台灣梅花鹿取食偏好及磨角行為 與墾丁森林稚樹族群變動之關係
葉定宏1,2) 郭耀綸3,4) 林照松1)
摘 要
恆春地區野放的臺灣梅花鹿已對墾丁高位珊瑚礁森林動態樣區植群的更新造成衝擊。本研究測試 野外台灣梅花鹿對墾丁高位珊瑚礁森林樹種葉片的取食偏好,並調查稚樹遭梅花鹿磨角情形,探討鹿 隻取食偏好與不同樹種幼齡稚樹族群變動率的關係。取食偏好結果顯示,梅花鹿對有效受測77樹種葉 片的啃食率具明顯差異,有9樹種全數葉片會被梅花鹿取食,適口性等級最高,例如山黃麻、稜果榕、
血桐等。然而,有12樹種葉片完全不被梅花鹿取食,適口性等級最低,包括紅柴、樹杞、海州常山、
厚殼樹、梧桐、白樹仔、石苓舅、大葉楠、台灣山桂花、粗糠柴、台灣梭羅樹及假酸漿。磨角痕跡調 查發現樹幹磨損程度隨植株徑級加大而減輕,遭嚴重磨損植株的徑級以小於2 cm的幼齡稚樹居多。紅 柴及毛柿分別為梅花鹿喜歡,以及避免磨角的樹種。梅花鹿對不同樹種的啃食率,與墾丁森林動態樣 區2001~2013年間,26樹種的幼齡稚樹族群減少率有極顯著正相關,梅花鹿適口性越高的樹種,其 稚樹族群減少率越大。梅花鹿取食偏好較高的樹種,若其族群空間分布集中在隆起礁岩生育地,則其 族群減少率較小,幼齡稚樹能躲避梅花鹿的取食傷害。此外,本研究也發現墾丁高位珊瑚礁森林71樹 種,由光合潛力代表的耐陰性,與梅花鹿取食偏好並不具顯著相關。
關鍵詞:啃食率、磨角痕跡、適口性等級、族群變動率、耐陰性。
葉定宏、郭耀綸、林照松。2021。台灣梅花鹿取食偏好及磨角行為與墾丁森林稚樹族群變動之關係。
台灣林業科學36(1):69-86。
1) 林 業 試 驗 所 森 林 恆 春 研 究 中 心 ,9 4 6 0 0 5 屏 東 縣 恆 春 鎮 墾 丁 里 公 園 路 2 0 3 號 H e n g c h u n R e s e a r c h Center, Taiwan Forestry Research Institute, 203 Gougyuan Rd., Kenting Village, Hengchun Township, Pingtung County 946005, Taiwan.
2) 國 立 屏 東 科 技 大 學 生 物 資 源 博 士 班 ,9 1 2 0 1 屏 東 縣 內 埔 鄉 學 府 路 1 號 G r a d u a t e I n s t i t u t e o f Bioresources, National Pingtung Univ. of Science and Technology, 1 Xuehfu Rd., Neipu Township, Pingtung 91201, Taiwan.
3) 國立屏東科技大學森林系,91201屏東縣內埔鄉學府路1號 Department of Forestry, National Pingtung Univ. of Science and Technology, 1 Xuehfu Rd., Neipu Township, Pingtung 91201, Taiwan.
4) 通訊作者 Corresponding author, e-mail:[email protected]
2021年1月送審 2021年5月通過 Received January 2021, Accepted May 2021.
Research paper
Browsing Preferences of and Fraying Behavior by Formosan Sika Deer (Cervus nippon taiouanus)
and Their Relationships with Changes
in Sapling Populations in the Kenting Karst Forest
Ding-Hong Yeh,1,2) Yau-Lun Kuo,3,4) Chao-Song Lin1)
【Summary】
The reintroduced Formosa Sika deer are impacting regeneration of forest vegetation in the Kenting Karst Forest Dynamic Plot (KKFDP). In this study, browsing preferences of deer of leaf- bearing twigs were tested, and saplings of various species were surveyed for fraying scars caused by deer to investigate the relationship between browsing preferences and rates of change of juve- nile populations of different tree species. Results showed that browsing rates significantly differed among 77 tested tree species. Twigs of 9 species were entirely browsed, including Trema orienta- lis, Ficus septica, Macaranga tanarius, etc., indicating that these species were the most palatable to deer. On the other hand, twigs of 12 species were not browsed at all, including Aglaia formosa- na, Ardisia sieboldii, Clerodendrum trichotomum, Ehretia acuminata, Firmiana simplex, Gelonium aequoreum, Glycosmis citrifolia, Machilus japonica var. kusanoi, Maesa perlaria var. formosana, Mallotus philippensis, Reevesia formosana, and Trichodesma calycosum, and thus were the least palatable. Surveys of fraying scars showed that damage to trees was alleviated as the diameter of the saplings increased, and the most severe fraying damage occurred in saplings with a diameter of ≤ 2 cm. The deer preferred saplings of Aglaia formosana and avoided saplings of Diospyros blancoi for their fraying behavior. The browsing rates of 26 tree species had significant positive correlations with rates of decline of juvenile populations between 2001 and 2013 in the KKDFP.
Results showed that the higher the palatability level of a tree species, the higher the rate of decline of its juvenile population. Declining rates of juvenile populations of species with higher palatabil- ity rates were lower if the population was primarily growing on exposed limestone habitats, which prevented browsing damage by deer. In addition, this study found that shade tolerance, represented by the photosynthetic capacity, of 71 tree species of the Kenting Karst Forest had no significant relationship with browsing rates of deer.
Key words: browsing rate, fraying scar, palatability level, population change rate, shade tolerance.
Yeh DH, Kuo YL, Lin CS. 2021. Browsing preferences of and fraying behavior by Formosan Sika deer (Cervus nippon taiouanus) and their relationships with changes in sapling popula- tions in the Kenting Karst Forest. Taiwan J For Sci 36(1):69-86.
緒 言
有蹄類動物族群近幾十年來在北半球溫帶 林擴張,影響局部地區樹種的密度及分布(Gill 1992, Côté et al. 2004, Skarpe and Hester 2008, Takatsuki 2009)。鹿科動物因對取食植物種類 具差異性偏好,致對生態系內不同植物物種的 群聚組成及族群數量具有深遠影響,被視為是 生態系的基石物種(keystone species) (Drayton and Primack 1996, DeCalesta and Stout 1997)。
較受鹿隻喜好的植物受損程度會較高,因此透 過差異性的取食偏好(browsing preference),
鹿隻可改變地被層植物的物種組成,較耐啃食 或 不 受 喜 好 的 植 物 因 而 成 為 主 要 的 組 成 物 種 (Nugent et al. 2001, Kuijper et al. 2010)。台 灣位處亞熱帶地區,因森林經營方式改變及社 會 環 境 變 遷 , 鹿 科 動 物 活 動 造 成 的 樹 木 危 害 也 逐 漸 普 遍 。 除 了 中 央 山 脈 分 布 的 水 鹿(Rusa unicolor swinhoei)之外(Wang et al. 2015),恆 春半島人為野放的臺灣梅花鹿(Cervus nippon taiouanus) (以下簡稱梅花鹿)已對墾丁高位珊瑚 礁森林植群也產生了危害(Wang et al. 2015)。
研 究 顯 示 梅 花 鹿 會 廣 泛 取 食 墾 丁 地 區 的 植物,且對不同植物的取食偏好具差異性(Hu and Wang 1994, Wang et al. 1998, Chen et al.
