行政院國家科學委員會專題研究計畫 成果報告
聚乙烯亞胺接枝對其載體特性及轉染效率的影響(2/2) 研究成果報告(完整版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 95-2221-E-011-079-
執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣科技大學化學工程系
計 畫 主 持 人 : 曾文祺
計畫參與人員: 博士班研究生-兼任助理:蘇玲鈺
碩士班研究生-兼任助理:張家健、莊智元
報 告 附 件 : 出席國際會議研究心得報告及發表論文
處 理 方 式 : 本計畫涉及專利或其他智慧財產權,2 年後可公開查詢
中 華 民 國 96 年 11 月 06 日
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※ 聚乙烯亞胺接枝對其載體特性及轉染效率的影響(2/2) ※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:5個別型計畫 □整合型計畫 計畫編號:NSC 95-2221-E-011-079-
執行期間: 2006 年 08 月 01 日 至 2007 年 07 月 31 日
計畫主持人:曾文祺 共同主持人:
計畫參與人員:蘇玲鈺、張家健、莊智元
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
5出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份 執行單位:國立臺灣科技大學化學工程系
中 華 民 國 96 年 11 月 05 日
Abstract
Branched polyethylenimine (PEI) is a cationic polymer capable of forming self-assembly complexes with DNA to become a highly efficient agent used in gene delivery. Conjugation through the primary amines of PEI is a most commonly used approach further to enable the targeting delivery or to improve the stability of the DNA-polymer complexes. An understanding of how the conjugation affects the transfection mechanisms can help design efficient polycationic vectors. In order to investigate the effects of conjugation, folate and the dextrans of molecular weight 1500 (dex-1500) and 10000 (dex-10000) were used to prepare three different types of PEI conjugates: dextran-PEI, folate-PEI, and folate-dextran-PEI, which were subsequently employed to form complexes with DNA. These conjugates were found to cause less cytotoxicity than the unmodified PEI as revealed by the MTT method, and to be able to deliver an approximate amount of ethidium monoazide labeled plasmid into the cells. The efficiencies of green fluorescent protein (GFP) expression mediated by these conjugates, however, were less efficient than those mediated by the unmodified PEI. A titration experiment suggested that conjugation through the primary amines of PEI resulted in the loss of buffering capacity, a major factor aiding the release of plasmid from the endosomes. When a quantitative relationship between the buffering capacity and transfection efficiency was examined, the minimal detection of GFP positive cells was noted to require a threshold of buffering capacity, around 50% of the unmodified PEI. In addition, the cytotoxicity could be also related to the buffering capacity in an approximately linear trend. It is thus concluded that the severe loss of buffering capacity by conjugation might be attributed to the inefficiency transgene expression mediated by the dextran-PEI conjugates.
Table of Contents
Introduction...5
Experimental Section ...6
Materials. ...6
Plasmid amplification and labeling...6
Preparation of folate-dextran-PEI conjugate. ...6
Preparation of folate-PEI conjugate...7
Preparation of dextran-PEI conjugate...7
Determination of the amount of primary amine on each PEI conjugate...7
Determination of the amount of folate on the PEI conjugate. ...7
Titration of the PEI conjugates. ...7
Measurements of the size and zeta potential of the DNA-polymer complexes...8
Cell culture...8
Cytotoxicity assay by MTT. ...8
Flow cytometric analysis. ...8
Results and Discussion ...9
Preparation of PEI conjugates...9
Transfection mediated by DNA-polymer complexes ...9
Physicochemical properties of the DNA-polymer complexes in the salt free and DMEM buffers ...10
Entry of plasmid delivered by the DNA-polymer complexes ...10
Measurements of buffering capacity...11
Measurements of cellular cytotoxicity...11
Effects of intracellular buffering capacity on cytotoxicity and transfection efficiency...12
Conclusions...12
References ...13
List of Figures and Tables
TABLE 1 THE ESTIMATED DEGREES OF CONJUGATION ON EACH PEI CONJUGATE 16
FIGURE 1 TRANSFECTION EFFICIENCIES OF THE PEI CONJUGATES AT DIFFERENT N/P RATIOS.
17 FIGURE 2 SIZES AND ZETA POTENTIALS OF THE DNA-POLYMER COMPLEXES IN THE DMEM
AND SALT-FREE BUFFERS. 17
FIGURE 3 MEAN FLUORESCENCE INTENSITIES OF THE CELLS AFTER INCUBATION WITH THE DNA-POLYMER COMPLEXES CONTAINING EMA-LABELED PLASMID 20 FIGURE 4 BUFFERING CAPACITIES OF THE PEI CONJUGATES 21 FIGURE 5 CELLULAR CYTOTOXICITIES OF THE DNA-POLYMER COMPLEXES 22 FIGURE 6 DEPENDENCE OF CYTOTOXICITY AND THE BUFFERING CAPACITY 23 FIGURE 7 DEPENDENCE OF TRANSFECTION EFFICIENCY AND BUFFERING CAPACITY 24
Introduction
Cationic polymer is a major type of synthetic vectors used in gene delivery1, 2. The positive charges of the polymers generally come from the amine groups, which play an important role in determining the vector efficacy. The amine groups participate in the formation of DNA-polymer complexes, the cellular entry of the complexes, and the intracellular processing of the complexes. In addition, the amine groups allow a way to chemically modify the vectors by conjugating other molecules onto the polymers.
Among the cationic polymers, such as polyethylenimine (PEI)3, 4, dendrimer5, polylysine6, chitosan7, PEI has been shown to be a most efficient agent for transfection. Branched PEI is consisted of 25% primary, 50% secondary, and 25% tertiary amines on average. The high density of amine groups enables PEI to form tighter and smaller self-assembly complexes with DNA through charge interactions8. The positive surface charges of the complexes allow the attachment of the complexes onto the negatively-charged proteoglycans of cell membranes9, and the subsequent entry into the cytosol via endocytosis. Inside the cytosol, the amine groups can further act as proton-sponge to induce the influx of chloride ions into the endosomes3, 10, thereby facilitating the release of DNA from the endosomes to get into the cytosol11-13. Once the released naked DNA or DNA-polymer complexes enter the nucleus, transcription begins.
Modifying the amine structures is usually employed to further promote the transfection efficiency mediated by DNA-polymer complexes or to enhance the stability of DNA-polymer complexes14, 15. A commonly used approach is to attach a pendant group onto the primary amines of the cationic polymers. Polylysine conjugated with polyhistidine on its -amine groups has been shown to be more efficient than the unmodified polylysine16, 17. The conjugated polylysine enhances the efficiency of transgene expression because the imidazole groups of polyhistidine can provide higher buffering capacity to more efficiently facilitate the endosomal release of DNA. In order to stabilize the DNA-polymer complexes, cationic polymers can be conjugated with the biocompatible polymers of no charge, such as polyethylene glycol15, 18-20 and dextran21, 22, to decrease the charge effects of the salts and proteins present in the extracellular environments. These stabilized vectors, however, generally less efficiently deliver plasmid into the cells for transgene expression because such modifications decrease the interactions of the vectors and the extracellular environments as well as the interactions with the cells. Targeted delivery is then used to restore the transgene expression by conjugating ligands onto either the primary amines of the cationic polymers or the distal ends of the previously conjugated polymers23-25, such as the use of RGD peptide26, mannose27, transferrin20, and folate18, 28, 29 for targeting endothelial cells, dendritic cells, hepatocytes, and folate-overexpression carcinoma, respectively.
Conjugation through primary amines is such a most commonly used method to modify polycationic vectors for gene delivery. Nevertheless, how the conjugation affects the cellular mechanisms of transgene expression in a quantitative way has so far received little attention.
