國立臺灣大學生物資源暨農學院植物病理與微生物學研究所 碩士論文
Department of Plant Pathology and Microbiology College of Bioresources and Agriculture
National Taiwan University Master Thesis
木瓜輪點病毒嚴重嵌紋壞疽系統及嚴重嵌紋系統感染性選殖 株之構築及不同系統間之比較
Construction of infectious clones from two strains of Papaya ringspot virus (PRSV) and comparative studies among different
isolates of PRSV
楊瑞春 Jui-Chun Yang
指導教授:洪挺軒 博士 Advisor: Ting-Hsuan Hung, Ph.D.
中華民國 101 年 8 月
August, 2012
ii
誌謝
能夠完成我在研究所的學業,實在是得益於身邊諸多親友的幫忙。首先要感 謝洪挺軒老師的指導,除開學術及實驗以外,也拓展我人生中的視野。感謝蘇鴻 基老師、洪挺軒老師、張龍生老師以及眾位學長姐在木瓜研究上所奠定的詳實基 礎,讓我能在論文撰寫過程中獲益良多。感謝口試委員張龍生老師及葉信宏老師
對我論文的指正及建議,與洪老師為論文內容悉心斧正。感謝119 室所有的成員,
宜霞學姐當初費心帶領我實驗入門,學姐亭君、小瑩和心玥的木瓜經驗談;謝謝 雅智學姐及雋軼的時刻相伴與討論,雅智學姐在實驗、平日事務上予以提點及協 助,還有雋軼總是不負所託地幫我做細菌培養,於此特別對他們致上感激之意;
學弟獻廣和秉祜在病徵辨認的協助讓我的實驗紀錄更加精確;實驗室三人組日常 上偶爾幫忙,以及與其他學弟妹相處之時也常帶給我歡樂氣氛。感謝葉信宏老師 實驗室時常提供各式酵素救急,正恩學長、Russ 學長在序列分析軟體使用上的建 議及解惑,還有其他成員的陪伴。感謝曾在實驗上給予我建議的眾多人士,學弟
逸軒借用酵素簡省我許多時間,各家廠商也不吝解答我的疑問。感謝b93 的同學
在這段時間的陪伴以及敦促,希望大家情誼長存。感謝我所認識的所有朋友,祝 福所有人海闊天空。
最後要感謝我的父母和所有家人,提供我不虞匱乏的人生、並支持我、包容 我至今的一切,在此獻上我誠摯的感謝。
iii
中文摘要
木瓜輪點病由木瓜輪點病毒(Papaya ringspot virus, PRSV)所引起,可視為影響木瓜
產業最重要的病害。臺灣於1975 年首次發現此病,病毒歷經數十年的演替至今田
間所存的PRSV 依病徵表現可細分為數種系統(strains),包括引起葉片出現嚴重嵌 紋病徵之嚴重嵌紋系統(severe mottling, SM strain),致使葉片嚴重嵌紋及畸形扭曲 之畸形系統(deformation, DF strain),以及造成葉片嚴重嵌紋並伴隨嚴重壞疽之嚴重 嵌紋壞疽系統(severe mottling wih necrosis, SMN strain)。臺灣田間的 PRSV 目前以 DF 系統最常見;而 SMN 系統則在冷熱交替之際造成木瓜植株快速萎凋,危害性 較高而被視為最具威脅性的系統。三種病毒系統雖具相當之序列相似性,但在木 瓜上呈現的病徵卻有明顯的差異,在分子學及病理學上頗有研究意義。本論文將 三系統之病毒基因體序列以two-step RT-PCR 各自增幅出 3 到 4 個 cDNA 片段,配 合片段間頭尾重疊區域剪接構築出全長感染性選殖株,並且將三種系統選殖株利 用胞外轉錄(in vitro transcription)合成完整的病毒 RNA transcripts,機械接種於木瓜 及白藜寄主。結果發現人工轉錄的病毒RNA transcripts 可感染木瓜並在葉部產生 典型病徵,但發病時間比原自然的病毒株約延遲一週;若將病葉汁液進行繼代接 種,則發病的時間和產生的病徵都與原自然的病毒株近似。將三種病毒系統在木
瓜呈現的病徵與過往記錄相比較,發現SMN 系統在後期出現絲狀葉,顯示其病毒
可能發生變異而導致病徵改變。經核苷酸定序後將三系統之全長選殖株序列進行 比對,發現以SM 系統與 DF 系統相似度 97.3%為最高,SMN 系統與兩者比對皆 為96.5%;各基因核苷酸序列相似度大都約在 96-99%,但 P3 基因的相似度稍低,
在95.8-96.7%,P1 基因低於 95%,5’-UTR 則在 93.0-94.2%。與各地已發表之 PRSV 分離株全長核酸序列進行比對,三系統與其他源自臺灣之分離株以及韓國分離株 較為近似,並與中國、泰國之分離株歸於同一關係群;與美洲和南亞地區分離株
親緣較遠,切合PRSV 的序列歧異度與分布地域相關之推論。本論文已成功獲得
iv
三種系統具感染力之全長cDNA 選殖株,對後續找尋病徵決定因子(symptom determinants)將可提供極大幫助,例如可針對系統間差異性較大的基因序列如 5’-UTR、P1、P3 進行重組置換,或針對特定基因如 HC-Pro 或 CI 進行序列變換,
藉以探討其致病機制。另外,由於先前所設計的DF 系統之專一性引子對經常也會
在SM 系統的 RT-PCR 試驗中出現微量增幅現象,本論文另行設計新的 DF 系統專
一性引子對,靈敏度與專一性俱佳,對日後PRSV 系統鑑別將更有助益。
關鍵字:木瓜輪點病毒;感染性選殖株;病毒系統
v
Abstract
Papaya ring spot caused by Papaya ringspot virus (PRSV) is considered to be the most important disease impacting papaya industry. This disease has occurred in Taiwan for decades since 1975. After the long-term genomic evolution in the field, there are 3 major pathological strains of PRSV in Taiwan at present time such as
SM (severe
mottling), DF (severe mottling with leaf-deformation) and SMN (severe mottling
with necrosis and quick decline) strains. DF is a predominant strain of PRSV in
the field of Taiwan now, and SMN should be the most destructive strain causing
quick decline in papaya plants especially when the season changes. SM, DF and
SMN strains have highly similar nucleotide sequences, but their incited
symptoms in papaya hosts are different. To investigate the relationship between
molecular and pathological characters, this thesis was dedicated to construct the
infectious clones for SM, DF and SMN strains of PRSV. Each intact infectious
clone was derived from the splicing of 3~4 amplified cDNA fragments with
two-step RT-PCR. The artificial full-length PRSV RNA transcripts could be harvested from theinfectious clones through in vitro transcription, and they were used in the
inoculation tests in either papaya orChenopodium quinoa hosts
. The results showed that the PRSV RNA transcripts derived from infectious clones could successfully infect papaya plants. However, the symptoms caused by the transcripts appeared 1 week later than those caused by natural PRSV inocula. The developing time of symptoms produced by the transcript-infected sap was similar to those caused by natural PRSV. The present SMN strain might incite “fern-leaf” in addition to original symptoms, which indicated mutation probably occurred in the genome of PRSV-SMN. Based on the alignment of full-length genomic nucleotide sequences among 3 different PRSV strains, SM and DFvi
have 97.3% homology; SM and SMN have 96.5% homology; and SM and DF have 96.5% homology. Individual gene analyses revealed that they were 95.8-96.7%, 94~95% and 93.0-94.2% similarity of nucleotide sequences in P3, P1 and 5’-UTR respectively. The rest of genes showed 96-99% similarity among 3 strains. The further alignment of full-length genomic nucleotide sequences among several PRSV isolates from different countries demonstrated that all isolates from Taiwan are close to those from Korea and they can be categorized into a phylogenic group with those from China and Thailand. The PRSV isolates from America and South Asia form another phylogenic group. It indicates that strain-diversity of PRSV corresponds with geographical difference. Constructions of infectious clones of 3 PRSV strains will be helpful to study key genes or regions associated with symptom determinants (such as 5’-UTR, P1, P3, HC-Pro and CI) through molecular recombination and substitution.
Besides, a new primer pair specific to the DF strain was developed in this thesis to improve the sensitivity and specificity and avoid non-specific amplification in the RT-PCR assay of PRSV-SM. It will provide a more reliable method for the identification among different PRSV strains.
