國立臺灣大學生命科學學院動物學研究所 碩士論文
Graduate Institue of Zoology College of Life Science
National Taiwan University Master Thesis
探討 DjAgo2 蛋白在渦蟲再生中所扮演的角色 Study on the role of Argonaute protein DjAgo2
in planarian regeneration
張智凱 Chi-Kai Chang
指導教授:朱家瑩 博士
Advisor: Chia-Ying Chu, Ph.D.
中華民國 101 年 2 月
February, 2012
i
ii
致謝
從前總是覺得致謝這樣的文章,在畢業的時候一定是信手拈來,隨處是感謝的字 句。然而在經過了這段不知是長還是短的時間後,回首一盼,要感謝的人和事真 是多到不知該如何下筆。首先要感謝的當然是朱家瑩老師的細心指導,不論是實 驗還是做人處世上,老師常強調的邏輯性思考、辨明是非、待人有禮,都成為是 大家為人處世的標竿。還要感謝老師在實驗方向的修正、論文內容改正上的勞心 勞力,更在忍受了我不成熟的實驗技術、不足的闡述能力以及癟腳的英文寫作能 力之餘,仍能不厭其煩的給予指導及教導。感謝陳俊宏老師、李心予老師、詹世 鵬老師,在研究方向上給予意見,以及口試和論文修改上給予很大的幫助。感謝 羅竹芳老師、齊肖琪老師、于宏燦老師,在我剛進學校時給予的建議。謝謝台大 科技共同空間的莊以君學姊、楊亞臻學姊、高毓鄖學姊、蕭君儀學姊、阿湯哥,
沒有你們的大力協助和被煩卻又逃不掉的意氣,我的實驗無法順利進行。還有要
感謝李心予老師實驗室的Milky 學長、沛翊學長、易謙學姊、翠華學姊、傳恩、
耆豪、岳謙、予農,常去麻煩你們和借藥品。感謝羅竹芳老師實驗室的詩婷學姊、
怡婷學姊教我進行miRNA 的實驗。感謝陳俊宏老師實驗室的岱威、亞凡、政儀、
哈哈、尚樺、易澤,常去找你們詢問無脊椎動物的問題和實驗方法,當然也借了 很多小東西。感謝吳益群老師實驗室的學長姐贊助抑制基因表現的方法。感謝丘 臺生老師實驗室的志翔學長的大力協助,並且讓我們可以常常去你們那邊聊天。
最後要感謝實驗室的夥伴們,勁文學長的樂觀和勤勉是在遇到挫折時鼓舞著我,
讓我得到前進的動力。感謝本嵂學長、芷均、妍君、郁文、捥恕陪伴著我的實驗 室生涯,並且讓實驗室總是處於沸騰的狀態。還有大學部學弟妹,弘毅、翊柔、
若晞、方怡、意清,解決你們的問題有時卻能藉此整理自己的思緒和認識自己。
最後的最後,感謝家人的支持與諒解,這段時間雖然無法參加並幫忙處理家裡的 事物,但我心與你們同在,也感謝你們與我同在。
iii
摘要
渦蟲是一種具有再生能力的扁形動物,能由身體一部分片段再生出包含腦部的完 整個體。渦蟲的再生能力建立在一群多功能成體幹細胞(neoblast)身上,他們可以
進行複製以及分化來替換身體所有的組織,而且這群細胞對γ-射線很敏感。Dugesia
japonica(東洋渦蟲)的 DjAgo2 蛋白是人類 Ago2 的同源蛋白質,在人類細胞上的研
究顯示hsAgo2 是 miRNA 和 siRNA 路徑中的關鍵因子,具有調控基因表現的能力。
本研究中,我們首先選殖DjAgo2 之完整全長 cDNA 並構築與 YFP 蛋白接合之重
組蛋白表現載體,將其表現在 HeLa cell 中。在螢光顯微鏡下觀察發現,DjAgo2
會和人類的P-body 組成物 RCK 等聚集於細胞質內,這現象和 hsAgo2 的表現很類
似。藉由整體原位雜交的實驗,發現 Djago2 高度表現在渦蟲的頭部和背中線上。
渦蟲尾部再生的實驗中顯示在blastema 以及 post-blastema 兩區 Djago2 的表現量在
再生時會上升,而 RT-qPCR 的結果也顯示其表現量在再生第三天的 blastema 和
post-blastema 中,分別高出平常的五倍和兩倍。藉由 FACS 來分離 neoblast 和已分
化細胞,發現Djago2 在成體幹細胞中的表現量較高,而且 γ-射線會使得部分表現
Djago2 的細胞消失。最後,用 Djago2 dsRNA 來餵食渦蟲以抑制蟲體中 Djago2 的 表現,渦蟲會失去再生的能力而且會喪失組織恆定性。RT-qPCR 的結果顯示表現 在幹細胞 (DjpiwiA)以及合成 DNA (Djpcna) 的基因表現量在抑制 Djago2 後會下降。
而且,M phase 的細胞減少以及 G2 phase 的細胞增加顯示抑制 Djago2 會使得成體
幹細胞的細胞周期停留在G2 phase 而無法進行有絲分裂。同時,也觀察到部分渦
蟲出現分化受到限制的現象。我們的實驗結果顯示,Djago2 在渦蟲成體幹細胞的 細胞複製、自我更新以及分化時都被需要的,可以藉由對於成體幹細胞的調控來 影響渦蟲的再生能力。
關鍵字:
渦蟲、再生、成體幹細胞、微核醣核酸
iv
Abstract
Planarians are capable of regenerating almost every part of their body, including the brain. Regeneration in planarians is mediated by a group of pluripotent adult somatic stem cells, called neoblasts, which can proliferate and differentiate to replace all tissues.
These cells are sensitive to γ-irradiation. DjAgo2 of Dugesia japonica is a homolog of hsAgo2, the key factor that regulates gene expression in miRNA and siRNA pathways.
In the study, DjAgo2 was cloned, and YFP-tagged DjAgo2 was expressed in HeLa cells.
DjAgo2 accumulates at specific foci that contain the P-body component, RCK. This pattern is similar to what we observed with hsAgo2. By whole-mount in situ hybridization (WISH), Djago2 was found to be highly expressed in regions of the brain and the middle dorsal line. Djago2 expression was increased in the blastema and post-blastema during regeneration. RT-qPCR results confirmed that respective Djago2 mRNA expression levels in the blastema and post-blastema were 5 and 2-times higher than that in the control at 3 days post-amputation. WISH data showed that a portion of Djago2-positive cells were depleted by γ-irradiation. We next examined Djago2 expression levels in neoblasts isolated by FACS. Our RT-qPCR data showed that the Djago2 level was higher in neoblasts than that in differentiated cells. Finally, Djago2 was silenced by feeding planarians Djago2 dsRNA. Depletion of Djago2 resulted in loss of the regeneration capacity and defects in tissue homeostasis in planarians. Djago2 silencing reduced levels of DjpiwiA, Djpcna, and DjpiwiC. Our data also showed that the ratio of M-phase cells was decreased and the ratio of G2-phase cells was increased in Djago2-silenced animals. This suggests that Djago2 silencing blocked the mitosis of neoblasts. Differentiation restriction was also observed in Djago2-silenced animals. Our data indicate that Djago2 is required for neoblast proliferation, and differentiation.
v
Keyword:
Planarian, regeneration, neoblast, Ago2, miRNA
vi
Contents
口試委員審定書………i
Acknowledgement………ii
Abtract (Chinese) ………iii
Abstract (English) ………iv
1. Introduction………..1
1.1 Planarians and regeneration………1
1.2 Blastema and neoblasts……….…………..………3
1.3 Small RNAs in planarian…………..………9
1.4 Argonaute proteins………..………13
2. Materials and Methods………..……16
2.1 Animals………16
2.2 5’ Rapid amplification of complementary (c)DNA ends (RACE) and 3’RACE ………16
2.3 Expression vectors………17
2.4 Cell culture and transfection………17
2.5 Immunofluorescence………18
2.6 Whole mount in situ hybridization………18
2.7 Preparation of single-cell suspensions for fluorescence-activated cell sorting analysis………19
2.8 Quantitative PCR……….…………20
2.9 RNA interference (RNAi) experiments ……….…………21
vii
2.10 Anti-phospho-histone H3 labeling……….………….…………22
3. Results……….24
3.1.1 Cloning of planarian Argonaute pretein-DjAgo2………24
3.1.2 The subcellular distribution of DjAgo2 is similar to that of human AGO2 localizing to P-body……….…………24
3.2.1 Expression pattern and level of Djago2 in planarian………25
3.2.2 Djago2 is highly expressed in regenerating region………26
3.3.1 γ-irradiation eliminates the Djago2-positive cells……….28
3.3.2 Djago2 is highly expressed in neoblasts ……….………30
3.4.1 Failure to regenerate in Djago-silenced animals……….…31
3.4.2 Tissue homeostasis defect was observed in long-term Djago2 RNAi treated animals……….……….………..33
4. Discussion………..35
4.1 DjAgo2 is a member of the Argonaute subfamily……….…35
4.2 Djago2 is up regulated in regenerating tissue during planarian regeneration….36 4.3 Expression level of Djago2 is higher in neoblasts than in differentiated cells…37 4.4 Djago2 is required for proliferation of neoblasts and differentiation of progenitor cells ……….……….………..38
5. References……….……….………42
6. Figures……….……….………..55
1
1. Introduction
1.1 Planarians and regeneration
Planarians are bilaterally symmetric metazoans in the class Turbellaria of the phylum Platyhelminthes. Planarians are small unsegmented flatworms that are not parasitic. They are commonly found in freshwater ponds or streams, and move by cilia on the ventral dermis. Planarians are triploblastic and acoelomate, which means they have three embryonic tissue layers, including an ectoderm, mesoderm, and endoderm, without a body cavity. While planarians have no specific circulatory, respiratory, or skeletal structures, they do have a digestive system with a pharynx and a blind gut, but no anus (Salo and Baguna, 2002). Planarians have a simple visual system on their heads.
