國立宜蘭大學生物技術研究所 碩士論文
Institute of Biotechnology National Ilan University
Master Thesis
以蛋白質體學方法分析腸炎弧菌對抗菌胜肽 之抗性機制
Proteomic analysis on the antimicrobial peptide- resistance mechanism of Vibrio parahaemolyticus
指導教授:陳威戎 博士 Wei-Jung Chen, Ph. D.
研究生:沈春榮 Chun-Jung Shen
中華民國九十七年七月
謝 誌
兩年的碩士班研究生生涯終於劃上句號了,回想這一路走來,要感謝 的人太多了,首要感謝指導老師 陳威戎老師在我剛邁進研究所的時候,願 意收留及指導我這個毫無蛋白質體學背景,完全從零出發的學生,給予我 許多的幫助、鼓勵與機會,在我實驗結果不如預期的時候,總是能從不同 的角度提出結果並非失敗的觀點。在我實驗遇到瓶頸的時候,總是能一面 幫我加油打氣一面修正實驗方法。提供許多難得的機會,讓我有管理實驗 室、親手操作質譜儀以及參與許多大型研討會的經驗。不僅關心學生的實 驗結果,也關心學生的作息時間及感情生活,使我與老師之間的相處有如 亦師亦友般的關係。也要感謝許多老師給予我的幫助,感謝林世斌老師及 郭村勇老師提供實驗所需的菌株,感謝台灣大學生化暨分子生物學研究所 暨基因體中心 周綠蘋老師提供 MALDI-TOF MS 以及莊景光學長耐心地教 導我儀器操作的方法及注意事項,感謝台灣大學生化暨分子生物學研究所 張智芬老師及謝青悅助教利用LC-tandem MS 幫我分析蛋白質點樣品。
感謝實驗室的成員們陪我度過兩年的實驗室生活,實驗室的生活總是 煩悶無聊的,幸好有你們陪我一塊說說笑笑,互相加油打氣。我認為我們 實驗室的特色就是氣氛輕鬆但不放鬆,做實驗方便但不隨便,而這樣的特 色全都依賴實驗室的所有成員們聚在一起而形成的。親和力十足的可婷師 母,在我擔任室長的時候,感謝妳給我的鼓勵與支持。大魔頭周宏達,總 是能夠準時地招集大夥一塊去吃飯。經常語出驚人的高高,感謝你傳授萃 取外膜蛋白的實驗心得。善解人意的筱倩,多虧妳好心地幫我蒐集資料,
讓我能夠專心看期刊,有如我的私人秘書一般。個性內斂的笑傑,時常提 供許多實用又方便的軟體。帥氣又和善的閔元,與你討論實驗讓我獲益良 多,經常點醒我許多沒注意到的細節。敢於追求的阿毛,當我不小心把自 己反鎖在宿舍門外時,感謝你不辭辛勞地載我去礁溪取得備鑰,讓我免於 露宿街頭的命運。名字比較帥的伊翔,總是不厭其煩地幫我下載期刊,讓 我能夠充實論文的內容。求知慾旺盛的貝貝、天真單純的若瑜、任勞任怨 的亞嫺,與妳們相處十分融洽,感謝妳們讓我能夠快速地融入實驗室生活。
抗壓性強的怡苓,幸好有妳陪我一塊度過那段有如無間地獄的瘋狂MIC 的 日子,妳真的是我最得力的幫手。心地善良且熱心助人的小斑,感謝妳在 我需要幫助的時候對我伸出援手,也感謝妳經常提供好吃又美味的吳媽媽 油飯。活潑開朗的育袖,妳的朝氣能夠讓實驗室充滿活力。不計形象極力 搞笑的妍妍,只要有妳在,實驗室的笑聲就不會間斷。個性率直的惠君、
氣質出眾的阿惪,感謝妳們經常提供各地名產,養肥了我的肚子。理解力 佳的邱衍、認真踏實的立蓁、熱情可愛的小甜甜、溫文儒雅的小盧子以及 神秘低調的非凡,感謝你們經常主動地幫忙插 tip、滅菌、清理垃圾以及清 洗被我堆積在水槽中的瓶瓶罐罐,你們的熱心讓實驗室擁有良好的環境。
實驗室的新進人員,大龍、維真、俊宏、薇如、孟修、志宏、綉琪,雖然 與你們相處只有短短一個月,不過在這段時間內和你們相處融洽,歡迎你 們加入 PSF 這個大家庭。接著還要感謝與我相處兩年的同窗好友們,感謝 彥均、小宮及小梁不吝分享你們的實驗心得,我會懷念我們每個星期固定 的午餐約會。感謝瑾玟熱心地與我分享抽取RNA 的實驗心得,讓我能夠免 去嘗試錯誤的階段而如期完成實驗。感謝讀書會的固定班底,卓臻、楹涵、
漢婷及筱婷,雖然過程中經常發生唇槍舌戰或不知不覺就聊起八卦,但是 與你們一塊讀書真的很有效率。
當然不能忘記一直在我身後支持我的家人及朋友,從我小的時候,爸 爸就以身作則地教導我學習如何當個有責任感、做事積極的人。當我離家 在外時,媽媽總是擔心我在外地的生活,讓我察覺到不管多大的孩子都少 不了媽媽的關愛及照顧。大妹妍伶總是適時地協助媽媽提供我經濟援助。
不管我離家多久,小妹珈毓總是會跟著爸爸一塊來接我回家。感謝你們長 久以來給予我的支持與鼓勵,讓我能夠心無旁鶩地取得碩士學位。也要感 謝與我相處十多年的死黨們,個子高大但不擅打籃球的弼凱、認真負責而 且和我一塊打電動會吵架的麵包以及從小玩到大的小白,感謝你們在我犯 錯時,指責我的不對,分享我的快樂,聆聽我的抱怨,與我一塊同仇敵愾,
你們是最講義氣的兄弟。
摘 要
腸炎弧菌是一株生長於入海口處的嗜鹽性革蘭氏陰性菌,同時也是造 成魚、蝦及貝類產生弧菌病的常見海洋病原菌。另外,腸炎弧菌也能夠藉 由人類攝食遭受腸炎弧菌汙染的未經烹煮之海鮮食物而導致人類產生細菌 性胃腸炎。由於抗生素具有有效殺菌的能力,因此人類廣泛地將抗生素使 用於治療細菌所引起的疾病。然而,細菌能夠藉由發展各種抗性機制而達 到抵抗抗生素的效果,使得我們必需尋找及發展能夠取代抗生素的治療 劑。抗菌胜肽是一種廣泛存在於自然界所有生物體中的短鍊胜肽,抗菌胜 肽主要是藉由破壞細菌細胞膜的方式達到殺菌的效果,因此,抗菌胜肽具 有取代抗生素的潛力。
在本研究論文中,藉由測定各種抗菌胜肽對腸炎弧菌的最小抑菌濃度 (minimal inhibitory concentrations;MICs),發現3種實驗室設計開發的抗菌 胜肽 (Q4、Q6及H1) 及天然存在的抗菌胜肽 (Pleurocidin) 具有有效抑制腸 炎弧菌生長的能力。藉由添加最小抑菌濃度一半的劑量作為起始濃度,並 隨著每次繼代增加2倍抗菌胜肽濃度的方式,嘗試誘導腸炎弧菌對抗菌胜肽 產生抗性,其結果顯示3種實驗室設計開發的抗菌胜肽確實不容易被腸炎弧 菌所抵抗。接著比較野生型腸炎弧菌與抗性菌株之間的膜蛋白表現量之差 異,並藉由質譜儀分析及生物資訊學進行蛋白質身份的鑑定。其結果顯示 外膜蛋白TolC、鞭毛蛋白、轉錄延長因子NusA、長鏈脂肪酸運送蛋白、轉 譯延長因子Tu、F1-ATP合成酶的α-次單位及二氫硫辛醯胺去氫酶等蛋白質 在抗性菌株中的表現量有增加的趨勢,經由即時定量聚合酶連鎖反應測定 上述蛋白質之mRNA在野生型腸炎弧菌與抗性菌株之間的表現量之後,我們 認為腸炎弧菌可能是利用多重抗藥性排出幫浦將菌體內的抗菌胜肽排出,
以及藉由增加膜的合成達到修復受損的細胞膜等方式,達到抵抗抗菌胜肽 的效果。在這場抗菌胜肽與病原菌共同演化的軍備競賽中,我們欲佔得先 機,唯有深入探究其可能之抗性機制,並加以反制。本碩士論文的發現提 供了一些初步的想法,我們仍將持續深入研究,以期知己知彼,百戰百勝。
Abstract
Vibrio parahaemolyticus is a halophilic gram-negative bacterium that occurs naturally in estuarine environments world-wide. V. parahaemolyticus is also a universal marine pathogen causing vibriosis in fish, shrimp and shellfish.
Consumption of uncooked seafood contaminated with a virulent strain of V.
parahaemolyticus can cause bacterial gastroenteritis of human. The extensive
use of classical antibiotics has led to the growing emergence of many resistant strains of pathogenic microorganisms. Therefore, the development of novel therapeutic agents that could overcome the resistance problem has become critical. Antimicrobial peptides (AMPs) is widespread present in all kinds of organisms. AMPs are of greatest potential to represent such a new class of antibiotics. In the present study, we measured the minimal inhibitory concentrations (MICs) of several AMPs against V. parahaemolyticus. Three synthetic AMPs (Q4, Q6 and H1) and a natural one from fish (Pleurocidin) have best antibacterial activity and selectivity. Incubation with serially increased concentrations of AMPs starting from MIC/2 during bacterial culture passages were performed for selection of resistant clones. We compared the differential expression patterns of the outer- and inner- membrane proteins (OMPs and IMPs) among wild-type and the resistant strains by two-dimensional gel electrophoresis (2DE). Protein spots significantly altered were then subjected to mass spectrometry analysis and protein identification using Mascot program.Two OMPs (TolC and flagellin) and Five IMPs (transcription elongation factor NusA, long-chain fatty acid transport protein, translation elongation factor Tu, α-subunit of F1-ATP synthase and dihydrolipoic dehydrogenase) were identified.
