行政院國家科學委員會補助專題研究計畫
█ 成 果 報 告
□期中進度報告
(計畫名稱)
登革病毒感染的致病機轉:動物模式
計畫類別:▇ 個別型計畫 □ 整合型計畫
計畫編號:NSC91-3112-B006-003-
執行期間: 91 年 5 月 1 日至 92 年 4 月 31 日
計畫主持人:黎煥耀
共同主持人:
計畫參與人員: 黃國珍、楊育靜
成果報告類型(依經費核定清單規定繳交):□精簡報告 ▇完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,▇一年□二年後可公開查詢
執行單位:國立成功大學
中 華 民 國 92 年 7 月 29 日
ABSTRACT
Dengue virus infection causes dengue fever, dengue hemorrhagic fever (DHF), and dengue shock syndrome (DSS). Thrombocytopenia is common in dengue fever, and is always found in DHF/DSS. The pathogenesis of thrombocytopenia is poorly understood. IgM anti-platelet auto-antibody was found in dengue patients. To further understand the relationship between anti-dengue virus antibody and anti-platelet antibody, monoclonal anti-dengue virus antibodies from the dengue virus infected mice that developed transient thrombocytopenia post dengue infection were generated. The analysis of a panel of monoclonal anti-NS-1 antibodies reveals three different patterns of platelet binding: strong, intermediate, or dull. Their isotypes are different, some are IgM while others are IgG1. Most of anti-platelet antibodies are cross-reactive
with NS-1 of dengue virus, and can be competitively inhibited by recombinant NS-1 protein, suggesting a molecular mimicry between dengue virus NS-1 protein and platelet. A clone, 13-F4-G5, preferentially bound activated platelets, can recognize two or three proteins around 150 kD on platelets. The binding to platelet would lyse the platelet in the presence of complement or enhance the ADP-induced platelet aggregation. Furthermore, some of these monoclonal
antibodies would also react with the cellular antigens of BHK. These studies suggest that dengue virus infection induces auto anti-platelet antibodies which thereafter may involve in the
manifestation of thrombocytopenia. A molecular mimicry between NS-1 and platelet is demonstrated.
中文摘要 登革病毒感染會引起登革熱或會致命的登革出血熱╱登革休克症候群,其致病機制一 直是研究的主題。我們過去的研究發現,登革病毒感染會誘導免疫反應的過度活化,造成 CD4/CD8 T-細胞比率的反轉,細胞激素的大量產生以及 T-細胞增殖反應受損。同時產生抗 血小板自体抗体,造成血小板減少,而內皮細胞或肝細胞受到病毒感染,直接及透過免疫 反應影響到內皮細胞的破壞,造成凝血和溶血的不平衡,形成登革出血熱。為瞭解登革病 毒感染產生自體抗体,造成血小板破壞減少的病變,我們利用融合瘤的技術從登革病毒感 染小鼠製造單株抗體,這些抗登革病毒的抗體可以和血小板結合。它的類型有 IgM 及 IgG1, 用登革病毒的 NS-1 可以中和它和血小板的結合,顯示登革病毒的 NS-1 和血小板間有分子 模擬的關係。這些抗血小板自体抗体,會增強活化血小板的凝集,在補体存在下也會溶解 血小板,這種抗血小板自体抗体的產生及作用可能是登革病毒感染造成血小板低下的主 因,也對於登革病毒感染和自体免疫力間的分子模擬提供了一個很好的模式供未來做更進 一步的研究。
INTRODUCTION
Dengue fever (DF) is an acute infectious disease caused by dengue virus which has four serotypes. It is characterized by biphasic fever, headache, pain in various parts of the body, rash, lymphadenopathy, and leukopenia. In most cases, the disease of dengue fever is self-limited. However, there is risk to progress into dengue hemorrhagic fever (DHF) or dengue shock syndrome (DSS) especially when cross infection of different serotypes occurs. DHF is a severe febrile disease characterized by abnormalities of hemostasis and increased vascular permeability, which in some instances results in DSS. DSS is a form of hypovolemic shock that is associated clinically with hemoconcentration and frequently leads to death if appropriate care is not given (2,3,9). Thrombocytopenia is common in dengue fever, and is always found in DHF/DSS. Its pathogenesis is poorly understood. La Russa and Innis reported dengue-virus-induced bone marrow suppression that depressed platelet synthesis (15). Wang et al, found that dengue-2 virus can bind to human platelets in the presence of virus-specific antibody (28). We also reported the presence of IgM anti-platelet auto-antibody in the sera of dengue patients, and its titer is higher in DHF/DSS patients than in DF patients (18). To further understand the relationship between anti-dengue virus antibody and anti-platelet antibody, a murine model of dengue virus infection was setup. Transient thrombocytopenia developed at 10-13 days after primary or secondary infection and was associated with the generation of anti-platelet antibody (12). A panel of
monoclonal antibodies was generated from these dengue virus-infected mice. In this study, it was reported that anti-dengue virus antibodies, especially anti-NS-1 ones, could cross-react with platelet. The molecular mimicry between dengue virus and self-antigens was discussed.
