行政院國家科學委員會專題研究計畫 成果報告
創傷弧菌 Vibrio vulnificus 菌體聚集因子於血清殺菌蛋 白的抗性角色
研究成果報告(精簡版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 95-2320-B-041-002-
執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 嘉南藥理科技大學生物科技系(所)
計 畫 主 持 人 : 陳昱仲
計畫參與人員: 碩士班研究生-兼任助理:鍾雅婷
處 理 方 式 : 本計畫涉及專利或其他智慧財產權,2 年後可公開查詢
中 華 民 國 96 年 12 月 03 日
目錄
前言 --- 2
目的 --- 4
研究方法 --- 5
結果與討論 --- 7
參考文獻 --- 10
成果自評 --- 13
研究計畫內容:
前言:
創傷弧菌(Vibrio vulnificus)是一株常存在海產食品的高致死率的致病菌,近年來有許多 國家也出現相當多的病例,包括美國、墨西哥、韓國及日本等(Bisharat et al., 1999; Hlady and Klontz, 1996; Paik et al., 1995),此菌的感染途徑常常是經由傷口侵入或由飲食造成感染,感 染後所造成的臨床症狀主要有三種:1.原發性敗血症(primary septicemia):其感染原因主要 是食入生的或未煮熟的海鮮類食品,創傷弧菌經由腸胃道而感染,臨床上病人會有發熱、寒 顫、嘔吐等症狀,常常導致菌血症及敗血性休克,其死亡率高達 50%以上(Blake et al., 1979;
Klontz et al., 1988; Chuang et al., 1992; Shapiro et al., 1998),原發性敗血症也常常伴隨著肌肉 軟組織及皮膚的壞死(Klontz et al., 1988; Chuang et al., 1992),除此之外,且此菌感染發病病 程相當快,往往發病 2 天後患者會休克,進而導致死亡(Blake et al., 1979; Klontz et al., 1988;
Chuang et al., 1992)。2.傷口感染(wound infection):通常是由於傷口直接碰觸含有創傷弧菌 的海水或海產食品,或是由於處理海產食品時遭刺傷而感染,感染部位會有紅腫及水泡,之 後很快造成組織壞死、潰爛或嚴重的蜂窩性組織炎,病程若惡化會發展成嚴重的次發性敗血 症而導致死亡,死亡率約為 25% (Blake et al., 1979; Klontz et al., 1988; Chuang et al., 1992;
Shapiro et al., 1998)。3. 腸胃炎:此症狀較輕微,常導致腹痛及下痢,往往會自然復原。創 傷弧菌的感染宿主的特異性一直是造成感染的主要因素,其感染的宿主往往具有潛在的慢性 疾病包括肝病或免疫系統低弱,都是容易感染的宿主(Tacket et al., 1984)。近來,美國疾病管 制中心指出,自 1996 至 2002 由於弧菌感染的病例已增加了 126% (Centers for Disease Control and Prevention),而台灣的臨床病例在近年也不斷地持續增加(Chuang et al., 1992; Chang et al., 1994; Hsueh et al., 2004)。
創傷弧菌的致病機制目前仍然尚未清楚,除了夾膜(Yoshida etal., 1985)以及我們所找到 的鉀離子吸收蛋白(TrkA; Chen et al., 2004)以外,仍未找到致病力強的致病因子。在菌體夾 膜方面,夾膜的多寡和創傷弧菌的致病力有直接的關係,缺少夾膜的菌株對小老鼠的致死力 遠小於野生株,致死菌量相差~10
4
(Yoshida et al., 1985)。最近研究報告指出,有關夾膜合成 與分泌過程所參與的基因都和創傷弧菌的致病力有關,不管是夾膜合成的基因或夾膜分泌通 道的基因(wza),任何一個基因被破壞均會使其對老鼠的致死率大大下降(Zuppardo and Siebeling, 1998; Wright et al., 1999)。在鉀離子吸收蛋白方面,我們研究報告指出,鉀離子吸 收蛋白 TrkA 除了在具有鉀離子吸收的功能外,它還是使菌株具有殺菌蛋白 polymyxin B、protamine 及血清殺菌作用的抗性能力,此基因缺失的突變株在小鼠感染模式致死力分析顯 示,經由腹腔感染或皮下感染途徑,TrkA 突變株的致死力都遠小於野生株約有~100 倍,顯 見 TrkA 在創傷弧菌感染的重要性(Chen et al., 2004)。