Real-time quantitative polymerase chain reaction (RT-qPCR) were also performed to determine the mRNA level of the above proteins. In conclusion, V.
parahaemolyticus may obtain their resistance against AMPs through multidrug
efflux transporter and repair of damaged cell mambrane by membrane biosynthesis.目 錄
謝誌‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅰ 中文摘要‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅲ 英文摘要 (Abstract)‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅳ 目錄 (Table of contents)‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅴ 表目錄 (List of Tables)‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅶ 圖目錄 (List of Figures)‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅷ 縮寫表 (Abbreviations)‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ Ⅹ 第一章 前言‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 1 一、腸炎弧菌‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 1 二、抗生素‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 2 三、抗藥性‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 5 四、抗菌胜肽‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 9 五、抗菌胜肽作用機制‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 10 六、細菌抵抗抗菌胜肽的機制‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 12 七、細菌的偵測系統‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 15 八、宿主-病原菌交互作用‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 18 九、蛋白質體學‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 19 第二章 實驗目的‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 21 第三章 實驗材料與儀器‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 22 實驗材料‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 22 實驗儀器‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 25 第四章 實驗方法‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 28 一、菌種培養‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 28 二、最小抑菌濃度‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 28 三、篩選抗性菌株‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 29 四、蛋白質萃取與分劃‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 30 五、SIF 及 SSF 的蛋白質萃取‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 30 六、蛋白質濃度定量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 31 七、二維凝膠電泳‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 32 八、膠體內水解‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 33
九、MALDI-TOF MS 分析‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 34 十、LC-ESI-Q-TOF-MS/MS 分析‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 35 十一、引子設計‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 36 十二、RNA 萃取‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 37 十三、RT-PCR‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 38 十四、RT-qPCR‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 39 第五章 結果‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 41 一、抗菌胜肽或抗生素對腸炎弧菌的最小抑菌濃度‧‧‧‧‧‧‧‧ 41 二、抗性菌株的篩選‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 42 三、腸炎弧菌膜蛋白的萃取及二維凝膠電泳分析結果‧‧‧‧‧‧‧ 43 四、MALDI-TOF MS 及 LC-ESI-MS/MS 的比對結果‧‧‧‧‧‧‧ 46 五、引子的設計與分析‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 47 六、即時定量聚合酶連鎖反應的結果‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 49 第六章 討論‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 51 一、抗菌胜肽的抗菌效果‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 51 二、抗性菌株對抗菌胜肽的抗性‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 51 三、萃取膜蛋白的限制‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 52 四、外膜蛋白TolC‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 53 五、鞭毛蛋白‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 54 六、轉錄延長因子NusA‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 55 七、長鏈脂肪酸運送蛋白‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 56 八、轉譯延長因子Tu‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 57 九、F1-ATP 合成酶的 α-次單位‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 57 十、二氫硫辛醯胺去氫酶‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 58 十一、腸炎弧菌抵抗抗菌胜肽的可能機制‧‧‧‧‧‧‧‧‧‧‧‧ 59 第七章 未來展望‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 61 第八章 圖表‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 62 第九章 參考文獻‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 91
表 目 錄
表4.1 每代菌液中所含有的抗菌胜肽之最終濃度‧‧‧‧‧‧‧‧ 29 表4.2 本研究論文中所使用的引子之序列、長度、GC 比率、熔點及
產物大小‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 37 表5.1 本論文研究所使用的抗菌胜肽之結構參數、分子量及最小抑
菌濃度‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 62 表5.2 有效抑制腸炎弧菌生長的抗菌胜肽或抗生素之最小抑菌濃度 63 表5.3 利用不同抗菌胜肽所挑選出來的可能抗性菌株‧‧‧‧‧‧ 64 表5.4 腸炎弧菌抗性菌株的最小抑菌濃度增加倍數‧‧‧‧‧‧‧ 65 表5.5 外膜及內膜蛋白質的鑑定結果‧‧‧‧‧‧‧‧‧‧‧‧‧ 66 表5.6 RT-qPCR 分析野生型腸炎弧菌與抗性菌株中標的蛋白之
mRNA 含量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 67
圖 目 錄