MATERIALS AND METHODS
Mice and cell culture. Breeder mice of BALB/c strain were purchased from The Jackson
Laboratory, Bar Harbor, ME or Charles River Japan, Inc. (Atsugi, Japan). They were maintained on standard laboratory chow and water ad libitum in the animal facility of the Medical College, National Cheng Kung University, Tainan, Taiwan. The animals were raised and cared for
following the guidelines set up by the National Science Council of the Republic of China. Six to twelve-week-old mice were used in all experiments. BHK and K562 cells were grown in DME medium containing 10% FBS.
Dengue virus preparation. A local isolate of dengue virus type 2 (PL046) was supplied by
the Institute of Preventive Medicine, Nan Kung, Taipei, Taiwan. Viruses were propagated in mosquito C6/36 cell line which were incubated in Eagle’s minimal essential medium containing 2% heat-inactivated FBS at 280C for 5 days (13). Each virus pool can obtain a titer of 1x108 PFU/ml by standard methods on BHK cells (25). To purify the dengue virus antigen, the pooled virus stocks were first concentrated by ultra-filtration of 10 kDa cut-off membrane, then
centrifuged at 10,000 x g for 10 min. The supernatant was further centrifuged at 100,000 x g for 3 h to pellet the dengue virus particle. This procedure can enrich virus stock to 1x1011 PFU/ml. The semi-purified dengue virus was used in the ELISA to screen the anti-dengue antibody.
Virus infected cells. BHK or K562 cells were maintained in Dulbecco’s modified eagle
medium containing 2% heat-inactivated FBS at 37℃and infected by either herpes simplex virus (KOS strain) or dengue virus at the m.o.i.=5. Cells were harvested at 48 h post infection and subjected to flow cytometric analysis. In some studies, BHK cells were grown in the Lab-Tek chamber slide system (Nalge Nunc International, Naperville, IL). They were infected at m.o.i. =1 for 48 h. The dengue antigen was detected with 13-F4-G5 mAb and second goat anti-mIgG peroxidase conjugate. A peroxidase stain with a reddish brown color was developed with an aminoethyl carbazole substrate kit (ZYMED Laboratories, San Francisco, CA), and
counterstained with 1% Evans blue. In the preparation of dengue virus infected cell lysates, C6/36 cell was maintained in Eagle’s minimal essential medium containing 2% heat-inactivated FBS at 28℃and was infected by dengue virus at m.o.i. =1. After 5 days of incubation, cells were harvested and then lysed in RIPA buffer, and used as the dengue virus antigen source.
Generation of monoclonal antibody from dengue-2-virus-infected mice. Groups of
BALB/c mice were inoculated intravenously with dengue virus (1 x 108 PFC). The mice were sensitized for 3 or 4 times at one-month interval with 1 x 107 PFC. Before the fusion, the mice were boosted with 1 x 106 PFC for three days. The splenocytes were fused with FO myeloma using 1% PEG as described previously (16). Several methods including ELISA on dengue virus or dengue virus infected cells, or anti-platelet binding by flow cytometry were used to screen the antibodies.
collected in sodium EDTA was centrifuged at 100 x g for 10 min at room temperature. The upper layer as platelet-rich plasma was removed to a 15-ml tube, mixed with 0.34% EDTA in
phosphate-buffered saline (PBS) and centrifuged at 1000 x g for 15 min. The pellets were washed 3 times with 0.34% EDTA-PBS and fixed in 10 ml of 1% formaldehyde in PBS at room
temperature for 10 min. The fixed platelet suspension was centrifuged at 1000 x g for 15 min. The pellets were washed in PBS twice and resuspended in 2 ml of PBS. The platelet count was determined using a hemacytometer. The anti-platelet antibody binding was determined with flow cytometric analysis as described previously (23). Platelets (2.5 x 106/0.1 ml) were incubated with monoclonal antibody for 60 min on ice, then washed twice with PBS. The second goat
anti-mouse IgG FITC-conjugated antibody (Cappel, Organon Teknika, N.V. Belgium) was added and the mixture was incubated for 40 min on ice. After washing twice with PBS, the platelets were suspended in PBS and analyzed by FACScan (Becton-Dickinson, Mountain View,CA) with excitation set at 488 nm. In some experiments, BHK or K562 cells infected with dengue virus at m.o.i.= 5 for 48 h were used. For intracellular staining, cells were treated with fixation buffer containing 4% paraformaldehyde followed by permeabilization buffer containing 0.1% saponin.
Platelet lysates. Normal human blood was collected in a tube containing 0.33% sodium
citrate at 9:1 ratio, then was centrifuged at 200 x g for 20 min under room temperature. The platelet-rich plasma (PRP) was collected and further centrifuged at 1000 x g for 10 min. The platelet pellet was gently resuspended in HEPES buffer (134 mM NaCl, 12 mM NaHCO3, 2.9
mM KCl, 0.34 mM Na2HPO4, 1 mM MgCl2, 10 mM hepes, 5 mM glucose, 0.3% BSA, pH 7.4),
and platelet number was counted with a hemacytometer. Platelets were activated by thrombin at concentration of 1 U/ml. After 5 min of incubation, the platelets were washed twice with HEPES buffer at 1000 x g for 10 min, and lysed in RIPA buffer (1% Triton X-100, 0.5% sodium
deoxycholate, 0.1% SDS, 150 mM NaCl, 1 mM EGTA, 50 mM Tris pH 7.6).
