行政院國家科學委員會補助專題研究計畫 █ 成 果 報 告
□期中進度報告
探討中樞神經系統受傷後活化態星形膠細胞中膠質原纖維酸性蛋白對 於硫化軟骨多醣表現之調控機轉研究
計畫類別:█ 個別型計畫 □ 整合型計畫
計畫編號:NSC 96 - 2321 - B - 038 - 003 - 執行期間: 96 年 8 月 1 日至 97 年 10 月 31 日
計畫主持人:楊維中 共同主持人:李怡萱
計畫參與人員: 劉為麟, 洪愉萱
成果報告類型(依經費核定清單規定繳交):█精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:
臺北醫學大學生物醫學材料研究所中 華 民 國 97 年 11 月 4 日
摘要
星形膠細胞(astrglial cells)為哺乳類動物中樞神經系統中含量很多的細胞形態。研 究發現,中樞神經受傷後會啟動一連串細胞性與分子性的修復機轉,包括星形膠細胞
(astrocytes)的活化以及細胞外基質分子(extracellular molecules)的大量表現,以 重建局部受傷的組織或軸突。而過量的細胞外間質聚集可能形成神經膠疤(glial scar), 會影響神經纖維之再生,並阻隔神經纖維之連結,影響神經訊號的傳導,使神經系統組織修 復失敗。組織免疫染色研究顯示,神經膠疤中富含活化的星形膠細胞表現高量的膠質原纖維 酸性蛋白(glial fibrillary acidic protein, GFAP)與硫化軟骨蛋白多醣(chondroitin sulfate proteoglycan, CSPG)成分分布一致。過量表現的CSPG於活化的星形膠細胞周圍, 是造成抑制神經纖維生長之關鍵物質。我們的研究發現,以初代星形膠細胞體外培養並以 AMPA+cyclothiazide刺激, 可誘發星形膠細胞之再活化,並表現高量的GFAP,且此活化的星 形膠細胞分泌大量的抑制神經纖維生長之CSPG neurocan 與已知體內受傷後星形膠細胞的 表現相似,顯示此模式可作為探討受傷後星形膠細胞之表現及影響。利用干擾RNA於活化的 星形膠細胞抑制GFAP的表現亦抑制了CSPG的表現。暗示,GFAP可能會調控CSPG的表現。此外 若以醣皮酯固醇藥物, Methylprednisolone(MP),能夠抑制GFAP以及CSPG的表現。neurocan 的大量表現可直接受到glucocorticoid receptor的調控,並透過c-Jun作用在neurocan的 promoter上,以調結 neurocan 之 mRNA level,並影響神經細胞纖維的再生。此研究有助 於了解如何藉由調控星形膠細胞的活化抑制神經膠疤的產生。以及找到更有效的促進中樞 神經組織再生及修復之藥物及治療策略。
關鍵詞: 星形膠細胞, 中樞神經系統, 中樞神經受傷, 硫化軟骨蛋白多醣
*本研究部份結果已發表於 Glia, 2008
Liu, W-L, Lee, Y-H, Tsai, S-Y, Hsu, CY, Sun, Y-Y, Yang, L-Y, Tsai, S-H, Yang, W-C V.*.
Methylprednisolone inhibits the expression of glial fibrillary acidic protein and chondroitin sulfate proteoglycans in reactivated astrocytes. Glia, 2008, (56):1390-1400.
Abstract
Astroglial cells are the abundant cell type in central nervous system (CNS). Formation of glial scar following CNS injury, which interferes with the axonal outgrowth and axonal guidance, remains a problem in neuronal regeneration. Immunohistochemistry studies in brain and spinal cord injury (SCI) animals have identified chondroitin sulfate proteoglycan (CSPG) molecules were co-localized with intensely glial fibrillary acidic protein (GFAP)-positive astrocytes in and immediately adjacent to the scar hampering neurite outgrowth and regeneration. Thus, up-regulation of CSPG following CNS injury has been demonstrated. Our previous studies have demonstrated that both mRNA and protein levels of several CSPGs were indeed significantly increased in reactivated astrocytes by AMPA with cyclothiazide excitotoxic induction mimicking injured condition and in rat with SCI.
Treatment with anti-inflammatory glucocorticoid, methylprednisolone (MP) could down-regulate GFAP and the expressions of CSPG. More interesting, the up-regulation of CSPG neurocan could be inhibited by introduction of siRNA to GFAP in reactivated astrocytes, suggesting that the astrocyte cytoskeleton molecule GFAP may regulate the CSPG expression. However, the regulatory
mechanism of GFAP on CSPGs is unclear. This study showed that the expression of CSPG neurocan can be directed regulated by MP. The regulatory mechanism is mediated by glucocorticoid receprot (GR). The responsive promoter region to glucocorticoid was identified. The transcription factor c-jun but not c-fos could bind to the predicted neurocan promoter region and the binding activity was down-regulated by MP treatment in a GR-dependent manner.