其它可能的致病因子尚有脂解酵素(lipase)、磷酸脂解酵素(phospholipase)、核酸分解酵 素(DNase)、蛋白質分解酵素(protease)、溶血酵素(hemolysin)、細胞毒素(cytolysin) (Kreger and Lockwood, 1981; Olive et al., 1986)和 type IV leader peptidase/N-methyltransferase (Paranjpye et al., 1998),尤其細胞毒素以及蛋白質分解酵素是在創傷弧菌的感染機制研究較清楚的,然而
它們的基因缺失菌株分別進行小鼠的致死性測試,結果它們的基因缺失菌株和野生株對小鼠 的半致死率(LD
50
)並無明顯差別,顯示細胞毒素以及蛋白質分解酵素均不是一個重要的致病 因子(Shao and Hor, 2000; Wright and Morris, 1991)。致病菌常常可在感染部位經由微血管侵犯到其它身體組織、器官,由於血液血清具有 殺菌作用,常常是避免細菌感染相當重要的防禦機制,因此致病菌都具有許多抵抗血清殺菌 作用的系統,這些抗血清殺菌作用的系統目前較明確的有莢膜(capsule),它可以避免血清殺 菌作用,以達到感染人體的目的。而血清之所以具有殺菌作用是因為含有一群抗菌蛋白 (cationic antimicrobial peptides) , 如 補 體 (complement) 、 bactericidal/permeability-increasing protein (BPI)及 defensin (Levy, 2000),這些抗菌蛋白的殺菌機制主要是抗菌蛋白會插入細胞 膜形成孔洞,導致菌體瓦解而死亡(如下圖)(Hankcock et al., 1995; Hankcock and Chapple, 1999),除此之外,在皮下組織、上皮細胞及免疫細胞如巨噬細胞(macrophage)及白血球 (neutrophil)也會分泌許多抗菌蛋白,這類型的抗菌蛋白包括有 defensins (Harder et al.,1997), azurocidin (Shafer et al., 1984; Pohl et al., 1990), melittin, magainins, polymyxin B, protamine sulfate, polylysin 及 cecropins (Vaara, 1992)。目前有些抗菌蛋白包括 ambicin (nisin), gramicidin S, polymyxin B 在 臨 床 或 商 業 上 已 被 使 用 , 然 而 創 傷 弧 菌 對 polymyxin B 或 serum complement 已具有抗性,其對這些抗菌蛋白的抗性機制目前仍然未清楚。
Model for antimicrobial peptide activity
目前致病菌對抗生素或抗菌蛋白的抗藥性機制雖然尚未清楚,近來已陸續有相關研究報 告指出,致病菌會受外在環境的不同而改變菌體性質,使其本身在惡劣的環境下繼續生存下 來而持續感染生物體(Dziejman and Mekalanos, 1995),在有關 Salmonella typhimurium 的研究報 告指出,此菌之所以具有抗菌蛋白(polymyxin B)抗性主要是因為環境調節因子(Environmental response regulator; PhoP)在抗菌蛋白的環境下會進行菌體性質的改變,使菌體具有抗菌蛋白的 抗性(Véscovi et al., 1996; Wősten et al., 2000),相似的研究結果也在 Pseudomonasa aeruginosa 發現,此菌的外膜蛋白 outer membrane H1 會受環境的影響而增加表現量,同時也會增強對抗 菌蛋白 polymyxin B 的抗性(Nicas and Hancock, 1980)。有關於菌體聚集而導致增加抗藥性的研 究報告甚少,目前大多僅限於菌體聚集現象的研究,如 Helaine 等指出在 Neisseria meningitidis 具有一個與纖毛蛋白 pilin 相似的蛋白叫 PilX,PilX 對於菌體的聚集是必要的因子,而且 PilX