The visual system includes a pair of eyespots with rhabdomeres and pigment cells (Sakai et al., 2000). The photoreceptors can sense light intensity and directionality but form no images (Sakai et al., 2000).
The regeneration ability of planarians is remarkable. A small amputated fragment from any part of their body can regenerate to form a complete worm (Morgen, 1901).
Studies on planarian regeneration were initiated several hundred years ago. Peter Simon Pallas was the first person to describe how a small piece of amputated planarian head
2
has the capability to regenerate to an intact body in 1766 (Brøndsted, 1969).
Experiments on planarian regeneration were performed by Dalyell and Johnson in the early nineteenth century. According to their results, Dalyell concluded that planarians may “almost be called immortal under the edge of the knife” (Brøndsted, 1969). After that, Harriet Randolph defined the minimal size of a fragment capable of regeneration as 1/279 of an intact worms (Randolph, 1897).
Thomas Hunt Morgan was the first person to systematically study planarian regeneration (Morgen, 1901). He described the regenerative ability and polarity of planarian regeneration. Although many animals can regenerate missing tails, arms, or legs, the planarian is one of the few animals that can even regenerate a head. In planarians, a new head is formed in 7 days after transverse amputation of the region posterior to the pharynx. New eyes are formed in 4 days of head regeneration. Therefore, it is a valuable model system for elucidating mechanisms that control cell and tissue replacement, a process that is important for the survival of most organisms (Slack, 2011).
Various species of planarians were characterized in detail as having either asexual or sexual strains. Some of them are commonly used to study regenerative phenomena in biological laboratories. For example, Dugesia ryukyuensis has been shown to exhibit the
3
capability of switching from asexual to sexual reproduction (Hoshi et al., 2003). The freshwater hermaphrodite, Schmidtea mediterranea, was well characterized by Alvarado's group in the past 10 years, and has emerged as a model system. In this study, I use Dugesia japonica as a model planarian. It is commonly distributed in the Far East, including Japan, Taiwan, Korea, China, and Primorskiy in Russia (Tamura et al., 1998).
In addition, D. japonica is the only species of freshwater planarian in Taiwan. The karyotype of D. japonica can be diploid, triploid, or mixoploid (Tamura et al., 1998).
For many research groups in Asia, D. japonica has become a suitable model system for studying the regeneration and developmental processes at the cellular and molecular levels.
1.2 Blastema and neoblasts
During planarian regeneration, lost parts of the body are rebuilt from the remaining tissues. The process of regeneration includes at least two events: blastema formation and pattern formation. The blastema is the pigment-free bud formed at a wound site after amputation. After blastema formation, cells in the newly grown tissues then differentiate into different cell types according to the developmental signals that determine the body's polarity. T.H. Morgan, who was the first researcher to perform
4
many experiments to understand how polarity is reestablished after amputation (Morgen, 1901). Recent studies showed that some signaling pathways regulate the dorsal-ventral axis and anterior-posterior polarity during regeneration. For example, SmedBmp4-1 molecules in the BMP signaling pathway are expressed in the midline region of intact animals, and regulate dorsal-ventral patterning (Reddien et al., 2007). SmedWntP-1, expressed in a few cells in the tail of intact animals, regulates anterior-posterior (AP) polarity during regeneration (Petersen and Reddien, 2009). Knockdown of the gene expression of β-catenin leads to ectopic heads at wound sites in all body regions (Adell et al., 2009; Chai et al., 2010; Gurley et al., 2008; Petersen and Reddien, 2008).
(Gentile et al., 2011) Source cells of the blastema were investigated for decades (Baguna and Romero, 1981; Slack, 1980). It is now accepted that the blastema is formed by proliferation of pluripotent stem cells, called neoblasts (Baguna et al., 1989). Neoblasts are small cells
5
of 5~10 μm. Neoblasts has a large nucleus surrounded with a thin cytoplasmic layer.
Many studies indicated that the blastema is not the proliferation site of neoblasts in planarians. During regeneration, cell division occurs in the region next to the blastema, called the post-blastema. It seems that damaged nerve fibers may release regulatory molecules into the extracellular space to regulate the process of regeneration(Salo and Baguna, 1984, 1989), even though no molecular evidence was shown. Cell proliferation of neoblasts is activated during regeneration. An early mitotic peak from slowly dividing cells with a long G2 phase, is observed at 8~12 h after amputation. The early mitotic peak reflects the existence of parts of neoblasts in the G2 phase, which await an appropriate signal to divide. The second mitotic peak occurs 2~4 days after amputation.
After cell proliferation, neoblasts migrate to the blastema and differentiate into various cell types. This process of restoring the amputated body part to an intact animal is completed within 2~3 weeks (Baguna et al., 1989).
Neoblasts fit the classical definition of a stem cell. They are small undifferentiated cells and are required for normal growth, self-renewal, and regeneration in planarians (Eisenhoffer et al., 2008). Neoblasts are localized in the parenchyma which lies beneath the muscle and surrounds the organs. Neoblasts comprise about 20%~30% of planarian cells, but only a small percentage of them have the capability for mitotic division
6
(Baguna and Romero, 1981; Baguna et al., 1989). Some of them are the immediate division progeny of neoblasts (Eisenhoffer et al., 2008). Neoblasts are described as adult stem cells, but they are unlike adult stem cells of mammals. In mammals, a tissue contains a small number of tissue-specific stem cells through which it can continuously renew itself. Fetal and adult stem cells are usually multipotent and can differentiate into limited cell types. However, the neoblasts of planarians are pluripotent stem cells that can renew all types of differentiated cells, whereas only mammalian embryonic stem cells exhibit pluripotency.
X-ray irradiation was used to inhibit planarians’ regenerative abilities by eliminating stem cells with mitotic ability (Bardeen and Baetjer, 1904). Wolff and Dubois performed serial experiments, with various portions of an animal shielded from irradiation (Wolff 1962). If the anterior region of an intact animal was irradiated and the region was amputated, the irradiated region became necrotic and died. The un-irradiated fragment of the same animal survived. If the anterior region of an intact animal was irradiated and the head was removed, the irradiated region was repaired and blastema was formed with a regeneration delay. This regeneration delay reflects neoblasts having to migrate through the irradiated region to the wound site. Therefore, planarians are like a culture dish. Although X-ray-irradiated planarians lose their regenerative ability,
7
transplantation of a neoblast-enriched cell population into irradiated animals can restore their regenerative ability (Baguna et al., 1989; Wagner et al., 2011). It indicates the regenerative ability of planarians depends on the neoblasts.
Planarians with developmental plasticity have the capability to grow and ‘de-grow’
depending on the nutritional status (Baguna and Romero, 1981; Lillie, 1900). A full-grown adult (~20 mm long) can shrink to a size that is much smaller (~1 mm) after starvation for months (Newmark and Sanchez Alvarado, 2002). Growth and de-growth in planarians occur due to changes in cell numbers (Baguna and Romero, 1981), not from changes in cell size. It is thought that the ratio of cells born by proliferation to cells lost by apoptosis changes during starvation. The phenotype of irradiated animals is similar to that of starved animals, which indicates that the activity of neoblasts is important for tissue homeostasis.
The mechanism of blastema formation in planarians differs from that of vertebrates.
