行政院國家科學委員會專題研究計畫 成果報告
人類基因體成長動力學模型
計畫類別: 個別型計畫 計畫編號: NSC94-2112-M-006-015- 執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 國立成功大學物理學系(所) 計畫主持人: 楊緒濃 共同主持人: 陳耀煌 計畫參與人員: 楊緒濃,陳耀煌, 報告類型: 精簡報告 處理方式: 本計畫可公開查詢 中 華 民 國 95 年 10 月 5 日行政院國家科學委員會補助專題研究計畫成果報告
人類基因體成長動力學模型
計畫類別:■ 個別型計畫 □ 整合型計畫 計畫編號:NSC 94-2112-M-006-015 執行期間:94 年 08 月 01 日至 95 年 07 月 31 日 計畫主持人:楊緒濃
共同主持人:陳耀煌 計畫參與人員: 成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告 本成果報告包括以下應繳交之附件: □赴國外出差或研習心得報告一份 □赴大陸地區出差或研習心得報告一份 □出席國際學術會議心得報告及發表之論文各一份 □國際合作研究計畫國外研究報告書一份 處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列 管計畫及下列情形者外,得立即公開查詢 □涉及專利或其他智慧財產權,□一年□二年後可公開查詢 執行單位:國立成功大學 中 華 民 國九 十 五
年十
月 二十五
日行政院國家科學委員會專題研究計畫成果報告
國科會專題研究計畫成果報告撰寫格式說明
Preparation of NSC Project Reports
計畫編號:NSC 94-2112-M-006-015 執行期限:94 年8 月1 日至95 年7 月31 日 主持人:楊緒濃 國立成功大學物理學系 共同主持人:陳耀煌 崑山科技大學電子工程系
摘要
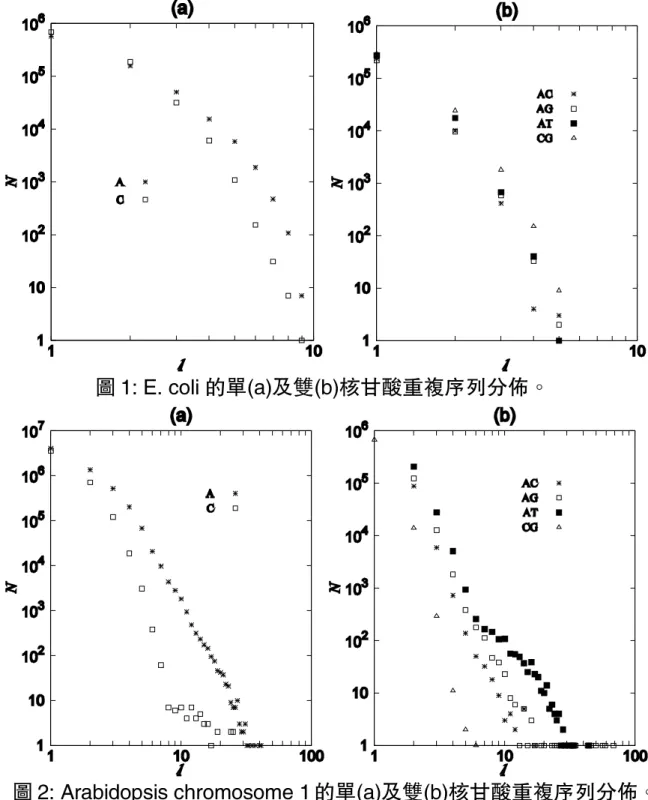
在這次的研究,我們對各種生物物種的基因組做其簡 單重複序列長度之分佈,並對其結果做一個合理的解 釋。我們將重複序列之長度分佈分為單元核甘酸的重複 序列分佈和雙元核甘酸的重複序列分佈。從這些分佈圖, 我們發現核甘酸 A 和 T 的相關重複序列有明顯的長距 離相關性。這個現象的主要原因是雙螺旋狀的去氧核糖 核酸裹的亙補核甘酸 A 和 T 是雙氫鍵,而亙補核甘酸 C 和 G 卻是三個氫鍵,也就是說,核甘酸 A 和 T 之間 的作用力比核甘酸 C 和 G 的作用力來的小。所以核甘酸 A 和 T 相關的重複序列比核甘酸 C 和 G 相關的重複序 列較容易演化,形成其分佈有長尾巴的性質。特別的是, 單核甘酸 A 和 T 重複序列的分佈之尾巴又比雙核甘酸 A 和 T 重複序列的長。核甘酸 C 和 G 相關的重複序列之分 佈都比較短。 1. 簡介 微衛星或稱之為簡單重複序列(SSRs),短縱列重複序列(STRs)是一很短 (一至六個鹼基)的核甘酸重複單元圖案。這些重複序列非常的廣泛分佈在真核 生物的基因組裹。因為重複序列分佈圖案在人類基因組能助於診斷或鑒定人類 基因疾病[1-12],所以能 了解各物種的基因組重複序列分佈之特性是非常 重要的。另外,重複序列之分佈對了解各生物物種的演化有很大的相關性[13-19]。 STRs 出現在去氧核糖核酸DNA序列的編碼區及非編碼區,不同區域 有不同的演化過程,主要是因為編碼區突變會受到抑制,所以不同區域的演 化程度非常不一樣的。在高等動(真核生物)基因組中的編碼區所佔的比例是相 當的小。例如,人類的基因組中的非編碼區,一般稱之為「垃圾基因」,佔全部 基因組的百分之九十七以上,所以在這次的研究褱,我們將對編碼區和非編 碼區不加於分辨。 2. 方法與材料 我們所分析的基因序列物種列於表 1,序列的取得從ftp://ftp.ncbi.nih.gov/genomes/。基因序列的檔案格式為 FASTA 格式,序列中 還包含著部分未確定的鹼基標著為”N”。從表 1 所列的物種基因組,我們將 計算它們的單及雙核甘酸重複序列的長度分佈。 表 1 3. 結果與結論 我們對表一的生物物種做分析,得到的各物種基因組的重複序列之長度 分佈,將其結果用以下圖表示之。以下之圖(a)為單元核甘酸的重複序列分佈, 圖(b)雙元核甘酸的重複序列分佈。從這些分佈圖,我們可以發現核甘酸 A 和 T 的相關重複序列有明顯的長距離相關性。這個現象的主要原因是雙螺旋狀的 去氧核糖核酸裹的亙補核甘酸 A 和 T 是雙氫鍵,而亙補核甘酸 C 和 G 卻是三 個氫鍵。也就是說,核甘酸 A 和 T 之間的作用力比核甘酸 C 和 G 之間的作用 力來的小。所以核甘酸 A 和 T 相關的重複序列比核甘酸 C 和 G 相關的重複序 列較容易演化,形成其分佈有長尾巴的性質。特別的是,單核甘酸 A 和 T 重 複序列的分佈之尾巴又比雙核甘酸 A 和 T 重複序列的長。核甘酸 C 和 G 相關 的重複序列之分佈都比較短。 分類名稱 本文中用名詞 染色體之數目 原核生物
Escherichia coli K12 E. coli 1 真核生物
Arabidopsis thaliana A. thaliana 5 Caenorhabditis elegans nematode 5+X Drosophila melanogaster fruit fly 6 Arms Homo sapiens human 22+XY Mus musculus mouse 19+XY Rattus norvegicus rat 20+X Saccharomyces cerevisiae yeast 16
圖 1: E. coli 的單(a)及雙(b)核甘酸重複序列分佈。
圖 3: Nematoda chromosome 1 的單(a)及雙(b)核甘酸重複序列分佈。
圖 5:Human chromosome 1 的單(a)及雙(b)核甘酸重複序列分佈。
圖 7:Rat chromosome 20 的單(a)及雙(b)核甘酸重複序列分佈。
圖 8: Yeast chromosome 1 的單(a)及雙(b)核甘酸重複序列分佈。 *參考文獻*
[1] Deka R. et al., Intra- and inter-population diversity at short tandem repeat loci in diverse populations of the world. In: Proceedings of the Third International Conference on DNA Finger Printing, Hyerabad, India, December 13-16, 1994., Electrophoresis 16 (1995) pp. 1659-1664.