Keywords: astrocytes, central nervous system (CNS), chondroitin sulfate proteoglycan (CSPG)
*Part of the data has been published in Glia, 2008
Liu, W-L, Lee, Y-H, Tsai, S-Y, Hsu, CY, Sun, Y-Y, Yang, L-Y, Tsai, S-H, Yang, W-C V.*.
Methylprednisolone inhibits the expression of glial fibrillary acidic protein and chondroitin sulfate proteoglycans in reactivated astrocytes. Glia, 2008, (56):1390-1400.
Introduction
Inhibition of axonal regeneration in the injured central nervous system (CNS) is mediated by axon growth-inhibitory ligands expressed in oligodendrocytes/myelin, reactive astrocytes in the lesion and degenerating tracts, and by fibroblasts in scar tissue. Astrocytes are the most abundant cells in the CNS and involved in a wide range of CNS pathologies, such as trauma, ischaemia, and neurodegeneration. Under the pathological condition, astrocytes may change their morphology and expression of many genes including the typical intermediate filament proteins, glial fibrillary acidic protein (GFAP), vimentin, and nestin. In response to CNS pathology or injury, cellular hypertrophy is a phenomenon referred to as reactive gliosis, which results in astrocyte proliferation and extensive hypertrophy of the cell body as increased production of intermediate filament protein such as GFAP.
The process of astrogliosis has been thoroughly studied in experimental models. The local activation of astrocytes in these models is mediated by cytokines and growth factors including interleukin (IL)-1, IL-6, transforming growth factor--alpha, transforming growth factor--beta1, tumor necrosis factor--alpha, gamma-interferon, basic fibroblast growth factor, etc. A well-known feature of
reactive astrocytes with increased GFAP expression was visualized around CNS injury lesion and the reactive astrocytes process extend towards the injury is a characteristic. However, the function of this reactive gliosis remains incompletely understood.
GFAP is a 50-kD intracytoplasmic filamentous protein that constitutes a portion of, and is specific for, the cytoskeleton of the astrocyte. Therefore, GFAP has been widely used as a marker of astrocytes and as a diagnostic marker for astrocyte derived human neoplasms. In addition, GFAP expression is a key step in the astrocytes differentiation and constitutes the hallmark response of astrocytes to injury. More over, hyperphosphorelation of GFAP and cytoskeleton reorganization were shown in brain injury such as Alzmeimer disease and infection, etc. (Funchal, Dos Santos et al.
2005). The NH2-terminal segment of GFAP has been presumed to participate in the regulatory
functions of these proteins. All four amino acid residues in the NH2-terminal region of GFAP that undergo phosphorylation are located here. GFAP transgenic animal studies have shown that GFAP is involved in complex cellular events such as cytoskeletal reorganization, maintenance of myelination, cell adhesion, and signaling pathways. Mice with GFAP deficiency has been conducted (Eliasson, Sahlgren et al. 1999). Glial scar formation appeared normal after spinal cord or brain lesions in GFAP knock out (GFAP-/-) or vimentin knock out (vimentin-/-) mice, but was impaired in GFAP-/-vim-/- mice that developed less dense scars frequently accompanied by bleeding (Pekny, Johansson et al. 1999), in addition, it suggested that vimentin may compensate the function of GFAP and some degree of functional overlap exists between these intermediate filament proteins. Another study reported that mice deficient in both GFAP and vimentin with a hemisection of the spinal cord have reduced astroglial reactivity associated with increased plastic sprouting of supraspinal axons including the reconstruction of circuits leading to functional restoration (Menet, Prieto et al. 2003). A study conduced in GFAP -/- and Vimetin -/- mice with entorhinal cortex lesions showed that only limited hypertrophy of cell processes with the volume of brain tissue the same as in wild-type mice and improved post-traumatic regeneration (Wilhelmsson, Li et al. 2004).