Platelet aggregation test. The platelet-rich plasma was collected as previously described.
The remaining portion was further centrifuged at 1200 x g for 10 min at room temperature to collect the platelet-poor plasma (PPP). Aggregation test was done with the instruction of the aggregometer (Hema Tracer 2, Niko Bioscience). Different concentrations of monoclonal antibody were incubated with PPP for 10 min before adding platelet agonist, ADP. Platelet aggregation was recorded for 10 minutes.
Western blot. Samples for electrophoretic analysis were disrupted for 5 min at 100o C in
0.125 M Tris-HCl (pH 6.8) containing 2.5% SDS, 5% 2-ME, and 10% glycerol. Bromophenol blue was added as a tracking dye. Separation gel with 12% acrylamide, 0.1% SDS, and 0.375% Tris-HCl (pH 8.8) and stacking gel of 4% acrylamide, 0.1% SDS, and 0.125% Tris-HCl (pH 6.8) were prepared. An equivalent amount of protein prepared from platelets was loaded into the gel. Electrophoresis was performed at 20 mA/gel until the bromophenol blue line reached the bottom of the gel. After electrophoresis, gels were transblotted to nitrocellulose paper with 25 mM Tris, 192 mM glycine in a transblot apparatus at 70 V for 3 h. After transfer, a small strip was cut and
stained with amido black to check the transfer of the protein. The nitrocellulose paper was then blocked with 1% BSA. The platelet proteins recognized by anti-platelet antibody were
determined. The goat anti-mouse IgG peroxidase conjugate (Cappel, Organon Teknika, N.V. Belgium) was used as secondary antibodies. The color was developed with PBS containing 0.05% 4-chlor-1-naphthol and 0.01% H2O2 (17).
Preparation of recombinant NS-1. The full length dengue-2 virus NS1 cDNA was cloned
to pRSET B expression vector (Invitrogen) to establish a pRSET-DVNS1 plasmid. After the sequences at 5’ and 3’ junctions were confirmed, this plasmid was then introduced into E. coli BL21(DE3)pLysS strain (Invitrogen). The recombinant NS1 (rNS1) proteins were induced by IPTG and purified with TALON metal affinity resin (Clontech). A single band was observed by SDS-PAGE analysis and the protein sequence was confirmed by an Applied Biosystems 477A autosequencer .
Antibody-mediated complement-dependent cytotoxicity of platelets. The platelet-rich
plasma was centrifuged on the BSA step-gradient (consisting of 50%, 25%, 17%, 12%, 10% BSA) at 1500 x g for 15 min at room temperature. The platelet fraction was collected and gel-filtrated with a Sepharose 2B (Sigma) column that was pre-equilibrated with HEPES buffer. Platelets (1 x 108 platelets) in 96-well plate were incubated with protein-G purified monoclonal antibodies in the presence of rabbit complement (1:25 dilution) for 4 h. The plate was then centrifuged at 1500 x g for 15 min to collect the supernatant. The supernatant (100 l) was transferred into 96-well ELISA plate and mixed with 100 l of the lactase dehydrogenase substrate (Boehringer
Mannheim, Mannheim, Germany). The OD was read at 450 nm after 30 min with ELISA reader. Cytotoxicity was calculated following the instruction of LDH cytotoxicity assay.
RESULTS
Different patterns of platelet binding by monoclonal antibodies derived from dengue- 2-virus-infected mice. Dengue virus infection can induce anti-platelet antibodies in human or
mice (12,18). The relationship between anti-dengue virus antibody and autoantibody was elucidated with monoclonal antibodies generated from dengue virus-infected mice. ELISA
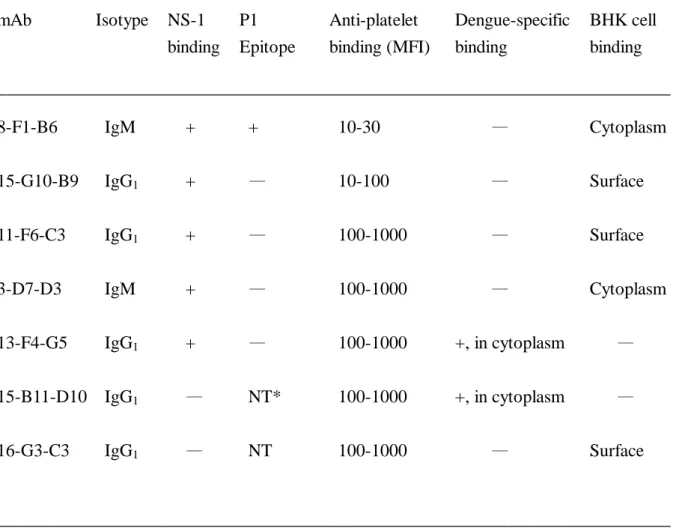
binding on either dengue antigen or dengue virus infected cell was used to screen the anti-dengue virus antibodies. We are particularly interested in the cross-reactive autoantibodies, therefore, the anti-platelet binding by FACScan analysis was also used to screen the monoclonal antibodies. More than 20 different clones were generated from several fusions. Among the anti-platelet antibodies, Figure 1 showed several monoclonal antibodies that react with human platelet. Three patterns of platelet binding were observed based on the degree of fluorescent intensity. Each category of platelet binding had several clones, only representative ones were shown. The strong binding clones are 11-F6-C3, 3-D7-D3, 13-F4-G5, 15-B11-D10, and 16-G3-C3; the intermediate binding, 15-G10-B9; the dull binding, 8-F1-B6. Their immunoglobulin classes are different; 3-D7-D3 and 8-F1-B6 are IgM while the rest are IgG1.