2009)。Hu and Wang (1994)於社頂梅花鹿復育 區圍籬內,藉跟蹤觀察法記錄5頭梅花鹿啃食之 植物種類,發現會取食151種植物,其中64種 為木本植物,有食葉者(browser)之傾向。Wang et al. (1998)在上述復育區內發現餵食的135種 植物中,有110種會被梅花鹿取食。Chen et al.
(2009)採取社頂地區植物測試台北動物園圈養 梅花鹿的食性偏好,發現餵食的217種植物中 有144種會被取食,並依據枝葉被取食程度區 分為:吃光葉片及芽(100%),吃掉大部分葉子 (99~51%)、吃掉少部分葉子(50~11%)、僅吃數 片葉子或淺嚐(10~1%)、完全沒吃(0%)等5類適 口性等級(palatability level)。該研究並調查社 頂地區植物在野外被梅花鹿取食的食痕,發現 172種植物中有141種會被梅花鹿取食。
墾 丁 高 位 珊 瑚 礁 自 然 保 留 區 位 於 恆 春 半 島 南 端 , 是 臺 灣 唯 一 保 存 較 完 整 的 高 位 珊 瑚
礁 原 始 森 林 。 於1996年在保留區內設立森林 動態樣區,並於2001年完成10 ha樣區的森林 植 群 調 查 , 後 續 定 期 監 測 森 林 組 成 結 構 之 長 期 變 化 。 監 測 結 果 發 現 , 由2001至2013的12 年 期 間 , 動 態 樣 區 內 幼 齡 稚 樹(胸徑1~2 cm) 的密度大幅減少,從2001年約1600株ha-1降至 2013年的770株ha-1,減少比例高達52% (Wang et al. 2015)。該期間不同樹種幼齡稚樹族群量 的 變 動 有 很 大 的 差 異 , 減 少 較 多 的 樹 種 如 血 桐(Macaranga tanarius)及蟲屎(Melanolepis m u l t i g l a n d u l o s a ) , 稚 樹 族 群 量 都 減 少 9 0 % 以 上 ; 但 有 少 數 樹 種 稚 樹 族 群 量 會 增 加 , 例 如 樹 杞(Ardisia sieboldii)及毛柿(Diospyros blancoi),族群量分別增加70及45% (Wang et al.
2015)。該森林原本茂密的林下地被層草本與木 本植群已變為極度稀疏,推測野放梅花鹿的活 動是地被植群及幼齡稚樹減少的重要原因。因 此,各樹種稚樹族群量的增減可能肇因於鹿隻 喜食與否,或對鹿隻取食有著不同的抵抗力或 耐受度(Wang et al. 2015)。
前述關於墾丁地區梅花鹿食性的研究,只 涵蓋墾丁高位珊瑚礁森林組成的部分樹種,或 是測試的情境較為人工化,故各項梅花鹿食性 偏好的研究結果並不完全一致,其樣本廣度也 不足以進行高位珊瑚礁森林幼齡稚樹族群量變 化與梅花鹿食性偏好之相關性分析。Lin et al.
(2017)曾於福山試驗林地投放木本植物葉片測 試野生臺灣山羌(Muntiacus reevesi micrurus)的 取食選擇,結果發現其取食偏好具有樹種間的 差異,可以區分為完全取食、部分取食及完全 不取食三類型。此結果顯示,以少量食物材料 於開放環境進行餵食測試也可以得知鹿科動物 的取食偏好特性。因此,本研究於恆春熱帶植 物園,測試非圈養狀態的梅花鹿,對墾丁高位 珊瑚礁森林眾多樹種的食性偏好或適口性是否 具顯著差異,並與先前研究互相印證。
除了梅花鹿葉片適口性的差異可能影響森 林幼齡稚樹族群量變化之外,森林樹冠開闊度的 長期變化,結合個別樹種耐陰性的差異(Kuo and Yeh 2015),也可能造成不同樹種的稚樹族群量 在同一期間有不同的變動趨勢。資源有效性假說
(resource availability hypothesis)解釋不同植物種 類間,具有不同程度的植物防禦物質成分,並預 期適應於資源良好環境的物種,相較適應於資 源貧瘠環境的物種,會有較快的生長速率,葉 片壽命較短,具有較少的化學防禦物質,因此 有較高的被取食率(Coley et al. 1985, Endara and Coley 2011)。研究發現可生長在低光環境的耐陰 樹種,通常會投資較高比例的化學防禦物質於葉 片,因此有較低的適口性,較能抵抗植食動物的 取食(Coley and Barone 1996)。梅花鹿為食性廣 泛的植食動物,取食行為可能也會受到葉片化學 防禦物質的影響,意味著樹種本身的生理功能性 狀(耐陰性),可能會間接影響不同樹種幼齡植株 被梅花鹿取食的機會,致幼齡稚樹族群量有不同 的變動程度及方向。
此外,食物取得的難易程度也可能會影響 該種類食物的被利用率。Wang et al. (1998)觀察 梅花鹿在野放初期,鹿隻並不會移動到高位珊 瑚礁礁岩頂部覓食,因此空間分布群聚在隆起 礁岩生育地的樹種,可能會受到地形的保護,
稚樹不易遭鹿隻取食危害,這或許也是共同影 響此處稚樹族群量動態變化的另一個因子。
本 研 究 目 的 為 測 試 野 外 梅 花 鹿 對 墾 丁 高 位珊瑚礁森林樹種葉片的取食偏好,並觀察其 磨角行為對林木的傷害樣式,探討鹿隻取食偏 好或磨角行為與樹種幼齡稚樹族群量變化的關 係。本研究提出的問題包括:(1)梅花鹿對墾丁 高位珊瑚礁森林不同樹種葉片的取食偏好(葉片 啃食率leaf browsing rate)有何差異?(2)梅花鹿 對不同樹種的取食偏好若具明顯差異,則葉片 啃食率與該樹種幼齡稚樹族群變動率之間是否 具顯著相關?(3)樹種的耐陰程度與其葉片啃食 率間是否具顯著相關? 若是,樹種的耐陰性是否 與稚樹族群變動率也具顯著相關?(4)主要分布 在隆起礁岩生育地的樹種,其幼齡稚樹族群減 少程度是否較小?