Traumatic injury in adult mammalian central nerve system (CNS) often results in the proliferation and reactivation of glial cells. The glial cells by secreting increased amount of extracellular matrix (ECM) response to CNS injury during the wound healing process. However, over-react inflammation may cause the secondary injury of the nerve tissue surrounding the lesion site. Since, the glial cells overproduced ECM may form astroglial scar, which block axonal regeneration and cause the impaired neuron function repair. Glial scar consists predominately of reactive astrocytes and proteoglycans where axons that are unable to regenerate past the lesion. The CSPG, neurocan, brevican, and phosphacan are produced by astrocytes in CNS. Proteoglycan, a class of glycoconjugates, is composed of a core protein and at least one glycosaminoglycan (GAG) chain that is covalently attached to the core protein. The GAG chain is a linear unbranched polymer consisting of repeating disaccharide units classified as dermatan sulfate (DS); chondrotin sulfate (CS); heparan sulfate (HS); or keratan sulfate (KS). It has been found that astrocytes can produce the four types of proteoglycans described at above (Johnson-Green, Dow et al. 1991). In the central nerve system (CNS) after injury, CSPGs were found to be up-regulated as a major component of gliotic scar tissues (McKeon, Jurynec et al. 1999; Jones, Margolis et al. 2003; Tang, Davies et al.
2003), which restrict the neurite out-growth (Levine 1994; McKeon, Jurynec et al. 1999; Tanaka, Nogawa et al. 2001; Asher, Morgenstern et al. 2002; Chen, Negra et al. 2002; Chen, Ughrin et al.
2002; Properzi, Asher et al. 2003). A study found the axonal regeneration in the spinal cord can be promoted by degradation of CSPG with chondroitinase ABC (Bradbury, Moon et al. 2002; Yick, Cheung et al. 2003), suggesting that the CS GAG is the potent inhibitor for neurite outgrowth.
However, the core protein of CSPG also possess inhibitory properties (Grumet, Flaccus et al. 1993;
Dou and Levine 1994; Balsamo, Ernst et al. 1995; Lilien, Arregui et al. 1999; Inatani, Honjo et al.
2001).
Materials and methods
Primary culture of rat astrocytes - Briefly, 1-to 2-d-old Sprague-Dawley rat brains were isolated and loosely homogenized in DMEM/F12 medium with 10% fetal bovine serum (FBS), filtered through 80-µm nylon mesh, and centrifuged at 1000 rpm for 10 min. The cells were grown in 75 mm flasks (1.5 brains per flask) until confluent. After separating the microgliaand oligodendrocytesby centrifugation, the tightly attached, highly purified astrocytes with glial fibrillaryacidic protein (GFAP)-positive Immunoreactivity were obtained. The astrocytes were then be trypsinizedand replatedonto 35 mm dishes and incubated with DMEM/F12 medium with 10% FBS in the CO2 tissue culture incubator at 37oC for the following experiments. The astrocytes will be cultured for short (2 – 14 days) and long term (>14 days) to represent the young and aged cells.
Actvation of astrocytes – In the generated C6 gloma cells containing GFAP Tet-On or GFAP Tet-Off expression vector, the expression of GFAP can be tightly controlled by the addition of Dox to over-express GFAP (GFAP Tet-On) or removal of Tc or Dox to turn off the
express of GFAP (GFAP Tet-Off). To stimulate the reactivation of primary cultured astrocytes, the cells were exposed to 200 μM AMPA and 100 μM cyclothiazide for 1 day in vitro (DIV).
MP Treatment of Reactivated Astrocytes- To study the effect of MP on reactivated astrocytes, astrocytes (1 x 105 cell/well) in a 24-well plate were pre-incubated with 0, 0.1, 1, 10, and 50 M MP for 30 min prior to AMPA/cyclothiazide treatment for 48 h. To study the regulatory
mechanism of MP, an antagonist of glucocorticoid receptors, RU486 (100 nM), was added to the reactivated astrocytes along with MP and incubated for 48 h. Each experiment was performed at least in duplicate and was carried out using at least three different astrocyte cultures.
RT-PCR - RT-PCR analysis of total RNA extract was performed using the desire primers.
RT-PCR will be performed using the Dyna RT-PCR system. Reaction components are as described in the manufacturer's protocol. Cycle and quantity titration will be performed to determine the linear range of amplification for each set of primers used. Total RNA will be prepared from cultures by directly lysing cells and mRNAs will be reversed transcribed into cDNA using oligo-dT and Superscript RNAase H-M-MLV reverse transcriptase. Proper amount of primers and 0.5 to 1 g of total RNA extract from the astrocytes cultured in various conditions will be used, and a 20-fold dilution of RNA for the reaction for GAPDH will be used for normalization. The temperature profile for PCR reactions will be tentatively planned. PCR reactions will be performed in a Perkin Elmer-Cetus Gene Amp System 9600. Reaction products will be analyzed on 12% acrylamide-TAE gels (Bio-Rad) and quantitated using Image analysis software. The cDNA will be sent to a biotech company for real-time RT-PCR to confirm the quantitation.