也會促進菌體利用纖毛附著於人體細胞(Helaine et al., 2005),但對抗藥性的關連並無著墨;對 於菌體聚集而增加菌體抗藥性的研究方面,目前較清楚的主要是在菌體生物膜的形成而增加 抗藥性,Drenkard 及 Ausubel 在其研究報告指出 Pseudomonas 對抗生素抗藥性主要可能是菌 體受到含有抗生素環境,藉由調節因子改變菌體的特性,包括菌體外形的改變,使得細菌轉 變為易形成生物膜(biofilm)的菌體,導致菌體對抗生素抗藥性的增加(Drenkard and Ausubel, 2002),顯示菌體形成生物膜極可能是致病菌對抗生素或抗菌蛋白產生抗藥性的機制之一。另 外,最近的研究報告也指出,在高抗藥性菌株 Enterococcus faecalis 的表面具有一個特殊的聚 集蛋白(aggregation substance; AS),此蛋白會使 E. faecalis 產生菌體聚集現象,並且會使菌體 產生抗藥性(Waters et al., 2004),由這些研究結果顯示,形成菌體聚集的機制似乎與細菌能抵 抗抗生素或抗菌蛋白的殺菌作用有著緊密關係。
目的:
創傷弧菌的致死率高、感染病程快速且其致病機制目前仍然尚未清楚,除了夾膜(Yoshida et al., 1985)以及我們所找到的鉀離子吸收蛋白(TrkA; Chen et al., 2004)以外,仍未找到致病力 強的致病因子做為往後在治療創傷弧菌感染的標的。創傷弧菌是株具有很強侵入能力的致病 菌,常經由皮下血管擴散至全身引起敗血症,由於創傷弧菌的致死性高,而且此菌易於含有 抗菌蛋白的血清中生長,因此創傷弧菌必然至少有一套抗殺菌的機制,使其能在人體內不斷 增值而造成感染。
先前的研究結果已知創傷弧菌具有抗菌蛋白 polymyxin B 及血清殺菌作用的抗性(Chen et al., 2004),因此,我們以抗菌蛋白 polymyxin B 處理菌體做為觀察菌株變化的模式,大部分來 自臨床的菌株經由 polymyxin B 處理後結果發現,大部分菌株是容易形成菌體聚集現象,而 且菌體並沒有瓦解破裂而死亡,相對地,無菌體聚集現象的菌株後來是較容易破裂死亡,並 且這些無菌體聚集現象的菌株幾乎都是屬於來自環境的菌株。在最近的研究報告也指出,經 過抗菌蛋白-乳鐵蛋白 lactoferrin 處理的 Staphylococcus aureus 會導致菌體外觀的改變且會使 得菌體發生聚集現象(Diarra et al., 2003),因此,這種菌體聚集現象或許是創傷弧菌之所以能 適應在血液血清中而不被抗菌蛋白殺死以及避免遭受免疫細胞吞噬殺死作用,而順利在人體 內迅速增值的重要機制之一。
另一方面,台灣是個海島型國家養殖業相當興盛,生食海產食品的人口也不段上升,加 上台灣肝病患者比率相當高,肝病患者是創傷弧菌感染最好發的寄主,近來,美國疾病管制 中心指出,自 1996 至 2002 由於弧菌感染的病例已增加了 126% (Centers for Disease Control and Prevention),而台灣的臨床病例在近年來也不斷地持續增加(Chuang et al., 1992; Chang et al., 1994; Hsueh et al., 2004),因此,對全球及台灣而言,解決創傷弧菌感染的課題卻是刻不容緩。
綜合以上所言,我們期望繼續(I)藉由轉位子基因突變株庫,以菌體聚集分析實驗持續篩 選更多的菌體聚集缺失突變株並進一步選殖菌體聚集基因,同時(II)以抗菌蛋白 polymyxin B 及血清抗性實驗進一步分析野生株及其菌體聚集缺失突變株在對抗菌蛋白與血清抗性能力的 差異,以了解菌體聚集相關基因在抗菌蛋白與血清抗性作用的角色,期望能尋找出有效的致 病因子當作設計殺菌藥物的標地蛋白,能更有效更快速殺死創傷弧菌或抑制其快速生長以達
到治癒效果,並進一步尋找有效抗原蛋白以製備疫苗,以期能達到預防創傷弧菌感染的目的,
並提供研究人員對其他致病菌在抗生素或抗菌蛋白產生抗性的可能機制的瞭解,能在致病菌 的感染控制有所助益。
研究方法:
臨床菌株的收集
由成功大學醫學院附設醫院收集臨床病株。
菌株的培養
E. coli DH5α與 Vibrio vulnificus 均以 LB 培養基或培養液進行菌株培養,培養溫度在 37℃下培養。
菌體聚集性(bacterial aggregation)測試
方法:2 ×10
7
CFU/ml 的轉位子突變株於含有 polymyxin B 的 LB 培養液,於 37℃下培 養 2 小時,觀察菌體聚集現象。菌體血清抗性(serum resistance)測試(Chen et al., 2004)
方法:50 μl 轉位子突變株菌液(2 ×10
5
CFU/ml PBS)和 50 μl 的 serum 或熱去活 化的 serum(56℃, 30 min)均勻混合下,於 37℃下培養 2 小時,取固定量在不 同稀釋濃度分析菌體的存活率。菌體於殺菌蛋白抗性(antimicrobial peptide resistance)測試