First, wound epithelium is formed by epidermal cell shape modifications in planarians, but it forms by cell proliferation in vertebrates. Second, mesenchymal cells are from preexisting neoblasts in planarians, but are from dedifferentiated cells in vertebrates (Newmark 2001). Parenchyma cells accumulate as neoblasts in the blastema (Betchaku, 1967, 1970), and the cytoplasmic processes of fixed parenchyma cell are related to
8
directional movements of regenerative cells by providing a contact guidance system (Hori, 1991). This indicates that differentiated cells regulate the behavior of adult stem cells by an extracellular substance.
For regeneration research, biologists tried to label these cells. Thymidine and 5’-bromo-2’-deoxyuridine (BrdU) are used to label cycling cells. Serine 10 of histone H3 becomes phosphorylated throughout condensing chromatin at the initiation of mitosis. Anti-H3P is used to label cells in the M phase (Hendzel et al., 1997). Recently, increasing numbers of genes were found in neoblasts. Djmcm2 is a member of the minichromosome maintenance (MCM) family of DNA replication factors (Salvetti et al., 2000). DjPCNA is a component of the DNA replication protein complex (Orii et al., 2005). DjRbAp48 is a component of different chromatin-modeling complexes (Bonuccelli et al., 2010). All three of those genes are specifically expressed in neoblasts and strongly expressed in the dorsal midline. Djvas-1, Djpabpc-1, and DjpiwiA are components of RNA granules, and are expressed in neoblasts (Rouhana et al., 2010).
Neoblasts can also be detected and isolated by fluorescence activated cell sorting (FACS) (Hayashi et al., 2006).
Several genes are known to be required for neoblast maintenance, e.g., Smedwi-2 (Reddien et al., 2005), Smedwi-3 (Palakodeti et al., 2008), Smed-bruli (Guo et al., 2006),
9
Dj-pum (Salvetti et al., 2005), Spoltud-1 (Solana et al., 2009), Smed-SmB
(Fernandez-Taboada et al., 2010), Smedinx-11 (Oviedo and Levin, 2007), Djmot (Conte et al., 2009), and Smed-CHD4 (Scimone et al., 2010). Several genes are found in progenitor cells, e.g., Smed-AGAT1, Smed-NB21.11e (Eisenhoffer et al., 2008), and Smed-p53 (Pearson and Sanchez Alvarado, 2010). Silencing of these gene blocks the
regenerative ability in planarians. According to those studies, we know that planarian regeneration depends on the presence and precise regulation of neoblasts, which differentiate to replace cells of any missing tissue (Rouhana et al., 2010). After over 200 years of studies, the mechanism of planarian regeneration is still unclear. But there are many markers to monitor the stages of neoblasts for research on planarian regeneration.
1.3 Small RNAs in planarian
In 1998, experimental introduction of RNA into cells can be used in certain biological systems to interfere with the function of an endogenous gene. In recent years, small, non-coding RNAs were shown to be essential players in almost all biological processes. Small RNAs (siRNA, miRNA, piRNA, tncRNA, snRNA) have the ability to regulate gene expressions in animal development, regeneration, stress response, and transposon silencing. Many small-RNA species have been identified. But most of the
10
biological functions of these species are still unclear. The miRNAs are endogenous RNAs with ~22 nucleotides that can play important regulatory roles in animals and plants by targeting mRNAs for cleavage or translational repression (Bartel, 2004).
Recently, functions of miRNAs were identified to be involved in regulation of cell cycle, cell proliferation, cell death, apoptosis, and fat metabolism in Drosophila (Brennecke et al., 2003), neuronal patterning in nematodes (Johnston and Hobert, 2003), and development of leaf and flower in plants (Aukerman and Sakai, 2003; Palatnik et al., 2003). miRNAs are also important to stem cells. Some miRNAs are specifically expressed in human and mouse embryonic stem cells (Houbaviy et al., 2003; Suh et al., 2004), and miRNAs have the ability to regulate the differentiation of hematopoietic stem cells in mice (Chen et al., 2004) and germline stem cells in Drosophila (Hatfield et al., 2005). Previous studies suggested that 90% of genes may be regulated by miRNAs in humans (Miranda et al., 2006).
The mature miRNAs are produced from precursor transcripts called primary miRNAs by RNA polymerase II (Cai et al., 2004; Lee et al., 2002). The pri-miRNA is processed into a 60~70-nucleotide pre-miRNA by Drosha in the nucleus (Lee et al., 2003). Pre-miRNAs have a two-nucleotide overhang at their 3' ends and a 5' phosphate group. Pre-miRNA is cleaved by Dicer to generate a duplex of miRNA and miRNA*
11
(Schwarz et al., 2003). Dicer is component of the RISC loading complex. RISC loading complex recruits Ago2 and transfers the miRNA duplex. Ago2 can then cleave the target RNA or repress translation (Kim et al., 2007; Liu et al., 2004). Most miRNAs pair with their targets through the sequence at the 5' end from the second to the eighth called the
“seed region” (Brennecke et al., 2005; Lewis et al., 2005). A single miRNA therefore can regulate many different mRNAs, whose sequences are complementary to seed region in miRNA (Lewis et al., 2003).
(Farrar et al., 2011) The piRNAs are the longest small RNAs with 25~32 nucleotides and they function in the germ line. Unlike siRNAs, piRNAs are mainly antisense. This suggests that piRNAs are produced from long, single-stranded precursor RNAs. Biogenesis of piRNA
12
is from sequences of piRNAs that are bound to Piwi, Aubergine and Ago3 (Brennecke et al., 2007). The biogenesis of piRNA occurs through an amplification mechanism of
“ping-pong” that is activated after transcription of transposon mRNA (Tabara et al., 2002). In planarians, miRNAs and piRNAs are predominant small RNAs (Friedlander et al., 2009).
Dasaradhi Palakodeti et al was the first group to study miRNAs in planarian. They reported the cloning and initial characterization of 71 miRNAs from the planarian Schmidtea mediterranea (Palakodeti et al., 2006). Marc R. Friedländer et al used
massive, next-generation sequencing methods to compare profile of small RNAs present in neoblasts, irradiated animals, and normal animals (Friedlander et al., 2009). The read numbers of piRNAs were predominant in sorted neoblasts (82%), and low in the irradiated sample (25%) (Friedlander et al., 2009). In contrast, miRNAs were predominant in the irradiated sample, but several miRNAs were enriched in neoblasts.
They used a reverse transcription quantitative polymerase chain reaction (RT-qPCR) to confirm the deep sequencing data. Ten miRNAs were up-regulated in neoblasts (let-7a, let-7b, miR-36b, miR-2a, miR-2d, miR-13, miR-71b, miR-756, and miR-2160).
Yi-Chien Lu et al used the same method to define the full complement of small RNAs present in irradiated animals, un-irradiated animals, a sexual strain, and an asexual
13
strain. Five miRNAs were identified that were reduced by at least 2-fold in irradiated animals (let-7a, mir-71b, mir-756, mir-13, and mir-752) (Lu et al., 2009). Qing-Nan Tian et al identified 8 miRNAs that were differentially expressed in regenerative tissues using an miRNA microarray (miR-71a, miR-756, miR-13, miR-281, miR-79, miR-2d-3p, miR-36, and miR-2b) (Tian et al., 2011). Cristina González-Estévez et al showed that 12 miRNAs were sensitive to irradiation by small RNA whole-mount in situ hybridization (WISH) (let-7c, mir-2c, mir-71c, mir-67, mir-2d, mir-71b, mir-752,
mir-7b, mir-36, mir-92, mir-124a, and mir-124b) (Tian et al., 2011). Those results indicated that miRNAs might play important roles in regeneration or neoblast maintenance and differentiation.
1.4 Argonaute proteins
The Argonaute protein family was first identified in plants, and were later found in bacteria, archaea, and eukaryotes (Cerutti and Casas-Mollano, 2006) . They are highly conserved among species, and many organisms encode multiple members of the family (Hock and Meister, 2008). Members of this family are defined by having a Piwi-Argonaute-Zwille (PAZ) domain and a P-element induced wimpy testis (PIWI) domain (Bohmert et al., 1998). Studies on Ago2 structure have shown that the PAZ
14
domain has a subdomain which displays oligonucleotide binding (OB)-like folding. It also showed that the PAZ domain can recognize 3' ends of single-stranded RNAs (Yan et al., 2003). The Mid domain is between the PAZ and PIWI domains and has cap-binding activity. The PIWI domain binds to the 5' end of siRNA and the target RNA.
The PIWI domain contains catalytic residues (DDH) with the capacity (Song et al., 2004) to cleave the mRNA strand between nucleotides 10 and 11 if the complementarity of the miRNA and mRNA is perfect (Yuan et al., 2005).