圖1.1 弧菌屬細菌與其特定宿主‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 1 圖1.2 抗生素的作用機制‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 4 圖1.3 細菌獲得外來抗藥性基因的途徑‧‧‧‧‧‧‧‧‧‧‧‧ 6 圖1.4 細菌抵抗抗生素的機制‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 8 圖1.5 抗菌胜肽造成細菌細胞膜損害的機制‧‧‧‧‧‧‧‧‧‧ 11 圖1.6 抗菌胜肽作用的機制‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 11 圖1.7 抗菌胜肽的作用機制與細菌發展的抗性機制‧‧‧‧‧‧‧ 13 圖1.8 細菌減少或中和表面負電荷的機制‧‧‧‧‧‧‧‧‧‧‧ 14 圖1.9 大腸桿菌的 σE活化路徑‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 16 圖1.10 大腸桿菌的 Cpx 系統之路徑‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 17 圖1.11 抗菌胜肽與細菌抵抗抗菌胜肽之間的共演化‧‧‧‧‧‧‧ 19 圖5.1 抗性菌株篩選實驗的最高代數之菌落生長情形‧‧‧‧‧‧ 68 圖5.2 野生型腸炎弧菌與抗性菌株 H1-Ⅶ-2 及 Ple-Ⅷ-2 的外膜蛋白
二維電泳圖‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 69 圖5.3 野生型腸炎弧菌與抗性菌株 H1-Ⅶ-2 及 Ple-Ⅷ-2 的內膜蛋白
二維電泳圖‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 70 圖5.4 野生型腸炎弧菌及 7 株抗性菌株的外膜蛋白二維電泳圖‧‧ 71 圖5.5 野生型腸炎弧菌及 7 株抗性菌株的內膜蛋白二維電泳圖‧‧ 72 圖5.6 在野生型腸炎弧菌與抗性菌株之間表現量有差異的外膜蛋白
質點‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 73 圖5.7 在野生型腸炎弧菌與抗性菌株之間表現量有差異的內膜蛋白
質點‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 74 圖5.8 使用 Mascot 軟體鑑定蛋白質點 O-1 的蛋白質身份‧‧‧‧‧ 76 圖5.9 使用 Mascot 軟體鑑定蛋白質點 O-2 的蛋白質身份‧‧‧‧‧ 77 圖5.10 使用 Mascot 軟體鑑定蛋白質點 I-1 的蛋白質身份‧‧‧‧‧ 78 圖5.11 使用 Mascot 軟體鑑定蛋白質點 I-2 的蛋白質身份‧‧‧‧‧ 79 圖5.12 使用 Mascot 軟體鑑定蛋白質點 I-3 的蛋白質身份‧‧‧‧‧ 80 圖5.13 使用 Mascot 軟體鑑定蛋白質點 I-4 的蛋白質身份‧‧‧‧‧ 81
圖5.14 使用 Mascot 軟體鑑定蛋白質點 I-5 的蛋白質身份‧‧‧‧‧ 82 圖5.15 RT-qPCR 分析 16S rRNA 在野生型腸炎弧菌和抗性菌株之間
的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 83 圖5.16 RT-qPCR 分析 tolC mRNA 在野生型腸炎弧菌和抗性菌株之間
的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 84 圖5.17 RT-qPCR 分析 fla mRNA 在野生型腸炎弧菌和抗性菌株之間
的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 85 圖5.18 RT-qPCR 分析 nusA mRNA 在野生型腸炎弧菌和抗性菌株之
間的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 86 圖5.19 RT-qPCR 分析 FATP mRNA 在野生型腸炎弧菌和抗性菌株之
間的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 87 圖5.20 RT-qPCR 分析 EF-Tu mRNA 在野生型腸炎弧菌和抗性菌株之
間的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 88 圖5.21 RT-qPCR 分析 atpA mRNA 在野生型腸炎弧菌和抗性菌株之
間的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 89 圖5.22 RT-qPCR 分析 E3 mRNA 在野生型腸炎弧菌和抗性菌株之間
的表現量‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧‧ 90
縮 寫 表
Abbreviation Full text
AMPs Antimicrobial peptides
BPB Bromophenol blue
BSA Bovine serum albumin
CHAPS 3-([ 3-Cholamidopropyl]dimethylammonio)-1-propane sulfonate CHCA α-Cyano-4- hydroxycinnamic acid
Ct Threshold cycle
DTT Dithiothreitol
E3 Dihydrolipoamide dehydrogenase
FATP Fatty acid transport protein
IAA Iodoacetamide
IEF Isoelectric focusing
IMPs Inner membrane proteins
LC-ESI-Q-TOF-MS/MS Liquid chromatography electrospray ionization quadrupole time-of-flight tandem mass spectrometry
M2A Magainin 2a
MALDI-TOF-MS Matrix-assisted laser desorption /ionization time of flight mass spectrometer MICs Minimal inhibitory concentrations
OMPs Outer membrane proteins
Ple Pleurocidin PMF Peptide mass fingerprinting
RT-PCR Reverse transcription- polymerase chain reaction RT-qPCR Real-time quantitative PCR
SDS-PAGE Sodium dodecyl sulfate-polyacrylamide gel electrophoresis SIF Sarcosinate-insoluble fraction SLS Sodium lauryl sarcosinate
SSF Sarcosinate-soluble fraction
TFA Trifluoroacetic acid
Tm Melting temperature
第 一 章 前 言
一、腸炎弧菌 (Vibrio parahaemolyticus)
腸炎弧菌為一種引起細菌性胃腸炎 (bacterial gastroenteritis) 的病原 菌,經常造成海島國家例如台灣、日本、東南亞地區、英國和荷蘭的食物 中毒事件。此菌引起的食物中毒
事 件 最 早 發 生 在1950年日本大 阪,造成272人集體食物中毒,其 中20人死於急性胃腸炎。1953年 日本學者藤野恒三郎 (Fujino T.) 等人從病人排泄物中成功將此菌 分離出來,後來經過多方的研 究,終於在1963年正式命名為腸 炎弧菌 (Pen et al., 1998)。另外,
腸炎弧菌若在含有人類或兔子血 液 的 特 殊 培 養 基 (Wagatsuma agar) 上產生溶血現象,則判定為 溶 血 型 (Kanagawa-positive) 腸 炎弧菌,而這種現象稱為神奈川 現象 (Kanagawa phenolmenon),
反之則為非溶血型 (Kanagawa- negative) 腸炎弧菌,溶血型腸炎 弧 菌 可 分 泌 耐 熱 性 的 溶 血 素 (hemolysin),所以只有溶血型的 腸炎弧菌會導致疾病。在海洋中 大約1%的腸炎弧菌屬於溶血型 的種類,而從病人身上分離出的
腸炎弧菌有96%是屬於會致病的 圖1.1、弧菌屬細菌與其特定宿主。
種類 (Pen et al., 1998)。 (摘自Reen et al., 2006)
腸炎弧菌為嗜鹽性的革蘭氏陰性菌,主要棲息於海岸或海河口之交接 處。腸炎弧菌可以獨自在海水中生存,也可以感染特定宿主,所以會對許 多水生生物造成弧菌病 (vibriosis),例如,魚類、甲殼綱動物、小鮑魚 (俗 稱九孔) ( Reen et al., 2006;圖 1.1),因此嚴重危害水產養殖業。另外,人類 如果誤食遭此菌汙染的海鮮,則會產生劇烈腹痛及下痢,嚴重者將導致死 亡。
二、抗生素 (Antibiotics)
在不同種類的微生物之間,為了互相對抗以求生存,所以製造能夠有 效抑制或殺滅其他微生物的物質,此種方式讓微生物在生存上獲得優勢,
至於微生物用來抑制或殺滅其他微生物的物質,將其稱為抗生素。在歷史 上第一個被發現的抗生素是青黴素 (Penicillin),為英國科學家弗萊明 (Alexander Fleming) 在1928年所發現的,他發現被青黴菌 (Penicillium
notatum) 污染的培養皿中,細菌無法生長在青黴菌菌落周圍,經過研究之
後發現黴菌製造並分泌青黴素,其可以有效抑制細菌生長,以致於造成細 菌在黴菌菌落周圍無法生長的現象。1940年弗洛理 (Howard Walter Florey) 和錢恩 (Ernst Boris Chain) 成功純化出青黴素,並從動物實驗和人體試驗的 結果證實青黴素能有效抑制細菌所引起的症狀,使得弗萊明、弗洛理和錢 恩三人獲頒諾貝爾生理醫學獎。在此之後由於許多科學家的投入,使得其 他抗生素有如雨後春筍般的出現,例如鏈黴素 (Streptomycin)、氯黴素 (Chloramphenicol)、新黴素 (Neomycin)、土黴素 (Oxytetracycline)、四環黴 素 (Tetracycline) 等等。由於原核 (Prokaryote) 與真核 (Eukaryote) 生物在 DNA複製 (DNA replication)、轉錄 (Transcription)、轉譯 (Translation) 上的 差異,所以抗生素可以非常專一性地作用於細菌。自從青黴素成功運用於 臨床之後,在眾多科學家的努力之下,迄今所發現的抗生素已達幾千種之 多,然而抗生素的作用機制主要包括以下五種(圖1.2):1、抑制DNA或RNA合成