方法:50 μl 轉位子突變株菌液(2 ×10
5
CFU/ml PBS)和 50 μl 的不同濃度的殺菌 蛋白溶液均勻混合下,於 37℃下培養 2 小時,取固定量在不同稀釋濃度分析菌 體的存活率。南方轉漬法(Southern blotting)
方法:將轉位子突變株染色體以 2~3 種限制酵素進行切割後,再進行電泳分離後轉漬 至硝化纖維膜,以抗生素 kanamycin 抗性基因作為 DIG 標誌的探針,觀察轉位 子在轉位子突變株染色體的數目及分佈狀況。
選殖菌體聚集相關基因
方法:聚集缺失突變株的染色體以限制酵素進行切割,同時以相同的限制酵素進行切 割 pUC19 以做為載體,並以 E. coli DH5α做為質體寄主,在 kanamycin 50μg/ml 的培養基篩選轉殖株,進行質體抽取後以轉位子序列設計的引子進行轉位子插 入點的核酸序列定序分析,以得知聚集相關基因的片段核酸序列。以此聚集相 關基因的片段核酸序列做為 DIG 探針,以菌落雜交法(colony hybridization)進行
創傷弧菌基因庫的選殖,並進一步進行序列分析以獲得菌體聚集基因的完整核 酸序列。
菌體聚集基因的序列分析
方法:將菌體聚集基因利用 BLAST (Altschul et al., 1997)分析其蛋白質序列。
菌體聚集基因回復實驗
方法:以引子利用 PCR 將完整的菌體聚集基因擴增,DNA 片段經限制酵素切割後以 pBC 做為載體,利用電穿孔方式(McDougald et al.,1994)將血清抗性基因轉入菌 體聚集基因剔除的突變株,以抗生素 chloramphenicol 5 μg/ml 及 rifampin 50 μ g/ml 培養基進行篩選基因回復菌株,以 PCR 及質體抽取分析基因回復結果。
選殖菌體聚集相關基因
方法:聚集缺失突變株的染色體以限制酵素進行切割,同時以相同的限制酵素進行切 割 pUC19 以做為載體,並以 E. coli DH5α做為質體寄主,在 kanamycin 50μg/ml 的培養基篩選轉殖株,進行質體抽取後以轉位子序列設計的引子進行轉位子插 入點的核酸序列定序分析,以得知聚集相關基因的片段核酸序列。以此聚集相 關基因的片段核酸序列做為 DIG 探針,以菌落雜交法(colony hybridization)進行 創傷弧菌基因庫的選殖,並進一步進行序列分析以獲得菌體聚集基因的完整核 酸序列。
菌體聚集基因的序列分析
方法:將菌體聚集基因利用 BLAST (Altschul et al., 1997)分析其蛋白質序列。
菌體聚集基因缺失菌株之構築
方法:構築的方式如同我們之前所發表的方法(Chen et al., 2004),以質體插入法利用一 次同源性互換將目的基因予以破壞,首先將欲破壞的基因片段接合到帶有抗生 素 kanamycin 抗性基因的質體後轉型送入 E. coli,利用接合生殖的方式,將此質 體帶入具有抗生素 rifampin 抗性的創傷弧菌(V. vulnificus),在創傷弧菌內會進行 同源性插入欲破壞的基因,產生特定基因缺失的突變株,這些突變株可藉在含 有抗生素 kanamycin 50 μg/ml 及 rifampin 50 μg/ml 的培養基予以篩選之。接 著以聚合酵素鏈反應(PCR)與南方轉漬法(Southern blotting)確認基因剔除突變 株。
菌體聚集基因回復實驗
方法:以引子利用 PCR 將完整的菌體聚集基因擴增,DNA 片段經限制酵素切割後以 pBC 做為載體,利用電穿孔方式(McDougald et al.,1994)將血清抗性基因轉入菌 體聚集基因剔除的突變株,以抗生素 chloramphenicol 5 μg/ml 及 rifampin 50 μ
g/ml 培養基進行篩選基因回復菌株,以 PCR 及質體抽取分析基因回復結果。
結果與討論:
(一) 創傷弧菌轉位子 Tn5 突變株庫的構築
我們已完成創傷弧菌轉位子 Tn5 突變株庫的構築,共收集有 6,548 株的轉位子突變 株。由 Southern blotting 分析得知,這些突變株幾乎全部都只有單一轉位子插入,而且都 不是重複複製的突變株(如 Figure 1)。
Figure 1 Analysis of transposon-integrated patterns of randomly selected V. vulnificus transposon mutants by Southern blotting.