The Argonaute protein family has two subfamilies, the Argonaute and PIWI. The Argonaute subfamily is similar to Arabidopsis thaliana Ago1, while the PIWI subfamily is similar to Drosophila melanogaster PIWI (Carmell et al., 2002). Argonaute subfamily proteins are expressed and bind to siRNAs or miRNAs for post-transcriptional gene silencing either by translational repression or by destabilization of the mRNA. PIWI proteins are involved in piRNA pathway. The mechanism of PIWI proteins is not clear.
In planarians, several PIWI proteins were identified. Smedwi-2 and Smedwi-3 have the ability to regulate neoblast proliferation (Palakodeti et al., 2008). Drpiwi-1 is essential for germline cell formation during sexualization of the planarian Dugesia ryukyuensis (Nakagawa et al., 2012). The Argonaute protein was identified in planarians. Djago2 silencing results in a failure to regenerate (Rouhana et al., 2010). However, the role of
15
Djago2 in planarian regeneration is still unclear.
16
2. Materials and Methods
2.1 Animals
The planarian D. japonica was captured in Jiaoxi Township, Ilan County, of northeastern Taiwan. The worms were maintained in aerated tap water at 18 °C for at least a month and fed chicken liver once a week. Replacement of the aerated tap water three times a week was used to control water quality. In this study, worms were used after 1 week of starvation.
2.2 5’ Rapid amplification of complementary (c)DNA ends (RACE) and 3’RACE
Total RNA was collected from intact planarians with the TRIzol reagent. 5' RACE-PCR began using mRNA as a template for a first round of cDNA synthesis with the superscript III First-strand Synthesis System (Invitrogen). An anti-sense gene-specific primer was used to recognizes Djago2 mRNA template in the 3' to 5' direction to generate a specific single-stranded cDNA product. Following cDNA synthesis, the enzyme terminal deoxynucleotidyl transferase (TdT), was used to add a string of deoxycytidine triphosphates (dCTPs) to the 3' end of the cDNA. A PCR reaction was performed using a secondary antisense gene-specific primer and an oligo
17
(dG) primer. The products of the PCR were cloned into a TA vector and sequenced. The 3' RACE-PCR used an oligo (dT) primer to generate cDNA. The PCR was then used to amplify 3' cDNA using a sense gene-specific primer and an oligo (dT) primer. The products of the PCR were cloned into the TA vector and sequenced. The first antisense gene-specific primer was GCATAATAAGCAGGAGCTGG, the secondary antisense gene-specific primer was CCGTCAAACACAGGTCTCTG, and the sense gene-specific primer was GGTGTTTGGGATATGAGAGG.
2.3 Expression vectors
Expression vectors with N-terminal YFP-tag was generated by PCR amplification of Expression vectors with an N-terminal yellow fluorescent protein (YFP) tag was generated by PCR amplification of Djago2 coding sequences from cDNA of intact planarians, followed by cloning into the XhoI and BamHI sites of pEYFP-C1 (BD).
2.4 Cell culture and transfection
HeLa cells were cultured in Dulbecco's minimal essential medium (DMEM) with 10% fetal bovine serum (FBS) at 37 °C with 5% CO2. Cells were transfected using Lipofectamine (Invitrogen) according to the manufacturer's protocol.
18
2.5 Immunofluorescence
Cells transfected with YFP-Djago2 were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) at room temperature for 20 min and permeabilized for 5 min with 0.25% (v/v) Triton X-100. Samples were washed three times with PBST (0.1% [v/v] Triton X-100 in PBS) and blocked for 30 min in PBST containing 2% (w/v) bovine serum albumin (BSA). Primary and secondary antibodies were diluted in blocking solution during incubation. Antibodies used in these experiments included anti-RCK/p54 and anti-YFP. Secondary antibodies against rabbit and mouse immunoglobulin G (IgG) were directly conjugated to Alexa Fluor dyes (Molecular Probes, Eugene, OR). Samples were washed three times with PBST after antibody incubation. After the final wash, samples were counterstained with Hoechst 33342 to visualize nuclei.
2.6 Whole mount In situ hybridizations
Animals 5 to 8 mm long were used for this experiment. Planarians were killed, and mucus was removed with 5% N-acetyl cysteine in 5/8 Holtfreter's solution for 5 min on slides, and then specimens were fixed in 4% formaldehyde for 20 min. Worms were
19
transferred into 1.5ml Eppendorf-tubes. Fixed samples were washed with 50% methanol and 100% methanol, and bleached overnight in methanol with 6% H2O2. Animals were washed with a serial dilution of methanol: PBST (0.3% triton X-100 in PBS). Animals were treated with 2 μg/ml of a proteinase K solution. Worms were then incubated at 56
°C for 2 h in pre-hybridization solution (50% formamide, 5× SSC, 1% Tween-20, and 1 mg/ml yeast torula RNA). Hybridization was performed at 65 °C in hybridization solution containing 50% formamide, 10% dextran sulfate, 5× SSC, 1% Tween-20, and 1 mg/ml yeast torula RNA with a DIG-labeled antisense RNA probe for 16 h. Animals were washed through a serial dilution into 2x SSC, then 0.2x SSC, and finally into MABT (100 mM maleic acid, 150 mM NaCl, and 0.1% Tween-20; pH 7.5). Animals were incubated with blocking solution (10% goat serum in MABT) for 2 h, and then with primary antibodies (1:4000 anti-digoxigenin-AP (Roche) diluted in blocking solution) overnight. Animals were washed 6 times with MABT, and the signal was developed with an NBT/BCIP mixture.
2.7 Preparation of single-cell suspensions for the fluorescence-activated cell sorting
(FACS) analysis
Planarians were cut into three fragments on ice, and rinsed with a calcium- and
20
magnesium-free solution (CMF: 2.56 mM NaH2PO4·H2O, 10.21 mM KCl, 14.28 mM NaCl, 9.42 mM NaHCO3, and 1% BSA). The fragments were cut into smaller pieces and treated with 0.25% (w/v) trypsin in CMF for several minutes at room temperature.
Samples were completely dissociated into single cells by gentle pipetting and then filtered through a 70-µm-pore size cell strainer (BD) and a 20-µm nylon net filter (Millipore) to remove tissue fragments. The single-cell suspensions were incubated with 13.5 µg/mL Hoechst 33342 (Sigma) and 0.5 µg/mL calcein-AM (Sigma) for 1 h at room temperature. Samples were pelleted by centrifugation at 1500 g for 2 min, and resuspended in CMF. A flow cytometric analysis used a FACSAria III cell sorter (BD).
2.8 Quantitative PCR
Total RNA was extracted using TRIzol, and cDNA was synthesized with the superscript III First-strand Synthesis System (Invitrogen). A real-time RT-PCR was performed using SYBR® FAST qPCR Kits (KAPA). Djactin was used for normalization. The following primers were used in the amplification reaction:
Djago2 forward, 5’-CCTGTAATATTTCTCGGTGCTGA-3’;
Djago2 reverse, 5’-CATCCATACTGCCTACAACAGC-3’;
Djactin forward, 5’-GGTAATGAACGATTTAGATGTCCAGAAG-3’;
21
Djactin reverse, 5’-TCTGCATACGATCAGCAATACCTGGAT-3’;
DjpiwiA forward, 5’-GGAGCCATAGGAGAAATCTCATTTG-3’;
DjpiwiA reverse, 5’-CGCTAATCCAAATCCGGGAAC-3’;
DjPCNA forward, 5-TGAGGCTATCACCATTACTGTTG-3’;
DjPCNA reverse, 5’-GGCTCGGTCATTTCAATAGTG-3’;
DjpiwiC forward, 5-GGACGTGACTACTTCTACCCAG-3’;
DjpiwiC reverse, 5’-CTTTGTGTGAAACATCGCACTG-3’
2.9 RNA interference (RNAi) experiments
Double-stranded (ds)RNA was used to knock down Djago2 expression (Sanchez Alvarado and Newmark, 1999). The Djago2 dsRNA sequence was cloned by a PCR.
The following primers were used: forward primer, GCCTAATACGAGACACTAT- AGAAGGTGTTTGGGATATGAGAGG; and reverse primer, GCCTAATACGAC- TCACTATAGGGTAACAATGCCGAAATTTGAT. These primers had a T7 promoter sequence. Djago2 dsRNA was synthesized with T7 polymerase for 3 h. After RNA synthesis, the sample was digested with DNase I for 15 min at 37 °C. Djago2 dsRNA was precipitated with ethanol, and resuspended in 20 μl of nuclease-free H2O.
Formation of dsRNA was confirmed by running 200 ng of these reactions in a 1.0%
22
agarose gel in TAE. This artificial food was mixed with 4 µg dsRNA in 25 μl of a 75%
chicken liver mix, 6 μl 2% ultra-low gelling agarose, and 1 μl red food color. The artificial food mix was allowed to solidify at 4 °C, and room-temperature food was fed to planarians. Worms were fed with RNAi food on days 1, 4, and 7. After the 3rd RNAi feeding, the heads and tails were immediately removed. After 9 days of regeneration, animals were fed and amputated again. The phenotype was monitored by a dissecting microscope.