Immunocytochemistry and Immunofluorescent Double Labeling – The cells will be rinsed twice with a control solution (120 mM NaCl, 5.4 mM KCl, 8 mM MgCl2, 1.8 mM CaCl2, 15 mM glucose, pH 7.4) and fixed for 30 min at 37 oC with 4% paraformaldehyde and 1% H2O2 in 20 mM phosphate buffer saline (16.4 mM Na2HPO4, 3.6 mM NaH2PO4, 0.9% NaCl, pH7.5). Fixed cells will be rinsed with 20 mM PBS containing 0.2% Tween 20 (PBST), and incubated sequentially with blocking solution, primary antibody, biotinylated secondary antibody, and avidin-horseradish peroxidase (HRP) reagent as described in the instructions of the Vectastain ABC kit (Vector Laboratories, Burlingame, CA). The anti-MAP-2 monoclonal antibody and the anti-GFAP monoclonal antibody will be properly diluted for primary antibody incubation. Color development of the immunoreactive cells was performed using diaminobenzidine tetrahydrochloride (DAB) in an HRP substrate DAB kit (Vector Laboratories). For colocalization double staining, FITC-conjugated anti-mouse IgG and Texas Red-conjugated anti-rabbit or anti-goat IgG will be used as the secondary antibodies for the respective primary antibodies.
Immunomicrographs were taken using an Olympus IX-70 inverted microscope equipped with fluorescent light source and an Olympus DP50 digital camera. Neuronal population will be determined by immunopositive staining of GFAP-positive cells for astroglial population. Image of colocalization experiment will be performed by superimposing the immunofluorescent image of FITC and Texas Red labelings in an off-line image analysis software (Image Pro-Plus v.5).
Promoter activity assay by luciferase activity- The neurocan promoter (~1 kb) / firefly luciferase constructs were transfected in rat astrocytes using lipofectamine 2000 (Invitrogen).
Standardization was achieved by cotransfection with pBL-null vector (Promega), which encodes renilla luciferase. Cells were harvested 12h and 24hr after transfection. The cells were lysed using Passive Lysis Buffer (Promega). The cells were then scraped from the plates, and debris was removed by a brief centrifugation. An aliquot of the cell lysate was mixed with the luciferase assay reagent Firefly and renilla luciferase activities were measured in the 20/20n Luminometer (Turner Biosystems) using Dual-Luciferase Reporter Assay System (Promega).
Chromatin-immunoprecipitation (CHIP) and PCR - Formaldehyde was added to the culture
medium of astrocytes (4 x 105) in a 35-mm culture dish to form a final concentration of 1%. Cells were incubated at room temperature for 10 min to allow cross-link transcription factors and DNA.
After sonication, chromatin (250 µl) were immunoprecipitated with 1 µl of anti-c-jun or anti-c-fos antibody (10 µg/µl) (Santa Cruz Biotechnology). The isolated DNA was purified by using PCR purification kit (Qiagen) and amplified by PCR by the desired primers.
Immuno-dot Blot
The expression of CS by reactivated astrocytes was analyzed using immuno-dot blots. Total protein (100 μg) from the conditioned media from astrocyte cultures was absorbed onto a PVDF membrane (Millipore) by mild vacuum pumping. Following blocking with 5% skim milk in tris-buffered saline (TBS, 50 mM Tris and 0.5 M NaCl, pH 8.0), the membrane was incubated with a monoclonal antibody against CS (CS56, 1:1000) in TBS with 0.1% skim milk for at least 1 h at 4°C. After three 10-min washes with TBST (TBS with 0.1% Tween-20), the blot was soaked in a solution containing alkaline phosphatase-conjugated rabbit anti-mouse IgG/M (Pierce, Rockford, IL, USA) secondary antibody (1:3000) in TBS with 0.1% skim milk for 1 h at room temperature. After extensive washing with TBST, the blot was incubated with NBT/BCIP for colorimetric development. The resulting dots were scanned for quantitative measurement of the intensity of each immunoreactive dot using image analysis software (Scion).
Western Blot Analysis- GFAP expression was analyzed in astrocyte cell lysate and CSPG expression was analyzed in astrocyte culture medium. GAPDH expression was used as the loading control. To analyze CSPG expression, conditioned medium (500 μl) was pre-incubated with 0.1 unit Chase ABC at 37°C for 3 h and then concentrated by Centricon-100 (Millipore, Bedford, MA, USA) for Western blot analysis. Whole-cell lysate, or an equal amount of total protein from the conditioned media as determined by a protein assay kit (BCA protein assay, Pierce, Rockford, IL, USA), was subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) followed by electroblotting onto a PVDF membrane (Millipore).