Keywords: papaya ringspot potyvirus; infectious clone; virus strain
vii
目錄
口試委員會審定書 i
誌謝 ii
中文摘要 iii
英文摘要 v
目錄 vii
表目錄 ix
圖目錄 x
壹、前言 1
貳、前人研究 4
一、木瓜輪點病毒分類及分子特性 4
二、木瓜輪點病毒之發生與危害 5
三、木瓜輪點病毒之傳播 6
四、木瓜輪點病毒之寄主範圍及病徵 6
五、臺灣境內木瓜輪點病毒系統及分布概況 7
六、木瓜輪點病毒各地病毒株之序列及其流變 8
七、Potyvirus 基因體及各基因區功能 10
參、材料與方法 15
一、試驗植物準備 15
二、木瓜輪點病毒系統來源與保存 15
三、木瓜輪點病毒之偵測方法 16
1. 病毒核酸萃取 16
2. Two-Step RT-PCR 16
3. One-Step RT-PCR 17
viii
4. PCR 產物電泳分析 17
四、木瓜輪點病毒 PRSV-SMN、PRSV-SM 之感染性選殖株構築 18
1. PRSV 核酸序列通用性引子對設計 18
2. Conventional PCR 18
3. PCR 產物之選殖 19
4. PRSV 序列全長選殖株之合成 20
五、胞外轉錄 (in vitro transcription) 21
六、接種試驗 21
七、選殖株之基因序列分析 21
肆、結果 23
一、木瓜輪點病毒基因體核酸片段增幅與選殖 22
二、全長選殖株之合成及胞外轉錄試驗 22
三、胞外轉錄感染力試驗及不同系統選殖株在寄主之病徵表現…………...23
四、偵測結果及新專一性引子對設計 25
五、三系統全長選殖株與以發表 PRSV 分離株之序列比對分析 25
伍、討論 28
陸、參考文獻 34
柒、圖與表 45
捌、附錄 67
ix
表目錄
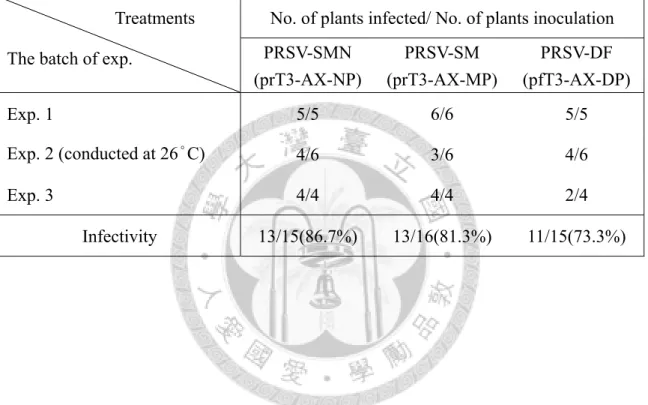
表一、木瓜輪點病毒(Papaya ringspot virus)基因體核酸片段之選殖所用引子對及其 基因產物 46 表二、由木瓜輪點病毒cDNA 選殖株所合成的 RNA transcripts 機械接種木瓜寄生
之感染率分析 47 表三、由木瓜輪點病毒cDNA 選殖株所合成的 RNA transcripts 機械接種木瓜寄主
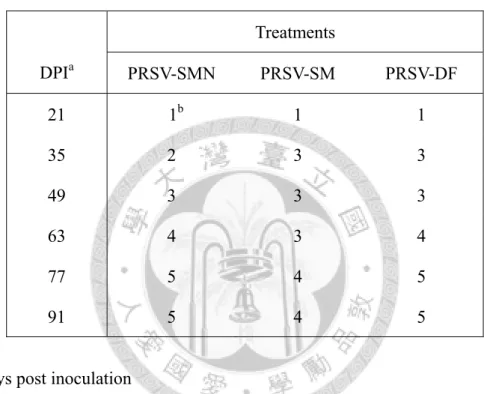
後之病徵發展 48 表四、由木瓜輪點病毒cDNA 選殖株所合成的 RNA transcripts 感染木瓜後,再將
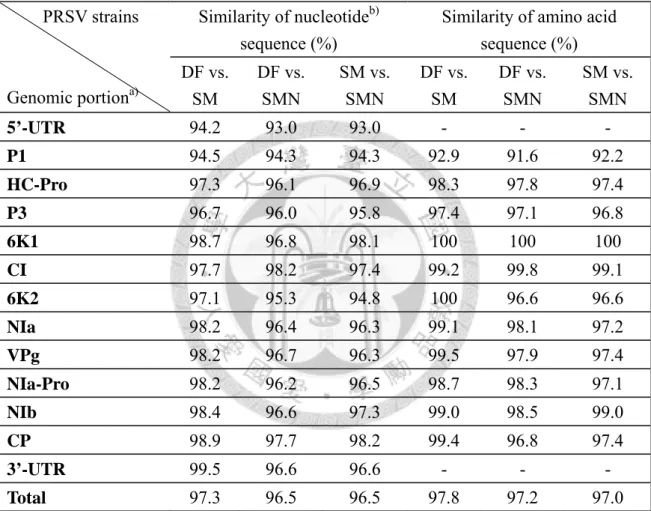
染病汁液繼代接種於木瓜寄主之病徵發展 49 表五、木瓜輪點病毒機械接種木瓜寄主後之病徵發展 50 表六、PRSV 三系統全長 cDNA 選殖株之全長基因體與個別基因的核苷酸及胺基酸 序列相似度分析 51
x
圖目錄
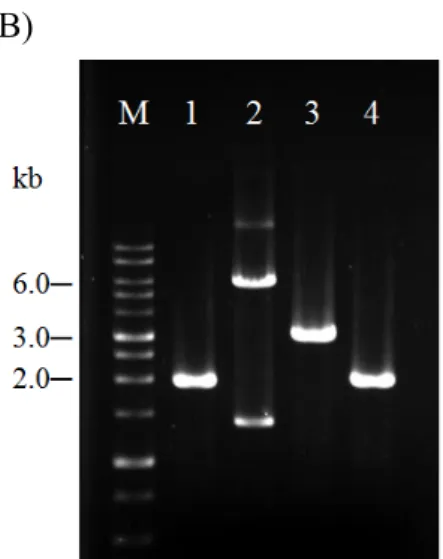
圖一、木瓜輪點病毒(PRSV)各系統分離株全長基因體序列選殖之策略 52 圖二、PCR 增幅之木瓜輪點病毒基因體核酸片段 53 圖三、木瓜輪點病毒(PRSV)全長基因體 cDNA 選殖片段以 XbaI 進行酵解後之電泳
分析 54 圖四、木瓜輪點病毒(PRSV)全長 cDNA 選殖株之胞外轉錄 RNA 產物電泳分析 55 圖五、木瓜輪點病毒嚴重嵌紋壞疽系統(PRSV-SMN)選殖株 pN-P、pN-A 胞外轉錄
產物感染台農二號木瓜28 天、70 天、84 天、98 天後之病徵表現 56 圖六、木瓜輪點病毒嚴重嵌紋系統(PRSV-SM)選殖株 pM-P 胞外轉錄產物在台農二
號木瓜於接種後28 天、70 天、84 天、98 天之病徵表現 57 圖七、木瓜輪點病毒畸形系統(PRSV-DF)選殖株 pD-P、pD-T 胞外轉錄產物在感染
台農二號木瓜28 天、70 天、84 天、98 天後之病徵表現 58 圖八、不同木瓜輪點病毒(PRSV)系統感染台農二號木瓜後期之全株外部病徵 59 圖九、PRSV 三系統之原病毒對照組在台農二號木瓜接種後 35 天之病徵表現 60 圖十、PRSV 三系統選殖株致病植株之繼代接種在台農二號木瓜引起之病徵 61 圖十一、由木瓜輪點病毒(PRSV) cDNA 選殖株所合成的 RNA transcripts 感染木瓜
後,再將染病汁液繼代接種於白藜寄主所產生之病徵 62 圖十二、針對各種木瓜輪點病毒(PRSV)之 cDNA 選殖株以 PRSV 通用型引子對及
系統專一型引子對進行RT-PCR 偵測 63 圖十三、新設計專一型引子對DF-626 於不同反應黏合溫度之木瓜輪點病毒(PRSV)
偵測效果比較 64 圖十四、由neighbour-joining 法建立各地區木瓜輪點病毒(PRSV)分離株基因體全長
核酸序列之親源樹狀關係圖,以 Potato virus Y 為外群 65 圖十五、以木瓜輪點病毒(PRSV) DF 系統為代表與世界各地區 PRSV 全長基因體
核苷酸序列之相似度比對 66
1
壹、前言
木瓜(Carica papaya Linnaeus,番木瓜)屬十字花目番木瓜科(Caricacrae),原產 於熱帶美洲,最初於墨西哥栽培。在17 世紀傳入亞洲,19 世紀末由中國引進臺灣,
由於木瓜栽培容易、栽植到結果僅需1 年,短期內就成為臺灣重要經濟果樹之一。
木瓜因其風味甜美,且富含木瓜酶、木瓜鹼、胺基酸及維生素,具有很高的食用 價值,廣為大眾市場接受。木瓜栽培範圍涵蓋全球熱帶地區,在中南美洲、亞洲、
非洲皆有大量種植,消費市場遍及世界各國。據世界糧農組織 (Food and Agriculture Organization, FAO) 統計,2010 年全球木瓜種植面積為 44 萬公頃,產量約 1,121 萬噸,每公頃產量約25 噸,以印度產量最高(42%),其次為巴西(17%)、奈及利亞 (6%)、印尼(6%)及墨西哥(5%)。而木瓜在臺灣可全年生產,盛產期為 11 月份,近 年種植面積為3,294 公頃,產量 129,322 噸,每公頃產量 41 噸,產地以台南縣、
屏東縣、高雄縣、嘉義縣為主(99 年農業統計年報, 2010)。
危害台灣的木瓜病害包括真菌類之木瓜白粉病(powdery mildew, Acrosporium
caricae Noack)、疫病(phytophthora blight, Phytophthora palmivora Butler, P.