2.10 Anti-phospho-histone H3 labeling
To detect mitotic cells, a rabbit anti-phospho histone H3 antibody (Millipore) was used to detect mitotic cells. Planarians were killed by ice-cold 2% HCl (0.5~1 min) and then fixed in Carnoy’s solution for 2.5~3 h at room temperature. After rinsing in MeOH, fixed specimens were bleached overnight in 6% H2O2 in MeOH at room temperature.
Planarians were either stored at -20 °C in MeOH or immediately rehydrated through an MeOH/PBSTx (0.3% Triton X-100 in PBS) series (75%, 50%, and 25% MeOH).
Samples were blocked and incubated overnight at room temperature in anti-H3P diluted 1:1000 in PBSTx + 10% goat serum. After multiple PBSTx washes (for at least 8 h), samples were post-fixed for 15 min at room temperature in 4% paraformaldehyde in
23
PBS and washed twice in PBSTx. Samples were incubated overnight at 4 °C in anti-rabbit Alexa 488 (1:1000) diluted 1:1000 in PBTx + 10% goat serum. Following three 10-min washes in PBSTx, samples were stained with Hoechst 33342 for 30 min.
24
3. Results
3.1.1 Cloning of Planarian Argonaute - Djago2.
The full length sequence of Djago2 was completed by 5’RACE and 3’RACE. The clone with 2787 nucleotides was isolated and sequenced (Fig.1B). The initial methionine was identified by comparison with the sequences of AGO2 ortholog from other organisms. The Djago2 encodes a protein composed of 929 amino acids.
Phylogenetic analysis showed DjAgo2 is a member of AGO subfamily and is closely related to Schmidtea mediterranea Ago2, Drosophila melanogaster Ago1 and human Ago2 (Fig. 1A). DjAgo2 contains conserved PAZ, Mid and PIWI domains. Sequence alignment of DjAgo2 with its ortholog in humans, hsAgo2, showed that is exhibited high similarity to human Ago2 with 76.5% identity, especially in the PIWI domain (88%) (Fig.1B). DjAgo2 also contains DDH, which are key residues for the catalytic activity of Ago2.
3.1.2 The subcellular distribution of DjAgo2 is similar to that of human Ago2
localized to P-bodies.
To characterize if the cloned DjAgo2 indeed exhibits similar function to its human
25
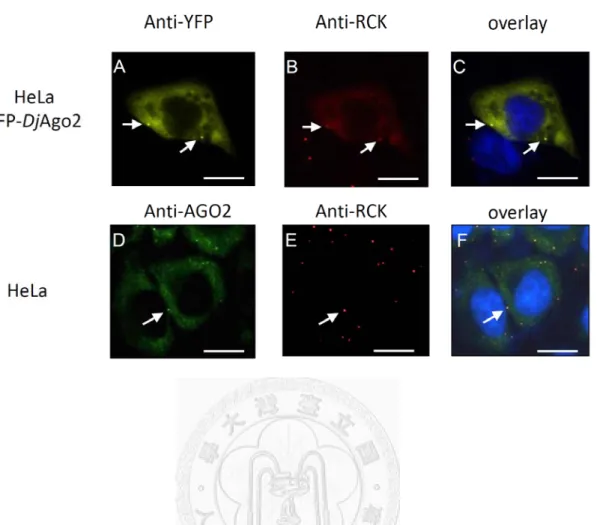
orthologue, I first examined localization of DjAgo2 in HeLa cells. Previous studies showed that Ago2 is localized to cytoplasmic mRNA P-bodies, and facilitates miRNA-induced gene silencing. Therefore, I hypothesized that if DjAgo2 is the functional planarian Argonaute protein, it should also localize to P-bodies. YFP-tagged DjAgo2 was expressed in HeLa cells, and its localization was confirmed by the human
P-body marker, RCK/p54 (Fig 2). YFP-DjAgo2 was expressed in cytoplasm, and accumulated at specific foci. RCK and YFP antibodies were used to label endogenous RCK and YFP-DjAgo2. HeLa cells which expressed YFP-DjAgo2 were co-stained with anti-YFP and anti-RCK. YFP-DjAgo2 co-localized with endogenous RCK (Fig. 2, panels a, b, c). This expression pattern is similar to that of human Ago2 (Fig 2, panels d, e, f)
3.2.1 Expression pattern and level of Djago2 in planarian.
To examine the expression of Djago2 in intact planarians, I used WISH to detect the mRNA of Djago2. WISH for Djago2 was performed with a digoxigenin-labeled antisense RNA probe. The RNA probe with ~1500 nucleotides in sequence length included the 3’ UTR and a partial sequence of the PIWI domain. Animals that were 5~8 mm long were used for this experiment. A sense RNA probe was used as the control,
26
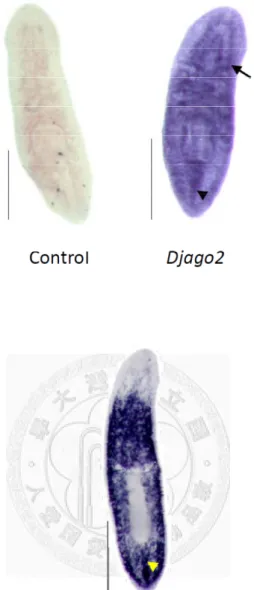
and no signal was detected after hybridization (Fig. 3A). This showed that the background signal was undetectable in our WISH system. The expression pattern of Djago2 was detected in the whole body and was readily detected in central nervous
system (CNS) and middle dorsal line. The expression pattern of Djago2 in intact planarians was similar to Djcbc-1 (Dμgesia japonica chromatoid body component 1) (Yoshida-Kashikawa et al., 2007). Djcbc-1 is a homolog of human RCK/p54. In planarians, Djcbc-1 is located in chromatoid bodies of neoblasts and chromatoid body-like structures in the CNS (data not shown). DjpiwiA is known to be a neoblast-specific gene, which is expressed throughout the planarian body. It is also expressed in cell clusters in the middle dorsal line (Fig. 3B).
3.2.2 Djago2 is highly expressed in the regenerating region.
I next examined the expression of Djago2 in regenerating animals. After amputation anterior and posterior to the pharyngeal region, the head, trunk, and tail were allowed to simultaneously regenerate. Expression patterns of Djago2 in the head, trunk, and tail were observed on day 3 after transverse amputation. Djago2 was expressed in cells of the CNS, middle dorsal line, and regenerating tissues (Fig. 4A).
This showed that Djago2 might be expressed in neoblasts. To observe Djago2 levels at
27
wound sites during planarian regeneration, worms were transversely amputated posterior to the pharynx, harvested and analyzed by WISH. Animals were harvested at various time points of 8 h, and 1, 2, 3, 5, and 7 days after amputation. Expression of Djago2 was increased in the regenerating region during regeneration until 3 days, and
the signal was lower on day 5 and 7 (Fig. 4B).
The expression levels of Djago2 at various regeneration time points were quantified by RT-qPCR. During planarian regeneration, the regenerating tissue, called the blastema, contains a mass of cells which is capable of replacing lost or damaged body parts. In previous studies, DjpiwiA-labeled neoblasts proliferated in the region of the post-blastema and then migrated to and differentiated into various cell types in the blastema (Umesono et al., 2011). Quantitative analysis of Djago2 was performed for tissues of the blastema and post-blastema during regeneration. Blastemas were harvested at various time points of 3, 5, and 7 days after transverse amputation at the post-pharyngeal region. Post-blastema samples of three more time points at 8, 24, 48 h post-amputation were harvested in the regenerating region. Expression levels of Djago2 were not examined in the blastema at 3, 5, or 7 days post-amputation because the newly formed tissues were too small. Control samples of the blastema and post-blastema were the tail and the region next to the first amputation, respectively. In total, 2 µg of RNA in
28
each sample was collected for reverse transcription to produce cDNA and was subjected to a qPCR analysis. In the post-blastema, expression of Djago2 was increased during regeneration, and Djago2 was respectively 2.5- and 3.2-fold higher than the control at 8 and 72 h post-amputation (Fig. 5A). In the blastema, expression of Djago2 was 6-fold higher than the control at 72 h post-amputation (Fig. 5B). Our results showed that the expression of Djago2 was increased in both the blastema and post-blastema during regeneration. Then I examined expression levels of DjpiwiA, a marker for neoblasts, in the blastema at 3, 5, and 7 days post-amputation. Expression of DjpiwiA did not rise up at 72 h after the first amputation (Fig. 5C). On days 5 and 7 post-amputation, expression levels of DjpiwiA were lower than the control. This confirmed that neoblasts had accumulated in the post-blastema but not in the blastema during regeneration.