[2] Dib C. et al., A comprehensive genetic map of the human genome based on 5,264 microsatellites, Nature 380 (1996) pp. 152-154.
[3] Han H.-J. et al., Genetic instability in pancreatic cancer and poorly differ-entiated type of gastric cancer, Cancer Research 53 (1993) pp. 5087-5089. [4] Lazzereschi D. et al., Microsatellite instability in thyroid tumours and tu-mour-like lesions, British Journal of Cancer 79 (1994) pp. 340-345.
[5] Rhyu M. G., Park W. S. & Meltzer S. J., Microsatellite instability occurs frequently in human gastric carcinoma, Oncogene 9 (1994) pp. 29-32.
[6] Orlow I. et al., Chromosome 9 allelic losses and microsatellite alterations in human bladder tumors, Cancer Research 54 (1994) pp. 2848-2851. [7] Eshleman J. R. & Markowitz S. D., Mismatch repair defects in human carcinogenesis, Hum. Mol. Genet. 5 (1996) pp. 1489-1494.
[8] Thomas D. C., Umar A. & Kunkel T. A., Mutation Research - Fundamen-tal and molecular mechanisms of mutagenesis 350 (1996) pp. 201--205. [9] Gurin C. C., Federici M. G., Kang L. & Boyd J., Causes and conse-quences of microsatellite instability in endometrial carcinoma, Cancer Re-search 59 (1999) pp. 462-466.
[10] Foster E. A. et al., Jefferson fathered slave's last child, Nature 396 (1998) pp. 27-28.
[11] Ohta T. & Kimura M., A model of mutation appropriate to estimate the number of electrophoretically detectable alleles in a finite population, Genet. Res. 22 (1973) pp. 201-204.
[12] Morral N., Nunes V., Casals T. & Estivill X., CA/GT microsatellite alleles within the cystic fibrosis transmembrane conductance regulator (CFTR) gene are not generated by unequal crossing over, Genomics 10 (1991) pp. 692-698.
[13] Dokholyan N. V., Buldyrev S. V., Havlin S. & Stanley H. E., Model of unequal chromosomal crossing over in DNA sequences, Physica A 249 (1998) pp. 594-599.
[14] Freudenreich C. H. Kantrow S. M. & Zakian V. A., Expansion and length-dependent fragility of CTG repeats in yeast, Science 279 (1998) pp. 853-856.
[15] Durrett R. T. & Kruglyak S., A new stochastic model of microsatellite evolution, J. Appl. Prob. 36 (1999) pp. 621-631.
[16] Lai Y. & Sun F., The relationship between microsatellite slippage muta-tion rate and the number of repeat units, Mol. Biol. Evol. 20 (2003) pp. 2123-2131.
[17] Moxon E. R. & Wills C., DNA microsatellites: Agents of Evolution?, Sci-entific American, Jan. (1999) pp. 72-77.
[18] Batzer M. A. & Deininger P. L., Alu repeats and human genomic diver-sity, Nature Reviews Genetics 3 (2002) pp. 370-379.
[19] Price A. L., Eskin E. & Pevzner P. A., Whole-genome analysis of Alu re-peat elements reveals complex evolutionary history, Genome Res. 14 (2004) pp. 2245-2252.