在DNA複製過程中,隨著解旋酶 (helicase) 將雙股DNA打開,造成 其他雙股DNA產生正超螺旋 (Positive supercoil) 結構,所以必須依賴 DNA旋轉酶 (DNA gyrase) 將雙股DNA切斷,並產生負超螺旋 (Negative supercoil) 而回復雙股DNA原本的結構。奎諾酮類 (Quinolones) 抗生素 能夠抑制第二型拓撲異構酶 (type Ⅱ topoisomerase) 的作用,在DNA複 製過程中,DNA旋轉酶 (DNA gyrase) 會與細菌DNA結合而形成一個複 合體,奎諾酮類抗生素能夠與此複合體進行結合,抑制複合體的瓦解,
進而阻斷DNA複製的進行,且造成細菌DNA斷裂 (Tran et al., 2005)。
Rifampin則可以藉由與細菌RNA聚合酶 (RNA polymerase) 的β-次 單位 (β-subunit) 結合,進而干擾轉錄作用的進行 (Villain-Guillot et al., 2007)。
2、抑制蛋白質合成
四 環 黴 素 可 藉 由 佔 據 細 菌 核 醣 體 中 的 胺 醯 tRNA 結 合 位 (Aminoacyl-tRNA site;A site),進而阻止胺醯tRNA的進入,達到干擾轉 錄作用的進行 (Connell et al., 2003)。
3、抑制細胞壁合成
在細胞壁合成過程中,青黴素結合蛋白 (penicillin-binding proteins;
PBPs) 協助轉胜肽酶 (transpeptidase) 執行胜聚醣 (peptidoglycan) 的聚 合作用,所以青黴素可藉由與青黴素結合蛋白作用而干擾細胞壁的合成 (Popham and Young, 2003)。
4、抑制新陳代謝路徑
Trimethoprim的結構與葉酸結構中的Pteridine類似,故能與葉酸競爭 二氫葉酸還原酶 (Dihydrofolate reductase) 的活性位置,進而干擾葉酸的 合成 (Tenover, 2006)。
圖1.2、抗生素的作用機制。
(摘自www2.bc.cc.ca.us/bio16/10_Antimicrobials.htm)
5、破壞細胞膜
多黏桿菌素B (Polymyxin B) 能夠在細菌細胞膜上形成孔洞,使得細 菌體內的物質流出而造成細菌死亡。
抗生素的發現無疑是人類醫學史上一個重要的里程碑,然而依照大自 然的法則,既然有微生物利用抗生素抑制其他微生物生長的現象,必然也 會有微生物抵抗抗生素以求生長的現象,否則特定菌種豈不遭受滅種的下 場,因此對於那些能夠抵抗抗生素的微生物,將其稱為抗藥性菌株。
三、抗藥性 (Antibiotic resistance)
醫院是最早出現抗藥性菌株的地方,因為醫院是大量使用許多抗生素 治 療 疾 病 的 場 所 (Levy, 1998),1930年代軍方醫院發現能抵抗磺胺劑 (Sulfonamide) 的化膿性鏈球菌 (Streptococcus pyogenes) (Levy, 1982),然 而,就在人類開始使用青黴素不久後,1940年代倫敦民間醫院發現能夠抵 抗青黴素的金黃色葡萄球菌 (Staphylococcus aureus) (Barber, 1948),相同 地,在開始使用鏈黴素治療肺結核之後,即在民生社區中發現能夠抵抗鏈 黴素的結核分枝桿菌 (Mycobacterium tuberculosis) (Crofton and Mitchison, 1948)。
在1950年代後期至1960年代初期之間,發現大腸桿菌 (Escherichia coli) 和腸道沙門氏菌 (Salmonella enterica) 等腸內菌出現多重抗藥性 (multiple drug resistance;MDR) 的現象 (Levy, 2001),這些多重抗藥性菌株導致嚴重 的臨床問題並造成死亡,特別是在開發中國家。然而抗性問題在當時並沒 有受到太多的重視,直到1970年代發現會造成呼吸道疾病的嗜血桿菌 (Haemophilus influenzae) 以及會造成泌尿生殖道疾病的奈瑟氏淋病雙球菌 (Neisseria gonorrhoeae) 皆具有抵抗安比西林 (Ampicillin) 的能力之後 (Elwell et al., 1977 and De Graaff et al., 1976),才使得人們開始注意到細菌抵 抗抗生素的嚴重性,另外,嗜血桿菌除了抵抗安比西林外,也會抵抗氯黴 素和四環黴素 (Marchall et al., 1984 and van Klingeren et al., 1977)。由於抗 生素的廣泛使用,導致許多不同菌種抵抗抗生素的能力逐漸提升,尤其是 在發展中國家,因為當地民眾不需處方籤就能購買抗生素。多重抗藥性菌 株造成治療上的困難,以致於必須同時使用多種不同的抗生素來治療疾病 (Iseman, 1993),因此抗藥性菌株會造成疾病治療上的危害及成本的負擔,
甚至導致無法治癒的現象。
細菌會為了適應環境的改變而調整基因的表現,任何改變對於細菌都 是一種壓力 (stress) 的來源,包括抗生素的處理。由於抗生素對細菌所產生 的壓力,導致細菌的基因發生突變,最後產生能夠抵抗抗生素的抗藥性基
因 (anti-drug genes) (Wang et al., 2001)。另外,細菌持續地接受抗生素所產 生的壓力,會導致細菌對抗生素的抗性從輕微抵抗發展成高度抵抗的能 力,因此抗生素的濫用造成對抗生素具敏感性的菌株死亡,同時也導致抗 藥性菌株的選殖 (Levy, 2002a)。然而,抗藥性基因藉由一些途徑從原本的 微生物體內散佈到其他不同種類的微生物體內 (Levy, 2002b) (圖1.3),這些 機制包括藉由噬菌體的感染而獲得抗藥性基因,此種機制稱為細菌轉染作 用 (Transfection)、藉由細菌之間的接合 (Conjugation) 作用而獲得帶有抗藥 性基因的質體或者藉由攝入裸露的DNA而獲得抗藥性基因,稱此種機制為 細菌轉形作用 (Transformation) (Levy and Marshall, 2004)。抗藥性基因通常 只會對抗單一家族或種類的抗生素,然而單一抗生素長期的使用,所誘導 出來的抗藥性菌株不僅能夠抵抗該抗生素,也會抵抗其他不同結構的抗生 素 (Levy, 2002a),雖然目前仍不清楚長期使用單一抗生素可以誘導出多重 抗藥性菌株的原因,但是這個現象是因為許多不同的抗藥性基因連接在同 一個轉位子或質體上,可能的解釋為經由誘導出來的抗藥性菌株似乎有從 其他菌株獲得其他抗藥性基因的傾向 (Levy and Marshall, 2004),由轉位子 將這些抗藥性基因集中在一個質體內,導致一個微生物體內帶有多種抗藥 性基因,而造成多重抗藥性菌株的出現。
圖1.3、細菌獲得外來抗藥性基因的途徑。(摘自Levy and Marshall, 2004)
人類將抗生素充分地應用在醫學、畜牧飼養、食品防腐等方面長達60 年,廣泛使用抗生素的結果造成人類與動物病原菌對抗生素的抗性增加,
目前發現細菌抵抗抗生素的機制包括以下五種方式 (Xu et al., 2006) (圖 1.4):
1、降低細胞膜對抗生素的通透性
由於抗生素抑制細菌生長的機制多半是藉由與細菌體內的特定蛋白 質結合,所以當細菌成功把抗生素阻擋在外,可使抗生素無法到達細菌 體內而與特定蛋白結合,例如綠膿桿菌 (Pseudomonas aeruginosa) 對於 β-內醯胺類 (β-lactam) 抗生素的通透性 (permeability) 很低 (Xu et al., 2006)。
2、將菌體內的抗生素排出
藉由排出幫浦 (efflux pump) 以主動運輸的方式將細菌體內的抗生 素運到細菌體外,例如大腸桿菌的AcrAB-TolC所組成的多重抗藥性排出 幫浦 (multidrug efflux transporter) 可以主動排出四環黴素、氯黴素和氟 奎諾酮類 (Fluoroquinolones) 的抗生素 (Chen et al., 2007)。
3、改變抗生素標的蛋白的結構
抗生素的標的蛋白之基因發生突變,使抗生素無法與其產物作用,
例如金黃色葡萄球菌的gyrA發生突變,可降低細菌對奎諾酮類抗生素的 敏感性 (susceptibility) (Hawkey, 2003)。
4、破壞或改變抗生素結構
藉由酵素分解抗生素,以破壞抗生素的結構,例如β-乙內醯胺酶 (β-lactamases) 可以破壞青黴素和頭孢菌素 (Cephalosporins) 的結構,或 是改變抗生素的結構,使其無法與標的蛋白作用,例如金黃色葡萄球菌 製造胺基糖磷酸轉移酶 (aminoglycoside phosphotransferase) 將胺基糖類 (aminoglycosides) 抗生素磷酸化,使其失去抗菌的活性 (Wright and Thompson, 1999)。
圖1.4、細菌抵抗抗生素的機制。(摘自www.scq.ubc.ca/?p=405)
5、改變代謝路徑
如果抗生素的標的蛋白是細菌代謝路徑上的酵素,藉由改變代謝路 徑, 可避開抗生素對細菌的影響,例如金黃色葡萄球菌可以藉由製造氯 黴素結合蛋白2 (penicillin-binding protein 2) 而避開萬古黴素 (Vanco- mycin) 抑制細胞壁合成的作用 (Hanaki et al., 1998)。
有趣的是細菌不僅發展出多種不同的抗性機制,就連其抗藥性基因也 有許多的差別,許多不同的抗藥性基因皆可以表現出抵抗同一種類抗生素 的抗性機制,例如,表現β-乙內醯胺酶的基因有數以千計之多,調控四環黴 素排出的抗藥性基因則有20種以上 (Levy et al., 1999)。另外,有一種以上 的抗性機制可以對抗相同的抗生素,例如,將四環黴素排出或改變核醣體 上四環黴素的作用位置皆為抵抗四環黴素的機制 (McMurry and Levy, 2000)。
四、抗菌胜肽 (Antimicrobial peptides;AMPs)
由於細菌能夠快速地發展抵抗抗生素的機制,抗生素的廣泛使用對細 菌而言,無疑是一種加速篩選抗性菌株的方式。多重抗藥性病原菌 (multiple antibiotic-resistant pathogens) 的出現,將會使人類面臨無藥可醫的困境。因 此,近年來國際間禁止使用抗生素在養殖業或是其他行業上,使得我們必 須尋找另一種可以用來取代抗生素的抗菌物質,越來越多的研究證實抗菌 胜肽具有有效的殺菌能力。
大約20 年前,科學家們從青蛙的皮膚、昆蟲的淋巴和人類的嗜中性白 血 球 中 發 現 含 有 一 種 帶 正 電 的 胜 肽 , 其 可 以 作 為 有 效 的 抗 微 生 物 劑 (Hancock, 2001),稱之為抗菌胜肽。迄今已經在自然界的所有生命體中發現 超過600 個抗菌胜肽,說明抗菌胜肽廣泛地存在於所有自然界的生命體中,