After blocking with 5% skim milk in TBS for 1 h, the blot was incubated with a monoclonal antibody against rat GFAP, neurocan (1F6 or 1D1), or phosphacan (3F8) (1:1000) in TBS with 0.1% skim milk for at least 1 h at 4°C. After three 10-min washes with TBST, the blot was soaked in a solution containing an alkaline phosphatase-conjugated goat anti-rat IgG secondary antibody (1:3000) in TBS with 0.1% skim milk for 1 h at room temperature. After extensive washing with TBST, the blot was incubated with NBT/BCIP for colorimetric development. The developed membrane image was scanned, and the intensity of the immunoreactive band was quantitatively measured by image analysis software (Scion).
Transfection of GR siRNA- The primary cultures of rat astrocytes (5 x 105 cells per well in a 24-well plate) were transfected with either control siRNA (a scrambled siRNA from Ambion) or siRNA against rat GR (siGR, sense, 5’-GCUACAGUCAAGGUUUCUGTT-3’, and anti-sense, 5’-CAGAAACCUUGACUGUAGCTC-3’ from Ambion) (10 nM) using Siport reagent (Ambion) in accordance with the manufacturer’s instructions. Forty-eight hours after transfection, cells were stimulated by AMPA+cyclothiazide, with or without MP pretreatment for another 48 h. The cells were then collected for RT-PCR analysis of GFAP and neurocan expression.
Statistical Analysis- Data are presented as the mean ± SD. Statistical analysis was performed by one-way ANOVA. P < 0.05 was considered statistically significant.
Results
MP Suppression of GFAP and CSPG Expression is GR-Dependent
To investigate the regulatory mechanism of MP in reactivated astrocytes, the GR-antagonist RU486 was added to MP-treated reactivated astrocytes and the expression of GFAP, neurocan, and extracellular CSPG was examined. MP-treatment suppressed mRNA (Fig. 1A) and protein (Fig. 1B) levels of GFAP, and this suppression was largely reversed by RU486. This suggests that the effect of MP on GFAP expression is mediated by GR. In addition, reduced expression of
extracellular CSPG by MP treatment in reactivated astrocytes was also reversed by RU486, as revealed by immuno-dot blot analysis (Fig. 2A). Furthermore, MP suppressed expression of neurocan (Fig. 2B) and phosphacan (Figure 2C) at both the mRNA and protein level, and this effect was reversed by RU486. This suggests that the inhibitory effect of MP on extracellular CSPG expression might also be mediated by GR.
It was possible that RU486 was having an effect by interacting with steroid hormone receptors other than the GR (Honer et al., 2003) To determine whether this was the case, we suppressed GR expression by introducing siRNA specific to the GR (siGR) into reactivated astrocyte cultures. We found that MP suppression of GFAP and neurocan expression was reversed by siGR transfection (Fig. 3). This confirmed that the effect of MP on GFAP and neurocan expression was mediated by the GR.
The transcriptional activity of predicted neurocan promoter was up-regulated after AMPA induction and reversely regulated by MP treatment.
Primary culture of astrocytes was transfected with a ~1 kb upstream region of neurocan promoter region followed luciferase reporter gene construct. The promoter activity was investigated in the cells with or without AMPA stimulation and the cells with or without MP pretreatment. The data (Fid. 4) showed that the transcriptional activity of predicted neurocan promoter was up-regulated after AMPA induction and reversely regulated by MP pre-treatment. It suggested that this region contains the responsoive element to MP. Further investigation by a predictive software indicated that the 1 kb upstream of neurocan gene contains several potential related transcription factor binding sites including AP-1 and NF-κB, etc.
MP down-regulated neurocan expression by suppressing c-jun rather than c-fos mRNA expression in a GR-dependent manner
Since c-jun and c-fos proto-oncogenes encode proteins that form a complex which regulates transcription on promoters containing AP-1 activation elements. The effect of MP on c-jun and c-fos mRNA expression in AMPA with cyclothiazide induced reactive astrocytes was investigated.