In this study, we used folate and the dextrans of molecular weight 1500 and 10000 to modify PEI, and prepared three different types of PEI conjugates: dextran-PEI, folate-PEI, and folate-dextran-PEI, each with various degrees of conjugation. These conjugates were used to form DNA-polymer complexes for transfection. Cellular mechanisms, from the cellular entry to the endosomal release, were investigated by the correlations among the physicochemical properties of the DNA-polymer complexes, the buffering capacity of each conjugate, the cellular cytotoxicity, the amount of intracellular plasmid, and the efficiency of transgene
expression. The physicochemical properties were indicated by the sizes and the zeta potentials of the complexes. The buffering capacity was determined by titration, and the cellular cytotoxicity was examined using the MTT method. The efficiency of transgene expression and the amount of intracellular plasmid were monitored using green fluorescent protein (GFP) and ethidium monoazide (EMA) labeled plasmid, respectively. Flow cytometry was used to measure the number of cells expressing GFP and the fluorescence intensity of EMA-labeled plasmid.
Experimental Section
Materials.
Polyethylenimine (PEI, MW 25000), sodium cyanoborohydride, 2-chloroethyl amine, dimethyl sulfoxide (DMSO), sodium periodate were obtained from Aldrich (Milwaukee, WI).
Dextran of molecular weight 10000, denoted by dex-10000 and dextran of molecular weight 1500, denoted by dex-1500 were from Fluka Chemie (Buchs, Switzerland). Folate, thiazolyl blue tetrazolium bromide (MTT), Dubecco’s modified Eagle medium (DMEM), and folate-deficient Dubecco’s modified Eagle medium were purchased from Sigma (St. Louis, MO). Dicyclohexyl carbodiimide (DDC), 2,4,6-trinitrobenzene sulfonic acid (TNBS), and 1-ethyl-3-(3-dimethylaminopropy) carbodiimide hydrochloride (EDC) were from Pierce (Rockford, IL). Ethidium monoazide (EMA) was purchased from Molecular Probes (Eugen, OR). All chemicals were used as received. Water was deionized by a Milli-Q water purification system (Bedford, MA). The pEGFP-C1 vector containing a mutated green fluorescent protein gene driven by a Cytomegalovirus promoter was from Clontech (Palo Alto, CA). Dialysis membranes with different molecular weight cutoffs were from Spectrum (Los Angels, CA). Fetal bovine serum and antibiotics of penicillin-streptomycin-amphotericin were from Hyclone (Logan, Utah).
Plasmid amplification and labeling.
The pEGFP-C1 vector was amplified in E. coli and purified as previously described. The purified plasmid was covalently labeled with EMA, which could provide a stable fluorescent reporter of the bound DNA. The labeling of plasmid with EMA by photoactivation was carried out as previously described30. Briefly, the mixture of EMA and plasmid was exposed to white light for 30 min. The unreacted EMA was removed by extraction with saturated n-butanol and dialysis against 2-l of distilled water six times using Spectra/Por membrane (MWCO = 15000) in the dark.
Preparation of folate-dextran-PEI conjugate.
Amination of dextran. Dextran (1.0 g) was dissolved in a solution of 2.2 g NaOH in 10 ml water. The mixture was incubated at 30 oC for 30 min. A weight of 2.6 g 2-chloroethylamine was added to the above mixture. The reaction was performed at 30 oC with constant shaking.
The extent of reaction was controlled by different reaction times: 12 and 18 h for dex-1500, and 12 and 24 h for dex-10000. After reaction, the reacted dextran was purified by repeated precipitation with 2-fold volume of methanol till the color became whitish before lyophilization.
Oxidization of amino-dextran. The vicinal hydroxyl groups of the above amino-dextran (0.6 g dex-1500 or 2.0 g dex-10000 in 20 ml water) were oxidized to a aldehyde group in the presence of sodium periodate (0.4 g)31. The reaction was carried out at 25 oC with constant shaking for 8 h in the dark. The oxidized amino-dextran was lyophilized after repeated
precipitation with methanol as described above.
Conjugation of folate to the oxidized amino-dextran. Folate (0.083 g) was treated with an equal molar amount of EDC in 12 ml DMSO at 25 oC for an hour before an addition of 25 ml sodium phosphate buffer (20 mM, pH 7.4) containing either 0.6 g dex-1500 or 2.0 g dex-10000. The reaction was carried out at 30 oC with constant shaking for 24 h in the dark.
After reaction, the mixture was concentrated by rotary evaporation followed by repeated methanol precipitation as described above.
Conjugation of folate-amino-dextran to polyethylenimine. The desired amount of folate-amino-dextran was reacted with 100 mg polyethylenimine in the presence of 320 mg sodium cyanoborohydride in 20 ml borate buffer (100 mM,pH 8.0). The reaction was carried out at 45 oC with constant shaking for 48 h. After reaction, the mixture was dialyzed against 2-l of distilled water 6 times using Spectra/Por membrane (MWCO = 15000) followed by lyophilization.
Preparation of folate-PEI conjugate.
The desired amount of folate (105, 315, or 425 mg) was mixed with a solution containing PEI (400 mg), pyridine (1 ml), and an equal molar amount of DDC in 20 ml DMSO18. The reaction was carried out at 30 oC for 24 h with constant shaking. The conjugated PEI was purified by dialysis against 2-l of distilled water for 6 times using Spectra/Por membrane (MWCO = 15000) followed by lyophilization.
Preparation of dextran-PEI conjugate.
PEI (1.0 g) was mixed with the desired amount of dextran plus 160 mg sodium cyanoborohydride in 25 ml borate buffer (100 mM, pH 8.0)4. The reaction mixture was incubated at 45 oC for 48 h with constant shaking. The PEI conjugate was lyophilized after the removal of sodium cyanoborohydride and the unreacted dextran by dialysis.
Determination of the amount of primary amine on each PEI conjugate.
The amount of primary amine remaining on each PEI conjugate was determined by TNBS.
Briefly, 0.5 ml of each sample (1 mg/ml) was mixed with 0.25 ml of TNBS in 0.1 M sodium bicarbonate buffer (pH 8.5). The mixture was incubated at 37 oC for 2 h followed by an addition of 0.25 ml of 10% SDS and 0.125 ml of 1 N HCl. The absorbance of each sample was measured at 335 nm by a spectrophotometer (Jasco, Tokyo, Japan). The degree of grafting was calculated by the difference in the amount of primary amine on unmodified PEI and that on each conjugate32.
Determination of the amount of folate on the PEI conjugate.
Folate was measured at its characteristic absorbance of 368 nm18. The amount of folate on the PEI conjugate was determined according to the calibration curve constructed by unreacted folate.
Titration of the PEI conjugates.
Unmodified and the PEI conjugates were diluted to a final concentration of 0.1 mg/ml with 0.1 N NaCl. The solution was adjusted to pH 8.0 before titration proceeded. An aliquot of 25 ml 0.1 N HCl was successively added into 30 ml of 0.1 mg/ml polymer solution, and the changes in pH were monitored by a pH meter (MP 220; Mettler, Todelo, Switzerland)
Measurements of the size and zeta potential of the DNA-polymer complexes.
A Zetamaster system (Malvern, Malvern, UK) was used to measure the sizes and zeta potentials of the DNA-polymer complexes prepared with either unmodified PEI or each conjugate as previously described. Briefly, the desired amount of each cationic polymer was mixed with 40 µg of DNA at an N/P ratio of nine, where the N/P ratio was used to indicate the charge ratio of the cationic polymer to DNA with N representing the number of amine in the polymer and P representing the number of phosphate in the DNA. The sizes and zeta-potentials of the DNA-polymer complexes were examined using a salt-free buffer (20 mM Hepes, 5.2% glucose, pH 7.0) and DMEM without phenol red.