3.3.1 γ-irradiation eliminates the Djago2-positive cells.
Since the increasing expression of Djago2 in regenerating tissues was synchronized with that of DjpiwiA, I hypothesized that Djago2 is expressed in neoblasts, and Djago2 expression is higher as a result of proliferation and differentiation of neoblasts. To check if Djago2 is expressed in neoblasts, I examined expression levels of Djago2 in both normal and neoblast-depleted animals. I depleted neoblasts in planarians by 90 Gy
29
γ-irradiation. Fourteen days after γ-irradiation, all animals displayed degeneration (Fig.
6A). All animals were dead at 3~4 weeks after irradiation, indicating a failure of tissue homeostasis. I used FACS to confirm if γ-irradiation eliminated neoblasts (Fig. 6B).
Single-cell suspensions were collected and filtered from dissociated animals. Staining with Hoechst 33342 reflects the DNA content in nuclei. The cytoplasm of living cells was stained with vital calcein-AM. In FACS, I first selected a population of particles which contained Hoechst 33342/calcein-AM double-positive cells. I separated those cells into three populations according to the DNA content and cell size. G2/M- (Fig 6B, p3) and G1/G0-phase cells (Fig. 6B, p4 and p5) could be distinguished by the Hoechst 33342 fluorescence signal. Our data showed that G2/M-phase cells (p3 in Fig 6B), which are highly proliferating neoblasts, had been eliminated at day 3 post γ-irradiation (Fig. 6B).
RT-qPCR analysis was used to examine the either Djago2 or DjpiwiA expression levels between irradiated and un-irradiated animals. Total RNA was collected from three animals in different treatments. A total 3μg RNA was collected from three animals and used for cDNA preparation. Our results showed that γ-irradiation eliminated the expression of DjpiwiA and reduced the level of Djago2 down to 23% as compared to the control. This demonstrated that γ-irradiation had eliminated neoblasts and reduced
30
Djago2 expression in intact animals (Fig. 6C). I also compared Djago2 expression
patterns between irradiated and un-irradiated animals by WISH (Fig. 7A). Djago2 was expressed in the CNS in both irradiated and un-irradiated animals. But the detected signals were weaker in the intact bodies of irradiated worms. Interestingly, signals in cell clusters of the dorsal middle line were totally eliminated by γ-irradiation, suggesting that irradiation indeed depleted Djago2-positive neoblasts in the dorsal middle line. Comparing the expression of Djago2 in irradiated and un-irradiated animals on day 3 post amputation, I found that the increased expression of Djago2 near the wound site had disappeared as well (Fig. 7B).
3.3.2 Djago2 is highly expressed in neoblasts.
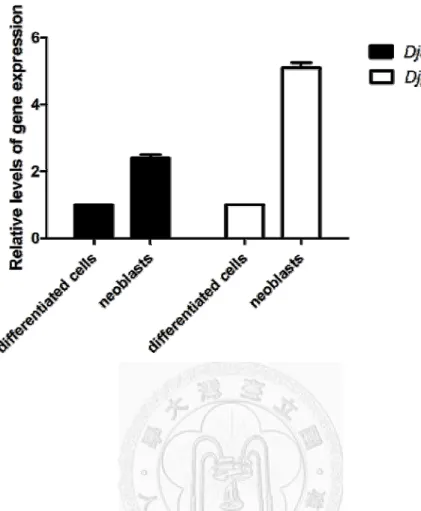
To confirm the expression of Djago2 in highly proliferating neoblasts, I collected G2/M-phase cells and analyzed their Djago2 levels by an RT-qPCR. Single-cell suspensions were prepared from 60 worms at ~1.5 cm in length for the FACS analysis.
Cells in population 3 (in the G2/M phase) were sorted as neoblasts. Cells in population 5 (in the G0/G1 phase) were sorted as differentiated cells. Cells in population 4, which contained part of the neoblasts, were not sorted. In total, 300 ng RNA was collected and subjected to cDNA synthesis. I compared Djago2 and DjpiwiA expressions between
31
neoblasts and differentiated cells. The Djago2 expression level was higher in neoblasts (Fig. 8).
3.4.1 Failure to regenerate in Djago2-silenced animals.
From previous data, I already knew that the Djago2 expression level is higher in regenerating tissue, and Djago2 is highly expressed in neoblasts. To check the function of Djago2 in planarian regeneration, I knocked down Djago2 by administering dsRNA.
I observed loss of the head and tail formation in planarian regeneration with one round of Djago2 dsRNA feeding, but not in control animals (Fig. 9B). Pleiotropic regeneration defects were observed in Djago2-knocked down planarians. Fifteen of 30 trunks were observed to have no blastema formation on day 7 after transverse amputation in the anterior and posterior regions of the pharynx (Fig. 9C). Seven of 30 trunks were observed to have limited regeneration with a single photoreceptor or no photoreceptor formation on day 7 after amputation. I extended the observation period to 14 days.
Three of seven trunks had formed a complete photoreceptor, others had formed an incomplete photoreceptor, while five of 30 animals died. In control animals, 29 of 30 trunks regenerated to intact animals, and one trunk died (Fig. 9C). Higher knockdown efficiency was observed by increasing the feeding times of Djago2 dsRNA (data not
32
shown), while I also found that animals began degenerating at the head region. However, most trunks did regenerate and died when I fed them Djago2 dsRNA more than five times.
I examined the expression of Djago2 in animals that had failed to regenerate by an RT-qPCR. The expression level of Djago2 was reduced to 11% after Djago2 silencing.
Expression levels of DjpiwiA, DjpiwiC, and Djpcna were also reduced to 3%, 16%, and 17% at 7 days after amputation (Fig. 9C). This suggested that depletion of Djago2 may eliminate a large population of neoblasts and also that Djago2 is required for maintaining the homeostasis of neoblasts.
I therefore checked if neoblasts were indeed eliminated in Djago2-depleted worms using a FACS analysis. Surprisingly, the FACS results showed that cells of population 3 (in the G2/M phase) were not depleted in animals that had failed to regenerate (Fig. 9D).
Moreover, the percentage of G2/M-phase cells had increased in animals that had failed to regenerate (Fig. 9E), suggesting that an undefined mechanism may be involved in the regenerating deficiency phenotype of Djago2-depletion.
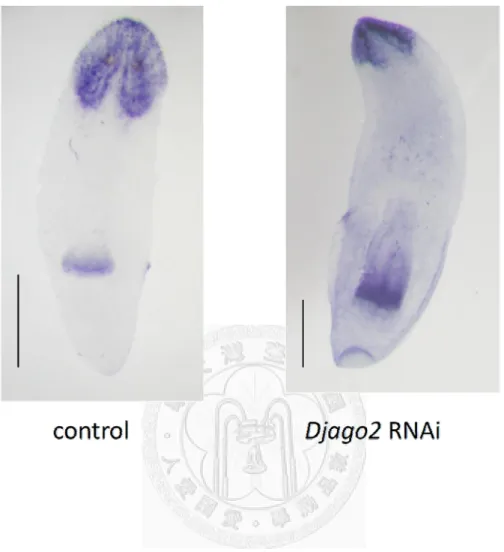
For those animals which showed incomplete regeneration with Djago2 RNAi, I further checked the pattern of formation of the head region to understand if DjAgo2 is also required for differentiation. Djndk is a head marker in planarians used to show the head
33
formation in control and limited-regeneration (RNAi) animals 14 days after amputation.
I examined expression patterns of Djndk by WISH. Compared to control RNAi animals, incomplete head formation was observed in Djago2 RNAi-treated animals (Fig. 10).
This result suggested that the differentiation potential from neoblast progeny was restricted in Djago2 RNAi-treated animals.
3.4.2 Tissue homeostasis defect was observed in long-term Djago2 RNAi-treated
animals.
As mentioned above, I observed the head-regression phenotype when I increased the Djago2 dsRNA feeding rounds. I therefore increased the dsRNA concentration in the RNAi food (6 mg of dsRNA in 25 μl of liver) to check if this degeneration phenotype resulted from the complete knockdown of Djago2. The head-regression phenotype was observed when RNAi food was given 3 to 6 times (Fig. 11A). This phenotype was similar to that of irradiated animals (Fig 6A). I examined the expression of Djago2 in head-degenerated animals by an RT-qPCR. The Djago2 expression level was reduced to 57%. Respective expression levels of DjpiwiA and Djpcna were reduced to 43% and 40% (Fig. 11B). These results suggested dysfunction of neoblasts in Djago2 RNAi-treated animals.