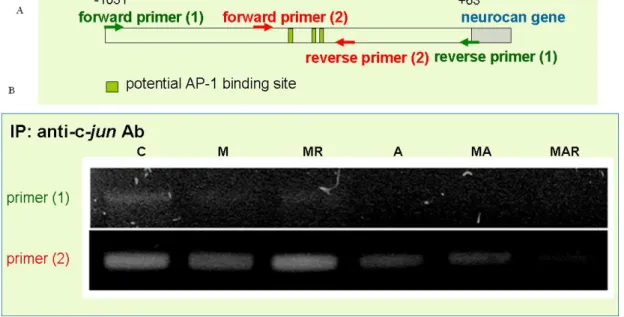
The mRNA level of c-jun but not c-fos was down-regulated by MP treatment in a glucocorticoid receptor-dependent manner. In addition, AMPA with cyclothiazide stimulation had no effect on c-jun and c-fos mRNA expression (Fig. 5). To demonstrate the effect of MP was mediated by the binding of c-jun to neurocan promoter, chromatin IP was carried out by using anti-c-jun antibody to immunoprecipitate c-jun binding DNA, followed by PCR using 2 different pairs of primer derived from neurocan promoter region. Figure 6 showed c-jun indeed bond to the neurocan promoter and the binding activity was down-regulated by MP treatment in a glucocorticoid receptor-dependent manner.
Discussion
MP treatment has been used to effectively reduce post-traumatic inflammation (Xu et al., 1998), which can cause secondary injury after SCI (Hsu and Dimitrijevic, 1990). In this report, we showed that treatment with MP attenuated the increased expression of GFAP and CSPGs in AMPA+cyclothiazide-reactivated astrocytes, suggesting that the neuroprotective effect of MP may be due not only to general reduction of inflammation, but may also be mediated through direct action on astrocytic cells. The effect of MP on reactivated astrocytes appeared to be mediated by GR, since the addition of the GR antagonist RU486 or GR-specific siRNA reversed MP suppression of GFAP and CSPG expression. The GR-mediated effect of MP may involve transcriptional regulation of 5'-upstream sequences of rat GFAP, which contains elements
involved in the GR-mediated response to corticosterone (Rozovsky et al., 1995). A putative AP-1 binding site has been identified in the 5'-flanking region of the mouse and human GFAP genes (Masood et al., 1993; Sarid, 1991). MP-induced downregulation of the inflammation-related transcription factor NF-κB and AP-1 has previously been demonstrated (Xu et al., 1998; Xu et al., 2001). In addition, there are also potential AP-1 and NF-κB binding sites in the 5'-upstream
region of the CSPG neurocan gene, as predicted by using a search engine that identifies
transcription factor binding sites. Therefore, it is possible that MP may regulate the expression of
neurocan through its actions on AP-1 and NF-κB signaling cascades.
Since our previous studies have shown that the up-regulated CSPG such as neurocan could be inhibited by introduction of siRNA to GFAP in AMPA with cyclothiazide induced reactive strocytes, the expression of CSPGs, especially neurocan and phosphacan, the two major inhibitory CSPGs, can be down-regulated by inhibition of GFAP expression in de-activated astrocytes. Our studies demonstrated down regulation of the CSPG neurocan can be modulated directly by GR-mediated MP, not necessary to be mediated by GFAP expression. The expression of CSPG neurocan can be down-regulated by c-jun mediated inflammatory pathway. Therefore, MP suppression on CSPG expression may be not solely mediate by GFAP inactivation. However, GR-independent regulation of the suppression of GFAP and CSPG expression by MP in
AMPA+cyclothiazide-induced reactivated astrocytes cannot be excluded. The effect of MP might also be mediated by its attenuation of excitotoxic activity of AMPA with AMPA receptors.
Astrocytes have functional AMPA receptors (Hosli and Hosli, 1993). Glucocorticoids can attenuate AMPA cytotoxicity by inhibiting AMPA receptor-associated channels in rat cortical neurons (Shirakawa et al., 2005) and in the nucleus of the solitary tract (Shank and Scheuer, 2003). Therefore, the MP-mediated attenuation of AMPA activity that inhibits
AMPA+cyclothiazide-induced astrocytes activation in astrocytes may occur.
In summary, this study suggests that MP treatment compared to non-selective removal of CS by chondroitinase ABC may preserve the better ECM structure that improves neuronal
regeneration and functional recovery after injury.
References
Asher, R. A., D. A. Morgenstern, et al. (2002) "Versican is upregulated in CNS injury and is a product of oligodendrocyte lineage cells." J Neurosci 22(6), 2225-36.
Balsamo, J., H. Ernst, et al. (1995) "The interaction of the retina cell surface
N-acetylgalactosaminylphosphotransferase with an endogenous proteoglycan ligand results in inhibition of cadherin-mediated adhesion." J Cell Biol 129(5), 1391-401.
