National Taiwan University Master Thesis
Partner Preference and Mating System of Taiwan Field Voles (Microtus kikuchii) – A Look at the Effects of Steroid Hormones
Chia-Chien Lee
Advisor: Yu-Teh K. Lin, Ph.D.
102 6
June 2013
R617
Delia
(
) (partner preference test)
[
] 24
3
4~5 (enzyme-
immunoassay method) (testosterone)
(progesterone) (estradiol) (corticosterone)
(p = 0.04) (p = 0.04)
(M. ochrogaster)
suspected to be monogamous. In monogamous animals, individuals should exhibit a strong social preference for a familiar partner versus a strange one. In this study, we examined the effect of cohabitation (pair-bonding) on the partner preference in Taiwan field voles. In the reciprocal experiments, all individuals were sexually naïve and cohabited with a heterosexual vole for 24 hr prior to the 3-hr partner preference trials.
We collected the feces of Taiwan field voles before and after the partner preference trials, and used the enzyme-immunoassay (EIA) method to analyze the concentration of steroid hormones, including testosterone of males, progesterone and estradiol of females, and corticosterone of all voles. The results showed that the voles did not spend more time in the partner chamber (male: p = 0.33; female: p = 0.098), but the female voles’
preference (p = 0.04) and time that the female voles spent in each chamber (p = 0.04) were both significantly related to if the preferred vole was the partner. In addition, the EIA results indicated that there was no significant relationship between steroid
hormones and partner preference. Also, the sexual dimorphism of the Taiwan field vole in body weight was reduced, and similar to the monogamous prairie vole (M.
ochrogaster). The results indicated that only female voles had partner preference, and
the steroid hormones did not affect the preferences. This study did not support that mating system of the Taiwan field voles is strictly monogamy.
Keywords: Taiwan field vole, mating system, partner preference test, EIA, steroid hormone, sexual dimorphism
... ii
Abstract ... iii
Introduction ... 1
Hypotheses ... 6
Materials & Methods ... 7
Voles and breeding colony ... 7
Sexual dimorphism in body weight ... 7
Partner preference trials ... 8
Testing apparatus ... 8
Partner preference tests ... 8
Hormonal analyses ... 10
Fecal sampling ... 10
Hormone extraction and EIA ... 10
Statistical analyses ... 11
Results ... 13
Sexual dimorphism in body weight ... 13
Partner preference experiments ... 13
Time in each chamber & Time in physical contact ... 13
Prefer or Not prefer ... 14
Time in the neutral chamber ... 14
Effects of hormones on preference ... 14
Discussions ... 16
Partner preferences ... 16
Steroid hormones ... 19
Sexual dimorphism ... 20
References ... 22
Appendix ... 43
Content of Tables
Table 1. Body weight of the Taiwan field vole at 90~180 days of age compared to three other microtine species at 90 days of age. ... 28 Table 2. Contingency table of logistic regression analyses using partner/stranger as
dependent variable and preference as independent variable. ... 29 Table 3. Results of linear regression analyses using time as dependent variable and
partner/stranger as independent variable. ... 30 Table 4. Effects of hormone (progesterone, estradiol, and corticosterone) concentration
of partner/stranger on the outcomes (preferred/non-preferred) of male
preference tests. ... 31 Table 5. Effects of hormone (progesterone, estradiol, and corticosterone) concentration
of partner/stranger on the outcomes (time in each chamber) of male preference tests. ... 32 Table 6. Effects of hormone (progesterone, estradiol, and corticosterone) concentration
of partner/stranger on the outcomes (time in physical contact) of male
preference tests. ... 33 Table 7. Effects of hormone (testosterone and corticosterone) concentrations of
partner/stranger on the outcomes (preferred/non-preferred) of female
preference tests. ... 34 Table 8. Effects of hormone (testosterone and corticosterone) concentration of
partner/stranger on the outcomes (time in each chamber) of female preference tests. ... 35 Table 9. Effects of hormone (testosterone and corticosterone) concentration of
partner/stranger on the outcomes (time in physical contact) of female
preference tests. ... 36
Content of Figures
Figure 1. The concept map of this thesis. ... 37 Figure 2. The partner preference apparatus is 135 × 25 × 50 cm3 in dimension, and
consists of three chambers. ... 38 Figure 3. A standard curve of corticosterone concentration. ... 39 Figure 4. Mean (± 1se) time the focal voles spent in the partner vs. the stranger
chambers during the 180-min partner preference tests. ... 40 Figure 5. Mean (± 1se) time the focal voles spent in physical contact with the partners
vs. the strangers during the 180-min partner preference tests. ... 41 Figure 6. Mean (± 1se) time that Taiwan field voles spent in the neutral chambers
during 180-min preference tests. ... 42
Introduction
A mating system is the way in which an animal society is structured in relation to sexual behavior. Mating systems in mammals can be generally classified as monogamy, polygamy, polyandry, or promiscuity based on the number of mates that each adult individual has (Wittenberger 1979, Clutton-Brock 1989). Monogamy, defined as a long- term association, and essentially exclusive mating relationship, between one male and one female occurs in less than 3% of mammalian species (Kleiman 1977, Wittenberger and Tilson 1980, Carter and Getz 1993). Some examples are common marmosets (Callithrix jacchus, Evans 1983), cotton-top tamarins (Saguinus oedipus, Price and McGrew 1991), and prairie voles (Microtus ochrogaster, Thomas and Birney 1979, Getz et al. 1981). Monogamy has been described in a variety of species exhibiting different life styles. Not a single ecological mechanism could explain the occurrence of monogamy in all mammalian species. However, a list of characteristics has been proposed (Kleiman 1977, Carter and Getz 1993, Carter et al. 1995) to determine if monogamy occurred. In a population with predominantly monogamy, one would observe at least some of the following phenomena:
1. Reduced sexual dimorphism in morphology and behavior (Dewsbury et al. 1980, Heske and Ostfeld 1990, Boonstra et al. 1993, Ostfeld and Heske 1993);
2. Long-term pair bonding persists throughout breeding and non-breeding seasons (Carter et al. 1995);
3. High paternal investment, compared to related species of other mating system (Oliveras and Novak 1986, Solomon 1993, Patris and Baudoin 2000);
5. Display aggressive behaviors by both sexes toward unfamiliar conspecifics for defending the nest and territory (Carter and Getz 1993, Carter et al. 1995, Back et al.
2002);
6. Home ranges overlap between only one male and one female, and range sizes are similar (Gaulin and FitzGerald 1988, Reichard 2003, Wu et al. 2012);
7. Social factors regulate reproductive physiology (e.g., estrus induction and ovulation) (Taylor et al. 1992, Carter et al. 1995).
Forming long-term heterosexual pair bonds is a pivotal characteristic among monogamous animals. Pair bonding represents an intense social attachment between one male and one female (DeVries et al. 1995, Carter et al. 1997). For small mammals, the observation and quantification of pair-bonding could rarely be assessed directly in the field. Researchers often use spacial relationships (home range overlaps) of animals via live-trapping or radio-telemetry to indirectly infer whether there is bonding between one male and one female (Getz et al. 1981, Getz and Hofmann 1986, Jike et al. 1988).
Laboratory studies could supplement field information. Researchers use the partner preference test to investigate whether a pair formed pair-bonding (Pierce and Dewsbury 1991, Williams et al. 1992, Winslow et al. 1993). In addition, genetic analyses,
specifically paternity assignments, of field populations could provide further information on genetic mating system. In a monogamous system, the incidence of multiple paternity should be very low (Wu et al. 2012).