針 對 上 述 各 項 問 題 , 本 研 究 提 出 的 假 說 為:(1)梅花鹿對於墾丁高位珊瑚礁森林不同樹 種葉片的取食偏好有差異,亦即梅花鹿對不同 樹種葉片的啃食率並不相同。(2)適口性較高的 樹種較易遭梅花鹿啃食葉片,其幼齡稚樹族群
的減少率會較大;適口性較低的樹種免於或僅 受輕微的啃食壓力,其幼齡稚樹族群量僅會微 幅減少甚或增加,因此各樹種葉片啃食率與其 幼齡稚樹的族群減少率間會呈顯著正相關。(3) 越耐陰的樹種其葉片具有較高的化學或物理性 防禦能力,令其適口性較低,故耐陰程度越高 的樹種,其葉片啃食率會較低,兩者呈顯著負 相關;耐陰程度越高的樹種其幼齡稚樹的族群 減少率也會較低。(4)相較於平坦谷地,珊瑚礁 礁頂環境較不適合梅花鹿活動;若為梅花鹿喜 好取食,但空間分布集中在隆起礁岩生育地的 樹種,因地形的保護效應,其幼齡稚樹族群的 減少率會較低。
材 料 與 方 法
一、試驗地及主要樹種
於2001年在墾丁高位珊瑚礁自然保留區 內,設立一處面積10 ha的森林動態樣區(Wu et al. 2011),標記胸徑≥ 1 cm木本植物種類、位置 及胸徑;又於2008及2013年進行過兩次複查,
記錄樣區內存活植株的種類及胸徑,計算各樹 種新增及死亡的數量。在此森林動態樣區內調 查到95種木本植物,數量前5名的優勢樹種分別 為黃心柿(Diospyros maritima)、鐵色(Drypetes littoralis)、紅柴(Aglaia formosana)、山柚 (Champereia manillana)、咬人狗(Dendrocnide meyeniana) (Wu et al. 2011)。恆春熱帶植物 園與上述10 ha森林動態樣區最近的距離約200 m,植物園範圍內約有1/3,面積約8 ha為展示 區,栽植多數非原生的木本植物,植物園其它 區域則為天然林,其樹種組成與上述森林動態 樣區類似。
二、梅花鹿對受測樹種葉片取食偏好測試 森林動態樣區內鹿隻族群量較多,受到梅 花鹿取食的歷程較植物園區久,2 m高度以下的 木本植物已經極為稀疏。如果在森林動態樣區 內以小量枝葉進行食性測試,可能多數的種類在 測試時間內即會被全部取食,鑑別不出適口性較 佳樹種之間偏好程度的差異。植物園區由於人為
活動頻繁,梅花鹿族群尚未大規模出現覓食,又 不至於稀少到只有少數鹿隻活動,造成測試失敗 或僅能測試少數個體的取食偏好。經評估選定此 熱帶植物園的展示區與天然次生林交界之處,
距離森林動態樣區約700 m 的地點(21°57’42.5”N 120°48’49.6”E)進行梅花鹿取食偏好測試,測試 時期自2017年7月至2019年5月。
本研究選取墾丁高位珊瑚礁森林85樹種,
測試梅花鹿對其葉片的取食喜好程度。試驗期 間每次選取7樹種各一枝帶有健康葉片的枝條供 測試。各枝條長度約30~60 cm,剪除破損葉片 後記錄可供餵食的葉片數量。受測枝條其中1枝 置於中間,其它6枝環繞於外側排列成六角形,
各枝條相距1.5 m。為防止葉片萎凋影響鹿隻取 食偏好,本研究於土面下埋入貯水之水瓶,再 將枝條末端插入水瓶內,令其持續吸水保持葉 片膨挺,並令枝條直立於地面模擬小苗樣態。
為避免受測枝條被鹿隻叼離無法記錄,測試時 以細繩及夾子將枝條與土面下水瓶連結固定。
各測試日均於傍晚放置新鮮枝條,次日早晨記 錄枝條遭啃食情形。已被啃食或葉片已枯萎的 枝條均將其更換,未被啃食且未枯萎的枝條則 繼續留用測試。供試枝條若都未遭啃食,最多 只做3次(天)的測試。自2017年7月中旬至2018 年9月共進行616次(枝)的測試,某日只要供試 7枝條中有任一枝的葉片被鹿隻啃食,該次即 視為有效測試,該7樹種各1枝條均視為有效樣 本;若某次任一枝條葉片均無被鹿啃食痕跡,
則 判 斷 該 次 無 梅 花 鹿 造 訪 ,7枝條均屬無效樣 本。為減少誤判梅花鹿是否有造訪測試區,自 2018年9月至2019年5月期間共328次的測試,
每次測試的7樹種枝條,均各自與梅花鹿喜食的 伴測樹種(血桐或蟲屎)枝條綁在一起,依伴測 枝條葉片有無被鹿啃食來判斷各受測枝條是否 為有效測試。本研究同一樹種有效枝條樣本要 達3枝(含)以上,才列為有效測試樹種。本研究 原本放置85樹種的枝葉供測試,其中屬於有效 測試的樹種計77種(Table. 1)。梅花鹿對各樹種 葉片啃食率的計算,是估算各有效測試樣本枝 條原有葉片數被取食多少比例,再將同一樹種 所有的有效樣本枝條葉片被取食比例平均,即
為鹿隻對該樹種葉片的啃食率。依照啃食率高 低再將受測樹種分成5個適口性等級(Chen et al.
2009)。
本研究另以Ivlev’s選擇指數(Ivlev’s index of electivity) (Strauss 1979)來計算梅花鹿對各 樹 種 葉 片 的 選 擇 偏 好 程 度 。 此 指 數 考 量 到 動 物 取 食 時 , 因 食 物 本 身 數 量 多 寡 可 能 會 影 響 取 食 偏 好 , 故 需 將 不 同 食 物 種 類 的 可 獲 得 性 (availability),即食物量,標準化。計算時將每 一有效受測枝條視為1單位食物量,如果枝條葉 片被取食比例為60%,則有0.6單位食物量被取 食。此指數計算公式為
E = (Ri – Pi) / (Ri+Pi) ...(1) 指數E代表選擇偏好度;Ri為i種植物被取 食量佔所有被取食量的比例(i種類被取食的單位 食物量總和/所有受測枝條的被取食單位食物量 總和);Pi為i種植物食物量在環境中佔所有潛在 食物量的比例,本研究引伸此參數為i種植物食 物量在測試實驗中佔所有測試食物總量的比例(i 種類有效測試枝條總數/所有受測枝條總數)。此 選擇指數介於 -1.0~1.0之間,負值表示梅花鹿 會避免選擇該種植物;0表示梅花鹿對取食該種 植物並無偏好性;正值表示梅花鹿會主動選擇 啃食該種植物。
三、梅花鹿磨角行為及偏好樹種調查
本研究於2017至2019,共3年期間進行三 項梅花鹿對林木樹幹磨角行為(fraying behavior) 的研究。第一項研究於2017年1~11月在恆春熱 帶植物園展示區,普查成樹樹幹磨痕狀態。第 二項研究是針對稚樹(胸徑≤ 10 cm)遭磨角傷害 的 調 查 , 分 別 在2017年11月、2018年11月及 2019年11~12月間進行,此期間為雄性梅花鹿發 情期,屬磨角行為高峰期;調查對象植株為植 物園展示區天然林步道兩側各5 m,以及森林邊 緣5 m範圍內的稚樹。第三項研究則探討梅花鹿 磨角時是否會選擇或避開某些特定樹種。
在調查項目方面,第一項研究記錄了樹幹 遭梅花鹿磨角的樹種及植株胸徑、磨痕離地高 度、輕微磨痕(磨損未達木質部)的長度、嚴重磨 痕(磨損至木質部裸露)的長度、磨痕位置的樹幹
Table 1. Leaf browsing rate, Ivlev’s index, and photosynthetic capacity (Amax, μmol CO2 m-2 s-1) of 77 tree species which were nibbled by Formosan Sika deer in the Kenting Karst Forest. Amax
provided by Kuo and Yeh (2015) and unpublished data. N/A denotes data not measured.