parasitica Dastur)、炭疽病(anthracnose, Glomerella cingulata)、黑點病(leaf spot, Asperisporium caricae Speg.);細菌類之黑腐病(black rot, Erwinia cypripedii Hori);
線蟲根瘤病(root knot, Meloidogyne spp.);病毒類之輪點病(ringspot, Papaya ringspot
virus, PRSV)、捲葉病(leaf curl, Papaya leaf curl virus, PaLCV)、畸葉嵌紋病(leaf
distortion and mosaic, Papaya leaf distortion mosaic virus, PLDMV)等。目前木瓜輪點 病當屬最重要的木瓜病害,且病毒感染後無藥可醫,使得多年生木瓜果樹淪為一 年生作物,是木瓜栽培業的最大限制因子。在台灣防治輪點病的方法主要為利用 網室栽培阻擋蚜蟲媒介傳播,以及栽植抗或耐木瓜輪點病的木瓜品系。隨分子育 種技術的進步,1998 年夏威夷開始商業栽培 Rainbow 及 SunUp 兩種轉基因木瓜 (transgenic papaya; genetically modified papaya, GM-papaya)品種(Gonsalves, 2004),2
國內則有學者利用農桿菌技術轉殖病毒鞘蛋白基因,育成抗木瓜輪點病毒轉基因 木瓜(Cheng et al., 1996; Bau et al., 2004)以及雙重抗木瓜輪點病毒及木瓜畸葉嵌紋 病毒的轉基因木瓜(Kung et al., 2009)。此外,過去也有以傳統育種法選拔出相對較 耐病的木瓜品種如台農5 號(Tainung No. 5, TN 5)與紅妃(Red Lady 786),近年來更 有台大園藝系選育出的高度耐病品種台大8 號(NTU 8)等(王, 1997;張, 1995;張 等,2006;Chang and Chung, 1992)。
木瓜輪點病毒 (Papaya ringspot virus, PRSV)隸屬於 Potyviridae (馬鈴薯 Y 病毒 科)、Potyvirus(馬鈴薯 Y 病毒屬),病毒顆粒外觀為長絲狀,大小約 760-800 × 12 nm,
基因體由約10kb 之單股(single strand)正譯性(positive sense)RNA 分子組成(De la Rosa and Lastra, 1983),可轉譯一個較大的聚合蛋白(polyprotein),並自行水解獲得 P1、HC-Pro、P3、6K1、CI、6K2、NIa、NIb 與 CP 等基因產物(Yeh et al., 1992),
近年又發現Potyviridae 另有一小分子病毒蛋白產物 P3N+PIPO (Chung et al., 2008)。PRSV 以其感染的寄主範圍可分為 PRSV-P 與 PRSV-W 兩種生物型
(biotypes),PRSV-P 能感染番木瓜科、葫蘆科及藜科,PRSV-W 只能感染葫蘆科與 藜科(Purcifull et al., 1984; Yeh et al., 1984),兩者在血清學上無法區分,序列比對之 相似度也極高(Bateson et al., 1994; Purcifull et al.,1984; Yeh et al., 1984);近來序列
資料增加,分析不同地域之PRSV 病毒分離株序列,也發現了決定寄主範圍的關
鍵因子位於NIa-Pro,其特定胺基酸經點突變後可使 PRSV-W 之單點突變病毒株感 染木瓜(Chen et al., 2008)。
臺灣田間的PRSV 按病毒在木瓜植株上產生之病徵型態區分為數種系統
(strains),如畸形系統(deformation, DF)、嚴重嵌紋系統(severe mottling, SM)、嚴重 嵌紋壞疽系統(severe mottling with necrosis, SMN)等(吉井,1986;林,1980;翁,
1981;廖,2004;蔡,1995)。不同系統間的病徵表現互有分別,環境與寄主的變 化亦有所影響(林,1980;廖,2004;蔡,1995)。早期田間流行系統與近來的系統
在生態組成上已有明顯變動,以往不易見到的DF 系統目前已相當普遍,SM 系統
3
則逐漸減少(廖,2004),顯示 PRSV 各系統在田間有大幅度的變化動態。本實驗室
保存了臺灣三個常見的PRSV 系統,在全長序列的比對分析可發現三系統間有高
達96-97%的相似度,然而仍保有相異之處。由前人對不同片段的序列比對中,發
現5’-UTR 的相似度在所有序列中最低,相似度只有 91.8-92.9%,P1 在所有基因中 相似度最低,為94.5-94.8%(李,2006;廖,2004);HC-Pro 在 Potyvirus 多項研究 中,證實其對於病徵發展及寄主互動上有相當程度的重要性(Dunoyer et al., 2004;
Kasschau et al., 1997; Mallory et al., 2002; Shiboleth et al., 2007; Yu et al., 2006)。為瞭
解病毒基因體序列與寄主病徵表現的關聯性,可藉由構築出PRSV 不同系統之全
長感染性選殖株(infectious clone)以進行研究,DF 系統之感染性選殖株已成功構築 並完成接種試驗(劉,2008),但其產生的病徵不顯著而有待改良。本論文接續構築 SM 與 SMN 系統的全長感染性選殖株,並也完成新的 DF 系統感染性選殖株之建 構,以利於將來對病毒序列進行基因操作,分析序列更動對病毒基因產物及寄主 因子產生之效應,在分子層次探討病原寄主間生理、病理的相關性。
4
貳、前人研究
一、木瓜輪點病毒分類及分子特性
在 ICTV (International Committee on Taxonomy of Viruses)分類系統中,PRSV 被劃分於Potyviridae(馬鈴薯 Y 病毒科)、Potyvirus(馬鈴薯 Y 病毒屬)之下,病毒顆 粒為長絲狀,長約760-800 nm,寬約 12 nm(De Bokx, 1965; 林,1980),基因體為 單股(single strand)正譯性(positive sense) RNA(De la Rosa and Lastra, 1983; Yeh and Gonsalves, 1985),大小約 10,317-10,335 個核苷酸(nts)。本實驗室保存之病毒分離 株總長為10,326 nts,有一個開放閱讀框架(open reading frame, ORF),由 nt 86 至 nt 10,120 區間可轉譯出 3,344 個胺基酸的聚合蛋白(polyprotein),並自行水解成數 種病毒蛋白,依序為P1 (63K)、HC-Pro (helper component-proteinase, 52K)、P3 (46K)、6K1 (6 kDa 1 protein, 6K)、CI (cylindrical inclusion protein, 72K)、6K2 (6 kDa 2 protein, 6K)、NIa (nuclear inclusion protein a, 48K)、NIb (nuclear inclusion protein b, 59K)與 CP (coat protein, 35K),NIa 又可水解成 VPg (genome linked viral protein, 21K) 及NIa-Pro(27K)( Gonsalves and Ishii, 1980; Purcifull and Edwardson,1967; Yeh and Gonsalves, 1985)。鹼基組成以 adenine 最多(31.2 %),其次為 uracil (27.0%)、guanine (23.8%)及 cytosine (18.0%)。5’-UTR(5’-untranslated region)共 85 nt,序列為 AU-rich 並以AUG 為轉譯起始碼;終止碼為 UAA,接續 206 nt 之 3’-UTR(3’-untranslated region) (Yeh et al., 1992)。Chung et al.於 2008 年發現 Turnip mosaic virus(TuMV)P3 基因區有一小段overlapping ORF,其 N 端之胺基酸序列與 P3 蛋白相同,中途則 讀框移位(frameshifting) 改以+2 閱讀框架(reading frame)轉譯,表現之基因產物命 名為PIPO (Pretty Interesting Potyviridae ORF, ~25K)。
依據 CMI/AAB 記載,PRSV 的熱不活化溫度(thermal inactivation point, TIP)為 54-60˚C,耐稀釋臨界點(dilution end point, DEP)為 10-3-10-4,生體外殘存時間 (longevity in vitro, LIV)約 8 小時(Purcifull et al.,1984)。
5
以PRSV 感染寄主之範圍,可分為兩種生物型(biotypes):木瓜輪點病毒木瓜 型(Papaya ringspot virus type papaya, PRSV-P)及木瓜輪點病毒西瓜型(Papaya
ringspot virus type watermelon, PRSV-W),PRSV-P 可感染番木瓜科(Caricaceae)、藜
科(Chenopodiaceae)及葫蘆科(Cucurbitaceae)作物,PRSV-W 無法感染番木瓜,但可 感染藜科與葫蘆科(Purcifull et al., 1984; Yeh et al., 1984),兩者間無法以血清學鑑別 差異(Bateson et al., 1994; Purcifull et al.,1984; Yeh et al., 1984)。二、木瓜輪點病毒之發生與危害
木瓜輪點病在1945 年證實由 PRSV 引起(Jensen, 1949),此病害最早紀錄可溯 及1929 年 Smith 氏於牙買加發現有類似病徵之木瓜病害,包括葉片輕微嵌紋、黃 化、新葉變小、老葉脫落等,而後在夏威夷歐胡島(Oahu island)陸續有相仿病害報 導,病徵除葉片嵌紋、幼葉捲曲,尚記錄嚴重受害病株有植株矮化現象、果實減 少或結有畸形果,並在果皮上造成中央為綠色之黃緣輪點病斑,病害命名由此而 來(Parris, 1938; Lindner et al., 1945),其後由 Jensen 氏鑑定為病毒所造成,正式定 名為木瓜輪點病毒(Papaya ringspot virus, PRSV)。PRSV-P 普遍發生於熱帶及亞熱 帶栽植有木瓜之地區,包括美國(Conover, 1962)、南美洲(De La Rosa and Lastra, 1983; Herold and Weibel, 1962; Smith, 1929)、加勒比海地區(Adsuar, 1947; Story and Halliwell, 1969)、非洲(Lana, 1980)、印度(Capoor and Varma, 1948)、泰國(Srisomchai, 1975)、臺灣(王等,1978)、日本、中國、菲律賓、澳洲(Thomas and Dodman, 1993) 及太平洋地區(Cook and Milbrath, 1971; Davis et al., 2005; Parris, 1938)皆有病害發 生之記錄。