There are two major categories of animal hormones involved in pair bonding:
neuropeptides and steroids. Neuropeptides including oxytocin and arginine vasopressin, have been confirmed to maintain the long-term bonding of male and female mammals (Insel and Hulihan 1995, Insel et al. 1998, Young et al. 1998, Cho et al. 1999, Young and Wang 2004). When the concentration of neuropeptides is high, the focal animal
would show greater partner preference (Williams and Carter 1992, Winslow et al. 1993, Williams et al. 1994, Cho et al. 1999). In contrast, when the receptors of neuropeptides are inhibited, a monogamous animal would not exhibit partner preference (Winslow et al. 1993, Liu et al. 2001).
On the other hand, steroids are synthesized from two classes of endocrine glands, the gonads and adrenal glands. The gonadal steroids, such as progesterone, estradiol, and testosterone, are related to the reproductive behavior and parental care, while the adrenal steroid hormones, such as cortisol, corticosterone, and adrenaline, secreted by adrenal cortex, are related to the homeostasis and stress. Previous studies (Carter et al.
1995, Carter et al. 1997) have shown differential effects of the two classes of steroid hormones on pair bonding of male and female mammals. For example, there was no difference in preference between intact vs. gonadectomized individuals (DeVries et al.
1997). Furthermore, mating could not facilitate partner preference formation in 24-hr or longer cohabitation periods (Williams et al. 1992). Thus partner preferences could develop in the absence of gonadal hormones. In naïve prairie voles, the effects of stress (3-min swimming) or corticosterone injections on partner preferences were sexually dimorphic. In previous studies, removal of the adrenal gland facilitated the development of partner preferences of females, but was followed by failure to form partner
preference of males (DeVries et al. 1995, DeVries et al. 1996).
The prairie voles (Microtus ochrogaster), distributed in midwest North America, have been a model species in the study of the mammalian mating system. The species has been categorized as monogamy (Thomas and Birney 1979, Getz et al. 1981) based on much research in ecology, behavior, physiology, genetics and neurobiology (Getz et
when given the choice of spending time with the paired vs. a stranger individual, often spent much more time with its previous partner (Pierce and Dewsbury 1991, Insel et al.
1995). In contrast, a similar microtine species, meadow voles (M. pennsylvanicus), do not exhibit such a partner preference (Lim et al. 2004). The mating system of the latter species has been categorized as promiscuity (Madison 1980, Gruder-Adams and Getz 1985). Studies (e.g., Lim et al. 2004) on neuropeptides of the prairie voles showed that individuals with higher concentration of oxytocin and vasopressin performed
significantly greater partner preference than other individuals. The densities of oxytocin (OTR) and vasopressin receptors (V1aR) in the ventral forebrain of prairie vole are also significantly higher than other mocrotine species, such as meadow voles (Microtus pennsylvanicus) and montane voles (Microtus montanus) with non-monogamy mating
systems (Insel and Shapiro 1992, Insel et al. 1994, Young et al. 1998, Lim et al. 2004).
On the other hand, in the study of steroid hormones in prairie voles, gonadal steroid hormones had no significant effect on partner preference. Whereas the effects of adrenal steroid hormone, corticosterone, was sex-dependent. It could increase males’ partner preference, but had an opposite effect on the females (Carter et al. 1997).
The Taiwan field vole (Microtus kikuchii) is an endemic species in Taiwan, distributed in high mountains at >2,500 meters in altitude. Past field and laboratory studies have supported the proposition that the mating system of Taiwan field voles is monogamy (Wu 1998, Yang 2011, Wu et al. 2012). The home range sizes of adults did not significantly differ between sexes in different seasons, and more than 70% home ranges showed overlaps with that of only one opposite sex (Wu 1998, Wu et al. 2012).
In addition, microsatellite DNA information suggested that a vole mated exclusively with the same heterosexual individual in a breeding season (Wu et al. 2012). In a partner preference experiment, Chen et al. (2006) found that Taiwan field voles, after a
3-month pairing period, spent significantly more time contacting with the paired partner than with an unfamiliar individual. Yang (2011) showed that male voles would provide direct care that enhanced growth, development and locomotion ability of pups, and improve the survival rate of offspring in a low temperature environment.
Although the above-mentioned studies supported that the Taiwan field vole is monogamous, I think the evidence is inconclusive for the following reasons. First, the home range studies did show overlaps between more than one male and female pairs in some cases (Wu et al. 2012). In addition, the trap spacing was 20-m and trapping interval 1-month in those studies, which may underestimate population density and the degree of home range overlaps among individuals. In fact, Quan et al. (2010) in a 6- week intensive (nearly daily) trapping study found home range overlaps among multiple adult males and females. Second, a study by Liang (2012) showed that trapping at nest sites sometimes capture more than a pair of adults particular during the early breeding season. Third, the partner preference experiment by Chen et al. (2006) used a 3-month pre-trial cohabitation. A long-term cohabitation could generate partner preference even in a non-monogamous species (Parker et al. 2001). In prairie vole, Williams et al.
(1992) showed that 24-hr cohabitation is sufficient to generate partner preference. Four, the neuroanatomy data (Chapell et al. unpublished data) showed that Taiwan field voles had unique receptor expression with similarities (V1aR binding in the lateral septum and ventral pallidum) and differences (OTR binding in the nucleus accumbens, NAcc) to those of prairie voles. OTR binding in NAcc is important for forming pair bonds in female voles, however, the receptor levels were low in Taiwan field voles. Finally, Wu et al. (2012) did not find multiple paternity in Taiwan field voles. Yet, the litter size of
reverse is not necessarily true. Based on those evidences, I suspect that Taiwan field voles may not be monogamous. Additional support is needed. In order to further clarify the mating system of Taiwan field voles, in this study I used partner preference
experiment with 24-hr cohabitation to examine pair bonding.
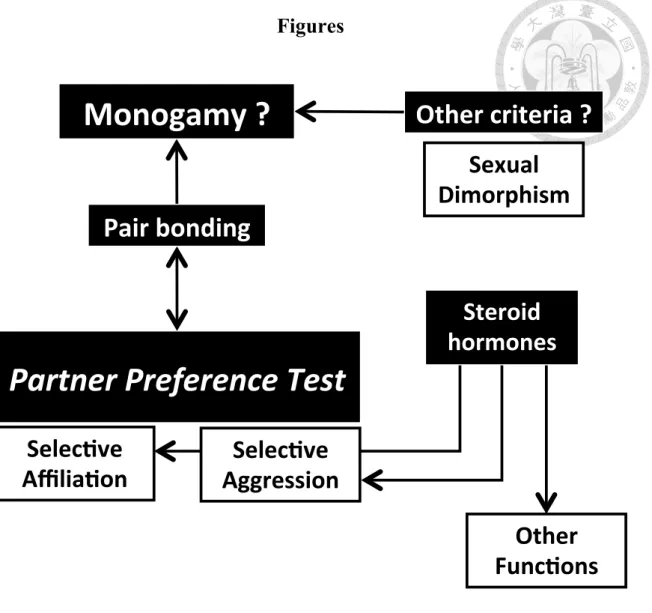
As I mentioned earlier, neuropeptide and steroid hormones may affect pair bonding. Because the effects of those hormones have not been examined in relation to partner preference in the Taiwan field vole, I attempted to examine steroid hormones in this study. Also, I compared the sexual dimorphism in body weight with those of other microtine species (Dewsbury et al. 1980) to further examine the mating system. Thus, the purposes of the study were to combine the partner preference tests with steroid hormone analysis, and use sexual dimorphism to provided new evidences for the mating system of Taiwan field voles. I provide a concept map for this study in Figure 1.