Species Browsing Ivlev’s Amax Species Browsing Ivlev’s Amax
rate (%) index rate (%) index
Antidesma pentandrum 100 0.46 14.1 Drypetes littoralis 48 0.13 10.5
var. barbatum Planchonella obovata 47 0.12 17.4
Boehmeria nivea 100 0.46 N/A Radermachera sinica 42 0.06 24.7
var. tenacissima
Dendrocnide meyeniana 100 0.46 23.0 Scolopia oldhamii 38 0.01 17.2 Ficus benjamina 100 0.46 22.0 Diospyros ferrea 37 0.00 12.0 Ficus irisana 100 0.46 16.7 Ficus superba var. japonica 33 -0.06 22.2 Ficus septica 100 0.46 23.4 Ficus caulocarpa 33 -0.06 20.6 Macaranga tanarius 100 0.46 31.9 Glochidion philippicum 33 -0.06 21.5 Schefflera octophylla 100 0.46 17.4 Syzygium formosanum 32 -0.07 10.9 Trema orientalis 100 0.46 27.7 Bischofia javanica 28 -0.14 22.7 Dracaena angustifolia 99 0.46 10.2 Melicope semecarpifolia 27 -0.15 16.3 Fraxinus insularis 99 0.46 22.0 Mallotus paniculatus 27 -0.16 29.4 Ficus ampelos 98 0.45 20.8 Ardisia quinquegona 20 -0.30 12.3 Palaquium formosanum 97 0.45 17.6 Gonocaryum calleryanum 19 -0.32 11.8 Koelreuteria henryi 93 0.43 19.1 Cryptocarya concinna 17 -0.37 12.5 Melanolepis multiglandulosa 92 0.43 31.0 Lagerstroemia subcostata 17 -0.38 25.7 Neonauclea reticulata 92 0.42 18.1 Litsea hypophaea 16 -0.39 16.7 Pisonia umbellifera 85 0.40 14.7 Heritiera littoralis 16 -0.41 17.7 Melia azedarach 80 0.36 34.6 Euphoria longana 15 -0.42 N/A Celtis formosana 78 0.35 19.6 Croton cascarilloides 14 -0.44 12.5 Ehretia dicksonii 75 0.34 20.6 Diospyros blancoi 6 -0.71 12.3 Gleditsia rolfei 75 0.34 25.1 Swietenia macrophylla 3 -0.84 17.5 Diospyros maritima 74 0.33 12.2 Viburnum odoratissimum 3 -0.85 16.8 Champereia manillana 73 0.33 12.4 Neolitsea parvigemma 2 -0.90 14.5 Gardenia jasminoides 72 0.32 16.5 Psychotria rubra 2 -0.91 13.8 Eriobotrya deflexa 71 0.32 16.6 Diospyros eriantha 2 -0.91 12.7 Premna serratifolia 71 0.31 27.7 Cinnamomum reticulatum 1 -0.96 16.9 Boehmeria wattersii 69 0.30 16.3 Aglaia formosana 0 -1.00 15.7 Murraya exotica 68 0.29 17.3 Ardisia sieboldii 0 -1.00 15.0 Beilschmiedia erythrophloia 67 0.29 13.5 Clerodendrum trichotomum 0 -1.00 N/A Liodendron formosanum 67 0.29 11.5 Ehretia acuminata 0 -1.00 27.6 Ficus microcarpa var. 67 0.29 23.9 Maesa perlaria var 0 -1.00 17.6
microcarpa formosana
Leea guineensis 66 0.28 20.1 Gelonium aequoreum 0 -1.00 13.6
Kleinhovia hospita 60 0.24 22.6 Glycosmis citrifolia 0 -1.00 10.7
Crateva adansonii subsp. 57 0.21 15.5 Machilus japonica var. 0 -1.00 14.9
formosensis kusanoi
Fraxinus griffithii 56 0.20 24.6 Firmiana simplex 0 -1.00 27.4
Ficus virgata 54 0.19 N/A Mallotus philippensis 0 -1.00 14.3
Sapindus mukorossi 51 0.16 19.4 Reevesia formosana 0 -1.00 13.2
Bridelia balansae 50 0.15 15.4 Trichodesma calycosum 0 -1.00 N/A
直徑。此外,也記錄磨痕的新舊。當年度造成 的磨痕為新磨痕,超過1年則為視為舊磨痕,部 分樹木則同時具有新、舊兩種磨痕。若磨痕傷 口潮濕、顏色較為鮮明有光澤、有鹿毛沾附、
磨損的樹皮纖維還連在樹幹上,則判斷為新磨 痕;若磨痕傷口顏色已變黯淡或已產生癒傷組 織,則判斷為非本季之舊磨痕。第二項針對稚 樹磨痕的研究,記錄了樹種、植株胸徑、環剝 程度、植株葉片是否枯死等項目。將嚴重磨痕 寬度佔樹幹周長的比例視為環剝比例,並將環 剝比例區分為1~29、30~59及60~100%三個級 距,各代表樹木遭輕度、中度及重度環剝。
第三項針對梅花鹿磨角時是否會選擇或避 開某些特定樹種的研究,是於2019年11~12月 在進行上述第二項研究時,一併調查被磨角樹 木相鄰半徑2 m內不同樹種植株是否也遭磨角 (Ramos 2006)。若已遭磨角植株半徑2 m內有植 株大小相近,但未遭磨角的相異樹種植株,則 假設梅花鹿曾經歷二擇一的選擇過程,並選取 該樹種進行磨角,如此完成一次較可信的選擇 事件。某樹種樹幹被梅花鹿磨角比例(磨角率)的 計算方法,為該樹種被磨角次數除以該樹種參 與磨角選擇事件的總次數,即除以被磨次數與 未被磨次數之加總。某樹種之磨角率越高,代 表梅花鹿越偏好選擇該樹種進行磨角。
四、幼齡稚樹族群變動率與梅花鹿取食偏好 之相關性分析
為探討墾丁高位珊瑚礁森林動態樣區內,
不同樹種幼齡稚樹(胸徑1~2 cm)族群量變動的 可能原因,本研究以該樣區在2001年幼齡稚樹
≥ 40株的26樹種,於2001~2013年12年期間的 族群變動率(Wang et al. 2004, Table 2)當應變 數(dependent variable),自變數(independent variable)則有三項。第一項自變數為梅花鹿對各 樹種葉片啃食率。第二項自變數為樹種的光合 潛力(photosynthetic capacity),代表各樹種的耐 陰性程度。第三項自變數為樹種在森林動態樣 區內群聚在隆起珊瑚礁礁岩的程度。此森林動 態樣區各樹種的空間分布類型,經分析得知上 述26樹種中有20樹種的個體會出現在隆起珊瑚
礁礁岩生育地(Wang et al. 2004),可計算礁岩集 中度數值。礁岩集中度計算方法為將供試某樹 種在動態樣區之隆起礁岩生育地的重要值指數 (IVI, Wang et al. 2004),除以同一樹種在森林動 態樣區共四類型生育地各自的重要值指數的加 總值(Table 2)。若該樹種僅分布在隆起礁岩生 育地,則數值為1;若完全沒有分布在隆起礁岩 生育地,則數值為0。本研究藉迴歸分析法,計 算供試樹種12年期間幼齡稚樹族群變動率,分 別與三項自變數間是否具顯著相關(p ≤ 0.05)。
結 果
梅花鹿對受測樹種葉片的取食偏好
墾 丁 高 位 珊 瑚 礁 森 林95種組成樹種中,
有85種在本研究進行梅花鹿食性測試,其中77 種有效測試次數達3次(含)以上(Table 1)。總共 測試944個枝條(次) 中,有505枝次屬於有效測 試,測試成功率約53%。在有效測試的77樹種 中,有65種會被取食(Table 1)。
將7 7 種 有 效 測 試 樹 種 的 平 均 啃 食 率 ( Ta b l e 1 ) 區 分 成 5 個 葉 片 適 口 性 等 級 , 適 口 性 第 一 級 , 平 均 啃 食 率 達1 0 0 % 的 樹 種 有 9 種,包括枯里珍(Antidesma pentandrum var.