PRSV-W 早先以 Watermelon mosaic virus 1 (WMV-1)命名(Webb and Scott, 1965),在血清學關係上與 PRSV-P 無法區分(Purcifull et al., 1984)。PRSV-W 侵襲 各地之葫蘆科作物,在美國(Webb et al., 1965)、墨西哥(Milne et al., 1969)、加勒比 海地區、澳洲(Greber, 1978)、歐洲、印度、東南亞、中東地區(Russo et al., 1979)
6
以及南美洲等地陸續有此病害報導(Purcifull et al., 1984; Yeh et al., 1984)。
國內於1975 年首度報導 PRSV-P 之發生記錄,在高雄縣燕巢、大樹和阿蓮鄉 發現病徵與輪點病相似之木瓜株(王等,1978;陳等,1976),經由陳等(1976)以電 子顯微鏡觀察病株組織,發現長約700 nm 之絲狀病毒顆粒,研判此病害應為 PRSV 所造成,在數年內疫情擴散至全台主要木瓜產地,使木瓜產業損失慘重(王等,
1978)。1987 年許氏等及張氏等調查國內瓜類作物病毒病時,發現 PRSV-W 在多種 瓜類發生比率高且分布甚廣,危害不容輕忽。
三、木瓜輪點病毒之傳播
PRSV 屬於嵌紋型(mosaic-type)病毒,傳播方式可經由機械摩擦及蚜蟲取食病 株(Adsuar, 1947),將帶有病毒之植物汁液接種至健康寄主。大部份學者認為 PRSV 無法經種子傳播(王等,1978;Capoor and Varma, 1948; Ishii and Holtzmann, 1963)。
PRSV 田間主要是藉媒介昆蟲以非永續性(non-persistent)方式傳播(Purcifull et al., 1984),PRSV-W 可由 15 屬 24 種蚜蟲傳播(Webb et al., 1965; Purcifull et al., 1984);
PRSV-P 可由 11 屬 21 種蚜蟲傳播(Jensen, 1949; Conover, 1964; Zettler et al., 1968),
以桃蚜(Myzus persicae Sulzer.)、棉蚜(Aphis gossypii Glover.)傳毒能力最佳,桃蚜獲 毒時間只需15-45 秒(Milne et al., 1969),在 1 小時內皆可保留傳毒能力,且蚜蟲數 越多,傳播率也會相對提高,因此蔓延速度主要取決於田間有翅蚜蟲密度之高低,
及病株之多寡與距離(Jensen, 1949; Conover, 1964; Zettler et al., 1968)。
四、木瓜輪點病毒之寄主範圍及病徵
PRSV 的寄主範圍涵蓋番木瓜科、藜科與葫蘆科,在番木瓜科及葫蘆科植物上 引起系統性感染,在藜科上則造成局部病斑(necrotic local lesions) (Purcifull et al., 1984)。PRSV-P 在傳播試驗中可感染 3 科 15 種植物(Jensen, 1949; Conover, 1964);
PRSV-W 無法感染木瓜,但可感染葫蘆科 11 屬及藜科 2 屬等 38 種植物(Purcifull et
7
al., 1984; Yeh et al., 1984)。PRSV-P 在木瓜上的病徵會因感染階段、植株生長勢、
株齡、環境及病毒系統之別而異。一般在葉片上起初會造成葉脈透化,後期會出 現葉片斑駁、嵌紋、畸形或黃化,葉片面積變小、葉肉組織縮減;葉柄及幼莖上 會有水浸狀條斑,幼葉常會皺縮或畸形;病株生長受阻及矮化,部分系統造成老 葉枯黃脫落只餘頂端新葉,或是自新葉萎凋而後全株死亡;不易開花結實,花瓣 上亦出現水浸狀輪紋(王,1995;Conover, 1964; Jensen, 1949; Purcifull et al., 1984)。
果實發育不良或畸形,表皮會有斑點、輪紋,且產量及品質明顯下降(Yeh et al., 1984)。感染 PRSV 之木瓜組織以電子顯微鏡觀察,可發現細胞質中有風車狀 (pinwheels)、束狀(bundles)及環狀(circulars)內含體(inclusions) (Zettler et al.,1968)。
在葫蘆科寄主矮南瓜(Cucurbita pepo L.)上,PRSV-P 部份強毒株可造成明顯的 葉片嵌紋及畸形、弱毒株則引起葉片輕微斑駁,PRSV-W 亦會引起葉片嵌紋、畸 形病徵;刺角瓜(Cucumis metuliferus cv. Accession 2459)、胡瓜(Cucumis sativus L.)、
蛇瓜(Trichosanthes anguina L.)與西瓜(Citrullus vulgaris S.)感染後會引起葉片斑 駁、嵌紋和葉脈壞疽(王等,1978;林,1980;Gonsalves and Ishii, 1980)。兩種生 物型皆可在指示植物紅藜(Chenopodium amaranticolor Coste and Reyn.)及白藜 (Chenopodium quinoa Wild.)接種葉造成局部病斑(Gonsalves and Ishii, 1980; Russo et
al., 1979; Yeh et al., 1984)。試驗上白藜多用於進行病毒單斑分離(蔡,1995;廖,
2004;Yeh et al., 1984),葫蘆科寄主在媒介昆蟲傳播試驗的效率較佳。
五、臺灣境內木瓜輪點病毒系統及分布概況
在臺灣地區的研究中,可依據PRSV-P 在木瓜上產生不同病徵型態,再區分為 數種系統(strain),目前田間主要有嚴重嵌紋系統(severe mottling, SM)、嚴重壞疽嵌 紋系統(severe mottling with necrosis, SMN)及畸形系統(deformation, DF),早期尚有 輕微嵌紋系統(mild mottling, M)、嚴重嵌紋畸形系統(severe mottling with
deformation, SMd)(吉井,1986;林,1980;翁,1981;關,1990)。嚴重嵌紋系統
8
(SM strain)接種木瓜後,於葉上產生嚴重嵌紋病徵,葉片明顯變小且低溫下出現畸 形葉,葉、莖、果實有水浸狀病斑;於白藜上產生褪色斑點之病徵,矮南瓜(C. pepo) 上則會產生葉脈透化、嵌紋病徵。嚴重嵌紋壞疽系統(SMN strain)造成罹病木瓜葉 片變小,於葉上產生嚴重嵌紋與細小壞疽斑點,下位葉快速黃化導致脫落,在高 低溫反覆交替後出現急速萎凋之病徵,是危害性最強系統;於白藜上產生的病徵 為褪色斑點,矮南瓜上則會產生輕微葉脈透化的病徵。畸形系統(DF strain)會造成 罹病木瓜葉片明顯變小,產生嚴重嵌紋及明顯畸形病徵,綠島(green islands)病徵明 顯,在感染後期出現絲狀葉;於白藜上產生的病徵為褪色性黃萎斑點,矮南瓜上 則會產生葉脈透化、嵌紋病徵。感染輕微嵌紋系統(M strain)之木瓜病株在低溫下 無病徵,高溫時葉片呈黃斑或輕微嵌紋;嚴重嵌紋畸形系統(SMd strain)會引起罹 病木瓜葉脈黃化、葉片萎縮、嚴重嵌紋及變形,因病徵類似故併入畸形系統(吉井,
1986;林,1980;翁,1981;廖,2004;蔡,1995)。木瓜輪點病自 1975 年於臺灣 南部發現後,在三十年間分化出數種系統,各系統互有消長:1978 到 1980 年間田 間以M 系統型病徵出現頻率最高,SMN 系統及 SM 系統較少;1980 年再度調查,
發現SM 系統佔有優勢,SMN 系統其次而 M 系統出現頻率減少(林,1980)。DF 系統至1986 年首次發現,往後十年內仍以 SM 系統佔大多數(吉川,1986;蔡,
1995),直至 2003 年發現 DF 系統有逐漸取代 SM 系統成為田間優勢系統之傾向 (廖,2004)。目前臺灣田間以 DF 系統較普遍,SM 系統族群數量略低,SMN 系統 雖然出現頻率低但病害嚴重度最高(廖,2004)。
六、木瓜輪點病毒各地病毒株之序列及其流變
目前 GenBank 共登錄 20 條各地 PRSV 病毒株基因體全長序列(附錄二),14 條 屬PRSV-P,分別來自夏威夷、泰國、臺灣、印度、墨西哥及中國;6 條為 PRSV-W,
來自臺灣、泰國、巴西、印度與韓國。PRSV-P 基因體長度在 10,317-10,332 nt,
PRSV-W 則為 10,323-10,335 nt。
9
將已發表全長序列之 PRSV 分離株進行基因序列比對分析,各地病毒株基因 體全長核酸序列相似度為83-89%,胺基酸序列則是 90-92%;病毒株之間核酸序列 變異度最高的區域為5’-UTR,相似度僅 64-84%,其次為 P1 序列,核酸及胺基酸 序列相似度為70-78%及 67-76%,而 polyprotein 其餘區域之核酸序列相似度在 85-94%、胺基酸序列相似度為 89-98%,3’-UTR 核酸序列相似度則是 92-96%
(Parameswari et al., 2007)。
而 PRSV 目前在臺灣常見的三個系統,DF、SM、SMN 做全長及個別基因序 列比對,發現三系統彼此之間核酸及胺基酸序列相似度達96-97%,其中 DF 與 SM 系統相似度為97.3%,SMN 系統與 DF、SM 系統相似度則為 96.8%及 97.0%。核 酸序列比對可發現在5’-UTR 相似度最低,只有 91.9-93.0%,P1 於 ORF 中核酸序 列相似度最低,為94.5-94.8%,胺基酸序列相似度為 92.2-93.1%。下游基因區之核 酸、胺基酸序列相似度為95-100%與 96-100%,3’-UTR 的核酸序列相似度則高達 99.5-100%(廖,2004;李,2006)。各區域的相似度趨勢大致上貼近其他 potyviruses 與PRSV 跨地域分離株的研究結果(Adams et al., 2005; Parameswari et al., 2007)。
將分散各地的PRSV 分離株進行序列與分子親緣分析(phylogenetic analysis),
可發現PRSV 之間的親緣關係與地域分布較具關連性,而非寄主感染能力(李,2006;
Bateson et al., 2002)。全長序列之親源樹狀關係圖(phylogenetic tree)將 PRSV 分為美 洲群與亞洲群,美洲群包括印度、墨西哥、巴西、夏威夷等地之分離株,亞洲群 則來自臺灣、泰國、中國、韓國。以基因歧異度最高的P1 比對,P1 胺基酸序列在 跨地域分離株間相似度為71%,同地域的 PRSV-P 與 PRSV-W 歧異度在印度為 16%、泰國為 18%、臺灣 8%;各地 PRSV-W 比對 P1 之歧異度為 10-35%,PRSV-P 則是0-35% (Bateson et al., 2002; Mangrauthia et al., 2008; Parameswari et al.,
2007)。PRSV 全長序列重組分析顯示 P1 區域是病毒基因重組發生熱點,也呼應 5’-UTR 及 P1 序列相較於其餘病毒基因有高度變異之現象(Noa-Carrazana et al., 2007; Mangrauthia et al., 2008)。以 CP 中央至 C 端區域(cCP)序列分析所得到的親
10
源樹狀關係圖推斷,PRSV 發源於印度及斯理蘭卡地區,先後發生兩次傳播事件:
PRSV-W 可能藉貿易活動,自南亞進入東亞及東南亞地區;發現新大陸後又隨航 海運輸傳入美洲並突變衍生PRSV-P。18 世紀初隨經濟作物大量推廣栽培,PRSV
進而散佈世界各處,在各地田間發生基因突變及重組,臺灣的PRSV 病毒株可能
源自泰國、墨西哥及夏威夷(Bateson et al.,2002; Gibbs et al., 2008; Mangrauthia et al., 2008)。
PRSV 包含兩種生物型,因全長序列資料陸續增加,對尋找決定寄主範圍的關 鍵因子亦有所斬獲,目前已發現其中之一位在NIa-Pro,PRSV-P 為 Lys2309、PRSV-W 是Asp2309,且進行Lys2309與Asp2309單一胺基酸置換確實可使PRSV-W 之單點突 變病毒株獲得感染木瓜能力,但一段時間後受試植株回復(recovery),且回復植株 汁液進行繼代接種(subculture)試驗,對健康木瓜不具感染能力(Chen et al., 2008)。