I tested four hypotheses:
1. The Taiwan field voles had weak sexual dimorphism in body weight.
2. The Taiwan field voles showed partner preference in both sexes.
a. The Taiwan field voles spent more time in the partner chamber.
b. The Taiwan field voles spent more time in physical contact with the partners.
3. The gonadal hormones (progesterone and estradiol) of female voles had no effect on partner preferences of male voles; the gonadal hormone (testosterone) of male voles had no effect on partner preferences of female voles.
4. The corticosterone had positive effects on male voles partner preference and negative effects on female voles partner preference.
Materials & Methods
Voles and breeding colony
Taiwan field voles (Microtus kikuchii) used in this study were laboratory-bred F1 or F2 generations that originated from wild stock trapped from an alpine meadow (24°08’36.4”N, 121°17’17.4”E) at the He-huan Mountains. All voles, except breeding pairs, were housed individually in polycarbonate cages (46 × 25 × 20 cm3) with 5-cm- thick aspen chip bedding (TAPVEI) and a 15-cm plastic tube as refuge. The breeding colony was maintained at 17 ± 0.5 °C room temperature, and on a 14:10-hr light/dark cycle (lights on at 0700). Sweet potatoes, rodent chow (LabDiet 5001), and water were provided ad libitum. Pups remained with their parents until 45 days of age, and were housed individually until testing. I recorded baseline information of each vole (sex, body weight, body conditions such as parasitism) weekly and breeding pair (litter size at birth, sex ratio at weaning) in the colony. All experimental voles were sexually naïve, and were tested at approximately 80 days of age.
Sexual dimorphism in body weight
I examined the sexual dimorphism in body weight using the information collected from litters containing at least one male and one female to control for the great variation in body weights among young from different litters (Dewsbury et al. 1980). I used the mean body weight between 90 and 180 days of age of each young, and excluded the weight during the partner preference trials. I obtained information from 8 litters that included 11 males and 10 females.
Partner preference trials
Testing apparatus
The partner preference testing apparatus (Fig. 2) is made of 0.5-cm thick transparent acrylic (135 × 25 × 50 cm3, LxWxH), and consists of three equal-sized chambers (45 × 25 × 50 cm3, LxWxH) divided by the same acrylic material. The two dividers each has a 7 x 7-cm passage hole that allow focal animal to move around chambers freely. At each end of the apparatus, a metal hook fixture allows tether
anchoring. Before each partner preference test, I placed new woodchip bedding and two rodent chow pellets in each chamber. A water-bottle was attached on the wall of each chamber. After each test, the apparatus was rinsed with 70% ethanol and scrubbed thoroughly with water to eliminate odors from previous test.
Partner preference tests
Each trial contained four periods: pre-test (5 days), cohabitation (24 hours), preference test (3 hours), and post-test (4 days) periods. For a male’s partner preference test, one male (focal animal) and two female (one partner and one stranger, stimulus voles) non-related adult voles were included. A female’s partner preference test would follow the same methods except that one female and two male non-related adult voles were included. In the pre-test period, the three voles were housed individually. I collected fecal pellets of each vole daily in the morning for 5 consecutive days. On the fifth morning, the focal male and a randomly chosen female (partner) from the duo were placed in a new cage and cohabitated for 24 hours. The remaining female (stranger) was moved to a new cage alone. On the sixth morning, the three animals would be moved to a behavioral testing room before the partner preference test started.
At the start of test, the partner and stranger were tethered and anchored to the opposite ends of the testing apparatus. The tethered voles acclimated in their respective chambers for 30 min before I introduced the focal vole. The focal vole was placed in the neutral (center) chamber and allowed to move freely in the apparatus. The behaviors of the three voles were video-recorded (Sony HDR-SR12) for 3-hr. At the end of 3 hours, the three voles were separated, and housed individually in the animal colony. Their fecal pellets were collected for the following 4 days in the morning. I condensed each 3- hr video using a 12:1 ratio speed transformation, and watched the full 15-minute videos.
I recorded the following behaviors of the focal voles: time in each chamber, time in physical (side-by-side) contact with each vole, and frequency of aggression toward each vole. An aggressive behavior occurred when the focal vole attacked, bit, or chased a stimulus vole (Ferkin 1988, Williams et al. 1992). I defined preference as a significant difference in time spent by the focal vole in physical contact with the partner vs. the stranger. I felt it’s a more conservative definition than the time spent in the partner vs.
the stranger chamber (Williams et al. 1992, Insel et al. 1995, Insel and Hulihan 1995).
Because the breeding of voles had not been productive, I used a method suggested by Lim et al. (2007) to reduce the number of animals required for trials. In those cases (8 cases for males; 4 cases for females), I performed two partner preference tests in one day, one in the morning and the other in the afternoon. The partner vole from the morning session would serve as the stranger vole in the afternoon, and vice versa. That way, I only needed four, instead of six, animals to complete 2 tests. Lim et al. (2007) showed that there was no measurable test order effect using such a method.
Hormonal analyses
Fecal sampling
I collected fecal pellets of a vole in the bedding during the pre-test and post-test periods. Because fecal pellets produced in a day reflect the physiological condition of a vole in the previous day (Hume et al. 1993, Pei et al. 2001), the fecal samples I
collected represented three periods: pre-test, preference test, and post-test periods. The feces produced on the 1st day post preference test were designated as reflecting the condition on the testing day. All fecal samples were stored at -20 °C before subjected to the analyses of concentration of progesterone, estradiol, testosterone and corticosterone.
Hormone extraction and EIA
All fecal samples were dried at 40 °C for 48 hours and ground into powder. I took 0.1 g of each fecal sample for the subsequent hormone extraction procedures. Each sample was first mixed with 0.8 ml of methanol and 0.1 ml ddH2O. After shaking (1250 xg) a sample for 30 minutes, 0.6 ml petroleum ether was added to each sample. Then, the samples were shaken for another 30 seconds, and centrifuged for 15 minutes at 1600 xg. The solution in the microtube was divided into three layers, from top to bottom, petroleum ether, methanol, and fecal remnant. Next, I transfered 0.6 ml of the hormone- containing methanol layer to a new microtube, and stored it at -20 °C until assayed.
The steroid hormone concentrations of the extracted samples were determined with the enzyme-immunoassay (EIA) method. The 0.6 ml samples were diluted using assay buffer, and mixed with steroid hormone-horseradish peroxidase coupler (HRP).
Then, the mixed solutions were added to a 96-well plate that was previously coated with the steroid hormone (say, progesterone) antibodies. The plate was shaken (100 rpm) for
20~30 minutes. The steroid hormone competed against HRP for antigen-binding sites at room temperature. Then, the plate was washed twice with washing buffer to remove non-binding antigens. Immediately, o-phenylenediamine (OPD) was added to each well, and sit for 20-min color reaction at room temperature. The reaction was stopped by the addition of 8 N sulfuric acid. The absorbency of samples was read with a dual
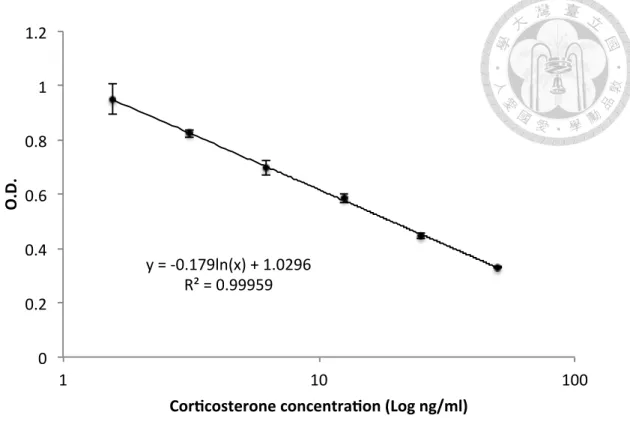
wavelength reader (490/630 nm), and compared with that of the steroid hormone standard curve (Fig. 3, an example of corticosterone EIA results).