Cell culture.
KB cells, a human epidermal carcinoma cell line, were maintained at 37 oC, 5% CO2, and 100% humidity in folate-deficient Dulbecco’s modified Eagle medium supplemented with 1% antibiotics of penicillin-streptomycin-amphotericin and 10% fetal bovine serum. Cells were seeded onto 35 mm petri dishes at 20,000 cells/cm2 16 h before transfection.
Transfection. The transfection reagent was prepared by mixing 2 µg plasmid with the desired conjugate at various N/P ratios (4, 6, 9, 12) in 0.2 ml of folate-deficient DMEM. The mixture was allowed to stand at room temperature for 15 to 30 min, and then 0.8 ml of folate-deficient DMEM was added to make the transfection reagents. For transgene expression, cells were cultured with the transfection reagents for 6 h and then replaced with culture medium for additional 18 h. For the measurement of plasmid delivery, EMA-labeled plasmid instead of unlabeled plasmid was used in the preparation of transfection reagent at the N/P ratio of 9. The cells were incubated with the EMA-labeled plasmid for 6 h at 37 oC and 4 oC to determine the amount of plasmid associated with the cells and that adhered onto the cell surfaces, respectively.
Cytotoxicity assay by MTT.
The transfected cells at the N/P ratio of 9 were washed twice with one ml of phosphate buffered saline (PBS; pH 7.2). Then, an aliquot of 1 ml PBS solution containing MTT (0.5 mg/ml) was added to the cultured cells. After 4 h incubation, DMSO was used to solubilize the colored formazan product, and the absorbance at 540 nm was measured.
Flow cytometric analysis.
The transfected cells were harvested with PBS containing 0.15% trypsin/1 mM EDTA. After the cells were detached, an equal volume of medium was added, and the cells were fixed in the presence of 1% paraformaldehyde for 10 min at room temperature. The fixed cells were then washed with 3 ml of PBS containing 1% paraformaldehyde three times. The final pellet was resuspended in PBS containing 1% paraformaldehyde and stored at 4 oC for flow cytometric analysis. For each sample, 20,000 events were collected by a flow cytometer (FACS Calibur; BD Biosciences, San Jose, CA) equipped with an argon-ion laser and a red diode laser. The analysis of the transfected cells was performed by the CellQuest software (BD Biosciences, San Jose, CA) as previously reported30.
Results and Discussion
Preparation of PEI conjugates
Three different types of conjugates were prepared in this study as listed in Table 1.
Dextran and folate reacted directly with PEI to produce dextran-PEI and folate-PEI conjugates, respectively. Dextran was conjugated onto PEI by a reductive reaction between the aldehyde group of dextran and the amine group of PEI. Two levels of dex-1500 conjugation and three levels of dex-10000 conjugation were generated: P-8LD and P-17LD;
P-1HD, P-11HD, and P-19HD. The former two denotations represented the conjugates containing around 8 molecules and 17 molecules of dex-1500 per PEI, respectively, and the latter three denotations represented the conjugates containing around 1 molecule, 11 molecules, and 19 molecules of dex-10000 per PEI, respectively. Folate was conjugated by an amide bond between the carboxyl group of folate and the amine group of PEI. Three levels of folate conjugation were synthesized: P-10FA, P-25FA, and P-35FA, which contained around 10, 25, and 35 molecules of folate per PEI, respectively.
For preparation of the folate-dextran-PEI conjugate, dextran was first modified with 2-chloroethylamine because 2-chloroethylamine could react with either the hydroxyl or the aldehyde group of dextran. After amination, one molecule of the dex-1500 and dex-10000 contained 1.5 and 7 amino groups on average, respectively. A further treatment of the amino-dextran with periodate resulted in 1.3 – 2.5 and 5 – 6 molecules of reactive aldehyde per dex-1500 and dex-10000 molecules, respectively. Folate was then conjugated onto the amino-end of the oxidized amino-dextran via an amide bond. Subsequently the aldehyde groups of the folate-dextran molecules reacted with the primary amines of PEI. By this way, a couple of folate-dextran-PEI conjugates were synthesized: P-7LD-7FA, P-8LD-31FA, P-20LD-21FA, P-1HD-1FA, P-9HD-21FA, P-9HD-35FA, and P-16HD-38FA, each of which possessed folate on the conjugated dextran.
Transfection mediated by DNA-polymer complexes
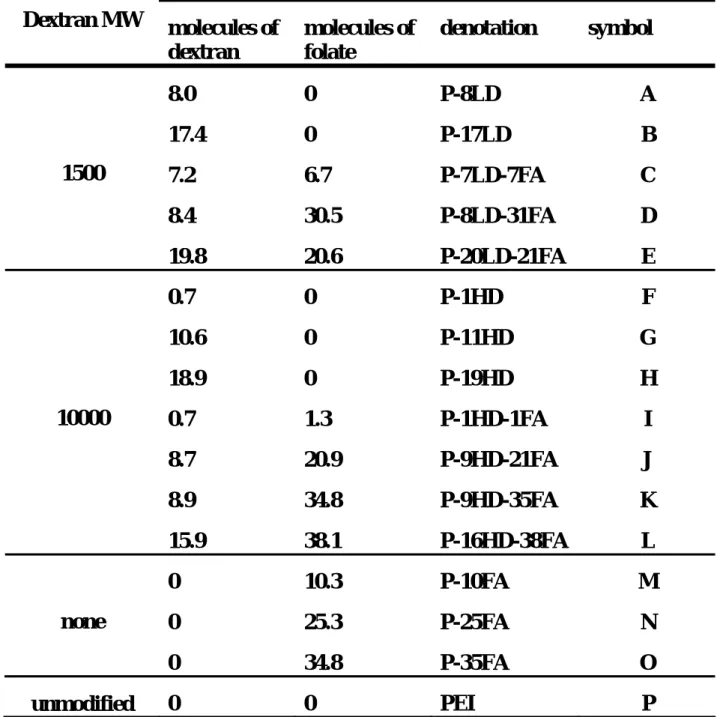
After the cells were incubated with various DNA-polymer complexes, flow cytometry was used to quantitate the fraction of the cells expressing GFP, which was set as an indicator of transfection efficiency in this study. The transfection efficiencies of certain polymers were found to exhibit a function of the N/P ratio. At the N/P ratio of 4, the GFP positive cells were very scarce under a fluorescence microscope for all the polymers, and were hardly detected by the flow cytometer (data not shown). When the N/P ratio was raised to 6, GFP positive cells could be detected mainly for the folate-PEI conjugates and unmodified PEI, the former of which showed very minimal efficiencies compared with the latter (Fig. 1A). An increase of the N/P ratio to 9 enabled some polymers, such as P-8LD, P-7LD-7FA, P-8LD-31FA, and P-1HD-1FA to produce some detectable GFP positive cells (Fig. 1B). At this charge ratio, the efficiencies of the PEI-folate conjugates and unmodified PEI were also increased. When the N/P ratio was increased to 12, the transfection efficiencies were further enhanced for the conjugates capable of producing GFP positive cells at the N/P ratio of 9 except for the unmodified PEI (Fig 1C).