七、Potyvirus 基因體及各基因區功能
Potyviridae 的病毒屬有 Brambyvirus、Potyvirus、Ipomovirus、Macluravirus、
Tritimovirus、Rymovirus、Bymovirus,在植物病毒分類中佔 30%,是全球性重要植
物病原。以 Potato virus Y 命名的 Potyvirus 是大宗植物病毒屬,其下病毒種泛稱為 potyvirus,在自然界主要由蚜蟲非永續媒介傳播,部分也可利用種子傳播。本屬病 毒為長絲狀、單顆粒 (monopartile),長度約 680-900 nm、直徑約 11-15 nm,基因 體為正譯性、約8-12 kb 單股 RNA,5’端共價鍵結 VPg,3’端有一段 poly(A) tail。鹼基組成以adenine 比例最高(23-44%),uracil(15.6-30.9%)次之,guanine(21-26 %) 與cytosine(14.9-28%)較低。基因體兩端有非轉譯區(untranslated regions),中間的 ORF 會轉譯一條長約 340-370 kDa 的 polyprotein,另有 overlapping ORF 轉譯約 25 kDa 融合蛋白(fusion protein);polyprotein 可在轉譯過程中由具 proteinase 活性之 P1、HC-Pro 與 NIa-Pro 自行水解,獲得約 10 個病毒蛋白產物(Dougherty and Carrington, 1988)。
11
以下就 Potyvirus 基因體各區域特性及其於感染進程中擔負功能做簡單整理。
5’-UTR 長度約 100-200 nt,具兩段 cap-independent regulatory
elements(CIREs),可形成二級結構 pseudoknots (PK),其中 PK1 序列與 18S rRNA 互補,與病毒3’端 poly(A) tail 共同促進 cap-independent translation,保守性片段如 UUUCA 及 CAA repeats 也可能參與 cap-independent translation (Carrington and Freed, 1990; Niepel and Gallie, 1999; Zeenko and Gallie, 2005),一些分散小片段亦證實會促 進轉譯作用(Kawarasaki et al., 2000)。
P1 為 polyprotein 最前端的蛋白,序列及長度變異度最高(Aleman-Verdaguer et
al., 1997; Shukla et al., 1994),C 端 Ser-type proteinase 辨認並水解 P1 及 HC-Pro 間
切位 (Verchot et al., 1992; Yang et al., 1998)。在感染細胞中 P1 會與 CI 結合,也可 於膜器上發現(Rodríguez-Cerezo and Shaw, 1991),移除 P1 之試驗結果指出 P1 協助 病毒之增殖及細胞間移動但並非必要(Verchot and Carrington, 1995)。P1 的 N 端對 病毒感染能力有重要影響(Moreno et al., 1999),此區域可與數種 potyvirus 蛋白協力 作用,與RNA 非專一結合並增益 HC-Pro 擾亂寄主的防禦機制(Kasschau et al., 2003)。HC-Pro 多以 homodimer 作用,可連結 CP、蚜蟲口針以行媒介傳播,具 proteinase 活性以及負責抑制寄主PTGS (post transcriptional gene silencing)機制,也參與病毒 細胞間移動,對病徵發展極為重要(Boevenik and Oparka, 2005; Kasschau et al., 1997)。HC-Pro 之 N 端 PTK motif 可結合 CP 之 DAG motif,KITC motif 則與蚜蟲 口針結合(Blanc et al., 1997);中央區域 domain A & B、FRNK motif 具 RNA binding 能力(Urcuqui-Inchima et al., 2000);C 端具類 Cys-type proteinase 活性可辨識 HC-Pro 及P3 間切位(Carrington et al., 1992),且可連同 P1 與 P3 抑制寄主 RNA silencing、
促進病毒增殖(Kasschau et al., 2003)。HC-Pro 作為 RNA silencing suppressor,可結 合寄主的small RNA,亦會干擾寄主防禦機制相關因子之運作,抑制 Dicer 對 siRNA 甲基化修飾、干擾RISC 運作,病毒得以增殖累積(Endres et al., 2010 ; Yu et al.,
12
2006),妨礙寄主 miRNA 調控也影響植株型態發展,導致病徵出現(Dunoyer et al., 2004; Mallory et al., 2002; Shiboleth et al., 2007)。報導指出 HC-Pro、CI、P1 皆會與 寄主因子專一性結合,擾亂葉綠體相關機制運作,破壞葉綠體結構、改變生理代 謝平衡而造成病徵發展(Cheng et al., 2008);HC-Pro 亦會與 Ca2+結合蛋白互動,影 響寄主訊息傳遞路徑,可能藉此協助病毒感染或是阻礙寄主防禦反應,且作用區 域會因病毒及寄主種類而異(Shen et al., 2010)。
P3 為帶疏水性區域之膜蛋白、與 6K1、6K2 皆無 RNA binding 活性。研究發 現P3 會在細胞質與 CI 結合(Rodríguez-Cerezo et al., 1993),在核內與 NIa、NIb 結 合(Langenberg and Zhang, 1997),並出現在 VRC(virus replication complex)(Guo et al., 2001)。P3 兩側各有一段疏水性區域,會同 6K1 聚集嵌入於內質網 (endoplasmic reticulum, ER)及高基氏體 (Golgi apparatu),與 VRC 之形成及移動有關
(Eiamtanasate et al., 2007; Cui et al., 2010)。P3 基因區以序列分析軟體發現小分子蛋 白P3N-PIPO 表現,其 ORF 在轉譯過程中途因讀框移位改以+2 閱讀框架轉譯,並 以UGA 為終止碼。此段區域在 Potyviridae 具保守性,突變會影響病毒複製能力、
減小寄主感染範圍(Chung et al., 2008)。P3N-PIPO 引導 CI 抵達原生質絲,共同協 助病毒細胞間移動(Wei et al., 2010)。有報導指出 P3 會影響病原性及病徵嚴重程 度,在 Plum pox virus (PPV)中 P3 及 6K1 可影響寄主範圍(Sáenz et al., 2000),P3 的N 端會與 RubisCO 結合,可能影響光合作用,與病徵發展相關(Lin et al., 2011)。
6K1 具有許多疏水性胺基酸,通常與 P3 結合並共同聚集嵌入 ER,單獨表現 則分散於細胞質及核內(Eiamtanasate et al., 2007)。P3 與 6K1 間的切位由 NIa-Pro 辨認,其水解效力略低,且移除該切位不影響病毒生存能力,但6K1 與 CI 之切位 移除會使病徵消失(Riechmann et al., 1995)。
CI 聚集在感染細胞之原生質絲或 ER 周圍,在細胞質累積成風車狀內含體 (pinwheel inclusions) (Carrington et al., 1998),具 RNA 及 NTP binding 能力,還有 NTPase 和 C 端 RNA helicase 活性區(Laín et al., 1991)。CI 之 NTPase 和 RNA helicase
13
可打開雙股RNA, helicase 活性區可結合 VPg 及寄主轉譯因子 eIF4E
(Tavert-Roudet et al.,2011),與病毒複製、轉譯以及細胞間移動相關。CI 由 P3N-PIPO 引領至原生質絲,與CP 及 VPg 合作引領組裝完成的病毒顆粒自 VRC 移動至原生 質絲、侵入鄰近細胞(Laín et al., 1991; Rojas et al., 1997; Wei et al., 2010)。
6K2 為跨膜蛋白,在中央有疏水性區域(Schaad et al., 1997),與 VPg、NIb、
CI 以及寄主轉譯因子在 VRC 中作用,且干擾寄主細胞的正常胞內運輸機制(Cotton
et al., 2009),與感染寄主能力相關(Moury et al., 2002)。6K2 嵌入 ER 並形成囊泡
(vesicle)至 Golgi apparatus,接著移動到葉綠體外膜並使其內陷形成 VRC,其內集 中病毒生理活動所需分子,提供反應平台(Wei et al., 2010)。NIa 與 NIb 皆會在細胞核內形成內含體(Dougherty and Parks, 1991)。NIa 可分 為N 端 VPg domain 與 C 端 proteinase domain;NIa-Pro、NIa、6K2-VPg-Pro、VPg 的形式皆有非專一性RNA 結合能力。NIa-Pro 具非專一性 DNase 與 Ser-type proteinase 活性(Carrington and Dougherty, 1987),可水解 polyprotein 多數切位,在 異種病毒間具專一性且無法互換(Parks and Dougherty, 1991);VPg 有 NTP binding activity,並與寄主因子 eIF4E 結合起始病毒轉譯(Miyoshi et al., 2008),可與 CI 結 合參與病毒細胞間及長距離移動(Schaad et al., 1997)。VPg 與病毒 RNA 之 5’端共 價鍵結,突變會使病毒完全喪失感染能力(Murphy et al., 1996)。VPg 有 2 段 nuclear
& nucleolar localization signals (NLS),可引導 VPg 進入細胞核、核仁及 Cajal body,
除了利用寄主合成機制,並干擾核仁與Cajal body 調控 small RNAs,擾亂寄主 PTGS 機制(Rajamäki and Valkonen, 2009)。在感染細胞中 VPg 及 NIa 會在細胞核累積,
6K2-VPg-Pro 則不易進入而位於細胞質 VRC (Cotton et al., 2009)。VPg 可做 primer 起始病毒複製,替代mRNA 前端 m7G 結構、在 VRC 內配合 NIb 及寄主轉譯相關 因子協助病毒複製及合成,亦干擾寄主轉譯合成引起病徵(Michon et al., 2006;
Miyoshi et al., 2008);VPg 也與寄主 Cys-rich protein 作用,其 PHD finger domain 可 能輔助病毒感染及移動(Dunoyer et al., 2004)。
14
NIb 具 2 段 NLS,可在核內形成內含體,也出現在細胞質及 VRC 內(Li et al., 1997)。具有 RNA-dependent RNA polymerase(RdRp)特有的 GDD motif,為病毒複 製必需(Klein et al., 1994),且會與寄主轉譯因子 eEF1A、PABP 及 Hsc70-3 結合(Wang