Statistical analyses
The three behaviors measured for the focal voles: time in each chamber, time in physical (side-by-side) contact with each vole, and frequency of aggression toward each vole, were analyzed using the Wilcoxon signed-rank test. In addition, I used Student’s t- test to compare the time spent in the neutral (center) chamber by focal males vs. females.
Differences were considered statistically significant when p < 0.05.
I used logistic regression analysis to examine if the choices of focal voles were influenced by the status of stimulus voles (partner or stranger). I used linear regression analyses to examine if the time spent in each chamber and in physical contact with each stimulus voles were influenced by the status of stimulus voles. I used logistic regression analysis to examine if the choices of focal voles were influenced by steroid hormone concentrations of themselves and stimulus voles. I included the concentration in the pairing time and the mean and the coefficient of variation (CV) of concentration before the pairing time. I used linear regression analyses to examine if the time spent in each chamber and in physical contact with each stimulus voles were influenced by hormone
performed power analyses (G*Power 3) to calculate appropriate sample sizes (Faul et al.
2007, Faul et al. 2009).
Results
Sexual dimorphism in body weight
The body weight of adult Taiwan field voles was 43.19 ± 3.38 g in males and 37.03 ± 3.54 in females (Table 1). The difference between adult males and females (6.16 ± 3.18 g) was significant (paired t-test; t = 5.47, p < 0.001). Clearly, there is body weight dimorphism. Compared to three other microtine rodents reported in Dewsbury et al. (1980, Table 1), all 4 species showed sexual dimorphism in body weight with males
heavier than females. The difference in the Taiwan field vole was 6.16 g, or a 16.6%
difference, which was similar to that of prairie voles (M. ochrogaster), and lower than those of montane voles (M. montanus) and meadow voles (M. pennsylvanicus).
Partner preference experiments
Time in each chamber & Time in physical contact
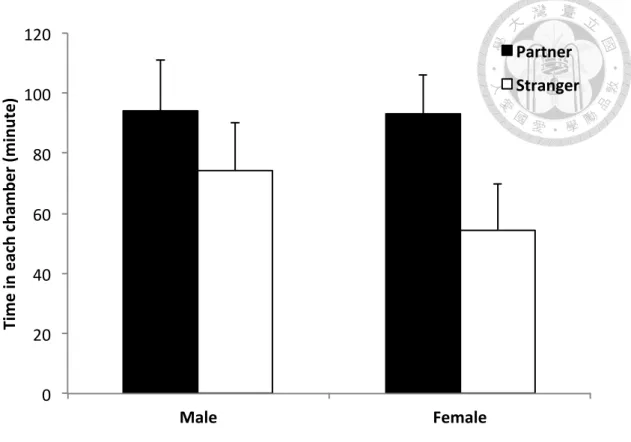
Although both male and female focal voles tended to spend more time in the partner chamber than in the stranger chamber, there was no significant difference
(Wilcoxon signed-rank test: p = 0.33 for males; p = 0.098 for females; Fig. 4). Similarly, both male and female focal voles tended to spend more time in physical contact with the partner than the stranger (Fig. 5). Although there were substantial differences, they were not significant (Wilcoxon signed-rank test: p = 0.18 for males; p = 0.27 for females). I did not observe much aggression between individuals during the trial. The focal females occasionally vocalized to either partners or strangers, but the focal males rarely did so.
Therefore, the aggression was not quantified and analyzed.
Power analyses indicated that one would need 151 and 26 replicates to see
difference in the time spent in physical contact with a stimulus vole in male and female focal voles, respectively.
Prefer or Not prefer
I used the time the focal vole spent in physical contact with the partner vs. stranger to determine which of them was preferred. Six of the 9 focal males chose their partners, while six of the 8 focal females chose their partners. The preference of focal males for partner vs. stranger females was random (Logistic regression; χ2 = 1.40, p = 0.24; Table 2). In contrast, the preference of focal females was not random (Logistic regression; χ2
= 4.22, p = 0.04; Table 2). The preference of focal females was significantly influenced by the status of stimulus voles, i.e., partner or stranger. Using the time in each chamber to define preference yielded the same results.
Linear regression analyses showed that the status of the stimulus voles (partner or stranger) significantly (p = 0.04) affect the amount of time a female focal vole spent in a chamber, whereas the status did not affect male focal voles (p = 0.55; Table 3). The status of the stimulus voles did not affect the amount of time a focal vole spent in
physical contact with a stimulus vole (Table 2; p = 0.27 for males; p = 0.18 for females).
Time in the neutral chamber
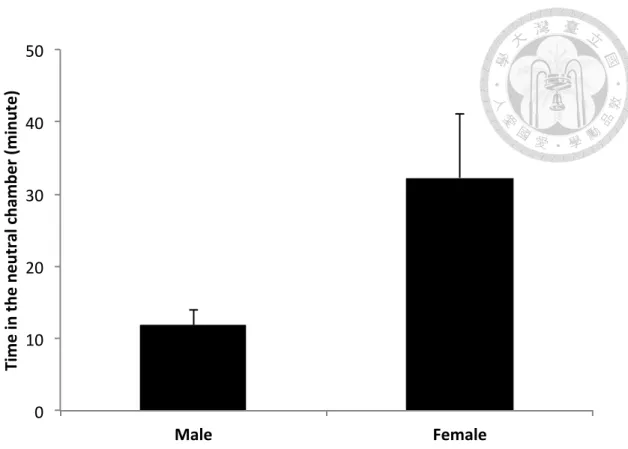
Female focal voles spent significantly more time in the neutral chamber than male focal voles (t-test; t = 2.22, p = 0.03; Fig. 6).
Effects of hormones on preference
I investigated the effects of steroid hormone concentrations on the outcome of preference tests. Three hormone parameters were included: average (over 5 days)
concentration before pairing, variation (CV over 5 days) of concentration before pairing, and concentration during pairing. Steroid hormones concentration of the
partner/stranger voles did not influence the outcome of preference tests of either male (Table 4) or female (Table 7) focal voles. Upon examining details, I found steroid hormones concentration of the partner/stranger voles did not influence the time spent in a chamber or the time spent in physical contact with a stimulus vole by either male (Table 5 and 6) or female (Table 8 and 9) focal voles.
Discussions
1. Partner preferences
The contingency table of partner preference tests indicated that female, but not male, Taiwan field voles showed significant preference for the cohabitation partner (Table 2). What does such a result mean? I can think of three potential explanations.
1. The result is inconclusive, because sample size is small.
2. The result is inconclusive, because cohabitation is too short.
3. The result supports that Taiwan field vole mating system may be polygyny.
In response to explanation 1, I admit the sample sizes were indeed very small.