Bradbury, E. J., L. D. Moon, et al. (2002) "Chondroitinase ABC promotes functional recovery after spinal cord injury." Nature 416(6881), 636-40.
Chen, Z. J., M. Negra, et al. (2002) "Oligodendrocyte precursor cells: reactive cells that inhibit axon growth and regeneration." J Neurocytol 31(6-7), 481-95.
Chen, Z. J., Y. Ughrin, et al. (2002) "Inhibition of axon growth by oligodendrocyte precursor cells." Mol Cell Neurosci 20(1), 125-39.
Dou, C. L. and J. M. Levine (1994) "Inhibition of neurite growth by the NG2 chondroitin sulfate proteoglycan." J Neurosci 14(12), 7616-28.
Eliasson, C., C. Sahlgren, et al. (1999) "Intermediate filament protein partnership in astrocytes." J Biol Chem 274(34), 23996-4006.
Funchal, C., A. Q. Dos Santos, et al. (2005) "Branched-chain alpha-keto acids accumulating in maple syrup urine disease induce reorganization of phosphorylated GFAP in C6-glioma cells." Metab Brain Dis 20(3), 205-17.
Gossen, M. and H. Bujard (1992) "Tight control of gene expression in mammalian cells by tetracycline-responsive promoters." Proc Natl Acad Sci U S A 89(12), 5547-51.
Gossen, M., S. Freundlieb, et al. (1995) "Transcriptional activation by tetracyclines in mammalian cells." Science 268(5218), 1766-9.
Grumet, M., A. Flaccus, et al. (1993) "Functional characterization of chondroitin sulfate
proteoglycans of brain: interactions with neurons and neural cell adhesion molecules." J Cell Biol 120(3), 815-24.
Inatani, M., M. Honjo, et al. (2001) "Inhibitory effects of neurocan and phosphacan on neurite outgrowth from retinal ganglion cells in culture." Invest Ophthalmol Vis Sci 42(8), 1930-8.
Inoue, S. and Y. Inoue (2001) "Developmental profile of neural cell adhesion molecule glycoforms with a varying degree of polymerization of polysialic acid chains." J Biol Chem 276(34), 31863-70.
Inoue, S., S. L. Lin, et al. (2000) "Chemical analysis of the developmental pattern of
polysialylation in chicken brain. Expression of only an extended form of polysialyl chains during embryogenesis and the presence of disialyl residues in both embryonic and adult chicken brains." J Biol Chem 275(39), 29968-79.
Johnson-Green, P. C., K. E. Dow, et al. (1991) "Characterization of glycosaminoglycans produced by primary astrocytes in vitro." Glia 4(3), 314-21.
Jones, L. L., R. U. Margolis, et al. (2003) "The chondroitin sulfate proteoglycans neurocan, brevican, phosphacan, and versican are differentially regulated following spinal cord injury." Exp Neurol 182(2), 399-411.
Levine, J. M. (1994) "Increased expression of the NG2 chondroitin-sulfate proteoglycan after brain injury." J Neurosci 14(8), 4716-30.
Lilien, J., C. Arregui, et al. (1999) "The juxtamembrane domain of cadherin regulates integrin-mediated adhesion and neurite outgrowth." J Neurosci Res 58(6), 727-34.
McKeon, R. J., M. J. Jurynec, et al. (1999) "The chondroitin sulfate proteoglycans neurocan and phosphacan are expressed by reactive astrocytes in the chronic CNS glial scar." J
Neurosci 19(24), 10778-88.
Menet, V., M. Prieto, et al. (2003) "Axonal plasticity and functional recovery after spinal cord injury in mice deficient in both glial fibrillary acidic protein and vimentin genes." Proc Natl Acad Sci U S A 100(15), 8999-9004.
Pekny, M., C. B. Johansson, et al. (1999) "Abnormal reaction to central nervous system injury in mice lacking glial fibrillary acidic protein and vimentin." J Cell Biol 145(3), 503-14.
Properzi, F., R. A. Asher, et al. (2003) "Chondroitin sulphate proteoglycans in the central nervous system: changes and synthesis after injury." Biochem Soc Trans 31(2), 335-6.
Sotelo, J. R., H. Horie, et al. (1991) "An in vitro model to study diabetic neuropathy." Neurosci Lett 129(1), 91-4.
Stocker, G., Z. Drzeniek, et al. (1996) "Proteoglycan synthesis in human and murine
haematopoietic progenitor cell lines: isolation and characterization of a heparan sulphate proteoglycan as a major proteoglycan from the human haematopoietic cell line TF-1."
Biochem J 317 (Pt 1), 203-12.