(二) 菌體聚集形成形成能力之分析
我們已由 2,150 株轉位子突變株中篩選出十一株菌體聚集形成能力降低的突變株 ADM (Aggregation-Defective Mutant)分別為 ADM-1 ~ ADM-11,這些突變株的菌體聚集 形成的現象與能力如 Figure 2 及 Table 1,Figure 2 顯示,野生株在抗菌蛋白的處理下,
菌體會形成聚集現象,然而,菌體聚集形成能力降低的突變株則不會形成菌體聚集的現 象。
Figure 2 Aggregation analysis of V. vulnificus ADM-1 strain and wild type after treatment of antimicrobial peptide polymyxin B
1A9 1A8 1A7 1A6 1A5 1A4 1A3 1A2 1A1
ADM-1 Wild type
(5 μg). Arrow showed aggregative bacteria.
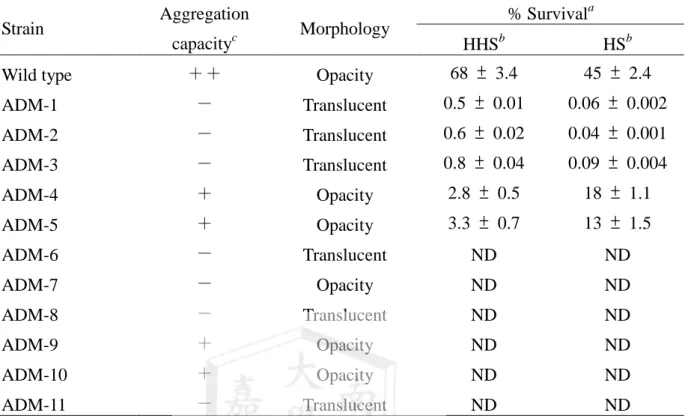
Table 1. Percentage of survival on exposure of V. vulnificus to complement heat-inactivated and non-heat-inactivated 50% normal human serum and the morphology and aggregation capacity of these strains
% Survival
a
Strain Aggregationcapacity
c
MorphologyHHS
b
HSb
Wild type ++ Opacity 68 ± 3.4 45 ± 2.4
ADM-1 - Translucent 0.5 ± 0.01 0.06 ± 0.002 ADM-2 - Translucent 0.6 ± 0.02 0.04 ± 0.001 ADM-3 - Translucent 0.8 ± 0.04 0.09 ± 0.004
ADM-4 + Opacity 2.8 ± 0.5 18 ± 1.1
ADM-5 + Opacity 3.3 ± 0.7 13 ± 1.5
ADM-6 - Translucent ND ND
ADM-7 - Opacity ND ND
ADM-8 - Translucent ND ND
ADM-9 + Opacity ND ND
ADM-10 + Opacity ND ND
ADM-11 - Translucent ND ND
a
The data corresponds to a representative experiment performed in duplicate. Data is given as the mean of survival relative to the original inoculums ± standard error of the mean.b
HHS, heated-inactivated human serum; HS, non-heated-inactivated human serum.c
++: bacteria aggregation (Strong); +: bacteria aggregation (Weak); -: bacteria lysis 這些突變株菌體聚集形成能力大小為 WT > [ADM4 ~ ADM5 ~ ADM9 ~ ADM10 ] >[ADM1 ~ ADM2 ~ ADM3 ~ ADM6~ADM-7~ADM-8~ADM-11]。
(三) 菌體聚集缺失突變株之外觀型態
突變株的外觀在 LB 固態培養基培養後觀察,結果如 Table 1 所示,菌體外觀呈現透 明(translucent) 的突變株是屬於夾膜缺失的菌體,如 Table 1 所示的 ADM-1、ADM-2 、 ADM-3、ADM6、ADM-8 與 ADM-11,相反地,外觀不透明(opacity)的突變株則是屬於 具有夾膜的菌體,如表 2 所示的 WT、ADM-4、ADM-5、ADM-7、ADM-9 與 ADM-10;
由 Table 1 顯示,菌體聚集生成的能力似乎與菌體有無夾膜(capsule)有關,形成菌體聚集 能力較差的突變株 ADM-1、ADM-2、ADM-3、ADM-6、ADM-8 與 ADM-11,菌體都呈 現透明(translucent)的外觀。