However, both preference parameters: the variations in the time spent in
partner/stranger chambers and the time spent in physical contact with partner/stranger had substantial variations. Power analyses indicated that the sample sizes required for generating enough power to reject null hypothesis were very high. Nevertheless, I did observe differential variation between males and females. We should consider it a biological reality. In response to explanation 2, there is no empirical study available that indicates the length of cohabitation required for examining partner preference. The only related study (Williams et al. 1992) I know of indicated that 24-hr cohabitation is sufficient to generate the preference for partner in prairie voles. Prairie vole is a
“model” monogamous species. Its mating system has been examined in the field and laboratory in numerous studies (Thomas and Birney 1979, Getz et al. 1981, Carter et al.
1995). I would like to argue that the use of 24-hr cohabitation, a conservative design, should be employed in the partner preference test. My notion was that if Taiwan field voles were as monogamous as prairie voles, we should be able to see partner preference in both males and females (Williams et al. 1992). Finally, I would like to propose explanation 3. I want to consider that the high variations of and differential variations
between males and females are biological reality, i.e., not a fraud due to experimental design. Such experimental results suggested that male Taiwan field voles have a strong potential to mate with multiple females when there is an opportunity. In a sense, the results support polygyny because females remain with one mate, while males do not.
Less than 3% of mammalian species examined show monogamy. Compared to other taxonomic groups, the requirement for females to stay with young (pregnancy and milk-nursing) gives males the opportunity to mate with other females in mammals.
Thus, it takes unique environmental situation for monogamy to occur. Monogamy may occur only when there is no environmental “polygamy potential” or there is no
opportunity to take advantage of what “polygamy potential” the environment affords (Wittenberger and Tilson 1980). For example, when male parental care is necessary for female reproductive success, or when an individual has less fitness with two or more mates than with one, the mating system is likely to show a “monogamy pattern”
(Wittenberger and Tilson 1980). Many ecological factors, such as resource availability and dispersion, mate availability and dispersion, and predation risk may affect mating system (Orians 1969, Kleiman 1977, Emlen and Oring 1977, Wittenberger and Tilson1980). There were many evidences that the mating system of the same species might be different in varied situation, such as habitats, seasons and population density (Getz et al. 1987, McGuire et al.1993, Cushing et al. 2001, Parker et al. 2001). The mating systems of a species may display intraspecific variations in response to different environmental conditions (Lott 1984). For example, the above mentioned monogamous prairie voles may display a polygamous mating system during winter breeding season
Kansas (Fitch 1957, Gaines and Johnson 1984, Swihart and Slade 1989). In contrast, promiscuous meadow voles may develop monogamous-like behaviors selective partner preference and stranger-directed aggression during the colder months of the year or under low population density during the summer breeding season (Madison et al.
1984, Parker et al. 2001). I propose that the mating system of Taiwan field vole is not monogamy, but polygyny. At least, it is strongly flexible depending on environmental conditions. In this study, all experimental voles were housed in the breeding room. They did not suffer stresses from food, weather and predators. Monogamy might not be necessary for Taiwan field voles. Only under some conditions, say circumstances that require bi-parental care, does monogamy arise.
Do field situations push the Taiwan field vole toward a monogamous mating pattern? Nearly all field studies on the ecology of Taiwan field voles came from
populations in a Yushan cane grassland in the He-huan Mountain. In such a habitat, Dr.
Liang-Kong Lin and colleagues found supports for a monogamy mating system (Wu 1998, Chen et al. 2006, Yang 2011, Wu et al. 2012). As mentioned earlier in the
introduction, I was not convinced by the available information. In the most recent study (Yang 2011), they found that paternal care was beneficial for the growth and
development of pups under a cold condition. However, the pup survival did not decrease without paternal care. In contradiction to their studies, my results support a non-monogamy mating system in Taiwan field voles. Three additional studies lend similar supports. Quan et al. (2010) in a 6-week daily trapping study found home range overlaps among multiple adult males and females, although it was in the alpine forest habitat. Second, Liang (2012) showed that trapping at nest sites often capture more than a pair of adults, particularly during early breeding season (March~June). Finally, the
density of receptors for OTR binding in the nucleus accumbens, a positive indication of pair bonding, was low in Taiwan field voles. The level was between those of prairie voles and meadow voles.
2. Steroid hormones
I did not find gonadal hormone concentration affect the outcomes of the partner preference tests of the Taiwan field voles. It conformed to the previous studies shown for the prairie vole (Williams et al. 1992, DeVries et al. 1997). Partner preferences in female prairie voles could be established after a 24 hr-cohabitation, and would not be enhanced further by mating or longer cohabitation periods (48 hr in this study)
(Williams et al. 1992). In both sexes, preferences were not influenced by the presence or absence of gonadal hormones (DeVries et al. 1997). Therefore, partner preferences can develop in the absence of gonadal hormones, and pair bonds could form without sexual experience. Also I did not find significant relationship between the partner preferences and the corticosterone, an adrenal steroid hormone, concentrations in the Taiwan field voles. The corticosterone was a different story on the partner preference of the prairie vole. DeVries et al. (1995, 1996) showed that exposure to the stress of swimming or injections of corticosterone facilitated the development of partner preferences in males, but not females. The response to stress or hormones of the
hypothalamic-pituitary-adrenal (HPA) axis could support the sexually dimorphic effects on preferences. Many studies indicated that the neuropeptides oxytocin and vasopressin could facilitate pair bonding and partner preferences in female and male prairie voles, respectively (Winslow et al. 1993, Williams et al. 1994, Insel and Hulihan 1995). Stress
treatments might inhibit preferences because they interfered with the release or action of oxytocin. It is possible that the non-significant results in my study were due to the use of a different methodology. In previous studies, the corticosterone treatments were removal of the adrenal gland or addition of artificial doses. I used natural fecal corticosterone concentration in this study. The range of variations in corticosterone concentration might not be enough to reveal the effects on pair bonding and resulting partner preferences.
3. Sexual dimorphism
According to a review by Dewsbury et al. (1980), males were significantly heavier than females at 90 days of age in 10 of the 13 muroid rodents, and the other 3 species were considered monogamous. All microtine rodents in their data (M. montanus, M.
pennsylvanicus, M. ochrogaster, and M. canicaudus), and the Taiwan field voles in the
current study, showed sexual dimorphism in body weight. In all cases, males are heavier than females. Among the 5 microtine species, the sexual dimorphism of the gray-tailed vole (M. canicaudus) was the smallest, and males had the small testes (relative to body size) and provided paternal care (Wolff et al. 1994). Although the gray-tailed voles exhibited above-mentioned characteristics of monogamy, the mating system was considered polygamy or promiscuity based on the social organization and the home ranges overlap pattern of the species (Wolff et al. 1994). Unlike the gray-tailed vole, the montane vole (M. montanus) and the meadow vole (M. pennsylvanicus) showed the largest sexual dimorphism, and the mating systems were considered polygamy or promiscuity (Madison 1980, Dewsbury 1981, McGuire and Novak 1986, Boonstra et al.
1993). The sexual dimorphism of the prairie vole (M. ochrogaster) was small, though not the smallest. The species had many characteristics of monogamy and was
considered monogamous during most situations (Thomas and Birney 1979, Getz et al.
1981). Although the Taiwan field vole had a very similar sexual dimorphism as the prairie vole, in light of the mating system of gray-tailed vole, we would need more evidence to establish the mating system of Taiwan field vole as monogamous.
The main purpose of this study had been to investigate the mating system of Taiwan field voles with partner preference test, and examine the effects of steroid hormone concentration on partner preference. I found males did not exhibit significant partner preferences, but females did. The preferences were not influenced by the concentration of steroid hormones. Sexual dimorphism was strong, yet the magnitude was similar to the monogamous prairie vole. In conclusion, my study showed that mating system of the Taiwan field voles is not strictly monogamy.
