon, V., 2020. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol.
Adv. 107545. In press.
[27]
Li, D., Wang, L., Zhao, Q., Wei, W., Sun, Y., 2015a. Improving high carbon dioxide tolerance and carbon dioxide fixation capability of Chlorella sp. by adaptive laboratory evolution. Bioresour. Technol. 185, 269-275.
[28]
Li, Y. H., Ou-Yang, F. Y., Yang, C. H., Li, S. Y., 2015b. The coupling of glycolysis and the Rubisco-based pathway through the non-oxidative pentose phosphate pathway to achieve low carbon dioxide emission fermentation.
Bioresou. Technol. 187, 189-197.
[29]
Liang, S., Altaf, N., Huang, L., Gao, Y., Wang, Q., 2020. Electrolytic cell design for electrochemical CO
2reduction. J. CO
2 Util. 35, 90-105.[30]
Lin, L. C., Berger, A. H., Martin, R. L., Kim, J., Swisher, J. A., Jariwala, K.,
Rycroft, C. H., Bhown, A. S., Deem, M. W., Haranczyk, M., Smit, B. 2012. In
silico screening of carbon-capture materials. Nat. Mater. 11(7), 633.
[31]
Lin, W. R., Lai, Y. C., Sung, P. K., Tan, S. I., Chang, C. H., Chen, C. Y., Chang, J. S., Ng, I. S., 2018. Enhancing carbon capture and lipid accumulation by genetic carbonic anhydrase in microalgae. J. Taiwan Inst. Chem. Eng. 93, 131- 141.
[32] Lin, W. R., Tan, S. I., Hsiang, C. C., Sung, P. K., Ng, I. S., 2019. Challenges and opportunity of recent genome editing and multi-omics in cyanobacteria and microalgae for biorefinery. Bioresour. Technol. 291, 121932.
[33]
Liu, C., Young, A. L., Starling-Windhof, A., Bracher, A., Saschenbrecker, S., Rao, B. V., Berninghausen, O., Mielke, T., Hartl, F. U., Beckmann, R., Hayer- Hartl, M., 2010. Coupled chaperone action in folding and assembly of
hexadecameric Rubisco. Nature. 463(7278), 197-202.
[34]
Luo, Z. W., Cho, J. S., Lee, S. Y., 2019. Microbial production of methyl anthranilate, a grape flavor compound. Proc. Nat. Acad. Sci. 116(22), 10749- 10756.
[35] Mikkelsen, M., Jørgensen, M., Krebs, F. C., 2010. The teraton challenge. A review of fixation and transformation of carbon dioxide. Energy Environ. Sci.
3(1), 43-81.
[36]
Mustafa, A., Lougou, B. G., Shuai, Y., Wang, Z., Tan, H., 2020. Current technology development for CO
2utilization into solar fuels and chemicals: A review. J. Energy Chem. 49, 96-123.
[37]
Ogle, J. M., Carter, A. P., Ramakrishnan, V., 2003. Insights into the decoding mechanism from recent ribosome structures. Trends Biochem. Sci. 28(5), 259- 266.
[38]
Salehizadeh, H., Yan, N., Farnood, R., 2020. Recent advances in microbial CO
2fixation and conversion to value-added products. Chem. Eng. J. 390, 124584.
[39]
Schada von Borzyskowski, L., Carrillo, M., Leupold, S., Glatter, T., Kiefer, P., Weishaupt, R., Heinemann, M., Erb, T. J., 2018. An engineered Calvin-Benson- Bassham cycle for carbon dioxide fixation in Methylobacterium extorquens AM1. Metab. Eng. 47, 423-433.
[40]
Show, K. Y., Lee, D. J., Chang, J. S., 2011. Bioreactor and process design for biohydrogen production. Bioresour. Technol., 102(18), 8524-8533.
[41]
Soufo, H. J. D., Reimold, C., Linne, U., Knust, T., Gescher, J., Graumann, P. L., 2010. Bacterial translation elongation factor EF-Tu interacts and colocalizes with actin-like MreB protein. Proc. Nat. Acad. Sci. 107(7), 3163-3168.
[42]
Stocker, T. F., Qin, D., Plattner, G.. K., Tignor, M. M. B., Allen, S. K., Buschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P. M., 2013. Climate change 2013: The Physical Science Basis.
[43]
Tan, S. I., Ng, I. S., 2020. New insight into plasmid-driven T7 RNA polymerase in Escherichia coli and use as a genetic amplifier for a biosensor. ACS Synth.
Biol. 9(3), 613-622.
[44] Thakur, I. S., Kumar, M., Varjani, S. J., Wu, Y., Gnansounou, E., Ravindran, S., 2018. Sequestration and utilization of carbon dioxide by chemical and biological methods for biofuels and biomaterials by chemoautotrophs: opportunities and challenges. Bioresour. Technol. 256, 478-490.
[45] Tubb, R. (2015). EIA: Annual energy outlook through 2040. Pipeline & Gas Journal, 1.
[46]
Tyo, K. E., Ajikumar, P. K., Stephanopoulos, G., 2009. Stabilized gene
duplication enables long-term selection-free heterologous pathway expression.
Nat. Biotechnol. 27(8), 760.