An optimal degree of folate conjugation seemingly existed to improve the transfection efficiency. A moderate degree of additional folate on either the dextran-PEI conjugates or unmodified PEI could enhance the transfection efficiency, but the efficiency dropped when the degree of folate conjugation was further increased; for example, the efficiencies of
P-7LD-7FA and P-25FA were the highest ones among the group: P-8LD, P-7LD-7FA, and P-8LD-31FA and another group: P-10FA, P-25FA, and P-35FA, respectively. The conjugates with dex-10000, however, failed to produce any detectable level of GFP positive cells at all of the N/P ratios except for the PEI conjugated with a very minimal level of dex-10000 and folate (P-1HD and P-1HD-1FA). Unmodified PEI was the most effective transfection agent at all the N/P ratios tested in this study, suggesting that the conjugation diminished the transfection effectiveness of PEI. In an effort to understand what factors altered the transfection effectiveness of PEI after conjugation, we investigated the physicochemical properties of the DNA-polymer complexes, the level of intracellular plasmid, and the buffering capacity of the PEI conjugates.
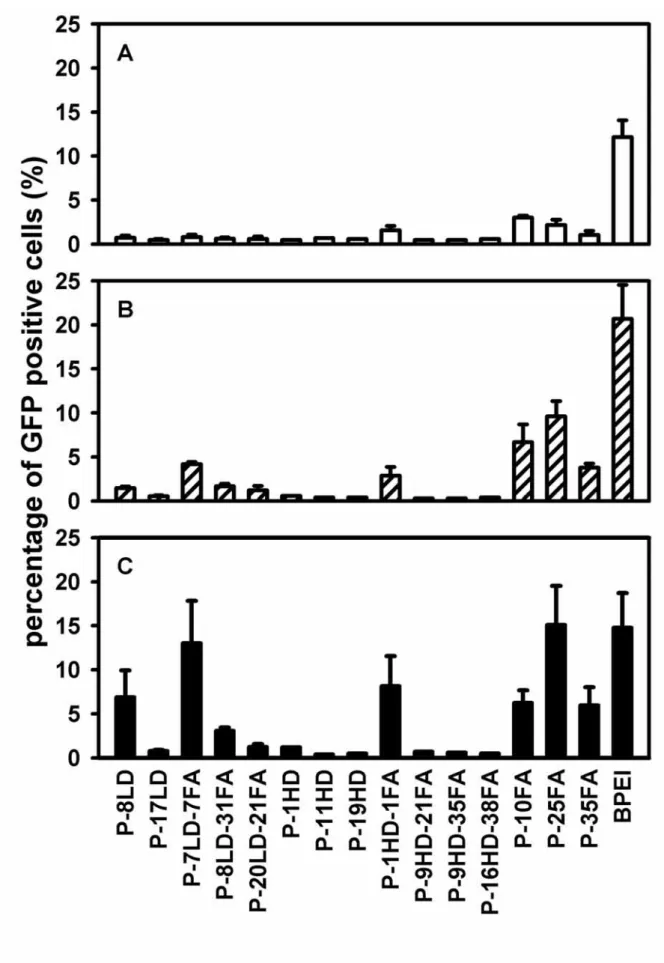
Physicochemical properties of the DNA-polymer complexes in the salt free and DMEM buffers
For the measurements of the physiochemical properties of the complexes, the DNA-polymer complexes were prepared at the N/P ratio of 9 in the salt-free and DMEM buffers, respectively. The sizes of the complexes were found to be below 200 nm in the salt-free buffer when PEI was conjugated with either dex-1500 or folate (Fig. 2A). But when the above complexes were prepared in the DMEM buffer, the sizes became larger than 1500 nm. On the other hand, the sizes of the complexes prepared with PEI conjugated with dex-10000 exhibited slightly larger sizes (200 – 400 nm) in the salt-free buffer (Fig. 2A). The sizes of these complexes, however, remained at the approximate magnitude (< 400 nm) when the DMEM buffer was used, indicating that the conjugation of dex-10000 could retard the salt-induced aggregation of complexes and thereby stabilized the complexes. These observations were consistent with a previous report22, which demonstrated that the conjugation of large dextran could better stabilize the DNA-polymer complexes than the conjugation of small dextran.
The conjugation of folate seemed to affect the zeta potentials of the complexes prepared in the DMEM buffer. For the approximate degrees of dextran conjugation, the polymers containing folate showed larger negative values of zeta potential in comparisons with those containing dextran alone; for example P-8LD-31FA vs. P-8LD, P-20LD-21FA vs. P-17LD, P-9HD-21FA vs. P-11HD, and P-16HD-38FA vs. P-19HD (Fig. 2B). These observations might reflect the nature of folate conjugation, which were on the exterior dextran of the complexes. The surface charges of the complexes were also more negative when dex-10000 was used than dex-1500, suggesting that dex-10000 provided a better charge shield.
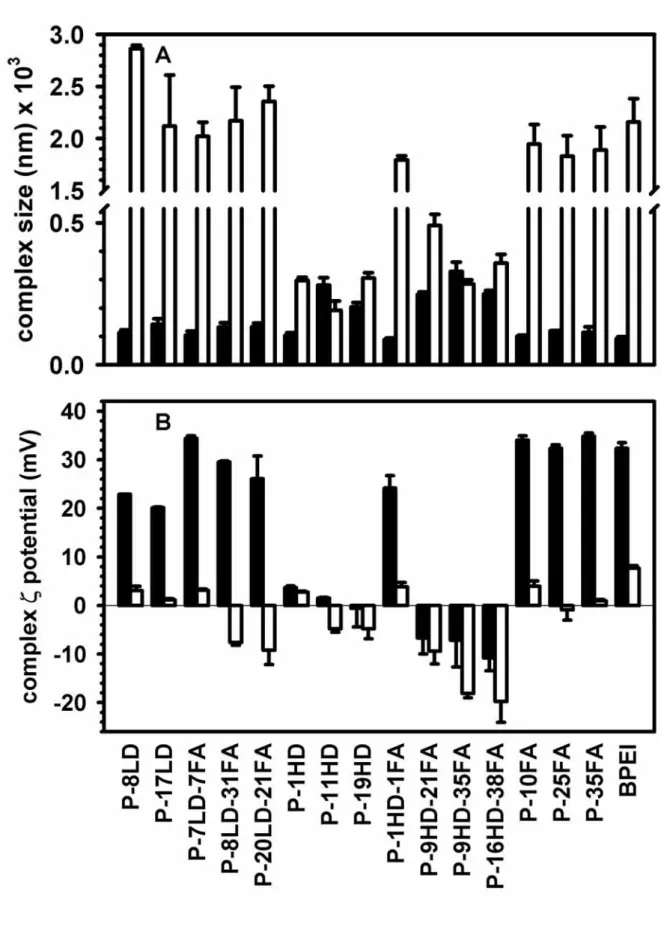
Entry of plasmid delivered by the DNA-polymer complexes
The cellular entry of DNA-polymer complexes was via endocytosis, which did not occur at the reduced temperature of 4 oC. At 4 oC the plasmid remained only bound on the cell membranes. The amounts of membrane-bound complexes were at the same level for all the polymers (p < 0.1) (Fig. 3A). The intracellular amounts of the delivered plasmid could be estimated by the differences in the mean fluorescence intensities of the cells incubated with EMA-labeled plasmid at 4 oC and those at 37 oC (Fig. 3B). In this study, flow cytometry was used to measure the changes in the cellular fluorescence intensity of each individual cell after the cells were incubated with the EMA labeled plasmid. Although the conjugation of folate was attempted to enable the targeted delivery of plasmid into the cells, the intracellular amounts of plasmid delivered by various polymers were of the same level (p < 0.1) irrespective of the grafting degrees and of the types of conjugation.