References
1998 (Microtus kikuchii)
2012
2006 (Microtus kikuchii)
49(1): 65-73
2011 (Microtus kikuchii)
2010 (Microtus
kikuchii)
Back, S. R., Beeler, L. A., Schaefer, R. L. and Solomon, N. G. 2002. Testing functional hypotheses for the behavior of resident pine voles, Microtus pinetorum, toward non- residents. Ethology, 108(11): 1023-1039.
Boonstra, R., Gilbert, B. S. and Krebs, C. J. 1993. Mating systems and sexual dimorphism in mass in Microtines. Journal of Mammalogy, 74(1): 224-229.
Carter, C. S., Devries, A. C. and Getz, L. L. 1995. Physiological substrates of mammalian monogamy: the prairie vole model. Neuroscience and Biobehavioral Reviews, 19(2):
303-314.
Carter, C. S., DeVries, A. C., Taymans, S. E., Roberts, R. L., Williams, J. R. and Getz, L. L.
1997. Peptides, steroids, and pair bonding.!Annals of the New York Academy of Sciences, 807: 260-272.
Carter, C. S. and Getz, L. L. 1993. Monogamy and the prairie vole. Scientific American, 268(6):
100-106.
Cho, M. M., DeVries, A. C., Williams, J. R. and Carter, C. S. 1999. The effects of oxytocin and vasopressin on partner preferences in male and female prairie voles (Microtus
ochrogaster). Behavioral Neuroscience, 113(5): 1071-1079.
Clutton-Brock, T. H. 1989. Mammalian mating systems. Proceedings of the Royal Society of London Series B-Biological Sciences, 236(1285): 339-372.
Cushing, B. S., Martin, J. O., Young, L. J. and Carter, C. S. 2001. The effects of peptides on partner preference formation are predicted by habitat in prairie voles. Hormones and
DeVries, A. C., Devries, M. B., Taymans, S. and Carter, C. S. 1995. Modulation of pair bonding in female prairie voles (Microtus ochrogaster) by corticosterone. Proceedings of the National Academy of Sciences of the United States of America, 92(17): 7744- 7748.
DeVries, A. C., DeVries, M. B., Taymans, S. E. and Carter, C. S. 1996. The effects of stress on social preferences are sexually dimorphic in prairie voles. Proceedings of the National Academy of Sciences of the United States of America, 93(21): 11980-11984.
DeVries, A. C., Johnson, C. L. and Carter, C. S. 1997. Familiarity and gender influence social preferences in prairie voles (Microtus ochrogaster). Canadian Journal of Zoology, 75(2): 295-301.
Dewsbury, D. A. 1981. An exercise in the prediction of monogamy in the field from laboratory data on 42 species of muroid rodents. The Biologist, 63: 138-162.
Dewsbury, D. A., Baumgardner, D. J., Evans, R. L. and Webster, D. G. 1980. Sexual
dimorphism for body mass in 13 taxa of muroid rodents under laboratory conditions.
Journal of Mammalogy, 61(1): 146-149.
Emlen, S. T. and Oring, L. W. 1977. Ecology, sexual selection, and evolution of mating systems. Science, 197(4300): 215-223.
Evans, S. 1983. The pair-bond of the common marmoset, Callithrix jacchus jacchus: an experimental investigation. Animal Behaviour, 31: 651-656.
Faul, F., Erdfelder, E., Buchner, A. and Lang, A. G. 2009. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4): 1149-1160.
Faul, F., Erdfelder, E., Lang, A. G. and Buchner, A. 2007. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2): 175-191.
Ferkin, M. H. 1988. The effect of familiarity on social interactions in meadow voles, Microtus pennsylvanicus: a laboratory and field study. Animal Behaviour, 36: 1816-1822.
Fitch, H. S. 1957. Aspects of reproduction and development in the prairie vole (Microtus ochrogaster). University of Kansas Publications, Museum of Natural History, 10: 129- 161.
Gaines, M. S. and Johnson, M. L. 1984. A multivariate study of the relationship between
in Microtus. Journal of Mammalogy, 69(2): 311-319.
Getz, L. L., Carter, C. S. and Gavish, L. 1981. The mating system of the prairie vole, Microtus ochrogaster: field and laboratory evidence for pair-bonding. Behavioral Ecology and Sociobiology, 8(3): 189-194.
Getz, L. L. and Hofmann, J. E. 1986. Social organization in free-living prairie voles, Microtus ochrogaster. Behavioral Ecology and Sociobiology, 18(4): 275-282.
Getz, L. L., Hofmann, J. E. and Carter, C. S. 1987. Mating system and population fluctuations of the prairie vole, Microtus ochrogaster. American Zoologist, 27(3): 909-920.
Getz, L. L., McGuire, B., Pizzuto, T., Hofmann, J. E. and Frase, B. 1993. Social organization of the prairie vole (Microtus ochrogaster). Journal of Mammalogy, 74(1): 44-58.
Gruder-Adams, S. and Getz, L. L. 1985. Comparison of the mating system and paternal
behavior in Microtus ochrogaster and Microtus pennsylvanicus. Journal of Mammalogy, 66(1): 165-167.
Heske, E. J. and Ostfeld, R. S. 1990. Sexual dimorphism in size, relative size of testes, and mating systems in North American voles. Journal of Mammalogy, 71(4): 510-519.
Hume, I. D., Morgan, K. R. and Kenagy, G. J. 1993. Digesta retention and digestive
performance in sciurid and microtine rodents: effects of hindgut morphology and body- size. Physiological Zoology, 66(3): 396-411.
Insel, T. R. and Hulihan, T. J. 1995. A gender-specific mechanism for pair bonding: oxytocin and partner preference formation in monogamous voles. Behavioral Neuroscience, 109(4): 782-789.
Insel, T. R., Preston, S. and Winslow, J. T. 1995. Mating in the monogamous male: behavioral consequences. Physiology & Behavior, 57(4): 615-627.
Insel, T. R. and Shapiro, L. E. 1992. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proceedings of the National Academy of Sciences of the United States of America, 89(13): 5981-5985.
Insel, T. R., Wang, Z. X. and Ferris, C. F. 1994. Patterns of brain vasopressin receptor distribution associated with social organization in microtine rodents. Journal of Neuroscience, 14(9): 5381-5392.
Insel, T. R., Winslow, J. T., Wang, Z. X. and Young, L. J. 1998. Oxytocin, vasopressin, and the neuroendocrine basis of pair bond formation. Vasopressin and Oxytocin: Molecular, Cellular, and Clinical Advances, 449: 215-224.
Jike, L., Batzli, G. O. and Getz, L. L. 1988. Home ranges of prairie voles as determined by
Kleiman, D. G. 1977. Monogamy in mammals. Quarterly Review of Biology, 52(1): 39-69.
Lim, M. M., Liu, Y., Ryabinin, A. E., Bai, Y., Wang, Z. and Young, L. J. 2007. CRF receptors in the nucleus accumbens modulate partner preference in prairie voles. Hormones and Behavior, 51(4): 508-515.
Lim, M. M., Wang, Z. X., Olazabal, D. E., Ren, X. H., Terwilliger, E. F. and Young, L. J. 2004.
Enhanced partner preference in a promiscuous species by manipulating the expression of a single gene. Nature, 429(6993): 754-757.
Liu, Y., Curtis, J. T. and Wang, Z. X. 2001. Vasopressin in the lateral septum regulates pair bond formation in male prairie voles (Microtus ochrogaster). Behavioral Neuroscience, 115(4): 910-919.