Cross-reactivity between dengue virus NS-1 protein and platelet or other self-antigens.
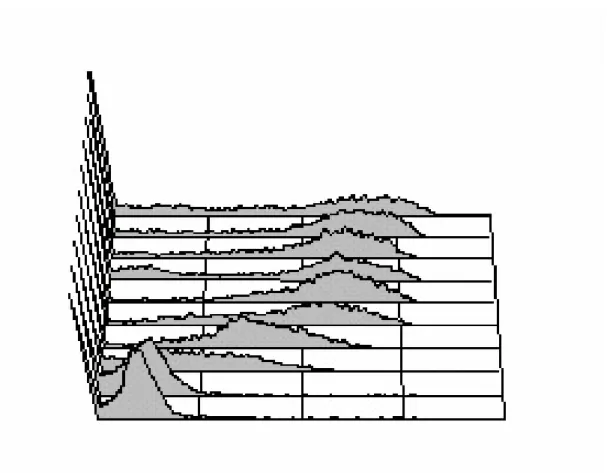
Among the anti-platelet antibodies, the dengue antigen specificity was determined. It was found that 13-F4-G5 can specifically recognize the dengue virus, but not herpes simplex virus antigen as shown by immunohistochemical staining and FACScan analysis on dengue virus infected BHK cells (Fig. 2B & 2C). To determine which protein is recognized, dengue 2 virus-infected C6/36 cell lysate was run on PVP membrane and stained with 13-F4-G5 antibody. As shown in Fig. 2A, it is the NS-1 from dengue virus infected C6/36 cell lysate that was bound by 13-F4-G5 on Western blot. Clones such as 8-F1-B6, 15-G10-B9, 11-F6-C3, 3-D7-D3, and 13-F4-G5 can bind recombinant NS-1 protein by ELISA, but 15-B11-D10 and 16-G3-C3 are not NS-1-reactive (data not shown). Using NS-1 to competitively block the binding, the platelet binding of 13-F4-G5 could be dose-dependently inhibited (Fig. 3). This suggests that a cross-reactive epitope between NS-1 and platelet antigen was recognized by 13-F4-G5 monoclonal antibody. The platelet
antigens recognized by 13-F4-G5 were then further determined by Western blot analysis on platelet lysate. Several bands with high molecular weight around 150 kDa were recognized by 13-F4-G5 (Fig. 4A), the binding were more intensive on thrombin (1 U/ml)-treated platelet than untreated platelet. This indicates that 13-F4-G5 preferentially recognize activated platelets. This preferential binding by 13-F4-G5 on thrombin-activated platelet was also demonstrated with FACScan analysis (Fig. 4B). The fluorescent intensity of 13-F4-G5 binding was higher in thrombin-treated platelet than non-treated platelet. Furthermore, P1 (amino acids 1-15) is the immunodominant linear epitope of NS-1 recognized by dengue patient sera (11). We tested whether P1 is also dominant in these anti-NS-1 antibodies. Only 8-F1-B6 recognized the
P1-peptide, the rest of anti-NS-1 antibodies are P1-peptide binding negative (data not shown).
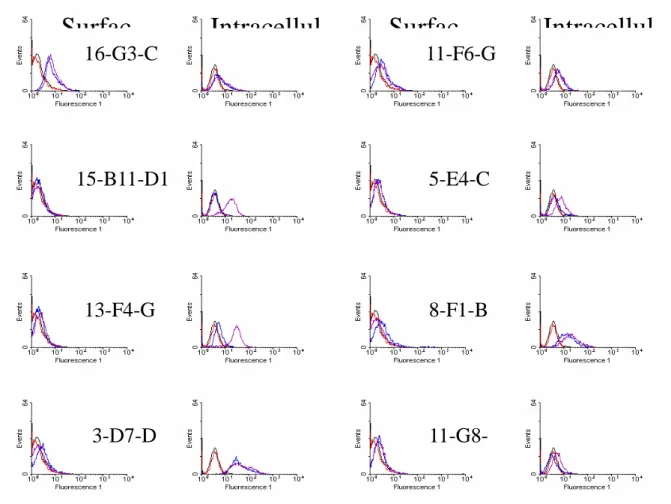
The cross-reactivity with other cellular antigens was further demonstrated by FACScan analysis on dengue virus infected cells. Surface or intracellular staining was used to localize the
antigen on membrane or in cytoplasm, respectively. Both 13-F4-G5 and 15-B11-D10 recognized dengue virus infection-induced antigen in cytoplasm (Fig. 5). But 3-D7-D3 would stain BHK intracellularly irrespective of the dengue virus infection, which suggests that 3-D7-D3 recognized cellular antigen of BHK. Similarly, 11-F6-C3, 16-G3-C3, and 15-G10-B9 could stain the BHK cells on the surface and intracellularly, which indicats that the cellular antigen of BHK
recognized by these mAbs is expressed on the cell surface. The 8-F1-B6 recognized cytoplasmic antigen of BHK cells. We have repeated the binding assay using K562 cell and obtained the same pattern of binding with these mAbs (data not shown). Individual clone has its own characteristic features with regard to the binding of P1, NS-1, platelet, or BHK cells, as summarized in Table 1. This suggests that cross-reactivity between dengue virus NS-1 protein and self-antigens does exist.