較特別的現象是 ADM-4 與 ADM-5 兩株菌株,雖然在 LB 培養基培養 48 小時後菌 落外觀呈現不透光性(opacity) (Figure A), 然而,菌體在培養初期,其夾膜的形成呈現不 均勻現象,夾膜於菌落表面呈現輻射狀(Figure B),此現象卻不同於野生株。
目前,我們正著手由 ADM-1、ADM-2、ADM-3、ADM-4 與 ADM-5 五株突變株選 殖出被轉位子破壞的基因,並進一步分析這些基因具有對殺菌蛋白與血清殺菌作用抗性 的角色;另一方面,我們要由 ADM-4 與 ADM-5 兩株突變株所選殖的聚集基因,進一步 探討在菌體夾膜呈現輻射狀所扮演的角色。
(四) 菌體聚集因子之選殖
目前由 11 株聚集缺失突變株選殖中選殖出 5 個菌體聚集基因,並進一步比對分析 這些聚集基因,由目前研究報告分析其可能的基因功能,分析結果如下表
Table 2. Homology of V. vulnificus ORFs to heterologous bacterial genes
Gene (from ADM) Organism Putative function % Amino acid
identity Accession no.
rmlA (ADM-2) Acitinobacillus
actinomtcetemcomitans
transferase
82 BAA94403
WcvA (ADM-3) V. cholerae O139 Nuclotide-sugar
epimerase 86 AAC46250
wza (ADM-1) E. coli Transport 61 BAE76576
orf1 (ADM-4) Unknown
orf2 (ADM-5) Unknown
(五) 菌體聚集因子於抗血清殺菌作用的角色
將野生株與菌體聚集基因缺失突變株(ADM-1~ADM-5)進行抗血清殺菌作用的比較 結果,如 Table 1 所示,這些菌體聚集基因缺失突變株的抗血清殺菌能力均明顯下降,顯 見這些菌體聚集基因使菌體具有抗血清殺菌作用的能力,其他的菌體聚集基因缺失突變 株仍持續分析中。
參考文獻:
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 25: 3389-3402.
Bisharat, N., Agmon, V., Finkelstein, R., Raz, R., Ben-Dror, G., Lerner, L., Soboh, S., and Colodner, R.
(1999) Clinical, epidemiological, and microbiological features of Vibrio vulnificus biogroup 3 causing outbreaks of wound infection and bacteraemia in Israel. Lancet 354: 1421-1424.
Blake, P.A., Merson, M.H., Weaver, R.E., Hollis, D.G., and Heublein, P.C. (1979) Disease caused by a marine Vibrio: Clinical characteristics and epidemiology. N Engl J Med 300: 1-5.
Bonner, J.R., Coker, A.S., Berryman, C.R., and Pollock, H.M. (1983) Spectrum of vibrio infections in a Gulf Coast community. Ann Intern Med 99: 464-469.
Centers for Disease Control and Prevention. (2003) Preliminary FoodNet data on the incidence of foodborne illnesses-selected sites, United States, 2002. Morb Mortal Wkly Rep 52: 340-343.
Chang, J.J., Sheen, I.S., Peng, S.M., Chen, P.C., Wu, C.S., and Leu, H.S. (1994) Vibrio vulnificus infection: report of 8 cases and review of cases in Taiwan. Chang Gung Med J 17: 339-346.