The intracellular amount of plasmid has been reported to be a critical factor in determining the level of transgene expression33. Large sizes of the complexes have been shown to increase the level of transgene expression by delivering more plasmid into the cells34. High surface charge of the complexes has also been shown to promote the cellular entry of plasmid. In this study, however, neither the larger sizes nor higher zeta potentials of the complexes resulted in higher amounts of intracellular plasmid. The approximately equal levels of intracellular plasmid might be presumably due to the saturation of the endocytic entry rate.
Measurements of buffering capacity
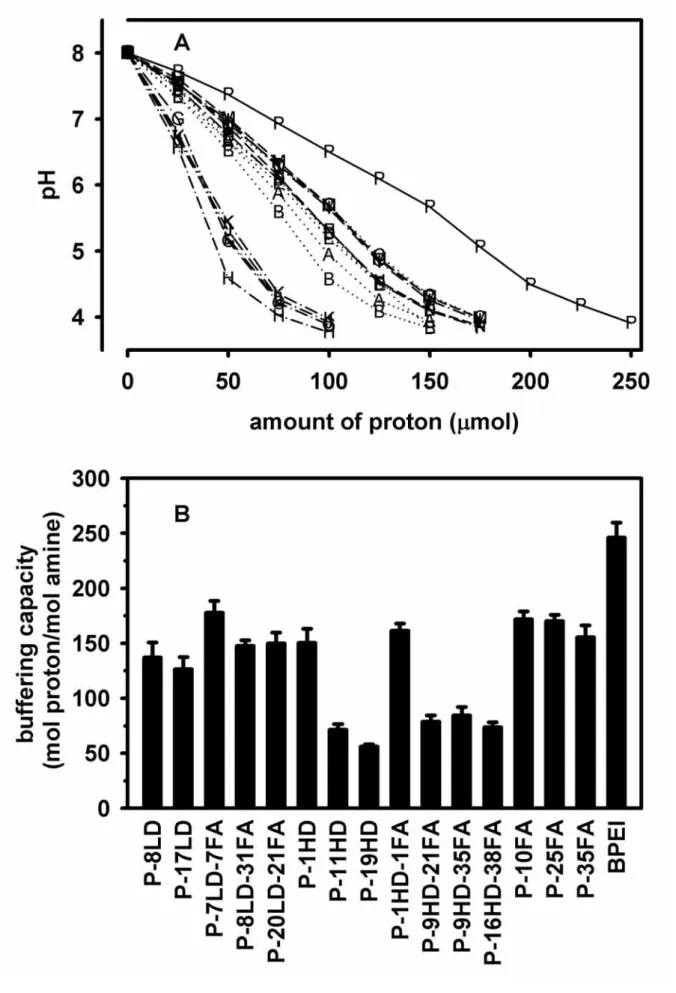
Because the buffering capacity of polycationic vectors affected the extents of the DNA-polymer complexes released from the endosomes, titration was used to examine whether the conjugation affected the buffering capacity of PEI. The polymers of high buffering capacity would undergo a smaller change in pH when the same amount of HCl was added into the polymer solutions during titration. Higher extents of changes in pH were found for the PEI conjugates than for the unmodified PEI (Figure 4A), indicating that the buffering capacity was reduced after conjugation. Because the pH of the endosomes was maintained around 5.5 to 6 by ATP-driven pumps, the buffering capacity of a polymer could be defined as the amount of proton required for a change in pH from 8.0 to 5.5 during in vitro titration in this study.
The extents of decreases in buffering capacity after conjugation seemed to depend on both the degrees of conjugation and the types of conjugated molecules (Fig. 4B). Large molecules of no charge, such as dex-10000, severely diminished the buffering capacity even under a very minimal level of conjugation. Only one conjugated molecule of dex-10000 was sufficient to result in 35% of buffering capacity loss (P-1HD), and the dextran-PEI conjugate possessed merely 1/5 of the original buffering capacity when 19 molecules of dex-10000 were conjugated onto PEI (P-19HD). For the approximate degree of dex-1500 conjugation, P-17LD retained half of the original buffering capacity. On the other hand, a higher degree of folate conjugation (P-35FA) still could retain more than half of the buffering capacity.
Besides, additional conjugation of folate onto dextran hardly further decreased the buffering capacity of the dextran-PEI conjugates, for example P-8LD-31FA vs. P-8LD, P-1HD vs.
P-1HD-1FA, and P-9HD-21FA vs. P-9HD-35FA .
The decreases in buffering capacity might be due to the hindrance of proton dissociation after conjugation as revealed by a previous report35. PEI conjugated with large-size molecules, such as dex-10000, might create higher hindrances to lower the extent of proton dissociation, and thus abolished the buffering capacity. The decreased buffering capacity might insufficiently disrupt the endosomes to aid the DNA release, and caused the low efficiency of transgene expression.
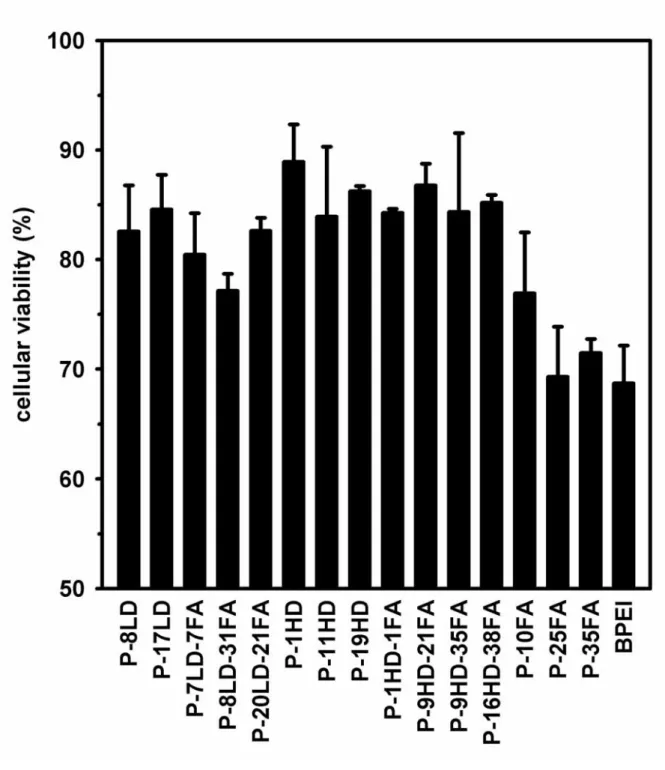
Measurements of cellular cytotoxicity
Low cell viability was found when the cells were transfected with PEI. Conjugation of either dex-10000 or dex-1500 could moderate the cellular cytotoxicity of the unmodified PEI (Fig. 5). Such improvements in cytotoxicity were the same for various levels of either dex-1500 or dex-10000 conjugation (p > 0.1). The direct conjugation of folate onto PEI, however, hardly promoted cell viability. Additional folate on the conjugated dextran was also found to have little effects on further improving the viability.