barbatum)、青苧麻(Boehmeria nivea var.
tenacissima)、白榕(Ficus benjamina)、澀葉 榕(Ficus irisana)、稜果榕(Ficus septica)、咬 人 狗 、 血 桐 、 江 某(Schefflera octophylla)、
山 黃 麻(Trema orientalis)。適口性第二級,
平 均 啃 食 率 在9 9 ~ 5 0 % 的 樹 種 有 2 9 種 , 包 括 番 仔 林 投(Dracaena angustifolia)、白雞油 (Fraxinus griffithii)、大葉山欖(Palaquium formosanum)、台灣欒樹(Koelreuteria henryi)、
皮 孫 木(Pisonia umbellifera)、黃心柿、山 柚、無患子(Sapindus mukorossi)等。適口性 第 三 級 , 平 均 啃 食 率 在4 9 ~ 1 0 % 的 樹 種 有 2 0 種,包括鐵色、樹青(Planchonella obovata)、
茄苳(Bischofia javanica)、白匏子(Mallotus paniculatus)、九芎(Lagerstroemia subcostata) 及小梗木薑子(Litsea hypophaea)等。適口性 第四級,啃食率在9~1%的樹種有7種,包括毛
柿、小芽新木薑子(Neolitsea parvigemma)、軟 毛柿(Diospyros eriantha)、九節木(Psychotria rubra)、土樟(Cinnamomum reticulatum)等。
適 口 性 第 五 級 , 葉 片 完 全 未 被 啃 食 者 計 有12 種,包括紅柴、樹杞、海州常山(Clerodendrum trichotomum)、厚殼樹(Ehretia acuminata)、
梧 桐(Firmiana simplex)、白樹仔(Gelonium aequoreum)、石苓舅(Glycosmis citrifolia)、大 葉楠(Machilus japonica var. kusanoi)、台灣山 桂花(Maesa. perlaria var. formosana)、粗糠柴 (Mallotus philippensis)、台灣梭羅樹(Reevesia formosana)、假酸漿(Trichodesma calycosum) (Table 1)。
供 試7 7 樹 種 的 I v l e v ’s 選 擇 指 數 最 高 為 0.46,為啃食率100%的那9樹種(Table 1)。供 試樹種Ivlev’s選擇指數最低為-1.00,為啃食率 0%的那12樹種。各樹種平均啃食率的排序與 Ivlev’s選擇指數的排序一致(Table 1)。
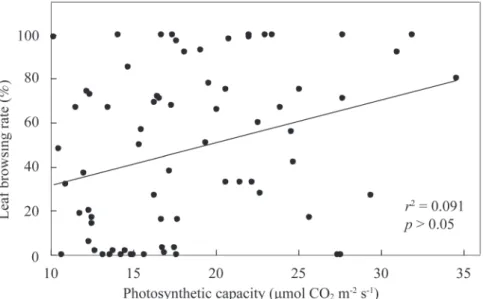
梅花鹿食性偏好測試的樹種中有72種曾測 定過光合潛力(Table 1),而此72樹種的光合潛 力與各樹種的啃食率並不具顯著相關(p > 0.05, r2 = 0.091) (Fig. 1),表示梅花鹿偏好取食的樹 種並不限於較不耐陰或較耐陰者,取食偏好與 樹種的耐陰性無關。
梅花鹿磨角行為及樹種偏好
2017年在植物園展示區內,共發現82樹 種 計1 8 5 株 林 木 遭 梅 花 鹿 磨 角 。 遭 磨 角 株 數
≥ 10株的樹種包括:穗花棋盤腳(Barringtonia racemosa) (16株)、毛土連翹(Hymenodictyon o r i x e n s e ) ( 1 4 株 ) 、 變 葉 木 ( C o d i a e u m variegatum) (13株);遭磨角株數≤2株的樹種高 達64種。因本項研究並未統計展示區內各樹種 可供磨角的總株數,因此上述數據並不足以顯 示梅花鹿摩角行為是否偏好某些特定樹種。
梅花鹿磨角行為對樹木造成的破壞樣式,
主要是在樹幹上造成帶狀磨痕,有176株(95%) 出現此類傷害;另有9株的磨角傷害樣式為戳刺 痕或細長劃痕。帶狀磨痕下緣離地高度在1~30 cm範圍者有47株,高度31~60 cm者有66株,高 度61~90 cm者有55株,高度範圍在91~120 cm 則僅8株。出現帶狀磨痕的176株其磨痕平均長 度為60 cm,以31~60及61~90 cm兩長度範圍的 株數較多,分別有70及62株。然而,磨損程度 達到木質部外露的嚴重磨痕,其平均長度為33 cm,以1~30及31~60 cm兩範圍的株數較多,分 別為69及91株。出現磨痕的林木胸徑以≤10 cm 的株數較多(62%),胸徑較大者遭鹿隻磨角的株 數較少(Fig. 2A)。樹木遭輕度、中度及重度環
Fig. 1. Relationships between the photosynthetic capacity and leaf browsing rate of 72 tree species in the Kenting Karst Forest.