Chen, Y.C., Chuang, Y.C., Chang, C.C., Jeang, C.L., and Chang, M.C. (2004) A K
+
uptake protein, TrkA, is required for serum, protamine, and polymyxin B resistance in Vibrio vulnificus. Infect Immun 72:629-636.
Chuang, Y.C., Yuan, C.Y., Liu, C.Y., Lan, C.K., and Huang, A.H. (1992) Vibrio vulnificus infection in Taiwan: report of 28 cases and review of clinical manifestations and treatment. Clin Infect Dis 15:
271-276.
Diarra, M.S., Lacasse, P., Deschenes, E., Grondin, G., Paradis-Bleau, C., and Petitclerc, D. (2003) Ultrastructural and cytochemical study of cell wall modification by lactoferrin, lactoferricin, and penicillin G against Staphylococcus aureus. J Electron Microsc 52: 207-215.
Drenkard, E., and Ausubel, F.M. (2002) Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation. Nature 416: 740-743.
Dziejman, M., and Mekalanos, J.J. (1995) Two-component signal transduction and its role in the expression of bacterial virulence factors. In Two-Component Signal Transduction. Hoch, J.A., and Silhavy, T.J. (eds). Washington, DC: American Society for Microbiology Press, pp. 305-317.
Hankcock, R.E.W., and Chapple, D.S. (1999) Peptide antibiotics. Antimicrob Agents Chemother 43:
1317-1323.
Hankcock, R.E.W., Falla, T., and Brown, M. (1995) Cationic bactericidal peptides. Adv Microb Physiol 37: 135-175.
Harder,J.,Barteis,J.,Christophers,E.,and Schrőder,J.M.(1997)A peptideantibioticfrom human skin.
Nature 387: 861.
Helaine, S., Carbonnelle, E., Prouvensier, L., Beretti, J.I., Nassif, X., and Pelicic, V. (2005) PilX, a pilus-associated protein essential for bacterial aggregation, is a key to pilus-facilitated attachment of
Neisseria meningitides to human cells. Mol Microbiol 55: 65-77.
Hlady, W.G., and Klontz, K.C. (1996) The epidemiology of Vibrio infection in florida, 1981-1993. J Infect Dis 173: 1176-1183.
Hsueh, P.R. Lin, C.Y., Tang, H.J., Lee, H.C., Liu, J.W., Liu, Y.C., and Chuang Y.C. (2004) Vibrio vulnificus in Taiwan. Emerg Infect Dis 10: 1363-1368.
Klontz, K.C., Lieb, S., Schreiber, M., Janowski, H.T., Baldy, L.M., and Gunn, R.A. (1988) Syndromes of Vibrio vulnificus infections: Clinical and epidemiologic features in Florida cases, 1981-1987. Ann Intern Med 109: 318-323.
Kreger, A., and Lockwood, D. (1981) Detection of extracellular toxin(s) produced by Vibrio vulnificus.
Infect Immun 33: 583-590.
Kumamoto, K.S., and Vukich, D.J. (1998) Clinical infections of Vibrio vulnificus: a case report and review of the literature. J Emerg Med 16: 61-66.
Levy, O. (2000) Antimicrobial proteins and peptides of blood: templates for novel antimicrobial agents.
Blood 96: 2664-2672.
McDougald, D., L. M. Simpson, J. D. Oliver, and M. C. Hudson. 1994. Transformation of Vibrio vulnificus by electroporation. Curr. Microbiol. 28:289-291.
Mead, P.S., Slutsker, L., Griffin, P.M., and Tauxe, R.V. (1999) Food-releated illness and death in the United States reply to Dr. Hedberg. Emerg Infect Dis 5: 841-842.
Nicas, T.I., and Hancock, R.E.W. (1980) Outer membrane protein H1 of Pseudomonas aeruginosa:
involvement in adaptive and mutational resistance to ethylenediaminetetraacetate, polymyxin B, and gentamicin. J Bacteriol 143: 872-878.
Oliver, J.D., Wear, J.E., Thomas, M.B., Warner, M., and Linder, K. (1986) Production of extracellular enzymes and cytotoxicity by Vibrio vulnificus. Diagn Microbiol Infect Dis 5: 99-111.