The effect of anti-platelet antibody on platelet function. When the platelet was incubated
with monoclonal antibody, we did not observe the platelet degranulation by 3H-serotonin release assay (data not shown). However, in the presence of complement, 13-F4-G5, 11-F6-C3, and 15-B11-D10-binding platelets would be lysed (Fig. 6). Moreover, anti-platelet mAb would enhance the ADP-induced platelet aggregation. At suboptimal amount of ADP (5 M), the platelet aggregation was enhanced by 13-F4-G5 in a dose-dependent manner (Fig. 7). 13-F4-G5 alone or together with goat anti-mIgG antibodies did not cause the platelet aggregation
spontaneously. Based on the data above, we conclude that anti-dengue virus, especially anti-NS1, antibodies would cross-react with platelet as well as cellular self-antigen, and cause their
dysfunction.
DISCUSSION
Thrombocytopenia is characteristic of dengue virus infection. Anti-platelet IgM
autoantibodies whose titers are higher in DHF/DSS patients than in DF patients (18) may involve in its pathogenesis. We used the dengue virus infected mice that developed transient
thrombocytopenia post infection to generate various monoclonal anti-dengue virus antibodies. Many mAbs that bind NS-1 of dengue virus are cross-reactive with platelets. The binding of platelets can be classified into strong, intermediate, and dull three patterns. Either IgM or IgG1
isotypes are present. Its binding to platelets would induce platelet lysis in the presence of complement, or enhance the ADP-induced platelet aggregation. Furthermore, cross-reactivity between NS-1 and platelet or cellular self-antigen was demonstrated.
Thrombocytopenia is common in dengue fever, and is always found in DHF/DSS. The pathogenesis of thrombocytopenia is poorly understood. Either thrombopoiesis in bone marrow was suppressed, or the platelet was destructed. La Russa and Innis suggested that
dengue-virus-induced bone marrow suppression depressed platelet synthesis and resulted in thrombocytopenia (15). Wang et al., found that dengue-2 virus can bind to human platelets in the presence of virus-specific antibody, and proposed that the immune-mediated clearance of
platelets was involved in the pathogenesis of thrombocytopenia in DHF/DSS (28). Virus such as Parvovirus infection is known to be associated with childhood idiopathic thrombocytopenic purpura (8). Falconar reported monoclonal anti-NS-1 of dengue virus could cross-react with human fibrinogen, thrombocytes and endothelial cells (5). We also found that IgM anti-platelet auto-antibody manifests in dengue patients, and its titer is higher in DHF/DSS than in DF patients. The presence of these autoantibodies would induce platelet lysis in the presence of complement (18). The cross-reactivity between dengue virus proteins, especially NS-1 and platelet was further demonstrated using monoclonal antibodies derived from dengue virus-infected mice in this study. Most of the anti-platelet mAbs are reactive to recombinant NS-1, but two of seven clones are NS-1 binding negative. The cross-reactivity between platelet and dengue virus antigens other than NS-1 is not excluded. Although human anti-platelet antibodies are IgM, the panel of mAbs contains IgG1 as well as IgM. The hybridoma fusions are derived from mice that have been
infected intravenously with dengue-2 virus three or four times. The immunoglobulin class is switch from IgM to IgG1, but indeed IgM after multiple infections with dengue virus can be
obtained. The generation of anti-platelet antibody during dengue virus infection will cause platelet destruction, and results in thrombocytopenia. The antibody-mediated deletion of platelet may play an important role on the manifestation of thrombocytopenia. Furthermore, the
molecular mimicry between dengue virus antigen and self-proteins is intriguing and needs further investigation. However, it raises a concern on the immune enhancement or memory of secondary infection. The high affinity of anti-platelet antibody generated after secondary immunization will increase the severity of thrombocytopenia caused by these pathogenic anti-platelet
auto-antibodies.
serotype of the second infection is different from that of the previous infection. The mechanisms involved in the pathogenesis of DHF/DSS remain poorly understood, although the
antibody-dependent enhancement (ADE) hypothesis is proposed to explain this unique clinical phenomenon. ADE is known as an in vitro observation when a subneutralization amount of antibody is present in the culture with virus (6,7). The virus will enter into the cells through the Fc receptor and the replication of virus in cells will be enhanced. When virion increases, immune deviation such as the production of anti-platelet auto-antibody is triggered. High virus load causes more immune activation. Secondary infection by different serotypes of dengue virus might have more virus load and stronger immune deviation compared to the primary one. The titer of anti-platelet antibodies is higher in DHF/DSS than DF probably because of the immune memory on antibody production. The immune memory or immune enhancement in secondary infection can boost more production of high affinity autoantibody.