包括細菌、真菌、植物及動物。大部份的抗菌胜肽皆以帶有訊號胜肽 (signal peptide) 且結構較大的前驅物 (precursor) 形式存在於生命體細胞中,再經 由蛋白質裂解 (proteolysis) 或基團添加 (例如糖基化 (glycosylation) 或鹵 化 (halogenation)) 等轉譯後修飾 (Post-translational modification;PTM) 作 用而成為具活性的抗菌胜肽 (Zasloff, 2002)。抗菌胜肽一般是由 12-50 個胺 基酸所組成且分子量小於10 kD (Hancock, 2001 and Ganz, 2001)。但是就其 胺基酸序列、二級結構及胜肽片段的大小都非常地不同 (Hancock, 2000 and Ganz and Lehrer, 1998)。其二級結構大致上可分成 α-螺旋結構 (α-helix structure),例如人類的 LL-37、兩棲動物的 Magainins、昆蟲的 Cecropins。
以及β-折板結構 (β-sheet structure),例如 Defensins (Peschel and Sahl, 2006)。
抗菌胜肽最廣為人知的結構就是其為一段具有正電荷及疏水性能力的 短鏈胜肽,其具有廣泛的抗微生物活性,可以對抗細菌、真核的寄生蟲、
病毒和真菌,更重要的是抗菌胜肽可以有效地對抗抗藥性細菌 (antibiotic resistant bacteria) (Hancock, 2001)。而抗菌胜肽在人體內主要存在於黏膜層 及皮膚組織中,做為人體防禦機制的第一道防線。另外,也被當作吞噬細 胞用來殺菌的武器之ㄧ (Peschel and Sahl, 2006)。
五、抗菌胜肽作用機制 (Action mechanisms of AMP)
由於細菌表面帶有大量的負電荷,例如革蘭氏陰性菌細胞外膜上脂多 醣 (lipopolysaccharide;LPS) 的磷酸基團或是存在於革蘭氏陽性菌細胞壁 中的脂磷壁酸 (lipoteichoic acid),所以最初對於抗菌胜肽殺菌機制的了解,
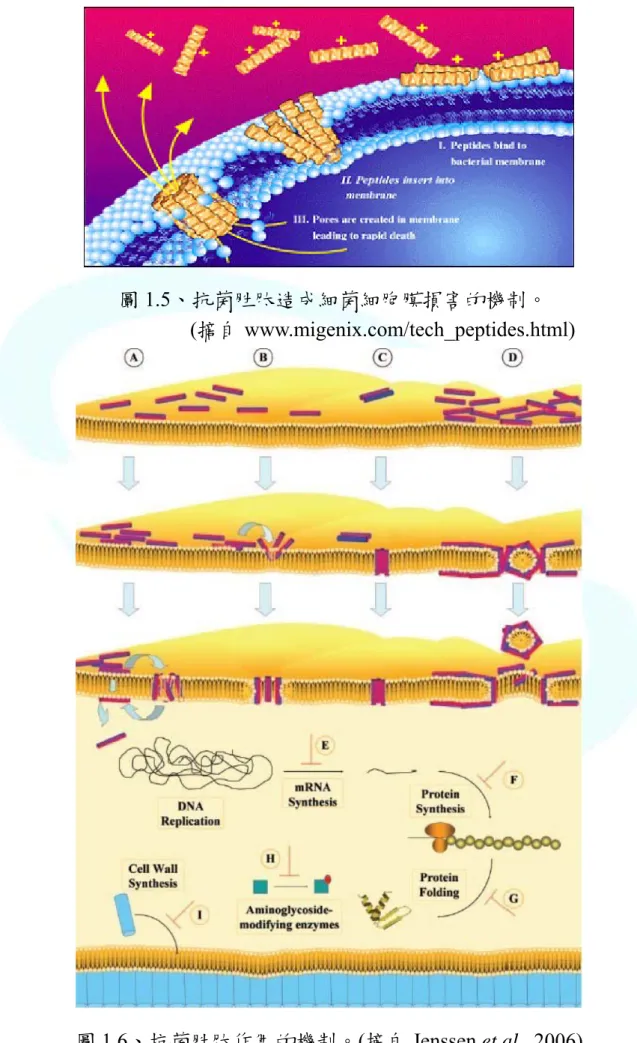
認為抗菌胜肽主要是藉由本身的正電荷與細菌表面的負電荷形成交互作 用,藉由Oriented circular dichroism (OCD) 實驗可知,隨著抗菌胜肽與細菌 細胞膜上脂質的混合比例不同,而有兩種不同的模式,稱為兩階段模式 (two-state model),包含 surface state (S-state) 及 insert state (I-state) (Haung, 2000)。S-state 主要是發生在抗菌胜肽與脂質的混合比例低的時候,此階段 的抗菌胜肽只能平行地吸附在細菌細胞膜表面,然而當抗菌胜肽與脂質的 混合比例到達一定的程度,抗菌胜肽則由平行吸附在細胞膜上轉成垂直插 入細胞膜,並聚集而形成孔洞,稱之為I-state。所以抗菌胜肽藉由在細菌細 胞膜上形成孔洞而造成細菌細胞膜的完整性被破壞,進而達到殺滅細菌的 效果 (圖 1.5)。
目前科學家們提出 4 種關於抗菌胜肽與膜之間的作用模式 (圖 1.6) , 分 別 是 聚 集 模 式 (Aggregate model) 、 超 環 面 孔 洞 模 式 (Toroidal pore model)、桶狀穿鑿模式 (Barrel-stave model) 和地毯模式 (Carpet model)。
由於某些抗菌胜肽 (例如 Magainin) 缺乏在細胞膜上形成通道的結 構,所以在膜中無特定的方向,故此類的抗菌胜肽在膜上所形成的孔洞不 僅差異很大也很不穩定,在通道瓦解之後,抗菌胜肽則會穿越細胞膜而進 入細胞質中 (Matsuzaki et al., 1996 and Matsuzaki, 1998),稱此種模式為聚集 模式。某些抗菌胜肽 (例如 Melittin 和 LL-37) 會以與細胞膜垂直的方向插 入細胞膜之後,使膜上脂質的親水性區域向內彎曲而形成孔洞,而抗菌胜 肽則排列在孔洞邊緣,稱此種模式為超環面孔洞模式。某些抗菌胜肽 (例如 Alamethicin 和 Gramicidin S) 在插入細胞膜之後,其疏水性區域會與細胞膜 的疏水性區域作用,而親水性區域則形成孔洞的內腔,稱此種模式為桶狀 穿鑿模式。某些抗菌胜肽 (例如 Ovispirin) 會平行地吸附在細胞膜表面,有
圖1.5、抗菌胜肽造成細菌細胞膜損害的機制。
(摘自 www.migenix.com/tech_peptides.html)
圖 1.6、抗菌胜肽作用的機制。(摘自 Jenssen et al., 2006)
如地毯一般,故稱為地毯模式,細胞膜上聚集了大量的抗菌胜肽,抗菌胜 肽會包圍細胞膜的一部份並形成類似微胞 (micelle) 的形式,細菌細胞膜則 因為一部份被抗菌胜肽帶走而形成一個孔洞,稱此種模式為地毯模式。以 上模式所造成的結果可以歸類為以下三種,1、短暫通道的形成。2、微胞 形成 (Micellarization) 或膜的溶解。3、穿越細胞膜。因此,依據抗菌胜肽 的作用模式可以概略地分成與膜作用 (membrane-acting) 和不與膜作用 (non-membrane acting) 兩種作用 (Jenssen et al., 2006)。
然而也有許多的證據指出某些抗菌胜肽可以與細菌中的特定目標作 用,包括抑制核酸合成、蛋白質合成、酵素活性及細胞壁合成 (Brogden, 2005 and Jenssen et al., 2006)。姑且不論抗菌胜肽精確的作用模式,抗菌胜肽的殺 菌活性幾乎普遍地需要與細菌細胞膜進行交互作用 (Hancock and Rozek, 2002)。
六、細菌抵抗抗菌胜肽的機制
目前發現抗菌胜肽的作用機制非常廣泛,其作用的對象包括了細菌的 外膜、細胞壁、細胞膜、細胞質及核酸,由此可知抗菌胜肽的作用機制不 只是損害細菌細胞膜。然而近年研究顯示細菌也會針對抗菌胜肽的作用機 制進行演化,不斷地發展出抵抗特定抗菌胜肽作用的抗性機制 (圖 1.7) ( Yeaman and Yount, 2003)。以下介紹幾種已經廣泛被研究的細菌抗性機制:
1、胞外降解 (Extracellular degradation)
細菌藉由釋放多種蛋白酶至環境中,分解環境中具有α-螺旋結構的 抗菌胜肽,以避免抗菌胜肽對細菌造成傷害。例如,金黃色葡萄球菌 (Staphylococcus aureus) 會製造 V8 及 aureolysin 等蛋白酶,以及沙門氏 菌 (Salmonella enterica serovar Typhimurium) 會製造外膜蛋白酶 PgtE (Peschel, 2002)。
圖1.7、抗菌胜肽的作用機制與細菌發展的抗性機制。
(摘自 Yeaman and Yount, 2003)
2、胜肽排出 (Peptide efflux)
除了製造蛋白酶以外,細菌也藉由製造位在細胞膜上的排出幫浦將 成功闖入細菌體內的特定抗菌胜肽排出於菌體外。而 exporter 主要藉由 辨認抗菌胜肽的特定胺基酸序列或是結構上的motif,進而將特定的抗菌 胜肽排出菌體外。例如,在奈瑟氏淋病雙球菌 (Neisseria gonorrhoeae) 及腦膜炎雙球菌 (Neisseria meningitidis) 中發現的多重抗藥性排出幫浦 MtrCDE 能夠排除 Protegrins 及 LL-37 (Peschel and Sahl, 2006)。
3、淨電荷改變 (Net charge alteration)
細菌為了有效對抗抗菌胜肽的多樣性,而發展出一套新的抗性機 制。藉由修飾細菌細胞膜或細胞壁中特定成份,達到降低或中和細菌表
面的負電荷,使得抗菌胜肽無法輕易靠近細菌。例如,在金黃色葡萄球 菌中,可藉由細胞壁中的磷壁酸 (teichoic acid) 與 D-丙胺酸結合或是藉 由細胞膜中磷脂醯甘油 (phosphatidyl glycerol;TG) 與 D-離胺酸結合,
而在沙門氏菌中可藉由 lipid A (為細菌外膜中脂多醣的成份之ㄧ) 與胺 基阿拉伯糖 (aminoarabinose) 結合,來達到減少表面負電荷的效果 (圖 1.8) (Peschel, 2002)。
圖1.8、細菌減少或中和表面負電荷的機制。
(摘自 Peschel, 2002)
七、細菌的偵測系統
細菌能夠適應的環境非常廣泛,例如腸炎弧菌可以在海水中生存,也 可以在人體中生存,然而兩者之間除了營養份多寡的差別外,最大的差別 在於鹽濃度的不同,所以細菌可以藉由調控特定基因的表現,使其能夠在 高鹽或低鹽的環境中生存。同樣地,細菌在人體中生存,必定會遭遇到抗 菌胜肽的攻擊,所以細菌則需要一套偵測系統來偵測所處環境中是否有抗 菌胜肽的存在,再藉由訊號傳遞的方式來調控特定基因表現,以達到保護 細菌免受抗菌胜肽所造成的傷害。
目前研究發現細菌有許多不同的偵測系統,依照訊號傳遞的方式可以 分成兩種類型,分別是調控膜內蛋白質裂解 (Regulated intramembrane proteolysis;RIP) 跟雙成份調控系統 (Two-component regulatory systems)。
1、調控膜內蛋白質裂解
細菌在不同的環境下會利用不同的 Sigma factor 來調控特定基因的 表現,這樣的特性使得細菌能夠在艱難的環境下存活。Sigma factor 為 RNA 聚合酶的次單位之ㄧ,當其與核心酵素 (core enzyme) 結合則稱為 全酵素 (Holoenzyme)。Sigma factor 具有辨認特定啟動子的功能,所以 能夠帶領 RNA 聚合酶至特定的啟動子,並使特定基因表現。細菌具有 多種alternative sigma factor,視情況不同而由特定的 sigma factor 決定基 因的表現。例如,在大腸桿菌中,一般主要是由σ70調控基因的表現,但 在低溫、高滲透壓或碳源不足的情況下,則由σS調控相對基因的表現。
其他的sigma factors 還有在熱緊迫 (heat shock) 下調控基因表現的 σE及 σ32,在氮源不足時調控基因表現的 σ54,調控鞭毛基因表現的 σ28。雖然 σE與σ32都是在高溫環境下調控基因表現,但是 σ32是在環境溫度稍微高 於細菌最適生長溫度的情況下調控基因表現,且受到細胞質中發生錯誤 折疊的蛋白質所誘導,而σE則在非常極端的高溫下調控基因表現,受到 膜間質或外膜上發生錯誤摺疊的蛋白質誘導。
圖1.9、大腸桿菌的 σE活化路徑。(摘自 Rowley et al., 2006)
在一般情況下,大腸桿菌的 σE會被位在細胞膜上的RseA 結合,當 膜間質或外膜存在錯誤折疊的蛋白質,此蛋白質會與細胞膜上 DegS 的 PDZ domain 結合,啟動 DegS 的蛋白質酶活性,進而切除 RseA 在膜間 質側的部份,接著由 YaeL 切除 RseA 在細胞質側的部份,SspB 將被切 下的部份送入ClpP-ClpX 複合體中進行分解,最後釋放 σE而調控特定基 因的表現 (Rowley et al., 2006) (圖 1.9)。由此對於 alternative sigma factor 調控細菌抵抗抗菌胜肽的機制有了初步的認識,推測這些受到alternative sigma factor 調控的基因產物應該能夠改善抗菌胜肽對細菌所造成的傷害 (Mathur et al., 2007)。而這種藉由裂解膜蛋白而釋放 sigma factor 的方式 則稱為調控膜內蛋白質裂解。