Paik, K.W., Moon, B., Park, C.W., Kim, K.T., Ji, M.S., Choi, S.K., Rew, J.S., and Yoon, C.M. (1995) Clinical characteristics of ninety-two cases of Vibrio vulnificus infections. Kor J Infect Dis 27:
355-365.
Paranjpye, R.N., Lara, J.C., Pepe, J.C., Pepe, C.M., and Strom, M.S. (1998) The type IV leader peptidase/N-methyltransferase of Vibrio vulnificus controls factors required for adherence to Hep-2 cells and virulence in iron-overloaded mice. Infect Immun 66: 5659-5668.
Pohl, J., Pereia, H.A., Martin, N.M., and Spitznagel, J.K. (1995) Amino acid sequence of CAP37, a human neutrophil granule-derived antibacterial and monocyte-specific chemotaxic glycoprotein structurally similar to neutrophil elastase. FEBS Lett 272: 200-204.
Reed, L.J., and Muench, H. (1938) A simple method of estimating the 50% endpoints. Am J HyG 27:
493-497.
Shapiro, R.L., Altekruse, S., Hutwagner, L., Bishop, R., Hammond, R., Wilson, S., Ray, B., Thompson, S., Tauxe, R.V., Griffin, P.M., and Vibrio Working Group (1998) The role of gulf coast oysters harvested in warmer months in Vibrio vulnificus infections in the United States, 1988-1996. J Infect Dis 178:
752-759.
Shafer, W.M., Martin, L.E., and Spitznagel, J.K. (1984) Cationic antimicrobial proteins isolated from human neutrophil granulocytes in the presence of diisopropyl fluorophosphates. Infect Immun 45:
29-35.
Shao, C.P., and Hor, L.I. (2000) Metalloprotease is not essential for Vibrio vulnificus virulence in mice.
Infect Immun 68: 3569-3573.
Tacket, C.O., Brenner, F., and Blake, P.A. (1984) Clinical features and an epidemiological study of Vibrio vulnificus infections. J Infect Dis 149: 558-561.
Vaara, M. (1992) Agents that increase the permeability of the outer membrane. Micro Rev 56: 359-411.
Véscovi, E.G., Soncini, F.C., and Groisman, E.A. (1996) Mg2+ as an extracellular signal: environmental regulation of Salmonella virulence. Cell 84: 165-174.
Waters, C.M., Hirt, H., McCormick, J.K., Schlievert, P.M., Wells, C.L., and Dunny, G.M. (2004) An amino-terminal domain of Enterococuus faecalis aggregation substance is required for aggregation, bacterial internalization by epithelial cells and binding to lipoteichoic acid. Mol Microbiol 52:
1159-1171.
Wősten M.M.S.M., Kox, L.E.F., Chamnongpol, S., Soncini, F.C., and Groisman, F.C. (2000) A signal transduction system that responds to extracellular iron. Cell 29: 113-125.
Wright, A.C., and Morris, J.G. Jr. (1991) The extracellular cytolysin of Vibrio vulnificus: inactivation and relationship to virulence in mice. Infect Immun 59: 192-197.
Yoshida, S., Ogawa, M., and Mizuguchi, Y. (1985) Relation of capsular materials and colony opacity to virulence of Vibrio vulnificus. Infect Immun 47: 446-451.
Zuppardo, A.B., and Siebeling, R.J. (1998) An epimerase gene essential for capsule synthesis in Vibrio vulnificus. Infect Immun 66: 2601-2606.
對於學術研究、國家發展及其他應用方面預期之貢獻
目前菌體於抗菌蛋白環境下形成菌體聚集現象而避免抗菌蛋白的殺菌作用之研究仍然 相當少,此研究計畫有助於瞭解細菌於抗藥性的抗性機制,進一步瞭解如何控制細菌的生理 性質,尤其近年來台灣的創傷弧菌感染病例是持續增加且台灣易受創傷弧菌感染的宿主(肝 病患者)相當多,除此之外,創傷弧菌在養殖漁業也常常導致魚類的感染,所以盡快瞭解創 傷弧菌致病的相關因子有助於治療創傷弧菌的感染患者或防範創傷弧菌的感染或避免養殖 業者的損失,以及瞭解創傷弧菌是否利用聚集的現象以利於其存活於海鮮食品的表面,因 此,此研究計畫預期將來能於抗藥性研究、醫學、養殖及食品領域上能有所貢獻。