Viruses have long been associated with inciting autoimmune disorders. Molecular mimicry is proposed to be one of the pathogenetic mechanisms for autoimmune disease. A similar
structure is shared between viral determinants and host antigens. The immune responses against the viral determinant are triggered post infection, which evoke a tissue-specific immune response that is presumably capable of eliciting cell and tissue destruction (21,22). Several virus proteins were mapped to self-antigens such as P2-C protein of coxsackie B virus and glutamate
decarboxylase in insulin-dependent diabetes (1,26), viral proteins and S-antigen in experimental autoimmune uveitis (24), coat protein of herpes simplex virus-type 1 and corneal antigens in autoimmune herpes stromal keratitis (29), transaldolase epitope and Epstein-Barr and herpes simplex virus type 1 capsid in multiple sclerosis (4), human cytomegalovirus late protein UL94 and endothelial cell in systemic sclerosis (20). In parvovirus infection, anti-parvovirus antibodies were reported to react with autoantigens including human keratin, collagen type II, thyroglobulin, single-strand DNA, cardiolipin, and ribonucleoprotein antigen Sm (19). Among the flavivirus, hepatitis C virus (HCV) can induce a number of diseases of presumed autoimmune background, like mixed cryoglobulinaemia, glomerulonephritis, panarthritis, arthritis, thyroiditis and skin lesions. In HCV-induced autoimmune hepatitis, anti-liver and kidney microsome autoantibodies directed against cytochrome P450 2D6 and cytotoxic T cells to HCV core 178-187 were found (14). The molecular mimicry between NS-1 of dengue virus and self-proteins of platelet is the first example in acute infection of dengue virus. It not only contributes to the pathogenesis of thrombocytopenia, but also raises a critical issue on long-term safety of dengue vaccine. The induction of autoimmunity by dengue virus should be taken into consideration in the dengue virus disease.
REFERENCES
1. Atkinson, M.A., M.A. Bowman, L. Campbell, B.L. Darrow, D.L. Kaufman, and N.K.
MacLaren. 1994. Cellular immunity to a determinant common to glutamate decarboxylase
and coxsackie virus in insulin-dependent diabetes. J. Clin. Invest. 94:2125-2129.
2. Bhamarapravati, N., P. Tuchinda, and V. Boonyapaknavik. 1967. Pathology of Thailand
haemorrhagic fever: a study of 100 autopsy cases. Ann. Trop. Med. Parasitol. 61:500-510.
3. Burke, D.S., A. Nisalak, D.E. Johnson, and R.M. Scott. 1988. A prospective study of
dengue infection in Bangkok. Am. J. Trop. Med. Hyg. 38:172-180.
4. Esposito, M., V. Venkatesh, L. Otvos, Z. Weng, S. Vajda, K. Banki, and A. Perl. 1999.
Human transaldolase and cross-reactive viral epitopes identified by autoantibodies of multiple sclerosis patients. J. Immunol. 163:4027-4032.
5. Falconar, A.K.I. 1997. The dengue virus nonstructural-1 protein (NS1) generates antibodies
to common epitopes on human blood clotting, integrin/adhesin proteins and binds to human endothelial cells: potential implications in haemorrhagic fever pathogenesis. Arch Virol
142:897-916.
6. Halstead, S.B. 1970. Observations related to pathogenesis of dengue hemorrhagic fever. VI.
Hypotheses and discussion. Yale J. Biol. Med. 42:350-362.
7. Halstead, S.B. 1988. Pathogenesis of dengue: Challenge to molecular biology. Science
239:476-481.
8. Heegaard, E.D., S. Rosthoj, B.L. Petersen, S. Nielsen, F. Karup Pedersen, and A.
Hornsleth. 1999. Role of parvovirus B19 infection in childhood idiopathic
thrombocytopenic purpura. Acta Paediatr. 88:614-617.
9. Henchal, E.A., and J.R. Putnak. 1990. The dengue viruses. Clin. Microbiol. Rev.
3:376-396.
10. Horwitz, M.S. and N. Sarvetnick. 1999. Viruses, host responses, and autoimmunity. Immunol. Reviews. 169:241-253.
11. Huang, J.H., J.J. Wey, Y.C. Sun, C. Chin, L.J. Chien, and Y.C. Wu. 1999. Antibody responses to an immunodominant nonstructural 1 synthetic peptide in patients with dengue fever and dengue hemorrhagic fever. J. Med. Vriol. 57:1-8.