剝的植株比例分別為47、27及26%。此三級傷 害程度的平均幹徑分別為20.0、5.7及4.3 cm,
遭重度環剝植株的平均幹徑最小,遭輕度環剝 植株的平均幹徑最大,顯示遭磨損程度會隨植 株徑級加大而減輕。在遭磨角總數185株的磨損 傷口中,判定為新傷口者有44%,判定為舊傷 口者則有43%,但有13%為舊傷口上再出現新 的磨痕,歸類為新舊混合傷口。
本 研 究 第 二 項 稚 樹 遭 鹿 隻 磨 角 傷 害 方 面 , 於2017、2018、2019三年期間分別調查 到133、355及223株,合計731株稚樹遭鹿隻 磨角,分屬40科97樹種,其中711株稚樹呈現 嚴重磨損。遭嚴重磨損植株的徑級以≤ 2.0 cm 者 居 多(59%),徑級2.1~4.0 cm 者佔25%,
8.1~10.0 cm較大徑級者株數僅為2% (Fig. 2B)。
新舊磨痕方面,有67%為新磨痕,舊磨痕只有 4%,但有29%的植株混合著新舊磨痕。遭磨 角稚樹株數≥ 20株的樹種,包括紅柴(94株)、
樹杞(61株)、春不老(Ardisia squamulosa) (42 株)、土楠(Cryptocarya concinna) (37株)、變 葉木(31株)、山柚(31株)、大葉楠(28株)、龍眼 (Euphoria longana) (20株)等。
在鹿隻磨角樹種選擇方面,2019年11~12 月調查到223次磨角事件中,遭磨角植株旁2 m 半徑內有形體類似,但卻未遭磨角的不同樹種 共出現41次事件,可視為為有效的對比事件。
其 中 可 供 有 效 對 比 的 樹 種 有25種,但只有紅
柴 、 樹 杞 及 毛 柿 此3樹種可供對比的次數≥10 次。紅柴遭磨角11次,未遭磨角2次;樹杞遭 磨角及未遭磨角各有6次;毛柿則22次均未遭 磨角。上述三樹種遭磨角機率分別為85、50、
0%,顯示紅柴為梅花鹿偏好磨角,毛柿為梅花 鹿避免磨角的樹種。
幼齡稚樹族群變動率與梅花鹿食性偏好之關係 於2001至2013年12年期間,墾丁高位珊 瑚 礁 森 林 動 態 樣 區 內26樹種之幼齡稚樹(胸徑 1~2 cm)的族群變動率,與梅花鹿對該樹種葉 片的啃食率間,具極顯著相關(p < 0.001),決 定係數(r2)為-0.524 (Fig. 3A)。梅花鹿偏好取 食的樹種,在此期間其幼齡稚樹的族群減少率 越高,族群偏向負成長。相反的,梅花鹿不喜 歡或避免取食的樹種,其幼齡稚樹的族群量降 低 程 度 較 低 , 甚 至 有 些 樹 種 幼 齡 稚 樹 族 群 量 有 增 加 的 表 現 , 例 如 樹 杞 、 毛 柿 、 石 苓 舅 、 樹青、裏白巴豆(Croton cascarilloides)、紅柴 及軟毛柿(Table 2)。此外,稚樹族群變動率與 樹種的光合潛力(耐陰性)間也呈顯著負相關(p
< 0.05,r2 = -0.236) (Fig. 3B)。然而,20種可 分佈在隆起礁岩生育地的樹種,其礁岩集中度 與其幼齡稚樹族群變動率(Table 2)間不具顯著 相關(p = 0.39),但上述樹種如果排除梅花鹿 不喜歡取食,葉片啃食率< 20% (Table 1)的樹 杞、毛柿、紅柴、軟毛柿、土楠、柿葉茶株萸
Fig. 2. The abundance of individual trees (a) and saplings (b) with different values of the diameter at breast height (DBH) which were used by Formosan Sika deer to fray their antlers in the Hengchun Botanical Garden. The value at the top of each bar represents the percentage.
(Gonocaryum calleryanum)等6樹種,則上述兩 因子間即具極顯著正相關(p < 0.01, r2 = 0.512, n
= 14) (Fig. 4)。此結果顯示,排除梅花鹿避開取 食的樹種後,族群空間分布越集中在隆起礁岩 生育地的樹種,則其幼齡稚樹族群減少率也越 低。
討 論
本研究梅花鹿葉片適口性的分級方式,採
用與Chen et al. (2009)在台北動物園測試圈養 梅花鹿食性偏好相同的分級法,將適口性依照 葉片啃食率由高至低分為1~5級。此方法採用目 視評估受測葉片被啃食的比例,估算法較為粗 略,會出現兩種植物葉片啃食率差距雖不大,
但會被區分在不同的適口性等級。因此,本研 究測試的植物若與Chen et al. (2009)調查之同種 植物之適口性等級前後僅差一級,則該植物視 為兩研究適口性分級結果一致。在兩研究都共 同測試的45種植物中,有31種(69%)的分級結
Fig. 3. Relationships of leaf browsing rate (a) and photosynthetic capacity (b) of 26 tree species with population change rate of juveniles in the Kenting Karst Forest. *: p ˂ 0.05; ***: p ˂ 0.001.
Apb: Antidesma pentandrum var. barbatum: Po: Planchonella obovata.
Fig. 4. Relationships of clumping on exposed limestone ratio of 14 tree species with population change rate of juveniles in the Kenting Karst Forest. ** p ˂ 0.01. Apb: Antidesma pentandrum var. barbatum; Po: Planchonella obovata.
果一致,其中有14種分級完全相同(Table 3)。
本研究葉片適口性較動物園測試結果晉一級(本 研究啃食率較大)的植物有9種,退一級者則有8 種,其餘14種的情況都是本研究植物適口性等 級較動物園測試者高,其中本研究適口性晉升 3級者,包括火筒樹(Leea guineensis)、台灣欒 樹 、 青 苧 麻 、 枯 里 珍 、 稜 果 榕 及 咬 人 狗 等6種 (Table 3)。Chen et al. (2009)也將社頂地區野外 梅花鹿啃食葉片的食痕出現率,在100~0%的 範圍中採用與上述相同的標準,區分適口性等
級。在與本研究共同測試的43種植物中,有36 種(84%)兩研究適口性分級結果一致,其中有 17種分級相同,本研究植物適口性晉一級有11 種,退一級則有8種(Table 3)。上述結果顯示梅 花鹿對不同種類的植物有明確的喜好或忌避。
Chen et al. (2009)也曾比較動物園圈養,以及野 外非圈養梅花鹿之食性偏好差異,發現35種共 同測試植物中有28種(80%)之適口性等級一致,
其餘7種均為野外梅花鹿的適口性高於動物園圈 養者。
Table 2. Population of juveniles of 26 tree species with a diameter at breast height of ≤ 2.0 cm in 2001 and 2013, population change rate, and ratio of population clumping on an exposed limestone habitat in the Kenting Karst Forest Dynamic Plot
Species Juvenile population Population Limestone 2001 2013 change rate (%) clumping ratio Ardisia sieboldii 491) 861) 761) 0.002)
Diospyros blancoi 161 232 44 0.43
Glycosmis citrifolia 104 133 28 N/A Planchonella obovata 107 128 20 1.00 Croton cascarilloides 41 44 7 N/A Aglaia formosana 808 752 -7 0.86 Diospyros eriantha 418 372 -11 0.29 Antidesma pentandrum var. barbatum 84 74 -12 1.00 Cryptocarya concinna 436 381 -13 0.00 Murraya exotica 76 59 -22 1.00 Liodendron formosanum 51 36 -29 N/A Drypetes littoralis 1128 715 -37 0.70 Beilschmiedia erythrophloia 258 156 -40 0.00 Palaquium formosanum 118 61 -48 0.75 Mallotus philippensis 99 50 -49 N/A Gonocaryum calleryanum 230 99 -57 1.00 Diospyros maritima 8501 3492 -59 0.48 Pisonia umbellifera 87 30 -66 0.13 Champereia manillana 256 85 -67 0.62 Leea guineensis 158 51 -68 0.00 Boehmeria wattersii 284 68 -76 N/A Dracaena angustifolia 278 41 -85 0.00 Dendrocnide meyeniana 568 81 -86 0.37 Ficus septica 107 14 -87 N/A Melanolepis multiglandulosa 938 21 -98 0.30 Macaranga tanarius 57 1 -98 0.27
1) Values provided by Wang et al. (2015).
2) Values provided by Wang et al. (2004).