Effects of intracellular buffering capacity on cytotoxicity and transfection efficiency
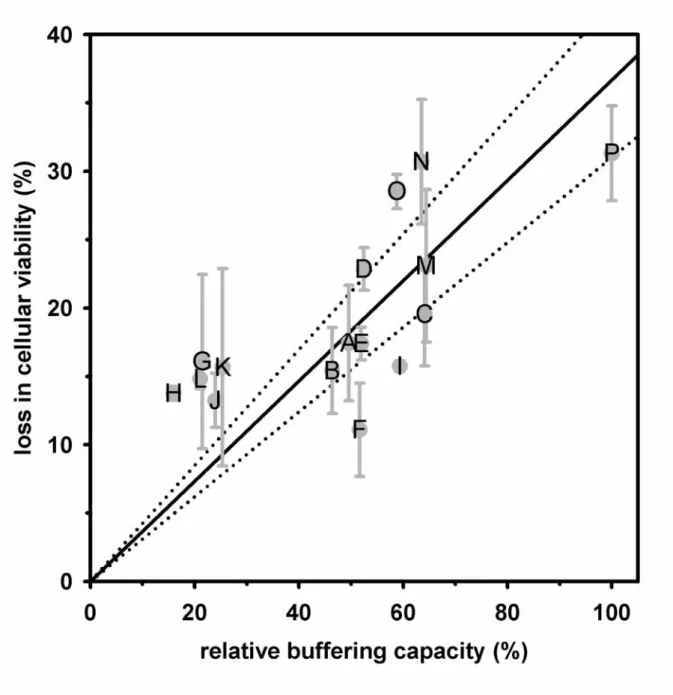
Several recent studies have demonstrated that buffering capacity is an important characteristic of PEI for its high efficiency of transgene expression10, 11, 13, 36. Conjugation onto the amines of PEI diminished the buffering capacity as revealed by other reports30, 37 as well as shown in this study, indicating that using the fluorophore-labeled PEI to monitor the intracellular amount of polymer might be unable to reflect the actual sponge effect of either unmodified PEI or the PEI conjugates. Therefore, we employed the intensity of EMA fluorescence to represent the intracellular amount of polymer delivered into each cell by assuming that the polymer was proportionally associated with the EMA-plasmid to enter across the cell membrane. Thus, the intracellular buffering capacity was defined as the multiplication of the EMA intensity (Fig. 3, panel B) and the buffering capacity of the corresponding polymer (Fig. 4, panel B), and was further expressed in terms of percentages by setting the intracellular buffering capacity of unmodified PEI as 100%. Fig. 6 revealed an approximately linear dependence between the loss in cellular viability and the relative intracellular buffering capacity when the relationship of these two parameters was examined.
The high buffering capacity constituted the sponge effect of PEI to disrupt the endosomes, and also induced high cellular cytotoxicity presumably due to the unbalances of intracellular protons and the disruption of intracellular organelles. On the other hand, the decreased buffering capacity might be unable to induce a sufficient influx of chloride ion into the endosomes, and therefore maintained high cell viability.
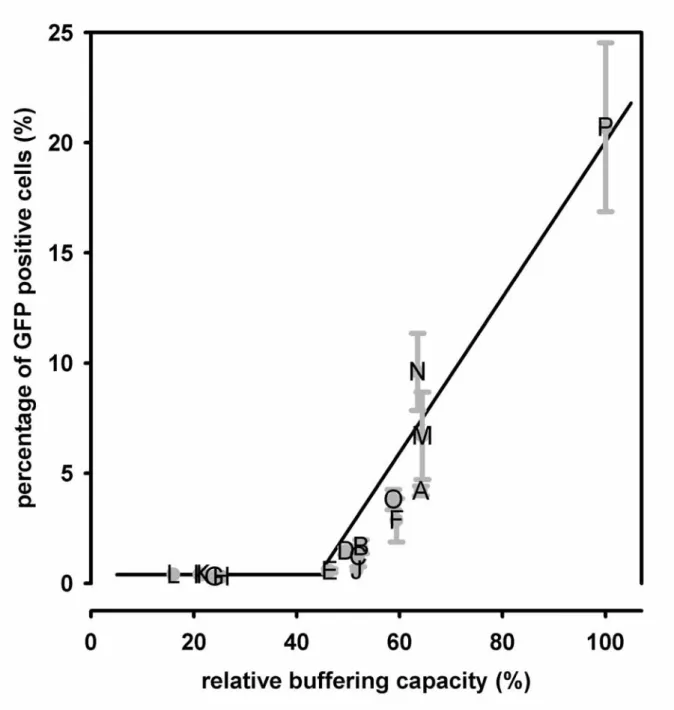
When the intracellular buffering capacity was used as a parameter to examine its relationship with the transfection efficiency, a threshold of intracellular buffering capacity, about 50% of the unmodified PEI capacity, was found for the minimal detection of GFP positive cells by flow cytometry (Fig. 7). Beyond the threshold, the percentage of GFP positive cells followed a trend increasing with the intracellular buffering capacity roughly in a linear way. It could be realized that the low intracellular buffering capacity was insufficient to disrupt the endosomes for the release of plasmid. Therefore, dextran-PEI and folate-dextran-PEI conjugates produced very minimal levels of transgene expression mainly due to the severely abolished buffering capacity whereas the folate-PEI conjugates still maintained the moderate percentages of GFP expressing cells due to the less decreases in buffering capacity.
Conclusions
In this study, the cellular cytotoxicity and the efficiency of transgene expression were found to quantitatively depend upon the buffering capacity of the PEI conjugates. The conjugation onto PEI through primary amine resulted in decreases in buffering capacity that might be attributed to the accompanying decreases in transfection efficiency and cell cytotoxicity. A high-degree conjugation of a large-size molecule, such as dex-10000, could stabilize the DNA-polymer complexes, and improve cell viability. Nevertheless such conjugation failed to produce detectable transgene expression presumably because of the significant decreases in buffering capacity. The quantitative descriptions of the dependence between buffering capacity and transfection efficiency as well as cellular cytotoxicity could be expected to provide a basis for the design of efficient polycationic vectors.
References
(1) Davis, M. E. Non-viral gene delivery systems. Curr. Opin. Biotechnol. 2002, 13, 128-131.
(2) Nishikawa, M.; Huang, L. Nonviral vectors in the new millennium: delivery barriers in gene transfer. Hum. Gene Ther. 2001, 12, 861-870.
(3) Boussif, O.; Lezoualc'h, F.; Zanta, M. A.; Mergny, M. D.; Scherman, D.; Demeneix, B.; Behr, J. P. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: polyethylenimine. Proc. Natl. Acad. Sci. USA 1995, 92, 7297-7301.
(4) Erbacher, P.; Bettinger, T.; Belguise-Valladier, P.; Zou, S.; Coll, J. L.; Behr, J. P.;
Remy, J. S. Transfection and physical properties of various saccharide, poly(ethylene glycol), and antibody-derivatized polyethylenimines (PEI). J. Gene Med. 1999, 1, 210-222.
(5) Tang, M. X.; Szoka, F. C. The influence of polymer structure on the interactions of cationic polymers with DNA and morphology of the resulting complexes. Gene Ther.
1997, 4, 823-832.
(6) Wolfert, M. A.; Seymour, L. W. Atomic force microscopic analysis of the influence of the molecular weight of poly(L)lysine on the size of polyelectrolyte complexes formed with DNA. Gene Ther. 1996, 3, 269-273.
(7) Leong, K. W.; Mao, H. Q.; Truong-Le, V. L.; Roy, K.; Walsh, S. M.; August, J. T.
DNA-polycation nanospheres as non-viral gene delivery vehicles. J. of Controll.
Release 1998, 53, 183-193.
(8) Dunlap, D. D.; Maggi, A.; Soria, M. R.; Monaco, L. Nanoscopic structure of DNA condensed for gene delivery. Nucleic Acids Res. 1997, 25, 3095-3101.
(9) Mislick, K. A.; Baldeschwieler, J. D. Evidence for the role of proteoglycans in cation-mediated gene transfer. Proc. Natl. Acad. Sci. USA. 1996, 93, 12349-12354.
(10) Sonawane, N. D.; Szoka, F. C., Jr.; Verkman, A. S. Chloride accumulation and swelling in endosomes enhances DNA transfer by polyamine-DNA polyplexes. J. Biol.