Lott, D. F. 1984. Intraspecific variation in the social systems of wild vertebrates. Behaviour, 88:
266-325.
Madison, D. M. 1980. Space use and social structure in meadow voles, Microtus pennsylvanicus. Behavioral Ecology and Sociobiology, 7(1): 65-71.
Madison, D. M., Fitzgerald, R. W. and McShea, W. J. 1984. Dynamics of social nesting in overwintering meadow voles (Microtus pennsylvanicus): possible consequences for population cycling. Behavioral Ecology and Sociobiology, 15(1): 9-17.
McGuire, B., Getz, L. L., Hofmann, J. E., Pizzuto, T. and Frase, B. 1993. Natal dispersal and philopatry in prairie voles (Microtus ochrogaster) in relation to population density, season, and natal social environment. Behavioral Ecology and Sociobiology, 32(5): 293- 302.
McGuire, B. and Novak, M. 1986. Parental care and its relationship to social organization in the montane vole (Microtus montanus). Journal of Mammalogy, 67(2): 305-311.
Oliveras, D. and Novak, M. 1986. A comparison of paternal behavior in the meadow vole Microtus pennsylvanicus, the pine vole Microtus pinetorum and the prairie vole Microtus ochrogaster. Animal Behaviour, 34: 519-526.
Orians, G. H. 1969. On evolution of mating systems in birds and mammals. American Naturalist, 103(934): 589-603.
Ostfeld, R. S. and Heske, E. J. 1993. Sexual dimorphism and mating systems in voles. Journal of Mammalogy, 74(1): 230-233.
Parker, K. J., Phillips, K. M. and Lee, T. M. 2001. Development of selective partner
species: implications for the mating system of the mound-building mouse Mus spicilegus. Behavioural Processes, 51(1-3): 35-43.
Pei, Y. X., Wang, D. H. and Hume, I. D. 2001. Selective digesta retention and coprophagy in Brandt's vole (Microtus brandti). Journal of Comparative Physiology B: Biochemical Systemic and Environmental Physiology, 171(6): 457-464.
Pierce, J. D. and Dewsbury, D. A. 1991. Female preferences for unmated versus mated males in 2 species of voles (Microtus ochrogaster and Microtus montanus). Journal of
Comparative Psychology, 105(2): 165-171.
Price, E. C. and McGrew, W. C. 1991. Departures from monogamy in colonies of captive cotton-top tamarins. Folia Primatologica, 57(1): 16-27.
Reichard, U. H. 2003. Monogamy: past and present. Cambridge University Press.
Solomon, N. G. 1993. Body size and social preferences of male and female prairie voles, Microtus ochrogaster. Animal Behaviour, 45(5): 1031-1033.
Swihart, R. K. and Slade, N. A. 1989. Differences in home-range size between sexes of Microtus ochrogaster. Journal of Mammalogy, 70(4): 816-820.
Taylor, S. A., Salo, A. L. and Dewsbury, D. A. 1992. Estrus induction in 4 species of voles (Microtus). Journal of Comparative Psychology, 106(4): 366-373.
Thomas, J. A. and Birney, E. C. 1979. Parental care and mating system of the prairie vole, Microtus ochrogaster. Behavioral Ecology and Sociobiology, 5(2): 171-186.
Williams, J. R., Carter, C. S. and Insel, T. 1992. Partner preference development in female prairie voles is facilitated by mating or the central infusion of oxytocin. Annals of the New York Academy of Sciences, 652: 487-489.
Williams, J. R., Catania, K. C. and Carter, C. S. 1992. Development of partner preferences in female prairie voles (Microtus ochrogaster): the role of social and sexual experience.
Hormones and Behavior, 26(3): 339-349.
Williams, J. R., Insel, T. R., Harbaugh, C. R. and Carter, C. S. 1994. Oxytocin administered centrally facilitates formation of a partner preference in female prairie votes (Microtus ochrogaster). Journal of Neuroendocrinology, 6(3): 247-250.
Winslow, J. T., Hastings, N., Carter, C. S., Harbaugh, C. R. and Insel, T. R. 1993. A role for central vasopressin in pair bonding in monogamous prairie voles. Nature, 365(6446):
545-548.
Wittenberger, J. F. 1979. The evolution of mating systems in birds and mammals. New York.
Wittenberger, J. F. and Tilson, R. L. 1980. The evolution of monogamy: hypotheses and
Wolff, J. O., Edge, W. D. and Bentley, R. 1994. Reproductive and behavioral biology of the gray-tailed vole. Journal of Mammalogy, 75(4): 873-879.
Wu, J. S., Chiang, P. J. and Lin, L. K. 2012. Monogamous system in the Taiwan vole Microtus kikuchii inferred from microsatellite DNA and home ranges. Zoological Studies, 51(2):
204-212.
Young, L. J. and Wang, Z. X. 2004. The neurobiology of pair bonding. Nature Neuroscience, 7(10): 1048-1054.
Young, L. J., Wang, Z. X. and Insel, T. R. 1998. Neuroendocrine bases of monogamy. Trends in Neurosciences, 21(2): 71-75.
28Tables 1. Body weight (in grams) of the Taiwan field vole at 90~180 days of age compared to three other microtine species at 90 days of age. Values in parentheses give the weight difference in relation to female weight. Body weight (mean ± sd)Sex Differences SpeciesLittersMaleFemaleMale − Femalep Reference M. kikuchii8 43.19 ± 3.3837.03 ± 3.546.16 ± 3.18 (16.6%)<.001This study M. ochrogaster3243.5 ± 6.337.3 ± 3.86.2 (16.6%)0.001Dewsbury et al. 1980 M. pennsylvanicus2553.8 ± 6.744.2 ± 9.79.6 (21.7%)0.001Dewsbury et al. 1980 M. montanus2542.4 ± 5.131.4 ± 5.011.0 (35.0%)0.001Dewsbury et al. 1980
Independent variable df Estimate SE Square p Partner Preferences of Males
Partner/Stranger 1 -1.20 1.02 1.40 0.24 Partner Preferences of Females
Partner/Stranger 1 -2.35 1.14 4.22 0.04*
*: p < 0.05
30
linear regression analyses using time as dependent variable and partner/stranger as independent variable. Dependent variableIndependent variableR-squaredfSSMSF Partner Preferences of Males !!!!!! Time in the chamberPartner/Stranger0.021 907.27 907.27 0.37 Time in physical contactPartner/Stranger0.08 1 3230.83 3230.83 1.30 Partner preferences of Females!!!!!! !Time in the chamberPartner/Stranger0.251 8037.798037.795.01 !!Time in physical contactPartner/Stranger0.12 1 2943.142943.141.96 *: p < 0.05
31
Results of categorical logistic regression analyses using outcome (preferred/non-preferred) as dependent variable and hormone Dependent variableIndependent variablesdfEstimateSEWald Chi- SquareAdjusted -preferred
Progesterone Mean before pairing1 -0.00050.0030.02 CV before pairing1 0.001470.020.004 Value during pairing1 0.0001580.00070.05 Estradiol Mean before pairing1 0.003080.0030.93 CV before pairing1 -0.01130.06 0.04 Value during pairing1 0.001020.0010.60 Corticosterone Mean before pairing1 0.0007270.003 0.07 CV before pairing1 -0.01170.04 0.10 Value during pairing1 0.000650.0010.26
32
5. Effects of hormone (progesterone, estradiol, and corticosterone) concentration of partner/stranger on the outcomes of male prefe tests. Results of linear regression analyses using outcome (time in each chamber) as dependent variable and hormone concentra independent variable. Dependent variable Independent variablesR-squaredfSSMSFAdjusted Time in each chamber
Progesterone!!!!!! Mean before pairing0.04521 1193.28 1193.28 0.661.00 CV before pairing0.03671 969.90969.900.531.00 Value during pairing0.01601 423.38 423.38 0.231.00 Estradiol!!!!!! Mean before pairing0.01471 388.67388.670.211.00 CV before pairing0.00071 18.6118.610.011.00 Value during pairing0.10841 2861.38 2861.38 1.70 0. Corticosterone!!!!! Mean before pairing0.01611 425.36425.360.231.00 !CV before pairing0.00171 45.8045.800.021.00 !!Value during pairing0.00591 154.76154.760.081.00
33 ntration of partner/stranger on the outcomes of male preference Dependent variableIndependent variablesR-squaredfSSMSFAdjusted Progesterone!!!!!! Mean before pairing0.01331 374.86374.860.19 CV before pairing0.00141 40.5740.570.02 Value during pairing0.00281 79.51 79.51 0.04 Estradiol!!!!!! Mean before pairing0.02021 568.19 568.19 0.29 CV before pairing0.03311 932.03932.030.48 Value during pairing0.03941 1109.071109.070.57 Corticosterone!!!!!! Mean before pairing0.13411 3776.68 3776.68 2.17 !CV before pairing0.04281 1206.421206.420.63 !!Value during pairing0.04061 1144.341144.340.59
34
Effects of hormone (testosterone and corticosterone) concentrations of partner/stranger on the outcomes of female preference Results of categorical logistic regression analyses using outcome (preferred/non-preferred) as dependent variable and hormone concentration as independent variable. Dependent variableIndependent variabledfEstimateSEWald Chi- SquareAdjusted Preferred/Non-preferred
Testosterone Mean before pairing1 0.001590.007170.05 1.00 CV before pairing1 0.02780.06050.211.00 Value during pairing1 0.006980.008190.73 1.00 Corticosterone Mean before pairing1 0.005340.005360.990. CV before pairing1 -0.02550.06030.18 0.67 Value during pairing1 0.007390.005291.950.