12. Huang, K.J., S.Y.L. Li, S.C. Chen, H.S. Liu, Y.S. Lin, T.M. Yeh, C.C. Liu, and H.Y.
Lei. 2000. Manifestation of thrombocytopenia in dengue-2-virus-infected mice. J. Gen.
Virol. 81:2177-2182.
13. Igarashi, A. 1978. Isolation of Singh’s Aedes albopictus cell clone sensitive to dengue and chikungunya viruses. J. Gen. Virol. 40:531-544.
14. Kammer, A.R., S.H. van der Burg, B. Grabscheid, I.P. Hunziker, K.M. Kwappenberg,
J. Reichen, C.J. Melief, and A. Cerny. 1999. Molecular mimicry of human cytochrome
P450 by hepatitis C virus at the level of cytotoxic T cell recognition. J. Exp. Med.
190:169-176.
15. La Russa, V.F., and B.L. Innis. 1995. Mechanisms of dengue virus-induced bone marrow suppression. Baillieres. Clin. Haem. 8:249-270.
gene products. II. Presence of both I-Jb and I-Jk suppressor factors in (nonsuppressor x nonsuppressor) F1 mice. J. Exp. Med. 155: 955-967.
17. Lei, H.Y., Y.L. Wang, S.C. Lee, and S.H. Chen. 1992. The effect of pepsin digestion in relation to pre-S region on hepatitis B surface antigen-induced hypersensitivity. J. Immunol.
148:3560-3566.
18. Lin, C.F., H.Y. Lei, C.C. Liu, H.S. Liu, T.M. Yeh, S.T. Wang, T.I. Yang, F.C. Sheu,
C.F. Kuo, and Y.S. Lin. 2001. Generation of IgM anti-platelet autoantibody in dengue
patients. J. Med. Virol. 63:143-149.
19. Lunardi, C., M. Tiso, L. Borgato, L. Nanni, R. Millo, G. De Sandre, A.B. Severi, and
A. Puccetti. 1998. Chronic parvovirus B19 infection induces the production of anti-virus
antibodies with autoantigen binding properties. Eur. J. Immunol. 28:936-948.
20. Lunardi, C., C. Bason, R. Navone, E. Millo, G. Damonte, R. Corrocher, and A. Puccetti. 2000. Systemic sclerosis immunoglobulin G autoantibodies bind the human
cytomegalovirus late protein UL94 and induce apoptosis in human endothelial cells. Nature Med. 6:1183-1186.
21. Manns, M.P., and P. Obermayer-Straub. 1997. Viral induction of autoimmunity: mechanisms and examples in hepatology. J. Viral Hepatitis. 4 (Suppl 2): 42-47. 22. Oldstone, M.B. 1998. Molecular mimicry and immune-mediated diseases. FASEB J.
12:1255-1265.
23. Schwarz, U.R., J. Geiger, U. Walter, and M. Eigenthaler. 1999. Flow cytometry analysis of intracellular VASP phosphorylation for the assessment of activating and inhibitory signal transduction pathways in human platelets. Thombosis Haemostasis 82:1145-1152.
24. Singh, V.K., H.K. Kalra, K. Yamaki, T. Abe, L.A. Donoso, and T. Shinohara. 1990. Molecular mimicry between a uveitopathogenic site of S-antigen and viral peptides: induction of experimental autoimmune uveitis in Lewis rats. J. Immunol. 144: 1282-1287. 25. Sukhavachana, P., A. Nisalak, and S.B. Halstead. 1966. Tissue culture techniques for the
study of dengue viruses. Bull. WHO 35:65-66.
26. Varela-Calvino, R., G. Sgarbi, S. Arif, and M. Peakman. 2000. T-Cell reactivity to the P2C nonstructural protein of a diabetogenic strain of coxsackievirus B4. Virology.
274:56-64.
27. Vreugdenhil, G.R., M.R. Batstra, H.J. Aanstoot, W.J. Melchers, and J.M. Galama. 1999. Analysis of antibody responses against coxsackie virus B4 protein 2C and the diabetes autoantigen GAD(65). J. Med. Virol. 59:256-261.
28. Wang, S., R. He, J. Patarapotikul, B.L. Innis, and R. Anderson. 1995.
Antibody-enhanced binding of dengue-2 virus to human platelets. Virology 213:254-257. 29. Zhao, Z.S., F. Granucci, L.Yeh, P.A. Schaffer, and H. Cantor H. 1998. Molecular
mimicry by herpes simplex virus-type 1: autoimmune disease after viral infection. Science
Table 1. Summary of anti-platelet mAb derived from dengue virus infected mice.
___________________________________________________________________________ mAb Isotype NS-1 P1 Anti-platelet Dengue-specific BHK cell binding Epitope binding (MFI) binding binding ___________________________________________________________________________ 8-F1-B6 IgM + + 10-30 — Cytoplasm 15-G10-B9 IgG1 + — 10-100 — Surface 11-F6-C3 IgG1 + — 100-1000 — Surface 3-D7-D3 IgM + — 100-1000 — Cytoplasm 13-F4-G5 IgG1 + — 100-1000 +, in cytoplasm — 15-B11-D10 IgG1 — NT* 100-1000 +, in cytoplasm — 16-G3-C3 IgG1 — NT 100-1000 — Surface ___________________________________________________________________________ *NT, not test
Legends of Figures
Fig. 1. Different profiles of anti-platelet binding with various mAb derived from dengue
virus-infected mice. Human platelet-rich plasmas in sodium EDTA was fixed with 1%
formaldehyde in PBS at room temperature for 10 min. Platelets (2.5 x 106/0.1 ml) were incubated with various monoclonal antibodies, followed by second goat anti-mouse IgG FITC-conjugated antibody. The binding of platelets was analyzed by FACScan at 488 nm.