Chem. 2003, 278, 44826-44831.
(11) Forrest, M. L.; Pack, D. W. On the kinetics of polyplex endocytic trafficking:
implications for gene delivery vector design. Mol. Ther. 2002, 6, 57-66.
(12) Godbey, W. T.; Wu, K. K.; Mikos, A. G. Tracking the intracellular path of poly(ethylenimine)/DNA complexes for gene delivery. Proc. Natl. Acad. Sci. USA 1999, 96, 5177-5181.
(13) Kichler, A.; Leborgne, C.; Coeytaux, E.; Danos, O. Polyethylenimine-mediated gene delivery: a mechanistic study. J. Gene Med. 2001, 3, 135-144.
(14) Thomas, M.; Klibanov, A. M. Enhancing polyethylenimine's delivery of plasmid DNA into mammalian cells. Proc. Natl. Acad. Sci. USA 2002, 99, 14640-14645.
(15) Kichler, A. Gene transfer with modified polyethylenimines. J. Gene Med. 2004, 6 Suppl 1, S3-10.
(16) Fajac, I.; Allo, J. C.; Souil, E.; Merten, M.; Pichon, C.; Figarella, C.; Monsigny, M.;
Briand, P.; Midoux, P. Histidylated polylysine as a synthetic vector for gene transfer into immortalized cystic fibrosis airway surface and airway gland serous cells. J.
Gene Med. 2000, 2, 368-378.
(17) Putnam, D.; Gentry, C. A.; Pack, D. W.; Langer, R. Polymer-based gene delivery with low cytotoxicity by a unique balance of side-chain termini. Proc. Natl. Acad. Sci. USA 2001, 98, 1200-1205.
(18) Benns, J. M.; Mahato, R. I.; Kim, S. W. Optimization of factors influencing the transfection efficiency of folate-PEG-folate-graft-polyethylenimine. J. Control.
Release 2002, 79, 255-269.
(19) Blessing, T.; Kursa, M.; Holzhauser, R.; Kircheis, R.; Wagner, E. Different strategies for formation of pegylated EGF-conjugated PEI/DNA complexes for targeted gene delivery. Bioconjug. Chem. 2001, 12, 529-537.
(20) Ogris, M.; Brunner, S.; Schuller, S.; Kircheis, R.; Wagner, E. PEGylated DNA/transferrin-PEI complexes: reduced interaction with blood components, extended circulation in blood and potential for systemic gene delivery. Gene Ther.
1999, 6, 595-605.
(21) Choi, Y. H.; Liu, F.; Park, J. S.; Kim, S. W. Lactose-poly(ethylene glycol)-grafted poly-L-lysine as hepatoma cell-tapgeted gene carrier. Bioconjug. Chem. 1998, 9, 708-718.
(22) Tseng, W. C.; Jong, C. M. Improved stability of polycationic vector by dextran-grafted branched polyethylenimine. Biomacromolecules 2003, 4, 1277-1284.
(23) Kircheis, R.; Kichler, A.; Wallner, G.; Kursa, M.; Ogris, M.; Felzmann, T.; Buchberger, M.; Wagner, E. Coupling of cell-binding ligands to polyethylenimine for targeted gene delivery. Gene Ther. 1997, 4, 409-418.
(24) Li, S.; Tan, Y.; Viroonchatapan, E.; Pitt, B. R.; Huang, L. Targeted gene delivery to pulmonary endothelium by anti-PECAM antibody. Am. J. Physiol. 2000, 278, L504-511.
(25) O'Neill, M. M.; Kennedy, C. A.; Barton, R. W.; Tatake, R. J. Receptor-mediated gene delivery to human peripheral blood mononuclear cells using anti-CD3 antibody coupled to polyethylenimine. Gene Ther. 2001, 8, 362-368.
(26) Harbottle, R. P.; Cooper, R. G.; Hart, S. L.; Ladhoff, A.; McKay, T.; Knight, A. M.;
Wagner, E.; Miller, A. D.; Coutelle, C. An RGD-oligolysine peptide: a prototype construct for integrin-mediated gene delivery. Hum. Gene Ther. 1998, 9, 1037-1047.
(27) Diebold, S. S.; Kursa, M.; Wagner, E.; Cotten, M.; Zenke, M. Mannose polyethylenimine conjugates for targeted DNA delivery into dendritic cells. J. Biol.
Chem. 1999, 274, 19087-19094.
(28) Lee, R. J.; Huang, L. Folate-targeted, anionic liposome-entrapped polylysine-condensed DNA for tumor cell-specific gene transfer. J. Biol. Chem. 1996, 271, 8481-8487.
(29) Li, S.; Deshmukh, H. M.; Huang, L. Folate-mediated targeting of antisense oligodeoxynucleotides to ovarian cancer cells. Pharm. Res. 1998, 15, 1540-1545.
(30) Tseng, W. C.; Tang, C. H.; Fang, T. Y. The role of dextran conjugation in transfection mediated by dextran-grafted polyethylenimine. J. Gene Med. 2004, 6, 895-905.
(31) Bernstein, A.; Hurwitz, E.; Maron, R.; Arnon, R.; Sela, M.; Wilcheck, M. Higher antitumor efficacy of daunomycin when linked to dextran: In vivo and in vitro studies.
J. Natl. Cancer Inst. 1978, 60, 379-384.
(32) Johnson, T. W.; Klotz, I. M. Preparation and characterization of some derivatives of polyethylenimine. Macromolecules 1974, 7, 149-153.
(33) Tseng, W. C.; Haselton, F. R.; Giorgio, T. D. Transfection by cationic liposomes using simultaneous single cell measurements of plasmid delivery and transgene expression.
J. Biol. Chem. 1997, 272, 25641-25647.
(34) Ogris, M.; Steinlein, P.; Kursa, M.; Mechtler, K.; Kircheis, R.; Wagner, E. The size of DNA/transferrin-PEI complexes is an important factor for gene expression in cultured cells. Gene Ther. 1998, 5, 1425-1433.
(35) Suh, J.; Lee, S. H.; Kim, S. M.; Hah, S. S. Conformational flexibility of polyethylenimine and its derivative. Bioorg. Chem. 1997, 25, 221-231.
(36) Bieber, T.; Meissner, W.; Kostin, S.; Niemann, A.; Elsasser, H. P. Intracellular route and transcriptional competence of polyethylenimine-DNA complexes. J Control.
Release 2002, 82, 441-454.
(37) Akinc, A.; Thomas, M.; Klibanov, A. M.; Langer, R. Exploring polyethylenimine-mediated DNA transfection and the proton sponge hypothesis. J.
Gene Med. 2005.
Table 1 The estimated degrees of conjugation on each PEI conjugate Degree of conjugation
Dextran MW molecules of dextran
molecules of folate
denotation symbol
8.0 0 P-8LD A
17.4 0 P-17LD B
7.2 6.7 P-7LD-7FA C
8.4 30.5 P-8LD-31FA D
1500
19.8 20.6 P-20LD-21FA E
0.7 0 P-1HD F
10.6 0 P-11HD G
18.9 0 P-19HD H
0.7 1.3 P-1HD-1FA I
8.7 20.9 P-9HD-21FA J
8.9 34.8 P-9HD-35FA K
10000
15.9 38.1 P-16HD-38FA L
0 10.3 P-10FA M
0 25.3 P-25FA N
none
0 34.8 P-35FA O
unmodified 0 0 PEI P
Figure Legends
Figure 1 Transfection efficiencies of the PEI conjugates at different N/P ratios.