35 female preference tests. Dependent variableIndependent variablesR-squaredfSSMSFAdjusted Testosterone!! !!!!! Mean before pairing0.00161 24.3424.340.011.00 CV before pairing0.00131 20.9520.950.011.00 Value during pairing0.01441 224.76224.760.121.00 Corticosterone!!!!!!! Mean before pairing0.06971 1084.581084.580.61.00 CV before pairing0.00001 0.0880.0880 1.00 Value during pairing0.11771 1830.231830.231.071.00
36 female preference tests. Results of linear regression analyses using outcome (time in physical contact) as dependent variable and hormone concentration as independent variable. Dependent variableIndependent variablesR-squaredfSSMSFAdjusted Time in physical contact
Testosterone!! !!!! Mean before pairing0.00141 22.5422.540.01 CV before pairing0.00131 19.8819.880.01 Value during pairing0.00001 0.600.600 Corticosterone!!!!!! Mean before pairing0.35581 5591.385591.384.42 CV before pairing0.11391 1790.351790.351.03 Value during pairing0.23881 3752.693752.692.51
Monogamy(?(
Partner'Preference'Test'
Selec.ve(
Affilia.on( Selec.ve(
Aggression(
Other(
Func.ons(
Other(criteria(?(
Sexual(
Dimorphism(
Steroid(
hormones(
Pair(bonding(
Figure 1. The concept map of this thesis.
Figure 2. The partner preference test apparatus is 135 × 25 × 50 cm3 (LxWxH) in dimension, and consists of three chambers (45 × 25 × 50 cm3, LxWxH). The two dividers each has a 7 x 7-cm passage hole.
y"="$0.179ln(x)"+"1.0296"
R²"="0.99959"
0"
0.2"
0.4"
0.6"
0.8"
1" 10" 100"
O.D.$
Cor(costerone$concentra(on$(Log$ng/ml)$
Figure 3. A standard curve of corticosterone concentration. !
!
!
!
!
!
!
!
!
!
!
!
!
0"
20"
40"
60"
80"
Male% Female%
Time%in%each%chamber%(minute)
Figure 4. Mean (± 1se) time the focal voles spent in the partner ( ) vs. the stranger ( ) chambers during the 180-min partner preference tests (male: n = 9; female: n
= 8). There was no significant difference between partner and stranger.
0"
10"
20"
30"
40"
50"
60"
70"
Male% Female%
Time%in%physical%contact%(minute)
Figure 5. Mean (± 1se) time the focal voles spent in physical contact with the partners ( ) vs. the strangers ( ) during the 180-min partner preference tests (male:
n = 9; female: n = 8). There was no significant difference between partner and stranger.
0"
10"
20"
30"
40"
Male% Female%
Time%in%the%neutral%chamber%(minu
Figure 6. Mean (± 1se) time that Taiwan field voles spent in the neutral chambers during 180-min preference tests (male: n = 9; female: n = 8). Female focal voles spent significantly more time in the neutral chamber than male focal voles (t-test; t = 2.22, p = 0.03).
(A) Male preference tests
No. Focal vole Partner Stranger
1 B011 B012 B020
2 B049 B042 B038
3 B047 B038 B042
4 B051 C002 C004
5 B043 C004 C002
6 C009 B046 B054
7 C001 B054 B046
8 C015 B056 B058
9 C017 B058 B056
(B) Female preference tests
No. Focal vole Partner Stranger
1 B018 B013 B017
2 B024 B027 B015
3 B030 B025 B029
4 B048 C003 B055
5 B060 C019 C021
6 B066 C021 C019
7 C008 B065 B061
8 B062 B061 B065
0"
200"
400"
600"
800"
1000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Progesterone)(ng/g))
Day)
B042)
0"
200"
400"
600"
800"
1000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Progesterone)(ng/g))
Day)
B038)
0"
500"
1000"
1500"
2000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Progesterone)(ng/g))
Day)
C002)
0"
1000"
2000"
3000"
4000"
5000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Progesterone)(ng/g))
Day)
C004)
0"
200"
400"
600"
800"
1000"
1200"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"11"
Progesterone)(ng/g))
Day)
B046)
0"
200"
400"
600"
800"
1000"
1200"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"11"
Progesterone)(ng/g))
Day)
B054)
0"
200"
400"
600"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Progesterone)(
Day)
0"
200"
400"
600"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Progesterone)(
Day)
(B) Estradiol
0"
200"
400"
600"
800"
1000"
1200"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Estradiol*(ng/g)*
Day*
B042*
0"
200"
400"
600"
800"
1000"
1200"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Estradiol*(ng/g)*
Day*
B038*
0"
500"
1000"
1500"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Estradiol*(ng/g)*
Day*
C002*
0"
500"
1000"
1500"
2000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Estradiol*(ng/g)*
Day*
C004*
0"
500"
1000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"11"
Estradiol*(ng/g)*
Day*
0"
200"
400"
600"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"11"
Estradiol*(ng/g)*
Day*
0"
200"
400"
600"
800"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Estradiol*(ng/g)*
Day*
B056*
0"
200"
400"
600"
800"
1000"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Estradiol*(ng/g)*
Day*
B058*
(C) Testosterone
0"
100"
200"
300"
400"
500"
600"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Testosterone((ng/g)(
Day(
C003(
0"
100"
200"
300"
400"
500"
1" 2" 3" 4" 5" 6" 7" 8" 9" 10"
Testosterone((ng/g)(
Day(
B055(