Boulanger et al. (2009)調查法國兩處溫帶 森林,不同年度鹿隻啃食木本植物的選擇性,
發 現 鹿 隻 的 食 性 偏 好 在 不 同 年 度 , 或 兩 處 森 林 均 呈 現 一 致 性 , 顯 示 因 不 同 啃 食 壓 力 致 鹿 隻 食 物 組 成 種 類 或 數 量 的 差 異 , 不 會 顯 著 改
變鹿隻對取食植物種類的選擇樣式(selectivity pattern)。然而,與Chen et al. (2009)台北動物 園鹿隻餵食測試結果比較,本研究部分植物測 試的適口性分級差異較大,可能是因為動物園 只測試少數圈養個體,可能僅反映固定的少數
Table 3. Comparisons of palatability levels of the same tree species by Formosan Sika deer investigated by this study and by Chen et al. (2009) in the Taipei Zoo and the Sheding area
Palatability level Palatability level
Species This Taipei Sheding Species This Taipei Sheding study Zoo area study Zoo area Antidesma pentandrum 1 (100)1) 5 3 Ficus superba var. japonica 3 (33) 4 1 var. barbatum
Boehmeria nivea 1 (100) 5 3 Mallotus paniculatus 3 (27) 2 N/A var. tenacissima var. paniculatus
Dendrocnide meyeniana 1 (100) 4 4 Scolopia oldhamii 3 (38) N/A 2 Ficus benjamina 1 (100) 3 2 Diospyros ferrea 3 (37) 4 N/A Ficus irisana 1 (100) 4 3 Glochidion philippicum 3 (33) 2 2 Ficus septica 1 (100) 5 4 Drypetes littoralis 3 (48) 4 3 Macaranga tanarius 1 (100) 2 1 Syzygium formosanum 3 (32) 3 4 Schefflera octophylla 1 (100) N/A2) 5 Radermachera sinica 3 (42) 3 N/A Trema orientalis 1 (100) 1 N/A Bischofia javanica 3 (28) 3 2 Fraxinus insularis 2 (99) 4 N/A Lagerstroemia subcostata 3 (17) 3 2 Palaquium formosanum 2 (97) 4 2 Litsea hypophaea 3 (16) 3 4 Koelreuteria henryi 2 (93) 5 3 Diospyros blancoi 4 (6) 3 4 Melanolepis multiglandulosa 2 (92) 1 2 Neolitsea parvigemma 4 (2) 4 4 Melia azedarach 2 (80) 3 N/A Psychotria rubra 4 (2) 4 N/A Celtis formosana 2 (78) 2 3 Diospyros eriantha 4 (2) N/A 4 Diospyros maritima 2 (74) 1 2 Cinnamomum reticulatum 4 (1) 4 4 Champereia manillana 2 (73) 3 3 Aglaia formosana 5 (0) 4 5 Gardenia jasminoides 2 (72) 4 2 Ardisia sieboldii 5 (0) 4 N/A Eriobotrya deflexa 2 (71) 4 N/A Clerodendrum trichotomum 5 (0) 5 4 Premna serratifolia 2 (71) 3 2 Ehretia acuminata 5 (0) 5 5 Murraya exotica 2 (68) N/A 2 Ehretia resinosa 5 (0) N/A 4 Ficus microcarpa 2 (67) 3 2 Maesa perlaria 5 (0) 5 5
var. microcarpa var. formosana
Leea guineensis 2 (66) 5 3 Gelonium aequoreum 5 (0) 5 4
Kleinhovia hospita 2 (60) 4 3 Machilus japonica 5 (0) N/A 4
var. kusanoi
Fraxinus griffithii 2 (56) 4 3 Mallotus philippensis 5 (0) 3 3 Ficus virgata 2 (54) 3 N/A Trichodesma calycosum 5 (0) N/A 5 Sapindus mukorossi 2 (51) N/A 3
1) Values in parentheses are leaf browsing rates.
2) Denotes not measured.
個體之取食偏好或忌避。本研究食性測試地點 位於恆春熱帶植物園開放空間,該處自動照相 機夜間可拍攝到數隻至高達10隻梅花鹿在測試 地點活動情形。此外,梅花鹿在人工圈養環境 的食性偏好有可能與野外環境不同。
本研究餵食測試的枝條僅受到梅花鹿1~3 天短期的啃食壓力,單一受測枝條被梅花鹿造 訪取食的次數有限,餵食測試的結果可能存在 個體取食行為之差異。然而,野外天然更新的 苗 木 會 受 到 梅 花 鹿 長 期 的 啃 食 壓 力 , 食 物 量 較 為 缺 乏 的 季 節 取 食 壓 力 較 大 , 苗 木 被 啃 食 情形較為嚴重。另外,梅花鹿對未曾覓食的種 類,會少量取食後再決定是否繼續取食(Hu and Wang 1994),野外苗木若遭到鹿隻大量造訪,
即使是適口性不佳的種類,也會累積較高的被 啃食率。因此,梅花鹿整體族群對此森林地被 植群及幼齡木的啃食壓力應會比短期測試結果 更加嚴重。本研究葉片啃食率小於10%的樹種 中,有部分樹種如台灣梭羅樹、石苓舅及白樹 仔,現地觀察到其野外幼齡植株葉片被啃食比 例已超過50%。此外,毛柿在本研究之葉片啃 食率僅6%,但在鄰近的毛柿造林地,其野外樹 苗的葉片啃食率也已超過50%。自2017年起作 者觀察到此森林動態樣區內毛柿樹苗葉片出現 食痕的頻度也逐年加大。
短 期 食 性 測 試 之 主 要 目 的 在 比 較 不 同 種 植物葉片適口性高低。然而單位時間內若造訪 的鹿隻數量眾多,令測試樹種遭極大的啃食壓 力,則大部分樹種的葉片啃食率可能都會接近 100%,因此僅適口性較低的樹種間才能比較 其適口性高低;相反的,如樹種間啃食壓力均 很輕微,則較能測出原本適口性即高的各樹種 間葉片啃食率的差異。Motta (1996)認為在鹿 隻密度較高的條件下,有些樹種會持續遭強烈 的取食壓力,然而有些樹種則是要待鹿隻較偏 好樹種的葉片即將耗盡時,才會開始遭啃食。
因 此 , 本 研 究 測 得 不 同 樹 種 葉 片 適 口 性 的 排 序,可供指示墾丁高位珊瑚礁森林面臨鹿隻嚴 重啃食壓力時,各樹種幼齡植株消失的時間順 序。此外,仍有一些因素會影響食性測試的結 果,例如整體的食物量對梅花鹿族群是否充足
(Gerhardt et al. 2013)、季節性造成的整體食物 來源改變(Dumont et al. 2005)、葉片成熟程度 為老葉或是幼葉(Coley 1980)等。因此,若是以 短期食性測試的結果來推論天然植群中,各樹 種可能受到的啃食壓力高低,仍需更多面向的 考量。
葉 片 的 某 些 功 能 性 狀 曾 被 提 出 作 為 預 測 動物取食偏好的指標(Coley 1985, Grime 1996, Forsyth 2005)。葉壽命(leaf lifespan)為樹種重 要 的 功 能 性 狀 之 一 , 此 性 狀 與 樹 種 的 耐 陰 性 呈顯著正相關(Reich et al. 1999, Wright et al.