Fig. 2. 13-F4-G5 mAb recognizes dengue virus NS-1 protein. (A) C6/36 was infected with dengue virus at m.o.i. =1 for 5 d. Cell lysate were prepared in RIPA buffer, and ran for western blot analysis with 13-F4-G5 mAb. (B) BHK cells infected with dengue virus at m.o.i.=5 for 48 h was stained intracellularly, cells sequentially treated with 4%
paraformaldehye and 0.1% saponin were incubated with 13-F4-G5 mAb. Secondary antibody was goat anti-mIgG-FITC conjugate. Shadow represents the background of second antibody conjugate. (C) BHK cells grown in Lab-Tek chamber slide was infected with dengue virus or herpes simplex virus at m.o.i.=1 for 48 h. 13-F4-G5 mAb was used to detect dengue infected cells. Goat anti-mIgG peroxidase conjugate was developed with an AEC substrate, the slide was counterstained with 1% Evans blue.
Fig. 3. Recombinant NS-1 inhibits the platelet binding of 13-F4-G5 dose-dependently. Purified mAb 13-F4-G5 (14 g) was incubated with various amounts of recombinant NS-1 for 30 min, then platelets (2.5 x 106/0.1 ml) were added, and the platelet binding was determined as described above. Percentage of inhibition and intensity of mean fluorescent intensity were presented. Control is the background of second antibody conjugate.
Fig. 4. 13-F4-G5 preferentially recognizes thrombin-activated platelets. (A) Platelets were activated by thrombin (1 U/ml). After 5 min of incubation, platelets lysate was prepared in RIPA buffer, and ran for western blot analysis with 13-F4-G5 mAb. (B) Thrombin (1 U/ml) activated or untreated platelets were incubated with 13-F4-G5, then were fixed with 1% formaldehyde. Second goat anti-mouse IgG FITC-conjugated antibody was added to detect the binding on FACScan. Shadow represents the background of second antibody conjugate.
Fig. 5. Flow cytometric analysis of various mAb binding to dengue virus infected BHK cells. BHK cells were infected with dengue virus at m.o.i.=5 for 48 h. The binding with various monoclonal antibodies was detected on FACScan. Both surface and intracellular staining were determined as described in Materials and Methods. Shadow represents the
background of second antibody conjugate.
were incubated with various monoclonal antibodies in the presence of rabbit complement (1:25 dilution) for 4 h. The lactase dehydrogenase released from damaged platelet was determined with OD450. Percentage of cytotoxicity was presented.
Fig. 7. Enhancement of ADP-induced platelet aggregation by 13-F4-G5. Platelet-poor plasma was prepared as described in Materials and Methods. Different concentrations of 13-F4-5 were incubated with PPP for 10 min. ADP (5 M)-induced platelet aggregation was detected with the aggregometer. 13-F4-5 (500 g) alone would not induce platelet aggregation, addition of 12 g goat anti-mIgG (GAM) has also no effect.
Fig. 1. Different profiles of anti-platelet binding with various mAb derived from dengue
Fig. 2. 13-F4-G5 mAb recognizes dengue virus NS-1 protein.
A. B.
Fig. 3. Recombinant NS-1 inhibits the platelet binding of 13-F4-G5 dose-dependently. 0 20 40 60 80 100 0 0.5 1 5 10 20 2' Ab NS1 (g) % inhi bi ti on 0 50 100 150 200 250 300 M F I inhibition MFI
Fig. 4. 13-F4-G5 preferentially recognizes thrombin-activated platelets.
A No Thrombin
Fig. 5. Flow cytometric analysis of various mAb binding to dengue virus infected BHK cells.
16-G3-C
11-F6-G
15-B11-D1
5-E4-C
13-F4-G
8-F1-B
3-D7-D
計畫成果自評: 本計劃在設立能因登革病毒感染可引起登革出血熱及登革休克症候群的動物感染上,其進 展相當緩慢,主要原因是由登革病毒感染至疾病有很多步驟,每一步驟都有它自己的調節 系統在控制,使其發展不致過度惡化,如果登革病毒不能在小鼠內快速複製,則不易啟動 此反應,因此不容易經由單一的注射登革病毒,就會引起登革出血熱/登革休克症候群。但 經由此研究,使我們瞭解登革病毒的 NS-1 可和血小板形成交? 反應,抗 NS-1 抗體會結合 血小板,造成血小板的破壞,這提供一個新的看法,可以讓我們對登革病毒感染和自體抗 體的關係做進一步的探討,對發展能造成登革出血熱及登革休克症候群的動物模式也刺激 新的想法。