Tanaka, K., S. Nogawa, et al. (2001) "Activation of NG2-positive oligodendrocyte progenitor cells during post-ischemic reperfusion in the rat brain." Neuroreport 12(10), 2169-74.
Tang, X., J. E. Davies, et al. (2003) "Changes in distribution, cell associations, and protein expression levels of NG2, neurocan, phosphacan, brevican, versican V2, and tenascin-C during acute to chronic maturation of spinal cord scar tissue." J Neurosci Res 71(3), 427-44.
Vogl-Willis, C. A. and I. J. Edwards (2004) "High-glucose-induced structural changes in the heparan sulfate proteoglycan, perlecan, of cultured human aortic endothelial cells."
Biochim Biophys Acta 1672(1), 36-45.
Wilhelmsson, U., L. Li, et al. (2004) "Absence of glial fibrillary acidic protein and vimentin prevents hypertrophy of astrocytic processes and improves post-traumatic regeneration." J Neurosci 24(21), 5016-21.
Yick, L. W., P. T. Cheung, et al. (2003) "Axonal regeneration of Clarke's neurons beyond the spinal cord injury scar after treatment with chondroitinase ABC." Exp Neurol 182(1), 160-8.
Figures and figure ledends
Figure 1. Effects of AMPA plus cyclothiazide (CYZ), methylprednisolone (MP), and RU486 treatment on the expression of glial fibrillary acidic protein (GFAP) in cultured astrocytes at A:
the mRNA level by RT-PCR and B: the protein level by Western blot analysis. The relative amounts of GFAP mRNA and protein in each group were normalized against GAPDH. * Significantly different (P < 0.05) compared to control cells that received no treatment.
Quantitatively measured data represent the mean ± SD of three independent experiments performed in triplicate.
Figure 2. Effects of AMPA plus cyclothiazide (CYZ), methylprednisolone (MP), and RU486 treatment on the expression of extracellular chondroitin sulfate proteoglycans (CSPGs), neurocan, and phosphacan. A: the total amount of secreted CSPGs in the culture medium were assayed by immuno-dot blot analysis, with quantitative data shown at the bottom. * Significantly different (P
< 0.05) compared to control cells that received no treatment. B: neurocan and C: phosphacan expression was determined at both the mRNA and protein levels in reactivated astrocytes cultured using the indicated conditions. The mRNA level was quantitatively measured by RT-PCR.
Quantitative data are presented as the mean ± SD. * Significantly different (P < 0.05) compared to the control. The protein levels of neurocan and phosphacan were analyzed by Western blotting (WB) using the monoclonal antibodies 1F6 (against the neurocan N-terminus), 1D1 (against the neurocan C-terminus), and 3F8 (against phosphacan).
Figure 3. Glucocorticoid receptor (GR) knockdown blocked the inhibitory effect of methylprednisolone (MP) on glial fibrillary acidic protein (GFAP) and neurocan mRNA expression in reactive astrocytes. The inhibitory activity of MP on GFAP and neurocan
expression was significantly blocked by the introduction of siRNA against GR (siGR) in AMPA plus cyclothiazide (CYZ)-induced reactivated astrocytes. Quantitative data are shown in the lower panel. Cultured astrocytes treated with scrambled siRNA served as the control. Open bars, ratio of GR/GAPDH mRNA levels; cross-hatched bars, ratio of GFAP/GAPDH mRNA levels;
black bars, ratio of neurocan/GAPDH mRNA levels. ∗ Significantly different (P < 0.05)
compared to the control. + Significantly different (P < 0.05) compared to AMPA+CYZ-induced cells.
Figure 4. Effects of MP on neurocan promoter activity in primary cultured astrocytes.
Primary culture of astrocytes were transfected with indicated ~1 kbneurocan promoter
region-luciferase constructs. Effect of MP on the promoter activity of neurocan was investigated in the cells with or without AMPA plus cyclothiazide (CYZ) stimulation. The data was shown as mean±SEM.
Figure 5. Effects of MP on the mRNA expression of c-jun and c-fos in primary culture astrocytes.
Total RNA was isolated from the cells cultured at the indicated condition. RT-PCR was carried out to investigate the mRNA expression of c-jun and c-fos, respectively.
Figure 6. Effects of MP on the binding activity of c-jun to neurocan promoter in primary cultured astrocytes. Chromatin IP was carried out in this study by using anti-c-jun antibody to
immunoprecipitated c-jun binding DNA, followed by PCR using 2 different pairs of primer derived from neurocan promoter region as shown in (A), the transcriptional regulation of c-jun on neurocan promoter was shown in (B).