et al., 2000),此外 NIb 也與 NIa-Pro 結合(Li et al., 1997),修飾 VPg 進而成為 primer
起始病毒複製(Puustinen and Mäkinen, 2004)。CP 為外鞘蛋白負責包裹病毒 RNA,參與病毒之蚜蟲傳播、細胞間移動及長距 離移動,同時調控病毒複製(Urcuqui-Inchima et al., 2001),與寄主範圍及病徵表現 密切相關(Moury et al., 2002)。CP 之中央區域可與 RNA 結合,和病毒組裝、細胞 間移動有關;N 端及 C 端暴露於外,與長距離移動有關(Dolja et al.,1995)。CP 中 央區域可與寄主的HSP70 結合,除協助組裝病毒顆粒,HSP70 亦與 HC-Pro 之 C 端共同修飾原生質絲結構(size exclusion limit, SEL)以通過原生質絲(Boevenik and Oparka, 2005)。CP 之 N 端 DAG motif 與 HC-Pro 之 N 端 PTK motif 結合,此處突 變則失去蚜蟲傳播能力(Andrejeva et al., 1999)。另外 CP 也與 NIb 互動調控病毒複 製進行(Hong et al., 1995)。CP 之中央及 C 端序列高度保守,N 端則與寄主及媒介 之專一性互動相關,變異度也較高(Ward and Shukla, 1991),現今分類上多依據 C 端CP (cCP)序列分析病毒間親緣演化關係(Gibbs et al., 2008)。
3’-UTR 序列組成為 AU-rich,其主要功能為維持病毒基因體在細胞內的穩定 及協助病毒複製、轉譯。3’-UTR 近末端有高度保守性片段 UAUGU,可能是 potential polyadenylation signal,該片段在酵母菌及 Nepovirus、Comovirus 的 3’-UTR 也有出 現(Maiss et al., 1989)。
15
參、材料與方法
一、試驗植物準備
使用木瓜來自健康種子繁殖的實生苗,木瓜品種為台農二號品系(Tainung No.2, TN2),種子浸於清水並每日更換,數日後移至潮溼環境催芽,依植株生長情況進 行換盆,最後移入七吋盆,並定期噴藥防治病蟲害與施加氮肥。
白藜種子經1%次氯酸鈉(NaOCl)溶液與 1% Tween 20 表面消毒 30 分鐘,於培 養皿內鋪上濾紙並以無菌水潤濕,再將消毒過的種子放入,待萌芽後移至穴盤中 生長,依植株生長與根系發育情況進行換盆,並定期噴藥防治病蟲害與施肥。
二、木瓜輪點病毒系統來源與保存
本實驗室保存在臺灣田間收集之 PRSV 三系統,包括嚴重嵌紋(SM)系統、畸 形(DF)系統與嚴重壞疽嵌紋(SMN)系統,系統接種原(innocula)來自本實驗室經三代
單斑分離之純系病毒系統的罹病木瓜葉或冷凍乾燥保存的粉末。接種時秤取2 g 的
罹病木瓜葉,以1:3-1:5 (w/v)之比例加入 0.05 M 磷酸緩衝液(K-P buffer: 0.1 M K2HPO4, 0.1 M KH2PO4, pH7.2; 0.1% DIECA),利用接種棒(spatula)及磨砂玻璃板予 以磨碎,作為接種液;或取病葉之冷凍乾燥粉末加入去離子水稀釋,均勻懸浮後 做為接種液。準備健康植株並選擇第三到第五完全展開葉,於接種葉上灑上少許 矽藻土(celite)或金剛砂(carborudum),以消毒過的接種棒沾取病毒接種液,順同方 向塗抹於葉片及葉柄,5 min 後以去離子水清洗葉面,最後以紙巾吸乾多餘水分,
即可置入隔離溫室生長,其間注意肥料管理與病蟲害防治,定期採樣偵測之。為 保有純系病毒可供試驗使用,每兩個月繼代接種病毒到新的木瓜植株。
如欲長期保存病毒樣本,可將罹病木瓜組織分裝並放置於-80˚C 冰箱,或是以 冷凍乾燥法處理:取新鮮病葉組織,加入5 倍重量 0.05 M 磷酸緩衝液,磨碎後在 4˚C 下 4,000g 離心 10 min,取上清液置入保存瓶中,在-80˚C 冰箱放至完全結冰,
16
再以真空冷凍乾燥器抽乾水分後用石蠟膜封口,於-20˚C 冰箱中保存。
三、木瓜輪點病毒之偵測方法 1. 病毒核酸萃取
此法為少量植物總核酸抽取法(total nucleic acid extract method),乃參考先前方 法 (Hung et al., 1999)並作些許修改。取 0.3 g 植物組織以液態氮於研缽中研磨成粉 末,加入1.8 mL 核酸萃取緩衝液(100 mM Tris-HCl, 100mM EDTA-2Na, 250 mM NaCl, pH8.0)混合均勻,再加入 0.3 mL 10% Sarkosyl(N-Lauroyl-sarcosine),倒入 1.5 mL 微量離心管中,於 55˚C 乾浴槽(BL3002; BasicLife Bioscience Inc., Taiwan)作用 1 hr。以微量高速離心機(Centrifuge 5415 R; Eppendorf AG, Hamburg, Germany)室溫 下6,000 g 離心 5 min,取上清液 0.8 mL 置於新離心管,並加入 100 µL 5 M NaCl 及100 µL 10% CTAB (hexadecyl-trimethyl-ammonium-bromide in 0.7 M NaCl),混合 均勻後65˚C 乾浴 10 min,再加入 600 µL CI (Chloroform: Isoamyl alcohol = 24: 1),
充分混合至乳狀。室溫下12,000 g 離心 5 min,取上清液 880 µL 置於新離心管中,
並加入600 µL PCI (Phenol: Chloroform: Isoamyl alcohol = 25: 24: 1),充分混合至乳 狀。室溫下12,000 g 離心 10 min,取上清液 700 µL 加入 420 µL isopropanol 輕輕混 勻。置於-20˚C 待 30 min,於 4˚C 下 12,000 g 離心 10 分鐘,倒掉上清液,管底沉 澱以500 µL 70%酒精(ethyl alcohol)潤洗,再用抽氣乾燥 10 min。最後以 150 µL TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0)充分溶解之,於-20˚C 保存。
2. Two-Step RT-PCR
反應過程分二步驟,先將病毒 RNA 反轉錄(reverse transcription)成互補股 DNA (complementary DNA, cDNA)再進行 PCR(polymerase chain reaction)。反轉錄過程如 下:取7 µL RNA template,加入 2 pmole Reverse primer 及 4 µL 2.5 mM dNTP mix (dATP, dTTP, dCTP and dGTP),65˚C 作用 5 min,立即置於冰上。加入 4 µL 5X 1st
17
strand buffer、2 µL 0.1 M DTT (1,4-dithiothreitol)及 1 µL reverse transcriptase (SuperScriptTM II Reverse Transcriptase, 200 U/µL; InvitrogenTM),在 42˚C 作用 1 hr 即可得到總體積20 µL cDNA 產物。
接續進行PCR 增幅,PCR 反應溶液含 2.5 µL 10X PCR buffer (with 15 mM MgCl2)、2.75 mM MgCl2、0.2 mM each dNTP、0.4 mM each primer (附錄三)、0.25 µL
Taq DNA polymerase (Super-Therm DNA polymerase, 1 U/µL; BERTEC)及 1 µL
cDNA template,加 ddH2O 調整反應總體積至 25 µL。將混合好之反應溶液放置 PCR 熱循環反應器內(GeneAmp 2720; Applied Biosystem Corp., Norwalk, CT, USA),PCR 溫度循環條件為:94˚C/3 min,[94˚C/1 min、annealing temp./1 min、72˚C/2 min]共 30 個循環,72˚C/10 min。PCR 增幅產物以 1.4% agarose gel 進行電泳膠體分析(Hunget al., 2000)。
3. One-Step RT-PCR
此偵測方法係將前述二個步驟合併進行。RT-PCR 反應溶液含 2.5 µL 10X PCR buffer (with 15 mM MgCl2)、2.5µL 5X 1st strand buffer (with 15 mM MgCl2)、5 mM DTT、2.5 mM MgCl2、0.2 mM each dNTP、0.4 mM each primer(附錄三)、0.25 µL Taq DNA polymerase、0.25 µL reverse transcriptase 及 2.5 µL template,以 ddH2O 調整反 應總體積至25 µL。RT-PCR 溫度循環條件為:42˚C/50 min 或 50˚C/35 min,94˚C/2 min,[94˚C/30 sec、annealing temp./30 sec、68˚C/45 sec]共 10 個循環,[94˚C/30 sec、
annealing temp./30 sec、68˚C/45 sec*,*:每循環增加 5 sec]共 25 個循環,68˚C/10 min。增幅產物以 1.4% agarose gel 進行電泳膠體分析(Hung et al., 2000)。
4. PCR 產物電泳膠體分析
PCR 反應結束後,取 10 µL PCR 產物和 2 µL 6X loading dye 混合均勻,在 1.4%
agarose gel 中進行電泳分析。依據 DNA marker (BIOLADDER® 100 bp DNA Ladder;
18
BasicLife Bioscience Inc.)判別產物分子量。電泳緩衝液為 0.5X TAE buffer(20 mM Tris-acetate, 0.5 mM EDTA, pH 8.0),以 100V 進行電泳。完成後膠體以 0.5 µg/mL EtBr (ethidium bromide)染色,使用數位化影像處理與分析系統(AlphaImager® AlphaEaseFC Software; Alpha Innotech Co., USA)觀察膠體內分離之 DNA 條帶。
四、 木瓜輪點病毒 PRSV-SMN、PRSV-SM 之感染性選殖株構築 1. PRSV 核酸序列通用性引子對設計
參照實驗室前人所發表 PRSV-SM 及 PRSV-SMN 的全長核酸序列(李,2006;
廖,2004),以引子設計軟體 PrimerSelectTM (DNASTAR, Inc., USA)與 Oligo 7 (Molecular Biology Insights, Inc., USA)分析兩系統序列,選取具特異性之序列合成 引子,並在部分引子5’端更動序列以導入限制酶切位(表一)。正向引子 T3F 在前端 外加T3 promoter 及一個 guanine,3’ UTR 所用 reverse primer 為 25 nt 之 oligo(dT) 接續限制酶 XbaI 切位序列,使 PCR 產物帶有 poly(A) tail(劉,2008)。
2. Conventional RT-PCR
先將萃取完成之病毒核酸做反轉錄以合成cDNA,目標片段之增幅選用具校正