The DNA-polymer complexes were prepared with the PEI conjugates as listed in Table 1, and then used to transfect cells. The transfection efficiency was indicated by the percentage of GFP positive cells measured by flow cytometry. The efficiencies of various conjugates were examined for the complexes formed at N/P ratio of 6 (panel A), 9 (panel B), and 12 (panel C). (mean ± S.D.; n = three independent transfection experiments)
Figure 2 Sizes and zeta potentials of the DNA-polymer complexes in the DMEM and salt-free buffers.
The synthesized PEI conjugates were used to form complexes with DNA in the presence of salt-free buffer (solid bars) and DMEM buffer (hollow bars). The sizes and zeta potentials of the complexes were displayed in panel A and panel B, respectively. (mean ± S.D.; n = three independent complex preparations)
Figure 3 Mean fluorescence intensities of the cells after incubation with the DNA-polymer complexes containing EMA-labeled plasmid
Flow cytometry was used to analyze the distribution of the fluorescence intensities of the cells incubated with the DNA-polymer complexes containing EMA-labeled plasmid. The mean values of the fluorescence distributions were used to indicate the cellular amounts of plasmid at 4 oC (panel A, solid bar) and 37 oC (panel A, hollow bar). The differences in the fluorescence intensities at 4 oC and 37 oC represented the intracellular amounts of the EMA-labeled plasmid delivered by the conjugates (panel B). (mean ± S.D.; n=three independent transfection experiments)
Figure 4 Buffering capacities of the PEI conjugates
Panel A displayed a typical titration curve of each conjugate titrated with 0.1 N HCl. The alphabets in panel A were referred to the conjugates listed in Table 1. The solid line (—) represented the unmodified PEI, the short-dash line (···) represented either the dex1500-PEI or folate-dex1500-PEI conjugates, the dash-dot line (−·−) represented either the dex10000-PEI or folate-dex10000-PEI conjugates, and the long-dash line (−−−) represented the folate-PEI conjugates. Panel B displayed the amounts of proton required to result in a change in pH from 8.0 to 5.5. (mean ± S.D.; n = three independent titration experiments)
Figure 5 Cellular cytotoxicities of the DNA-polymer complexes
Cytotoxicity was assayed by the MTT method, and expressed as the percentages of cell viability by setting the untransfected cells as 100%. (mean ± S.D.; n = three independent transfection experiments)
Figure 6 Dependence of cytotoxicity and the buffering capacity
Loss in cellular viability was used to indicate the cytotoxicity by setting the untransfected cells as 0%. The relative buffering capacity was defined as the multiplication of the mean fluorescence intensity of intracellular EMA-plasmid (Fig. 3, panel B) and the buffering capacity of the corresponding conjugate (Fig. 4, panel B) by setting the unmodified PEI as 100%. The solid line showed a linear regression between the loss in cellular viability and the relative buffering capacity, and the dash line showed the linear regression with 99% of confidence. The alphabets indicated each conjugate as listed in Table 1.
Figure 7 Dependence of transfection efficiency and buffering capacity
The relative buffering capacity was defined as the multiplication of the mean fluorescence intensity of intracellular EMA-plasmid (Fig. 3, panel B) and the buffering capacity of the corresponding conjugate (Fig. 4, panel B) by setting the unmodified PEI as 100%. The relationship showed that a minimal value of the relative buffering capacity was required for a detectable level of GFP positive cells.
Figure 1 Transfection efficiencies of the PEI conjugates at different N/P ratios.
Figure 2 Sizes and zeta potentials of the DNA-polymer complexes in the DMEM and salt-free buffers.
Figure 3 Mean fluorescence intensities of the cells after incubation with the DNA-polymer complexes containing EMA-labeled plasmid
Figure 4 Buffering capacities of the PEI conjugates
Figure 5 Cellular cytotoxicities of the DNA-polymer complexes
Figure 6 Dependence of cytotoxicity and the buffering capacity
Figure 7 Dependence of transfection efficiency and buffering capacity
計畫成果自評
本研究內容與原計畫相符並達成預期目標,研究成果已於學術期刊發表。
出席國際學術會議心得報告
計畫編號 95-2221-E-011-079-
計畫名稱 聚乙烯亞胺接枝對其載體特性及轉染效率的影響(2/2)
出國人員姓名 服務機關及職稱
曾文祺
國立臺灣科技大學化學工程系 會議時間地點 July 20 – July 24, 2007 美國波士頓
會議名稱 第二十一屆蛋白質協會年會
發表論文題目
Assessment of Hydrogen Bonds between Maltooligosyltrehalose Synthase and Maltopentaose by Computer Simulation and Site Directed Mutagenesis(摘要編 號為 499)
參加會議經過及與會心得
2007 年 6 月 20 至 24 日於波士頓舉行的第廿一屆蛋白質年會,此次年會的主軸是從出生 至死亡(from birth to death),藉以標顯蛋白質在生物體中所拌演重要的角色。此次年會共有 609 篇摘要,近千人與會。
會議首日由劍橋大學的 Dr. Fersht 致開幕詞揭開序幕,當日有两大主題分別是蛋白質摺 疊(protein folding)與系統生物學(Systems Biology)。傍晚則是本次大會的 plenary lecture,演講 者是來自美國加州大學舊金山分校的 Professor Peter Walter,講題為”未摺疊蛋白質的應答:由 內質網至細胞核的胞內訊息”。 內容主要介紹內質網對細胞內蛋白質摺疊的重要性,當蛋白 質摺疊超過其可負載的程度,將產生對 mRNA splicing 的訊息調控。一旦內質網蛋白質摺疊 的能力與其負載間無法取得平衡時,將進一步誘發細胞凋亡。此一未摺疊蛋白質的應答對細 胞的存亡至為重要,也對許多疾病如病毒感染、糖尿病及癌症影響至鉅。
次日年會探討的主題有四分別是蛋白的結構與動力學、蛋白質體學、蛋白質錯誤摺疊與 聚集及酵素催化與配位體辨認,隔日接著登塲的四個主題為蛋白質轉譯、蛋白質設計、分子 媒介與單分子研究,接續再一日的主題為蛋白質品質控制與蛋白質資訊學得許,最終一日以”
由生物物理學至生物學: 研究細胞內蛋白質” 為主題,劃下此次大會的句點。大會中並頒獎 表揚在蛋白質領域中做出重要研究貢獻的學者,分別是 The Stein and Moore Award 的 Professor Paul Schimmel(來自 The Scripps Research Insititute);The Hans Neurath Award 的 Professor Robert Sauer(來自 MIT)及 The Christian B. Anfinsen Award 的 Professor Carl Frieden(來自 Washington University)。
個人參加本次年會的內容,標題為”Assessment of Hydrogen Bonds between
Maltooligosyltrehalose Synthase and Maltopentaose by Computer Simulation and Site Directed
質結合時氫鍵的位置,並以實驗及酵素動力學的研究,與預測結果做確認,深感興趣。特別 是來自美國康乃爾的 Dr. Daniel Ripoll,由於其研究背景相似,對糖基的模擬所使用的力學力 塲參數,特別進一步深入討論與交換意見。由於糖基上的氧原子具有長電子對,其力學力塲 參數較之於核酸與蛋白質,來得複雜。日前並無適當可靠的力塲參數可與核酸與蛋白質同時 使用。因此可將目前的現有不同力塲參數做一比較,以決定何者較為與實驗結果相近。與本 次大會中,有一些利用力學模擬的研究,著重在於了解蛋白質摺疊機制並預測其摺疊結果,
以提供對蛋白質變性的參考。這些研究也提供了以後以力學模擬研究的其他方向。