34
Since neoblasts are the only mitotic cells in planarians as far as is known, I labeled mitotic cells with anti-phospho-histone 3-serine 10 (H3P), to confirm if the depletion of Djago2 affected the M-phase of mitosis. As shown in Fig. 11C, numbers of
H3P-positive cells dramatically decreased in Djago2-silenced animals. However, cell cytometric data showed that G2/M-phase cells were not significantly reduced in degenerating animals (Fig. 11D). Similar to animals that had failed to regenerate (Fig.
9E), the ratio of G2/M-phase cells even increased in degenerating animals compared to the control (Fig. 11E). A summary of these results indicated that RNAi of Djago2 blocked dividing neoblasts.
35
4. Discussion
4.1 DjAgo2 is a member of the Argonaute subfamily.
Argonaute proteins are highly conserved in many organisms. Members of the Argonaute protein family are characterized by the presence of PAZ (Piwi-Argonaute-Zwille), Mid, and PIWI domains. The protein sequence of DjAgo2 is most similar to Drosophila Ago1 and human AGO2, and DjAgo2 has homology to the Argonaute subfamily (Fig. 1). The PAZ domain of DjAgo2 has 79.7% identity with Drosophila Ago1 and 75.7% identity with human AGO2. The PIWI domain of DjAgo2
has 84.8% identity with Drosophila Ago1 and 85.8% identity with human AGO2. The PAZ domain is bound to the 3’ end of small RNA. The PIWI domain is bound to the 5’
end of small RNA. This shows that DjAgo2 might have small RNA-binding affinity.
The PIWI domain of DjAgo2 contains the catalytic residue, DDH, which is an essential motif for cleavage activity. This indicates that DjAgo2 potentially has mRNA-cleavage activity. In Drosophila, Ago1 functions in the miRNA-mediated translation repression pathway (Miyoshi et al., 2005), and Ago2 plays major roles in the RNAi pathway. Both of them have slice activity, and are involved in different pathways. The protein sequence of DjAgo2 is more closely related to Drosophila Ago1 than to Ago2, suggesting that the
36
function of DjAgo2 is more related to miRNA pathways. In addition, I found that when YFP-tagged DjAgo2 was expressed in HeLa cells, it also localized to P-body and co-localized with human RCK, a P-body component (Fig. 2). Since human Ago2 is localized to cytoplasmic P-bodies which are mRNA decapping and degradation sites (Hutvagner and Simard, 2008). Our result suggests that DjAgo2 might have a role in miRNA-mediated mRNA degradation.
4.2 Djago2 is up regulated in regenerating tissue during planarian regeneration.
The expression level of Djago2 increased at the wound site (Fig. 4A). During regeneration, the expression level of Djago2 increased in the regenerating region from 8 hours to 3 days after amputation, and then decreased (Fig. 4B). In a quantification analysis, regenerating tissue was separated into two regions: the blastema and post-blastema. Post-blastema tissue contains many neoblasts which proliferate and differentiate into progenitor cells. These cells migrate to the blastema, and then differentiate into various types of differentiated cells. In the blastema, progenitor cells do not undergo mitosis. During regeneration, the expression of Djago2 was increased 2.5-fold at 8 h post-amputation, and then had declined to 1.7-fold higher than the control at 48 h post-amputation (Fig. 5B). The expression of Djago2 reached a peak at
37
72 h post-amputation (Fig. 5B). This phenomenon is similar to the change in neoblast proliferative activity during regeneration(Salo and Baguna, 1984). The increased level of Djago2 is caused by the aggregation and proliferation of neoblasts. In the blastema, the expression of Djago2 had increased to 6-fold higher than the control at 72 h post-amputation, and then dropped back to the control level (Fig. 5C). In the blastema, elevation of Djago2 may be caused by a wound response of differentiated cells or the differentiation of progenitor cells. To clarify if the increase of Djago2 is a result of wound response or a signal from progenitor cells, we examine the expression level of Djago2 at 72 h post amputation in irradiated animals. The expression of Djago2 was not
higher in irradiated animals on day 3 post-amputation (Figs. 4A, 7B). Therefore, the increased level of Djago2 did not result from a wound response of differentiated cells, but from differentiation of progenitor cells.
4.3 Expression level of Djago2 is higher in neoblasts than in differentiated cells.
My data show that Djago2 is expressed in clustering cells which are located in the middle dorsal line (Fig. 2A). DjpiwiA is also expressed in the middle dorsal line (Fig.
3C). FACS data showed that 90 Gy of γ-irradiation eliminated neoblasts (Fig. 7B), while Djago2-positive cells in the middle line were also eliminated. RT-qPCR data
38
confirmed these results. The eliminated cells were neoblasts. The expression level of Djago2 was lower in irradiated animals (Fig. 6C) and was higher in neoblasts (Fig. 8).
4.4 Djago2 is required for proliferation of neoblasts and differentiation of
progenitor cells.
In my data, three phenotypes were observed in Djago2-silenced animals. First, failure of regeneration was observed after Djago2 RNAi food was administered 3 times (Fig. 9B). Parts of the trunk could not form a blastema at the wound site, and animals died within 14 days after amputation. This phenotype was similar to silencing of Djbruli, Smedwi-2, Smed-smB (Fernandez-Taboada et al., 2010), DjPCNA (Orii et al., 2005),
and DjRbAp48(Bonuccelli et al., 2010). No blastema formation means no neoblasts or no proliferation. In Djago2 RNAi worms, expression levels of DjpiwiA, Djpcna, and DjpiwiC were lower on day 7 post-amputation (Fig. 9D) with no blastema formation
compared to control animals. DjpiwiA and DjpiwiC were used as neoblasts markers. The number of neoblasts decreased in Djago2-silenced animals.
In the second phenotype, parts of trunk fragments could form a blastema, but failed to form a pair of normal photoreceptors. This phenotype was similar to those of Djcbc-1 and Djtrans-2a. This indicates that Djago2 may be required for differentiation of
39
progenitor cells. Animals exhibiting limited head regeneration with a single photoreceptor were used to examine the ability of progenitor cells to differentiate. In a comparison of expression patterns of Djndk between normal animals and Djago2 RNAi-treated animals, an incomplete brain was observed in animals with only one photoreceptor (Fig. 10). This indicated that Djago2 silencing limited the differentiation capability of progenitor cells. However, we still need to check which types of differentiated cells can’t be formed from progenitor cells in Djago2 silenced animals.
The regeneration of many organs other than brain also needs to be monitored to confirm that Djago2 silencing limits the ability of progenitor cells to differentiate to other terminally differentiated cells (Oviedo and Levin, 2007).
In the third phenotype, homeostasis defects were observed when dsRNA was fed more than 3 times. Levels of DjpiwiA, Djpcna, and DjpiwiC decreased in both degenerating and regeneration-failed animals. Surprisingly, our FACS data showed that the number of G2/M-phase cells was increased in failed-regeneration animals (Figs. 9F, 11E). Since the mitotic cells can be also labeled by anti-H3P antibody, we found that the amounts of anti-H3P-positive cells were reduced in homeostasis-defective animals (Fig.
11C). This shows that Djago2 silencing blocked the mitosis of neoblasts. It implied that, in Djago2-silenced animals, the cell cycle of neoblasts was stopped in the G2 phase.
40
These data suggest that head regression is resulted from a deficiency of neoblasts. My data showed that Djago2 silencing led to animal regression and death, which might have been caused by apoptosis (Naoghare et al., 2011).
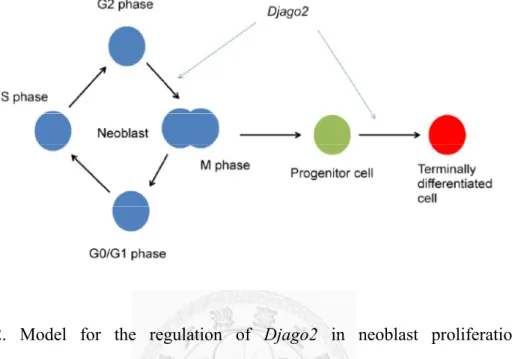
Taken together, our data suggest that Djago2 is required for planarian regeneration and tissue homeostasis. Djago2 is also required for proliferation and self-renewal of neoblasts and differentiation of progenitor cells (Fig. 12). In previous studies, inhibition of many RNA granule components, including Djbruli, Djvas-1, Djcbc-1, Djupf-1, Djxrn-1, Djedc-4, Djdicer and translation initiation factors, leds to lethality or limited
regeneration (Anderson and Kedersha, 2006; Rouhana et al., 2010). This suggests that post-transcriptional regulators are required for stem-cell proliferation and planarian regeneration. Since Djago2 is an orthologue of hsago2 with high conservation, it is possible that Djago2 also mediates small RNA-mediated gene silencing that regulates neoblast proliferation and progenitor cell differentiation. In the future, involvement of Djago2 in planarian miRNA- or siRNA-mediated gene regulation should be further
confirmed. We expect to identify the miRNAs regulated by Djago2 in planarians by the immunoprecipitation of Djago2. In my preliminary data, several miRNAs were found to be up-regulated in regenerating tissue. We will further confirm if these miRNAs are associated with Djago2, and are required for planarian regeneration. Many following
41
experiments, such as using miRNA inhibitors to block the function of miRNA in planarians, will be conducted in the near future.