(proofreading)功能之聚合酶。使用 Advantage® 2 PCR kit (Clontech Laboratories, Inc.) 進行PCR,反應溶液含 5 µL 10X Advantage 2 PCR Buffer、0.25 mM each dNTP、0.4 mM each primer、1 µL 50X Advantage® 2 Polymerase Mix 以及 5 µL cDNA
template,添加 ddH2O 調整總體積至 50 µL。PCR 溫度循環條件為:94˚C/1 min,
[95˚C/30 sec、63˚C/1 min、72˚C/1 min per kb]共 25 個循環,加入 0.5 U Taq DNA polymerase 再以 72˚C 作用 10 min。PhusionTM High-Fidelity DNA Polymerase (Finnzymes Oy)之反應溶液配置為 10 µL 5X Phusion HF Buffer (with 7.5 mM
MgCl2)、2.5 mM MgCl2、0.25 mM each dNTP、0.5 mM each primer、0.5 µL Phusion DNA polymerase(2 U/µL)與 3µL cDNA template,以 ddH2O 調整總體積至 50 µL。
19
PCR 溫度循環條件為:98˚C/30 sec,[98˚C/5 sec、63˚C/12 sec、72˚C/40 sec per kb]
共25 個循環,72˚C/10 min。反應完成之 PCR 產物使用 PCR Clean-up Kit (GeneMark) 純化,最後以45 µL elution solution 溶離核酸而收集之。純化完成的 PCR 產物加入 5 µL 10X PCR buffer、0.2 mM dATP 及 1 U Taq DNA polymerase,在 72˚C 作用 20 min。膠體純化 PCR 產物時使用 0.8-1% agarose gel 進行電泳,以 DNA marker (AccuRuler 1 kb DNA RTU Ladder; Maestrogen, Inc.)判別產物之分子量,再用乾淨刀 片切下目標片段,利用QIAquick® Gel Extraction Kit (QIAGEN Group)予以純化。
3. PCR 產物之選殖
PCR 產物利用 pGEM®-T Vector System I (Promega)進行選殖,步驟為將 3 µL 純化產物、5 µL 2X Rapid Ligation Buffer、1 µL pGEM®-T Vector (50 ng/µL)與 1 µL T4 DNA Ligase (3 U/µL)混合,在 16˚C 進行接合反應(ligation)過夜。隨後進行轉型 作用(transformation),將接合產物送入 Escherichia coli strain JM109。反應產物加入 大腸桿菌勝任細胞(ECOSTM 9-5 E. coli competent cell; Yeastern Biotech Co., Ltd.)中 混合,冰浴10 min 後在水浴槽(WB201; Kansin Instruments Co., Ltd., Taiwan)以 42˚
C 熱休克(heat shock) 45 sec,再將菌液置於冰上 5 min。經藍白篩(blue-white screen) 挑選成功轉入載體之菌株,先在1.5% LB 固態培養基(LB plate,含 50µg/mL
ampicillin)上加入 40 µL X-gal (40 mg/mL in DMF)及 40 µL 100mM IPTG 均勻塗佈 後,經heat-shock 處理之菌液塗佈於 LB plate/Amp 中,在培養箱(TU-400; YIH DER Instruments Co., Ltd., Taiwan)以 37˚C 培養 13-16 hr。隨機挑選白色菌落分別接種於 LB plate/Amp 及 1 mL 液態培養基(LB broth,含 50 µg/mL ampicillin),37˚C 培養 12-16 hr (LB broth/Amp,200 rpm 震盪),保存於 4˚C。
培養好的1 mL LB culture 利用 Gene-SpinTM Plasmid MiniPrep Kit (Protech)純化 微量質體(plasmid),按流程操作並以 50µL elution solution 溶離質體。獲得之微量 質體以限制酶酵解確認目標片段之存在,即可大量培養進行後續實驗,可使用
20
Gene-SpinTM Plasmid MiniPrep Kit 抽取 4 mL LB culture,溶離於 100 µL elution solution;或培養 30-50mL LB culture,以 AxyPrep Plasmid Midiprep Kit (AXYGEN) 抽取質體,最後以300 µL elution solution 溶離。
4. PRSV 全長序列選殖株之合成
以MapDraw TM (DNASTAR, Inc., USA)程式分析 pGEM®-T Vector 及病毒序 列,選取適當限制酶切位。利用限制酶(Restriction Endonucaleases; New England Biolabs®, Inc.)進行酵解,反應溶液含純化質體、1X NEBuffer、1X BSA (Bovine Serum Albumin),以 ddH2O 調整體積,加入限制酶並設定適當反應溫度作用 1 hr;
進行部分酵解時1 µg 質體加入 0.1 U 限制酶,在反應溫度作用 75-90 min。經膠體 純化之目標片段用T4 DNA Ligase (6 U/µL; New England Biolabs®, Inc.)行接合反 應,將欲接合之片段、1 µL 10X Ligation Buffer 與 1 µL T4 DNA Ligase 配置成 10 µL 反應液,16˚C 過夜後送入 E. coli 勝任細胞。
五、胞外轉錄(in vitro transcription)
將帶有全長序列之選殖株質體以 XbaI 酵解 2-3 hr,得到之線狀質體以 PCI 處 理去除蛋白質,加入0.1 倍體積 3M NaOAc (sodium acetate)、3 倍體積 99.5%酒精 與1 µL glycogen (20µg/µL; Roche)混合均勻,置於-20˚C 過夜。隔日 4˚C 下 12,000 g 離心 15 min,移去上清液再加入 500 µL 70%酒精,4˚C 下 12,000 g 離心 15 min,
倒掉上清液並抽氣乾燥10 min,核酸沈澱物溶於 6 µL DEPC-H2O。使用 T3 mMESSAGE mMACHINE® Kit (Ambion®)進行 in vitro transcription 以合成 capped RNA transcripts,步驟為取 6 µL 產物加入 2 µL 10X Reaction Buffer、10 µL 2X NTP/CAP mix、2 µL enzyme mix,置於 37˚C 反應 30 min。加入 1 µL 30mM GTP solution 於 37˚C 反應 30 min,再加 1 µL 30mM GTP solution 置於 37˚C 反應 2 hr。
取1 µL 產物做電泳分析確認後進行接種。若欲定量需去除 template DNA,將 RNA
21
產物加入1 µL TURBO DNase,在 37˚C 反應 15 min 後以 PCI 處理,再以 0.6 倍體 積isopropanol 或 2 倍體積 99.5%酒精沉澱,以 TE buffer 或 DEPC-H2O 溶解產物。
六、接種試驗
胞外轉錄所得之 RNA transcripts 進行機械接種到寄主植物木瓜及白藜,每片 葉片接種至2-3 µg RNA:選取約 3-4 片展開葉之木瓜幼苗,接種第一、第二片展 開葉(Chiang and Yeh, 1997);白藜在生長至約 7-8 片展開葉時接種。使用經漂白水 清洗及高壓滅菌消毒之玻璃棒與金剛砂進行機械接種,接種後置於溫室及植物恆 溫生長箱(26˚C)(GC-101; FIRSTEK, Taiwan)隔離觀察,4 週後取接種葉之上位葉進 行核酸純化,並以RT-PCR 檢測,所使用引子對有通用型引子對 PRSV 476,與不 同系統之專一型引子對SMN-466、SM-517、DF-598,分別進行增幅及電泳分析。
七、選殖株之基因序列分析
選殖株的病毒全長序列皆委外定序(Tri-I Biotech, Taiwan),並經 NCBI 的 BLAST®(Basic Local Alignment Search Tool)程式做比對分析,參考前人發表序列得 到選殖株所嵌入之病毒全長基因體cDNA 序列,並使用 Clustal X (Larkin, et al., 2007)進行多重序列比對(multiple sequence alignment)分析,所得結果輸入
PAUP*(Sinauer Associates, Inc.,USA)作親緣關係演算,套用 neighbor-joining 法及 Hasegawa-Kishino-Yano (HKY) model、並以 Potato virus Y (GenBank accession No.
AB270705)為外群,產生 PRSV 分離株之親源樹狀關係圖。病毒各基因區域之序列 相關比較分析使用分析軟體MegAlignTM (DNASTAR, Inc., USA)以 J. Hein method 進行比對,SimPlot 則以 Kimura’s two parameter model 演算(Lole, et al., 1999)。
22
肆、結果
一、木瓜輪點病毒基因體核酸片段增幅與選殖
接種於台農2 號(TN2)木瓜保存之木瓜輪點病毒(PRSV) SMN、SM、DF 三系
統純系病毒株,分別抽取其核酸並合成cDNA 以作為後續構築選殖株之用。病毒
cDNA 以不同引子對(表一),進行各片段之 PCR 增幅,並將 PCR 產物選殖於載體 pGEM-T Vector(圖一)。
SMN 系統首次以 Advantage 2 polymerase 進行 PCR 增幅,獲得的病毒核酸片
段依序為:引子對T3F/R1968 增幅出的 T3F-HC-NA (2.0 kb)片段、引子對 F1748/R8546 增幅出的 HC-Nb-NA (8.6 kb)及引子對 F6727/PolyA-XbaI 增幅出的 Na-AX-NA (3.6 kb),分別以電泳確認其片段長度(圖二 A),並以 T-A cloning 套組 構築出prT3F-HC-NA、prHC-Nb-NA 和 prNa-AX-NA 三選殖株。因定序發現序列 準確度不足,於是再次增幅病毒核酸片段,以Phusion DNA polymerase 配合通用 性引子對T3F/R1968 增幅出 T3F-HC-NP (2.0 kb)、引子對 NcoI+F1577/R7359 增幅 出NP1-Na-NP (5.8 kb)、引子對 F6518/R9629 合成 Na-CP-NP (3.1 kb)與引子對 F8353/PolyA-XbaI 合成 Nb-AX-NP (2.0 kb),分別以電泳確認其片段長度(圖二 B)。
利用T-A cloning 方式選殖到 prT3F-HC-NP、prNa-CP-NP 以及 prNb-AX-NP 三選殖 株,prNP1-Na-NP 運用引子 NcoI+F1577 及 PCR 產物 3’端所含限制酶切位 NcoI、
BspEI 進行 directional cloning。