然而外膜蛋白 (outer membrane proteins;OMPs) 似乎可以調控許多 格蘭氏陰性菌的σE表現量,將可以表現正常OmpU 的質體送入 ompU 突 變的霍亂弧菌 (Vibrio cholerae) 中,可回復 σE的表現量,且改善 ompU 突變株對抗菌胜肽的敏感性。
2、雙成份調控系統
雙成份調控系統是細菌廣泛使用來偵測並對胞外壓力做出反應的 一套訊號傳遞系統,系統中的成員主要有 2 個,一個為具有跨膜結構的 感應器激酶 (sensor kinase),另一個為位於胞內的反應調節者 (response regulator)。CpxAR 系統是與調節封套壓力 (envelope stress) 有關的典型 雙成份調控系統 (DiGiuseppe and Silhavy, 2003),在大腸桿菌遭受某些封 套壓力的情況下,Cpx 系統可以控制超過 100 種蛋白質的表現,其中有 些蛋白質也會受到σE的調控。
圖 1.10 為 Cpx 系統的訊號傳遞路徑,當細菌遭受封套壓力的情況 下,會促使CpxP 與 CpxA 分離,此一分離使得 CpxA 的激酶活性被活化,
CpxA 會先磷酸化自己,接的再將磷酸根轉移至 CpxR 身上,獲得磷酸根 的 CpxR 則轉變成活化態,可以藉由特異性地結合在某些基因上游的辨 認位置 (recognition site) 而調控下游基因的表現 (Rowley et al., 2006)。
圖 1.10、大腸桿菌的 Cpx 系統之路徑。
(摘自 Rowley et al., 2006)
八、宿主-病原菌交互作用 (Host-pathogen interaction)
宿主-病原菌交互作用就像是一場宿主與病原菌之間的攻防戰,包括病 原菌如何適應宿主及宿主如何防止病原菌在體內形成菌落化 (colonization) 及對組織造成傷害。許多研究證實大多 數的病原菌對特定宿主具有高度的 適應能力,這樣的能力使得病原菌能在宿主體內形成菌落化,並對組織造 成傷害。許多病原菌或多或少都帶有針對特定抗菌胜肽的抗性能力,所以 病原菌可以藉由抵抗特定抗菌胜肽的作用而使其能夠適應特定的宿主 (Kraus and Peschel, 2006)。
當然,宿主也會發展能有效對抗細菌抗性機制的抗菌胜肽,例如,人 類β-防禦素 3 (Human β-defesin 3;hBD3) 被發現能夠有效地抑制具有降低 表面負電荷能力的金黃色葡萄球菌生長,經由序列分析發現人類β-防禦素 3 帶有+10 價,故推測宿主藉由提高抗菌胜肽的正電荷,使其能與細菌表面 殘留的負電荷交互作用。不過,由於抗菌胜肽的序列大小有限,所以如果 提高抗菌胜肽的正電荷,相對地會減少抗菌胜肽的疏水性能力,也就是說 人類 β-防禦素 3 可能藉由其他的機制進行殺菌。進一步研究發現人類 β-防 禦素3 會取代自溶素 (autolysins) 在細胞壁的結合位 (anchoring site),使自 溶素被釋放且造成細菌細胞壁的瓦解。自溶素是細菌在進行細胞分裂時用 來瓦解細胞壁的陽性酵素。由此我們可以了解抗菌胜肽藉由提高本身正電 荷及發展新的殺菌機制來對抗細菌降低表面負電荷的策略 (Peschel and Sahl, 2006)。
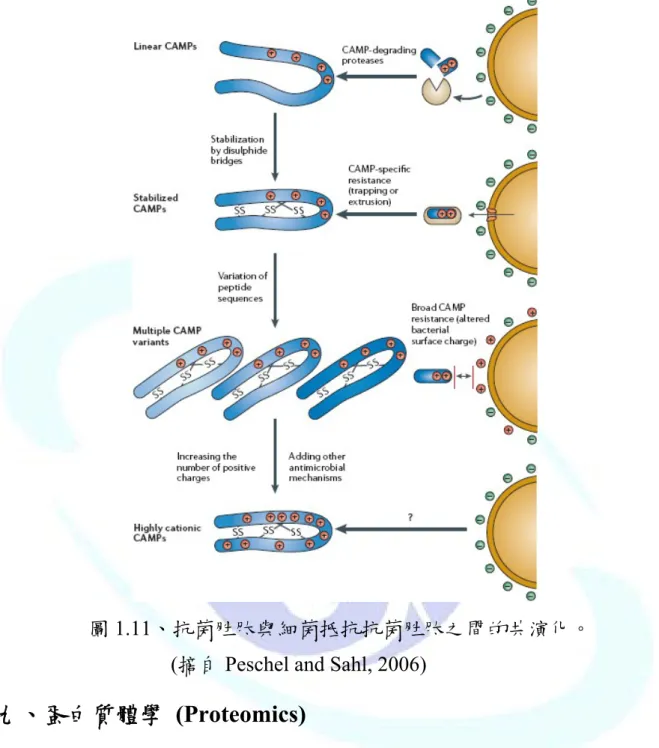
每當細菌發展出新的抗性機制對抗抗菌胜肽時,宿主也會跟著發展出 能夠逃避或抵抗細菌抗性機制的抗菌胜肽來對抗細菌。由此我們可以了解 宿主的抗菌胜肽與細菌抵抗抗菌胜肽的機制是不斷地進行演化,所以宿主- 病原菌交互作用也可以說是抗菌胜肽與抗菌胜肽抗性菌株的共同演化 (Co-evolution) (圖 1.11) (Peschel and Sahl, 2006)。
圖1.11、抗菌胜肽與細菌抵抗抗菌胜肽之間的共演化。
(摘自 Peschel and Sahl, 2006)
九、蛋白質體學 (Proteomics)
2001 年美國 Celera 生技公司完成人類基因定序,驚訝地發現人類基因 序列中僅含有3 萬至 4 萬個基因,不如原先所預期的 10 萬多個基因,顯示 有相當多的生物功能資訊是儲存在基因轉錄後的產物 (mRNA),而 mRNA 所攜帶的功能訊息必須再透過其轉譯後產物 (蛋白質) 才能執行其生理功 能。因此,以基因序列訊息資料庫為基礎的生物資訊學搭配新一代質譜技 術整合發展出蛋白質體學,並開啟了後基因體時代 (Post genomic era) 的大 門 (王和吳,2003)。
由於組織或細胞內的蛋白組成異常複雜,原核生物和真核生物可能分 別由數千或上萬個不同的蛋白質所組成,現階段的質譜儀仍無法一次分析 數以千計的蛋白質,因此在樣品進行質譜分析之前,必須藉助適當的蛋白 質分離技術降低樣品複雜程度。搭配質譜儀技術的蛋白質分離技術,大致 可分為 gel-base 或 gel-free 兩種,gel-base 指的是利用二維凝膠電泳 (Two- dimensional gel electrophoresis;2-DE) 分離蛋白質混合物,再以質譜儀分析 鑑定膠體所呈現出的蛋白質。而 gel-free 則是利用液相層析法 (liquid chromatography) 分離蛋白質或胜肽,再以液相層析-電噴灑串聯式質譜儀分 析蛋白質 (陳,2005)。
蛋白質體學在後基因體時代中是一門具有高準確性、高效能及高產率 的蛋白質分析方法,提供我們對於標的細胞在特定情況下基因表現的了解 (Peng et al., 2005)。藉由二維凝膠電泳分析細胞萃取物的總蛋白質時,由於 單一細胞中不同蛋白質的含量差異可以到達107-1010倍,導致含量豐富的蛋 白質會覆蓋住許多低含量蛋白質,以致於使其無法被偵測。因此,針對不 同的研究目標來使用前分群策略,可以偵測到更多的蛋白質,提升目標蛋 白質體的完整性,如此才能以功能性蛋白質體學來深入瞭解各種生理、病 理機制。前分群的方法一般可分為從組織中分離特定細胞、次細胞分群 (Subcellular fractionation)、階段萃取方式 (Sequential extraction method)、特 定轉譯後修飾的蛋白之純化。然而,藉由前分群策略所獲得的部份蛋白質 體則稱為次蛋白質體 (Subproteome) (陳,2005)。
第 二 章 實 驗 目 的
本研究室自2005 年起,嘗試以天然界存在之近千種抗菌胜肽序列為模 版,設計合成出一系列人工抗菌胜肽,經測試其對 7 種革蘭氏陽性菌與 11 種革蘭氏陰性菌 (含 4 種弧菌屬細菌) 之抗菌活性與專一性,並同時評估抗 菌胜肽之溶血活性後,挑選出數條對腸炎弧菌此一重要水生病原菌具高度 抗菌活性與專一性,且同時具低溶血活性的抗菌胜肽,為求日後能實際將 抗菌胜肽施用於生物體,取代目前抗生素的作用,我們希望先期探討病原 菌對抗菌胜肽可能產生的抗性機制,進而研擬對策回應之,以避免抗菌胜 肽重蹈抗生素的覆轍。由於革蘭氏陰性菌的外膜是直接與外在環境接觸的 部位,因此外膜上的蛋白質能夠反應革蘭氏陰性菌因應環境改變而表現的 封套壓力反應 (Envelope-stress response) (Walsh et al., 2003),而革蘭氏陰性 菌內膜上的蛋白質則主要是與代謝、訊號傳遞等相關的蛋白質。因此,本 研究將利用蛋白質體學方法探究野生型腸炎弧菌與能對抗菌胜肽產生抗性 之抗性菌株之間的膜蛋白表現之差異,以深入了解其可能的抗性機制。
第 三 章 實驗材料與儀器
實驗材料
一、菌株:
Vibrio parahaemolyticus
二、藥品:
1、菌種培養液及培養基
1 Difco Tryptic Soy Agar Soybean-Casein Digest Agar, USP (TSA)、
Bacto Tryptic Soy Broth Soybean-casein Digest Medium (TSB) 購 自Becton Dickinson (USA)
2 Sodium Chloride (NaCl) 購自 Wako (Japan) 2、抗菌胜肽及抗生素
1 所有人為設計或天然存在的抗菌胜肽皆由科羅耐國際科技公司 代為合成
2 Ampicillin、Kanamycin、Penicillin G 及 Tetracycline 購自 Sigma (USA)
3、蛋白質分劃 (Fractions)、萃取及定量
1 N-Lauroylsarcisine Sodium salt Solution 購自 Fluka (Switzerland)
2 3-([3-cholamidopropyl]dimethylammonio)-1-propane sulfonate (CHAPS)
、DL-Dithiothreitol (DTT)、Thio-urea、Bovine serum albumin (BSA) 購自Sigma (USA)
3 Acetone 購自 Mallinckrodt (USA)
4 Bromophenol blue 、 IPG buffer 購 自 Amersham Biosciences (Swenden)
5 Glycerol、Sodium Dodecyl sulfate (SDS) 購自 Merck (Germany) 6 Urea 購自 Wako (Japan)
7 95% Ethonal 購自慶發 (Taiwan)
8 Bio-Rad Protein Assay Reagent Dye Reagent Concentrate 購自 Bio-Rad (USA)
9 dialysis membrane Spectra/Pro 6, MWCO: 8kDa 購自 SPECTRUM (USA)
4、二維凝膠電泳
1 0.5 M Tris-HCl (pH 6.8)、1.5 M Tris-HCl (pH 8.8) 購自 Amresco (USA)
2 30% acrylamide-0.8% bis-acrylamide、Precision Plus Protein All Blue Standards 購自 Bio-Rad (USA)
3 Ammonium persulfate、TEMED (N,N,N’,N’-tetramethyl ethylene diamine) 購自 Amersham Biosciences (Swenden)
4 Isopropyl alcohol (isopropanol) 購自 Merck (Germany)
5 Iodoacetamide (IAA) 購自 Amersham Biosciences (Swenden) 6 Mineral oil 購自 Pharmacia (Swenden)
7 GelCode Blue Stain Reagent 購自 Pierce (USA) 8 Tris (base) 購自 J. T. Baker (USA)
9 Glycine 購自 Wako (Japan)