2005)。葉壽命較長的耐陰樹種,其葉片通常 有較高的結構性防禦功能(Wright and Cannon 2001),也會投資較多的化學防禦物質在葉片 (Coley and Barone 1996)。在低光環境下,幼 齡植株能固定的碳水化合物很有限,若葉片常 被 動 物 取 食 利 用 , 則 植 株 將 遭 嚴 重 傷 害 。 因 此,耐陰樹種幼齡植株的葉片常具適口性低的 特性(Hallik et al. 2009)。另一方面,葉壽命較 短,光合潛力較高的不耐陰或陽性樹種,其葉 片氮含量較高(Wright et al. 2005),但投資在 葉片的化學防禦物質較少,因此較容易受到植 食動物取食(Matsuki and Koike 2006)。然而,
分析墾丁高位珊瑚礁森林72樹種的光合潛力與 各樹種被梅花鹿取食的程度,卻發現這些樹種 光合潛力及由此代表的耐陰性,與梅花鹿取食 偏好並不具顯著相關。此森林有些陽性樹種葉 片大幅度被梅花鹿取食,例如山黃麻、血桐、
白榕等,但也有部分陽性樹種如厚殼樹及梧桐 則完全不被取食;另一方面,毛柿、台灣梭羅 樹、石苓舅等光合潛力較低,耐陰程度較高的 樹種,的確不為梅花鹿喜食,但梅花鹿卻也偏 好取食某些較耐陰的樹種,例如枯里珍、黃心 柿、山柚等(Table 4)。因此,墾丁高位珊瑚礁 森林樹種的耐陰性並不能供作梅花鹿對樹種葉 片適口性的指標。
綜合本研究食性測試結果及現地觀察,推 測墾丁高位珊瑚礁森林適口性較低的樹種可能 具有幾方面的特性,包括(1)具毒性或複雜的次 階代謝物質,例如裏白巴豆(Xu 2018)、台灣土 沉香(Excoecaria formosana) (Kumar 2013)、
紅柴(Paritala 2015);(2)具有特殊的味道,例 如海州常山、珊瑚樹(Viburnum odoratissimum) 及樟科(Lauraceae)的樹種;(3)特定的分類群,
例如紫金牛科(Myrsinaceae)的樹杞、小葉樹杞 (Ardisia quinquegona)及台灣山桂花。Lin et al.
(2017)也發現有多重因素會影響臺灣山羌的取 食偏好。葉片木質素成分比例的高低,可以解 釋臺灣山羌對受測樹種葉片的取食程度,但有 一些樹種則為例外。葉片質地及富含揮發性精 油的葉片也可能會影響臺灣山羌的取食偏好(Lin et al. 2017)。另外,該研究測試顯示紫金牛科 的樹杞及台灣山桂花均屬於臺灣山羌高取食率 的樹種,而本研究結果則顯示梅花鹿不喜歡取 食樹杞及台灣山桂花葉片。
鹿科動物每年定期落角,新生的茸角在硬 化過程中,外層茸皮會壞死,因此鹿隻會藉由 磨角行為來清除茸角上壞死的茸皮,藉以提升 舒適感(Geist 1998)。此外,藉由磨角製造樹幹 上的視覺(磨痕)及氣味信號,可在發情期向其 它雄鹿傳遞領域性訊息,達到展示及溝通之目 的(Massei and Bowyer 1999),磨痕及氣味也可 能具有後續向雌鹿溝通的作用(Johansson and Liberg 1996)。本研究假設空間位置為鹿隻選 擇磨角植株的決定因子,或是當誘發磨角行為 的情境出現時,鹿隻才會就近選擇一株適合的 樹木去磨角。假設鹿隻進行磨角行為時只會利 用就近方便的植株,那麼選擇行為發生的空間 尺度應不大。本研究針對遭鹿隻磨角之植株,
其半徑2 m內徑級相近的不同樹種進行兩兩對 比,區別磨角行為是否具樹種偏好,結果發現 紅柴遭梅花鹿磨角的比例高達85%。Chen and Wang (1999)於鄰近的社頂地區也調查到紅柴遭 磨角比例為15%,此數值高於該樹種株數佔調 查區域總株數的比例(3.2%),且紅柴遭磨角的 Ivlev’s指數為0.66,顯示梅花鹿對選擇紅柴來磨 角具有正向偏好,此發現與本研究調查結果一 致。相反的,Wang et al. (2019)在墾丁高位珊 瑚礁森林動態樣區調查到紅柴遭梅花鹿磨角比 例僅2.0%,低於此樹種背景株數比例的4.6%,
且紅柴遭磨角的Ivlev’s指數低至-0.40,顯示梅 花鹿避免選擇紅柴進行磨角。然而,Wang et al.
(2019)調查對象多為較大徑級林木,而本研究 僅分析胸徑≤ 10 cm的稚樹,可能因此造成兩研 究梅花鹿利用紅柴來磨角的偏好傾向不一致。
此外,本研究發現毛柿為梅花鹿避免選用來磨 角的樹種,此結果與Wang et al. (2019)的調查 結果一致。Gill (1992)認為鹿科動物選擇特定 樹種磨角,是因為該樹種所具備的物理特性;
Tompson (1969)報導麆鹿(roe deer, Capreolus capreolus)喜好選用具彈性、單幹無分枝的稚樹 (sapling-size)來磨角;Larner (1977)認為梅花 鹿似乎喜好胸徑大於30 cm而樹皮平滑的植株磨 角,選擇紅豆杉(yew)、花楸(rowan)更勝於橡 樹(oak)。毛柿具有粗糙硬質的樹皮,又為耐陰 樹種不易天然修枝,枝下高極低,常具有低處 側枝,似乎符合上列文獻歸納的規則。此外,
毛柿的側枝與主幹通常呈90度夾角,似乎比其 它側枝斜上生長的樹種更容易卡住鹿角分叉,
可能會阻礙梅花鹿磨角過程的順暢度。墾丁國 家公園梅花鹿復育區蔡木生先生認為,梅花鹿 偏好選擇樹幹較具彈性的小樹去磨角,例如紅 柴、過山香(Wang et al. 2019)。本研究調查時 也發現紅柴常有幹徑小於0.5 cm的幼齡植株遭 磨角,與上述報導吻合。推測梅花鹿磨角時採 取身體長軸正對樹幹的方向,上下擺動頭部讓 鹿角磨擦樹幹,鹿角上磨擦位置移動的軌跡呈 弧形。如果的彈性不足,樹幹就無法對鹿角施 加足夠的反作用力;但若樹幹太硬,則鹿角移 動的弧形軌跡可能就會受阻,梅花鹿頭部擺動 的幅度無法達到預期的距離。
鹿 隻 對 磨 角 樹 木 的 選 擇 , 會 受 到 植 株 大 小 、 灌 叢 或 藤 蔓 的 干 擾 等 因 素 影 響( R a m o s 2006)。對梅花鹿而言,選用不同樹種植株來磨 角,可能對其內在感受差別不大,不至於出現 嚴重的負面反應,也不至於妨礙磨角的主要功 能,因此樹種間的被取代性較高。相對的,食 物品質良窳與其內在感受及生存有關,有毒或 次階代謝物含量高的植物,可能會造成身體不 適,或出現嚴重的負面感受。因此,食物品質 與可食性,對動物的取食選擇應更具影響力。
此外,鹿隻一天中耗費較長的時間進行覓食,
但 用 較 少 的 時 間 進 行 磨 角(Braza 1986, Chen