42
5. References
Adell, T., Salo, E., Boutros, M., and Bartscherer, K. (2009). Smed-Evi/Wntless is required for beta-catenin-dependent and -independent processes during planarian regeneration. Development 136, 905-910.
Anderson, P., and Kedersha, N. (2006). RNA granules. J Cell Biol 172, 803-808.
Aukerman, M.J., and Sakai, H. (2003). Regulation of flowering time and floral organ identity by a MicroRNA and its APETALA2-like target genes. Plant Cell 15, 2730-2741.
Baguna, J., and Romero, R. (1981). Quantitative-Analysis of Cell-Types during Growth, Degrowth and Regeneration in the Planarians Dugesia-Mediterranea and
Dugesia-Tigrina. Hydrobiologia 84, 181-194.
Baguna, J., Salo, E., and Auladell, C. (1989). Regeneration and Pattern-Formation in Planarians .3. Evidence That Neoblasts Are Totipotent Stem-Cells and the Source of Blastema Cells. Development 107, 77-86.
Bardeen, C.R., and Baetjer, F.H. (1904). The inhibitive action of the Roentgen rays on regeneration in planarians. Journal of Experimental Zoology 1, 191-195.
Bartel, D.P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell
43
116, 281-297.
Betchaku, T. (1967). Isolation of planarian neoblasts and their behavior in vitro with some aspects of the mechanism of the formation of regeneration blastema. J Exp Zool 164, 407-433.
Betchaku, T. (1970). The cellular mechanism of the formation of a regeneration
blastema of fresh-water planaria, Dugesia dorotocephala. I. The behavior of cells in a tiny body fragment isolated in vitro. J Exp Zool 174, 253-279.
Bohmert, K., Camus, I., Bellini, C., Bouchez, D., Caboche, M., and Benning, C. (1998).
AGO1 defines a novel locus of Arabidopsis controlling leaf development. EMBO J 17, 170-180.
Bonuccelli, L., Rossi, L., Lena, A., Scarcelli, V., Rainaldi, G., Evangelista, M., Iacopetti, P., Gremigni, V., and Salvetti, A. (2010). An RbAp48-like gene regulates adult stem cells in planarians. J Cell Sci 123, 690-698.
Brøndsted, H.V. (1969). Planarian regeneration. Pergamon Press. Oxford.
Brennecke, J., Aravin, A.A., Stark, A., Dus, M., Kellis, M., Sachidanandam, R., and Hannon, G.J. (2007). Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128, 1089-1103.
Brennecke, J., Hipfner, D.R., Stark, A., Russell, R.B., and Cohen, S.M. (2003). bantam
44
encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 113, 25-36.
Brennecke, J., Stark, A., Russell, R.B., and Cohen, S.M. (2005). Principles of microRNA-target recognition. PLoS Biol 3, e85.
Cai, X., Hagedorn, C.H., and Cullen, B.R. (2004). Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 10, 1957-1966.
Carmell, M.A., Xuan, Z.Y., Zhang, M.Q., and Hannon, G.J. (2002). The Argonaute family: tentacles that reach into RNAi, developmental control, stem cell maintenance, and tumorigenesis. Gene Dev 16, 2733-2742.
Cerutti, H., and Casas-Mollano, J.A. (2006). On the origin and functions of RNA-mediated silencing: from protists to man. Curr Genet 50, 81-99.
Chai, G., Ma, C., Bao, K., Zheng, L., Wang, X., Sun, Z., Salo, E., Adell, T., and Wu, W.
(2010). Complete functional segregation of planarian beta-catenin-1 and -2 in mediating Wnt signaling and cell adhesion. J Biol Chem 285, 24120-24130.
Chen, C.Z., Li, L., Lodish, H.F., and Bartel, D.P. (2004). MicroRNAs modulate hematopoietic lineage differentiation. Science 303, 83-86.
Conte, M., Deri, P., Isolani, M.E., Mannini, L., and Batistoni, R. (2009). A mortalin-like
45
gene is crucial for planarian stem cell viability. Developmental Biology 334, 109-118.
Eisenhoffer, G.T., Kang, H., and Sanchez Alvarado, A. (2008). Molecular analysis of stem cells and their descendants during cell turnover and regeneration in the planarian Schmidtea mediterranea. Cell Stem Cell 3, 327-339.
Farrar, G.J., Millington-Ward, S., Chadderton, N., Humphries, P., and Kenna, P.F.
(2011). Gene-based therapies for dominantly inherited retinopathies. Gene Ther.
Fernandez-Taboada, E., Moritz, S., Zeuschner, D., Stehling, M., Scholer, H.R., Salo, E., and Gentile, L. (2010). Smed-SmB, a member of the LSm protein superfamily, is essential for chromatoid body organization and planarian stem cell proliferation.
Development 137, 1055-1065.
Friedlander, M.R., Adamidi, C., Han, T., Lebedeva, S., Isenbarger, T.A., Hirst, M., Marra, M., Nusbaum, C., Lee, W.L., Jenkin, J.C., et al. (2009). High-resolution profiling and discovery of planarian small RNAs. Proc Natl Acad Sci U S A 106, 11546-11551.
Gentile, L., Cebria, F., and Bartscherer, K. (2011). The planarian flatworm: an in vivo model for stem cell biology and nervous system regeneration. Dis Model Mech 4, 12-19.
46
Guo, T., Peters, A.H., and Newmark, P.A. (2006). A Bruno-like gene is required for stem cell maintenance in planarians. Dev Cell 11, 159-169.
Gurley, K.A., Rink, J.C., and Sanchez Alvarado, A. (2008). Beta-catenin defines head versus tail identity during planarian regeneration and homeostasis. Science 319, 323-327.
Hatfield, S.D., Shcherbata, H.R., Fischer, K.A., Nakahara, K., Carthew, R.W., and Ruohola-Baker, H. (2005). Stem cell division is regulated by the microRNA pathway. Nature 435, 974-978.
Hayashi, T., Asami, M., Higuchi, S., Shibata, N., and Agata, K. (2006). Isolation of planarian X-ray-sensitive stem cells by fluorescence-activated cell sorting.
Development Growth & Differentiation 48, 371-380.
Hendzel, M.J., Wei, Y., Mancini, M.A., Van Hooser, A., Ranalli, T., Brinkley, B.R., Bazett-Jones, D.P., and Allis, C.D. (1997). Mitosis-specific phosphorylation of histone H3 initiates primarily within pericentromeric heterochromatin during G2 and spreads in an ordered fashion coincident with mitotic chromosome
condensation. Chromosoma 106, 348-360.
Hock, J., and Meister, G. (2008). The Argonaute protein family. Genome Biol 9, 210.
Hori, I. (1991). Role of fixed parenchyma cells in blastema formation of the planarian
47
Dugesia japonica. Int J Dev Biol 35, 101-108.
Hoshi, M., Kobayashi, K., Arioka, S., Hase, S., and Matsumoto, M. (2003). Switch from Asexual to Sexual Reproduction in the Planarian Dugesia ryukyuensis. Integr Comp Biol 43, 242-246.
Houbaviy, H.B., Murray, M.F., and Sharp, P.A. (2003). Embryonic stem cell-specific MicroRNAs. Dev Cell 5, 351-358.
Hutvagner, G., and Simard, M.J. (2008). Argonaute proteins: key players in RNA silencing. Nat Rev Mol Cell Biol 9, 22-32.
Johnston, R.J., and Hobert, O. (2003). A microRNA controlling left/right neuronal asymmetry in Caenorhabditis elegans. Nature 426, 845-849.
Kim, K., Lee, Y.S., and Carthew, R.W. (2007). Conversion of pre-RISC to holo-RISC by Ago2 during assembly of RNAi complexes. RNA 13, 22-29.
Lee, Y., Ahn, C., Han, J., Choi, H., Kim, J., Yim, J., Lee, J., Provost, P., Radmark, O., Kim, S., et al. (2003). The nuclear RNase III Drosha initiates microRNA processing. Nature 425, 415-419.
Lee, Y., Jeon, K., Lee, J.T., Kim, S., and Kim, V.N. (2002). MicroRNA maturation:
stepwise processing and subcellular localization. EMBO J 21, 4663-4670.
Lewis, B.P., Burge, C.B., and Bartel, D.P. (2005). Conserved seed pairing, often flanked