IPG strip Immobiline DryStrip, pH 4-7, 7cm 購自 GE Healthcare (USA) (formerly Amersham Biosciences)
5、膠體內水解及質譜儀分析
1 Ammonium bicarbonate 購自 Sigma (USA) 2 Acetonitrile (ACN) 購自 Merck (Germany)
3 Sequencing Grade Modified Trypsin 購自 Promega (USA) 4 Trifluoroacetic acid (TFA) 購自 Riedel-deHaen (Germany) 5 α-Cyano-4-hydroxycinnamic acid (CHCA) 購自 Sigma (USA)
6 Sequazyme peptide mass standards kit 購自 Applied Biosystems (USA)
7、RNA 萃取
1 TRIzol reagent 購自 invitrogen (USA)
2 Chloroform、Isopropyl alcohol (isopropanol) 購自 Merck (Germany) 3 Diethyl pyrocarbonate (DEPC) 購自 Sigma (USA)
4 95% Ethonal 購自慶發 (Taiwan) 8、逆轉錄和即時定量聚合酶連鎖反應
1 所有 primers 皆由明欣生物技術有限公司代為合成
2 oligo-d(T)18 primer、10 mM dNTPs Mix 購自 GeneTeks BioScience Inc. (Taiwan)
3 SuperScriptTM Ⅲ Reverse Transcriptase 購自 invitrogen (USA) 4 FastStart Universal SYBR Green Master (Rox) 購 自 Roche
(Switzerland)
實驗儀器
一、菌種培養、最小抑菌濃度及篩選抗性菌株
1、無菌操作台 Laminar flow, Chu An Instruments CO. (Taiwan) 2、恆溫培養箱 Incubator E600L, Deng Yng Instruments CO. (Taiwan) 3、分光光度計 Spectrophotometer Heλios β, Thermo Electron corporation
(USA)
4、高速離心機 High Speed Refrigerated Centrifuge 6500, KUBOTA (Japan)
5、微量盤分析儀 Microplate reader SpectraMax M2, Molecular Devices (USA)
二、蛋白質分餾 (Fractions)、萃取及定量
1、高速離心機 High Speed Refrigerated Centrifuge 6500, KUBOTA (Japan)
2、超音波細胞破碎機 Ultrasonic Cell Disrupter SONIFIER 150D Micro, BRANSON (USA)
3、超高速離心機 Ultracentrifuge CP-MX Series CS 150 GXL, HITACHI (Japan)
4、微量盤分析儀 Microplate reader SpectraMax M2, Molecular Devices (USA)
5、冷凍真空乾燥機 Freeze Dryer with shell freezer, Ilshin Lab.co., Ltd (Korea)
三、二維凝膠電泳
1、等電點焦集電泳儀 Ettan IPGphor Ⅱ, GE Healthcare (USA)
2、等電點電泳膠條陶瓷容槽 Strip Holder, 7 cm, GE Healthcare (USA) 3、蛋白質電泳套組 Bio-Rad Mini-PROTEAN 3 Electrophoresis Module
Assmebly, Bio-Rad (USA)
4、電源供應器 Bio-Rad PowerPac Basic, Bio-Rad (USA)
5、平台式全彩雷射掃描器 Epson perfection V750 PRO, Epson (USA) 四、膠體內水解及質譜儀分析
1、雜交烘箱 Hydridization oven DHO-101, Firstek (Taiwan)
2、超音波洗淨機 Ultrasonic cleaner 5510R-DTH, BRANSON (USA) 3 、 離 心 式 真 空 濃 縮 乾 燥 機 Speed Vac Univapo 100H, UniEquip
(Germany)
4 、 基 質 輔 助 雷 射 脫 附 游 離 飛 行 時 間 質 譜 儀 Matrix-assisted laser desorption/ionization time of flight mass spectrometer (MALDI-TOF- MS) voyager DE-STR BiospectrometryTM Workstation, Applied Biosystems (USA)
(感謝台大醫學院生化暨分子生物學研究所暨基因體中心 周綠蘋教 授提供)
5 、 液 相 層 析 電 噴 灑 四 級 棒 飛 行 時 間 串 聯 式 質 譜 儀 liquid chromatography electrosprey ionization quadrupole time of flight tandem mass spectrometer (LC-ESI-Q-TOF-MS/MS) 1200 Series capillary LC pump, Agilent (USA), Finnigan LCQDECA Ultra-Sensitive Benchtop MSn System, Thermo Electron corporation (USA)
(委託台大醫學院生化暨分子生物學研究所 張智芬教授實驗室及鎂 陞科技股份有限公司代為分析)
五、RNA萃取
1、高速離心機 High Speed Refrigerated Centrifuge 6500, KUBOTA (Japan)
2、分光光度計 Spectrophotometer Heλios β, Thermo Electron corporation (USA)
六、逆轉錄和即時定量聚合酶連鎖反應
1、PCR加熱循環器 PCR thermal cycler T-CY, Creacon technologies (Holland)
2、溫度梯度即時核酸定量分析系統 iCycler iQ5 multicolor Real Time PCR detection system, Bio-Rad (USA)
第 四 章 實 驗 方 法
一、菌種培養 (Bacterial culture)
腸炎弧菌的冷凍保存菌液在室溫下解凍後,移至無菌操作台中,以微 量吸管管嘴 (pipette tip) 尖端沾取少量菌液並塗抹在含有 1.5% NaCl 的 TSA 固體培養基上。將此培養基置於 28℃恆溫培養箱中培養約 8-9 小時,
即可從培養基上獲得腸炎弧菌的單一菌落,之後則視實驗的需求而將菌落 接種在適當體積的培養液中。如果腸炎弧菌菌液是要用於測定最小抑菌濃 度的實驗,則將單一菌落接種在裝有3-4 mL 含 1.5% NaCl 之 TSB 培養液 的養菌管中。若是用於抗性菌株的篩選,則將菌落接種在裝有5 mL 含 1.5
% NaCl 之 TSB 培養液的養菌管中。若是要進行萃取腸炎弧菌蛋白質的話,
則將菌落接種在裝有200 mL 含 1.5% NaCl 之 TSB 培養液的 1 L 滅菌錐形 瓶中。最後將接種後的培養液置於28℃恆溫培養箱中隔夜培養。
二、最小抑菌濃度 (Minimal inhibitory concentrations;MICs)
最小抑菌濃度從字面上的解釋為抑制細菌生長所需的最小濃度,然而 根據以下的實驗處理方法,可以將最小抑菌濃度更精確地定義為將細菌遲 滯期 (Lag phase) 延長 48 小時所需要的最小濃度。在 3-4 mL 的腸炎弧菌菌 液經過隔夜培養後,添加1 mL 菌液至測定管 (cuvette) 中,並利用分光光 度計在600 nm 波長下測量腸炎弧菌菌液的濁度 (OD600)。接著則以菌液的 濁度推測菌液中可能的菌落數,假設 OD600=1,則表示菌液中的菌落數約 為109 CFU/mL,若 OD600=2,則表示菌液中的菌落數約為 2×109 CFU/mL,
以此類推。在得知菌液中的菌落數之後,以含1.5% NaCl 的 TSB 培養液將 腸炎弧菌菌液的菌落數稀釋成104 CFU/mL,並添加 135 μL 經稀釋後的菌液 至96 孔盤中。之後則分別添加 15 μL 經過連續稀釋的抗菌胜肽及抗生素至 96 孔盤中,劑量濃度以 640 μg/mL 為最大濃度,並逐步 2 倍稀釋成 320、
160、80、40、20 μg/mL,所以當 15 μL 的抗菌胜肽或抗生素與 135 μL 菌液 混合後,在菌液中的抗菌胜肽或抗生素濃度則為64、32、16、8、4、2 μg/mL。
當抗菌胜肽或抗生素分別與菌液混合後,將96 孔盤移至 28℃恆溫培養箱中 培養48 小時。最後利用微量盤分析儀在 600 nm 波長下測量菌液的的濁度。
在本論文研究工作中所使用的抗菌胜肽分別為14 種實驗室所設計開發的抗 菌胜肽及2 種天然存在的抗菌胜肽 (周,2007),抗生素則分別為安比西林、
卡那黴素 (Kanamycin)、青黴素 G (Penicillin G) 及四環黴素。
三、篩選抗性菌株 (Selection of AMP-resistant strains)
在獲得抗菌胜肽對腸炎弧菌的最小抑菌濃度之後,選擇能夠有效抑制 腸炎弧菌生長的抗菌胜肽作為篩選抗性菌株實驗的誘導劑,並以最小抑菌 濃度的一半劑量為起始濃度進行誘導腸炎弧菌對抗菌胜肽產生抗性。抗性 菌株的誘導方法乃依據Peng 等人在 2005 年發表的研究報告,並稍做修飾。
首先添加最小抑菌濃度的一半劑量之抗菌胜肽至5 mL 腸炎弧菌菌液中,並 在28℃恆溫培養箱中培養 30 分鐘,之後取出 500 μL 的菌液至裝有 125 μL 甘油 (glycerol) 的微量離心管中,並放置在-20℃下保存。接著以微量吸管 管嘴尖端沾取少量菌液塗抹至含有 1.5% NaCl 的 TSA 培養基中,再將該 培養基置於 28℃恆溫培養箱中隔夜培養。接著重複上述動作,再次添加抗 菌胜肽至原先的菌液中,使菌液中的抗菌胜肽濃度達到原先的 2 倍,每次 繼代都增加兩倍的抗菌胜肽濃度,共繼代 10 次 (表 4.1)。最後,從最高繼 代次數的培養基中挑選單一菌落,並測定其最小抑菌濃度,倘若其最小抑 菌濃度比原始菌株的大,則視為抗性菌株。
表4.1. 每代菌液中所含有的抗菌胜肽之最終濃度。
Table 4.1. The antimicrobial peptides concentration of every passages.