SM 及 DF 系統皆以 Phusion DNA polymerase 行增幅,而 prNP1-Na-DP 之構築 改用 NheI 切位進行 directional cloning。SM 系統所得質體為 prT3F-HC-MP、
prNP1-Na-MP、prNa-CP-MP 還有 prNb-AX-MP。DF 系統在選殖過程中,帶有 NIa、
NIb 部分序列之片段無法順利篩選出與其他質體同方向的 cDNA 選殖株,因此改用 pfNa-CP-DP,其餘選殖株則是 prT3F-HC-DP、prNP1-Na-DP 以及 pfNb-AX-DP。
23
二、全長選殖株之合成及胞外轉錄試驗
為構築 PRSV 全長基因體 cDNA 選殖株,利用病毒序列原有的限制酶切位搭 配pGEM-T Vector 做選殖,並使全長 PRSV 基因體序列前端帶有 T3 promoter 及一 個guanine,尾端連接約 25nt 之 poly(A)及 XbaI 切位(圖一 A)。PRSV-SMN 系統構 築出全長序列選殖株共有2 株:prT3-AX-NA (= pN-A)及 prT3-AX-NP (= pN-P);
PRSV-SM 系統構築成 prT3-AX-MP (= pM-P);PRSV-DF 系統為 pfT3-AX-DP (=
pD-P)。
三種系統之全長基因體cDNA 選殖株以限制酶 XbaI 酵解後得到約 10.3 kb 病 毒全長序列外加3.0 kb 載體 pGEM-T Vector,合計約 13.3 kb 之線狀 DNA 分子,以 1% agarose gel 進行膠體分離所獲分析結果,其分子量對照 DNA marker 及既有之 DF 系統全長選殖株 pT3-NP( = pD-T) (劉,2008)與預期相符(圖三)。將經酵解之全 長cDNA 選殖株以 T3 RNA polymerase 做 in vitro transcription 並加入 DNase 處理,
經PCI 純化測得 5 µg 質體轉錄後可得約 60-70 µg RNA 產物,RNA transcripts 進行 電泳分析,可觀察到預期分子量之RNA 分子(圖四)。
三、胞外轉錄感染力試驗及不同系統選殖株在寄主之病徵表現
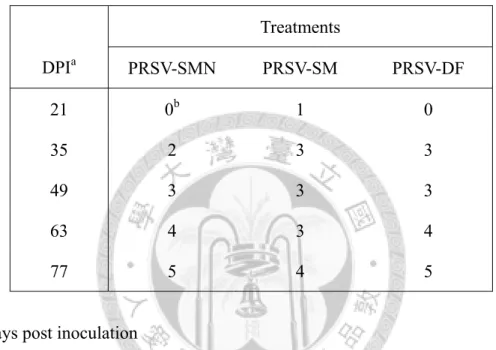
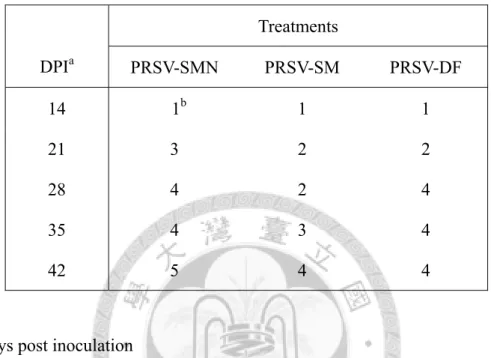
取生長至 3-4 片展開葉之 TN2 木瓜幼苗,每片展開葉接種 3 µg RNA
transcripts、每株木瓜苗接種 2 片葉子。接種後第三週所有系統之上位葉開始有輕 微葉脈透化(vein clearing)現象且葉片輕微黃化。SMN 系統初期造成葉脈透化及葉 面輕微嵌紋(mild mottling),續而新生葉葉脈褪色、葉緣向上捲曲(curling);中期新 葉葉緣向上捲曲、葉片變厚、新葉面積縮小且植株矮化(stunting);後期下位葉脫落,
僅存頂端絲狀葉(fern leaf),其葉肉縮減只餘葉脈,且植株嚴重矮化、根系亦發育 不良。SM 系統起初使木瓜葉脈透化及嵌紋(mottling);中期葉片嵌紋、黃化且葉面 積縮小,主脈周圍有綠帶;後期造成葉片嚴重嵌紋(severe mottling)、葉片縮小且葉 面散布綠島(green island)但葉片型態無顯著改變,植株有矮化現象。DF 系統前期
24
導致葉脈透化及葉面嵌紋斑駁、且新葉之葉面皺折不平,其後新生葉脈白化、葉 緣向上捲曲;中期葉片變厚、扭曲畸形(deformation),葉肉組織萎縮而植株矮化;
中後期畸形葉出現綠島塊斑,新生葉呈絲狀(fern leaf, shoe stringing);後期頂梢簇 生(rosette)絲狀葉、綠島逐漸褪去,植株與根系發育明顯受阻(圖五~八)。
不同系統選殖株之RNA transcripts 分批接種至 TN2 健康植株,其感染發病率
依接種時期之氣溫變化有些許差異,溫度較低時三個病毒系統發病率皆為100%,
若氣溫升高則DF 系統之感染發病率有下降情形,於恆溫生長箱內(26˚C)三系統在 木瓜寄主上感染發病率亦有下降趨勢(表二)。將上述發病進展之病徵嚴重度,依病 徵級數分作6 級(0 級~5 級),自發病後隔週進行記錄(表三)。
接種 PRSV 三系統之 TN2 在第二週時均開始出現輕微葉脈透化情形。SMN 系 統前期使系統葉葉脈透化、黃化、下位葉脫落,頃後只餘頂梢新生葉且植株嚴重 矮化,新生葉萎縮、後期轉為絲狀;SM 系統前期造成葉脈透化、葉面嵌紋,後期 葉片明顯嵌紋、縮小、植株矮化;DF 系統初期引起葉脈透化、葉面輕微嵌紋,中 期葉片黃化、縮小,中後期葉面畸形並出現綠島,後期新生葉呈絲狀且植株矮化(圖 九)。將上述發病進展之病徵嚴重度,自發病後每週進行記錄病徵級數(表五)。
三系統感染性選殖株接種之發病木瓜植株,將病葉汁液再以機械方式感染株 高約20 cm 之健康 TN2 植株。三週後 SM 系統感染者即出現輕微嵌紋病徵,隨後
嵌紋程度加劇,中期葉片嚴重嵌紋、植株矮化;SMN 系統接種後第四週出現病徵,
發病中期下位葉脫落,後期下位葉皆脫落萎凋、只餘新生葉;DF 第四週時葉面輕 微嵌紋、而後出現葉面皺折不平情形,中期葉片嚴重嵌紋,中後期造成新生葉畸 形且末端呈絲狀,後期畸形葉可見綠島。依據發病進展做病徵嚴重度之分級記錄,
自發病後隔週進行記錄(表四)。將三系統繼代接種至 30 cm 高之 TN2 健株,數週 後SM 系統發生葉片嚴重嵌紋;DF 系統葉片嚴重嵌紋伴隨畸形,且出現綠島;SMN 系統葉片之嚴重嵌紋病斑出現壞疽、下位葉黃化脫落 (圖十)。
將三系統選殖株之RNA transcripts 接種至單斑寄主白藜上,病徵表現較原病
25
毒的自然分離株為差,SMN 與 SM 系統經過 14 天後僅在接種藜葉上觀察到一個褪
色黃萎病斑、接種DF 系統之葉片未出現局部病斑。自發病木瓜繼代接種至白藜,
SMN 與 SM 系統在白藜葉上造成少量局部病斑,病斑邊緣不明顯,DF 系統在白藜 上產生病斑能力較低,且病斑與健康部位之間的邊界模糊 (圖十一)。
四、偵測結果及新專一性引子對設計
接種第四週,待三種系統皆於寄主木瓜上產生明顯病徵後,摘取少量系統葉 以總核酸抽取法萃取植物體之核酸,並以通用型引子對PRSV-476,與各系統專一 型引子對SMN 466、SM 517、DF 598 做分子鑑定(圖十二)。使用引子對 PRSV-476 做RT-PCR 檢測時,電泳分析可發現有特異性 DNA 條帶增幅(476 bp);以 SMN 與 SM 兩系統專一性引子對 SMN-466、SM-517 對三系統做 RT-PCR 增幅,SMN 與 SM 兩系統均有系統專一性的條帶出現,證實其 RNA transcripts 成功感染並可在寄 主體內增殖移動;然而引子對DF-598 對三系統之純系樣本進行 RT-PCR 檢測時,
除DF 系統有專一性增幅外,SM 系統也有非專一性增幅。因此本論文重新設計出
DF 系統專一性引子對 DF-626 進行系統間專一性增幅之測定,其對 DF 系統之敏感 性及專一性較引子對DF-598 為佳(圖十三)。
五、 三系統全長選殖株與已發表 PRSV 分離株之序列比對分析
將三系統感染性選殖株進行定序,所得全長序列再與前人已發表序列比較(附 錄四),可發現同系統間在核酸及胺基酸序列皆維持高度一致性,僅有 DF 系統與 選殖株pfT3-AX-DP、pT3-NP 在核酸位置 8182-8189 發現位移;SMN 系統之選殖 株prT3-AX-NP、prT3-AX-NA 與先前本研究室發表的序列相比則是在部分區域出 現核酸序列單點替換,多數集中於6K2、VPg 和 3’-UTR 基因區,並在病毒蛋白 P1、6K2、NIa 及 CP 之胺基酸序列導致非同義突變(nonsynonymous mutation)。
三系統選殖株的基因區域序列做系統間個別比對(表六),與原發表的病毒株序
26
列相似度頗一致(李,2006;廖,2004),在基因體前段之序列保守性較低,5’-UTR 及P1 核酸序列相似度為 93-94%;在 NIa 以下的後段基因體核酸序列相似度則為 96-99%。整體而言,DF 系統與 SM 系統間相似度最高,全長核酸序列相似度為 97.3%,除了 5’-UTR、P1 及 P3 之外的核酸序列相似度皆有 97%以上,胺基酸序 列更在98-100%;SMN 系統與另外兩系統之全長核苷酸相似度皆為 96.5%,和 DF 系統之polyprotein 胺基酸相似度為 97.2%、與 SM 系統之胺基酸相似度為 97.0%。
各病毒基因之胺基酸的序列相似度在系統間大都在97-100%,僅在 P1 相似度 低於93%,P3 序列相似度略低,但多在 97%上下。6K1 之序列相似度在三系統間 皆為100%,6K2 因 SMN 系統在該區域有核酸序列變動,與另外兩系統之序列相 似度為96.6%,DF 與 SM 兩系統間仍為 100%。NIa 蛋白之 VPg 在 SMN 系統中亦 有胺基酸序列更動,SMN 及 SM 系統間序列相似度為 97.4%。
將全長感染性選殖株之核酸序列與國內外發表之PRSV 分離株進行比對,以
neighbour-joining 法套用 HKY model 分析病毒全長基因體、並以 Potato virus Y (GenBank accession No. AB270705)為外群繪出親源樹狀關係圖(phylogenetic tree)(圖十四),可看出 PRSV 約略依不同地域形成兩大關係群,美洲群(American group)包括來自夏威夷(S46722; EU126128)、巴西(DQ374152; DQ374153)、墨西哥 (AY231130)及印度(EF017707; EU475877)的 PRSV-P 與 PRSV-W,亞洲群(Asian group)病毒株則涵括泰國(AY010722; AY162218)、韓國(AB369277)、中國(EF183499;
HQ424465)及臺灣(X97251; EU882728; AY027810; DQ340769; DQ340770;
DQ340771) (Bateson et al., 2002; Mangrauthia et al., 2008; Noa-Carrazana et al., 2007)。本次研究所構築的 5 組 PRSV 全長感染性選殖株如預期劃分在各病毒系統 下,且與臺灣的永康分離株(P-YK)、P-5-19 及嘉義分離株(W-CI)較為近似,另外韓 國發表之分離株(W-KR)也與我們的 SM 系統親緣關係相當接近。
以DF 系統之全長核苷酸序列為主與 SMN、SM 系統做比對,並將其餘 PRSV 分離株依親源樹之關係群歸為美洲或亞洲群,以Kimura’s two parameter model 做
27
全長序列相似度繪圖(similarity plot)(圖十五)。其中可看出美洲群與 DF 系統之序列 歧異度最高,且在P1、P3 之 C 端、VPg 之 C 端、NIa-Pro 之 C 端、NIb 之 N 端及 CP 之 N 端序列變異度較大;亞洲群在大部分區域與 SMN、SM 系統差異不大,僅 在CI 之 N 端、6K1 及 CP 之 C 端的序列相似度較兩系統略低。在三系統間以 SM 與DF 系統的胺基酸序列較為接近,但兩者在 P1 之中央區域、P3 中央到 C 端區域 變異度較大,CI 基因以下序列則相似度極高;SMN 與 DF 兩系統之核苷酸序列只 有在P1 之 N 端及 CI 基因區,與 SM 系統有較高相對歧異度,而胺基酸序列於 P1、
HC-Pro 之 N 端、P3 中央區域、VPg 之 C 端、NIa-Pro、CP 之 N 端則與 DF 系統相 似度較低。此外SMN 及 SM 兩系統在 P1、HC-Pro 之 N 端、P3 中央到 C 端區域、
VPg 之 C 端、NIa-Pro、CP 之 N 端區域胺基酸序列變異度較高。