The numbers of subculture AMPs
Ⅰ Ⅱ Ⅲ Ⅳ Ⅴ Ⅵ Ⅶ Ⅷ Ⅸ Ⅹ
Q4 2 4 8 16 32 64 128 256 512 1024
Q6 2 4 8 16 32 64 128 256 512 1024
H1 2 4 8 16 32 64 128 256 512 1024
Pleurocidin 4 8 16 32 64 128 256 512 1024 - Unit:μg/mL
四、蛋白質萃取與分劃 (Protein extraction and fractionation)
在獲得抗性菌株之後,接著就要利用蛋白質體學的方法分析抗性菌株 與野生型菌株在內膜與外膜蛋白表現上的差異。內膜與外膜蛋白的分離方 法乃依據Huang 等人在 2006 年的研究報告,並稍做修飾。主要是利用月桂 醯肌胺酸鈉 (sodium lauryl sarcosinate;SLS) 對內膜蛋白具有高度溶解性,
但是對外膜蛋白則不具溶解性的特性,因此達到將內膜與外膜蛋白分離的 效果 (Filip et al., 1973;Mintz and Fives-Taylor, 2000)。
將隔夜培養後的200 mL 菌液 (OD600值介於1.6-2.0) 添加在 250 mL 的 離心瓶中,利用高速離心機以4,000 xg 離心 15 分鐘,去除上清液並收集細 菌團塊。接著以40 mL 的 0.15 M NaCl 溶液清洗菌體,再重複上述的離心動 作以便收集細菌團塊,總共清洗3 次。之後以 5 mL 的 0.15 M NaCl 溶液懸 浮細菌團塊並轉移至50 mL 離心管。將菌液置於冰上並以超音波細胞破碎 機破壞菌體,輸出頻率為23 KHz,輸出功率為 50 W,以震一秒停一秒的方 式避免震盪時產生高溫,且持續進行 2 分鐘。在超音波破壞菌體之後,利 用高速離心機以5,000 xg 離心 20 分鐘,將細胞殘骸和未被破壞的細菌集中 在離心管管底,因此必需小心地抽取上清液。使用超高速離心機將上清液 以100,000 xg 離心 40 分鐘,離心結束後去除上清液並保留沉澱團塊。添加 3 mL 的 2%月桂醯肌胺酸鈉溶液懸浮沉澱物,並在室溫下靜置 1 小時,使 其充分作用。再次使用超高速離心機將上述樣品溶液以100,000 xg 離心 40 分鐘,離心結束後分離上清液與沉澱物。此上清液為肌胺酸鹽溶解性分劃 (Sarcosinate-soluble fraction;SSF),可將上清液置於冰上以等待進行有機溶 劑沉澱蛋白質的萃取方法或置於-20℃中保存。而沉澱物則是肌胺酸鹽不溶 性分劃 (Sarcosinate-insoluble fraction;SIF)。
五、SIF 及 SSF 的蛋白質萃取
添加100 μL 的復水緩衝溶液 (Rehydration buffer) 懸浮 SIF 團塊,復水 緩衝溶液是由7M 尿素 (urea)、2M 硫脲 (thio-urea)、2% (w/v) CHAPS、2
% (v/v) IPG 緩衝液和 0.002% (w/v) 溴酚藍 (bromophenol blue;BPB) 所 配置而成,並在使用前添加18.2 mM 二硫蘇糖醇 (DL-dithiothreitol;DTT)。
接著將裝有 SIF 溶液的微量離心管管口蓋上透析膜,並以橡皮筋將透析膜 綁在微量離心管管口。利用5,000 mL 的二次水對 SIF 溶液進行微量透析,
透析期間需放置在4℃下,且過程中需換水 4 次。透析結束後可利用桌上型 離心機將蛋白質溶液樣品集中於微量離心管底部,並將微量離心管管口封 上封口膜 (parafilm)。在封口膜上戳洞並放進-80℃冰箱中 10-20 分鐘,使液 態的蛋白質樣品凝固成固態。利用冷凍真空乾燥機將透析過後的 SIF 溶液 乾燥成粉末,冷凍真空乾燥機的作用原理為創造出一個低溫低壓的環境,
在此環境下,樣品內的水份將依據昇華的原理,即由固態不經液態而直接 變成氣態,之後再加以排除,即可達到乾燥的作用。接著添加100 μL 的復 水緩衝溶液回溶SIF 粉末,並進行蛋白質定量。
至於萃取 SSF 的蛋白質之方法,則是利用有機溶劑能夠降低水活性的 原理,使蛋白質親水性區域的水合度降低,因此導致蛋白質聚集在一起而 沉澱的現象。此方法乃依據Huang 等人在 2006 年的研究報告,並稍作修飾。
添加4 mL 的冰丙酮至 15 mL 離心管中,在添加 1 mL SSF 溶液至該離心管 中,使SSF 溶液與冰丙酮充分混合,之後置於-20℃冰箱放置隔夜,使其充 分作用。使用高速離心機以 12,000 xg 離心 20 分鐘,將經由丙酮所沉澱下 來的蛋白質集中在離心管底部,接著去除上清液。之後添加5 mL 冰酒精清 洗 SSF 的沉澱團塊,重複上述離心動作且去除上清液,總共清洗 3-6 次。
將清洗過後的沉澱團塊放置在室溫下風乾,以便去除殘留的酒精。之後添 加200 μL 的復水緩衝溶液溶解乾燥後的沉澱團塊。接著進行微量透析和冷 凍真空乾燥,如之前所述。最後使用200 μL 的復水緩衝溶液回溶 SIF 粉末,
並進行蛋白質定量。
六、蛋白質濃度定量 (Protein concentration determination)
配製牛血清白蛋白 (bovine serum albumin;BSA) 標準溶液,使其濃度
分別為0.2、0.4、0.6、0.8、1.0 μg/mL。添加 200 μL 經 5 倍稀釋的蛋白質分 析試劑於96 孔盤中,再分別添加 10 μL 經連續稀釋的 BSA 標準溶液或待測 蛋白質樣品與上述試劑混合。之後利用微量盤分析儀以595 nm 波長測量上 述混合溶液的吸光值。將連續稀釋的BSA 標準溶液之吸光值繪製成標準曲 線,並求得線性公式。最後將待測樣品的吸光值代入公式,即可求得待測 樣品的蛋白質濃度。
七、二維凝膠電泳 (Two-dimensional gel electrophoresis)
在直流電場中,帶負電的粒子會往正極方向移動,而帶正電的粒子則 往負極方向移動,稱此現象為電泳 (electrophoresis)。二微電泳乃是依據樣 品中蛋白質的等電點 (Isoelectric point;pI) 及分子量皆不盡相同的原理而 達到分離樣品蛋白的效果。
等電焦集法 (Isoelectric focusing;IEF) 乃是在陶瓷容槽 (strip holder) 中加入兩性物質 (ampholyte buffer),當通以直流電時,兩性物質會在由聚 丙烯醯胺膠體組成的等電點電泳膠條 (IPG Drystrip) 內形成一個連續的 pH 梯度,而在膠體內的蛋白質則會朝著與自身所帶電荷相反的電極方向移 動,直到抵達與等電點相同的pH 值處才停止,因此達到依據蛋白質等電點 不同而分離的效果。首先取70 μg 的外膜蛋白樣品或 140 μg 的內膜蛋白樣 品 (體積皆不能超過 125 μL),並添加含有兩性物質的復水緩衝溶液至體積 125 μL。將此蛋白質溶液添加於陶瓷容槽中,然後蓋上 7 公分長度的等電 點電泳膠條,並加入礦物油以覆蓋膠條。將陶瓷容槽移至等電點焦集電泳 儀 (IPGphor) 上,首先給予 30 伏特的低電壓進行復水 (rehydration) 14 小 時,使乾燥的等電點電泳膠條因為吸收樣品溶液而澎潤,此步驟的目的為 讓樣品蛋白及復水緩衝液進入膠條中。在復水程序完畢後,接著給予300、
500、1,000 伏特的電壓,分別跑 0.5 小時,最後再提升到 5,000 伏特之後累 積13,400 伏特小時,此步驟為讓蛋白質跑到等電點位置。
在等電點焦集電泳結束後,將膠條浸泡在額外添加1% (w/v) DTT 的平