一、前言
植物體的發育從不具有生殖能力的幼年期轉變為具有生殖能力的成熟時期,如此 逐漸改變的過程可以稱之為相的轉變(phase change),這樣的改變在草本植物可能只 需幾個星期或幾個月,在木本植物可能是需經過多年的轉變。在發育生物學的領域,
植物被歸為後胚胎植物,可經由一群未分化且具有潛力分化成側生器官之頂端分生組 織(SAM)來調控這些相的轉變。相的轉變可以由植物荷爾蒙吉貝素(gibberellin, GA3, Rogler and Hackett, 1975)或細胞分裂素(cytokinin)(Bouriquet et al., 1985)的刺激、
枝條連續性的培養在含有細胞分裂素的培養基中(Brand and Lineberger, 1992;Fouret et al., 1885;Walker, 1986;)、頂端分生組織的培養(Bon and Monteuuis, 1991),以及 在體內(in vivo)(Muzik and Cruzado, 1958)或體外(in vitro)(Ewald and Kretzschmar, 1996;Huang et al., 1992a, 1992b;Revilla et al., 1996)培養下,將老樹頂端枝條嫁接 至年輕砧木(rootstocks)等方式都可以取得植物相的轉變或稱年輕化(rejuvenation)
的轉變。
相的轉變在針葉樹木本植物紅杉之研究,即是以嫁接的方式來達到。從 1976 年 開始,以老樹枝條頂端經五次連續性嫁接至年輕種子苗後,不論在外部型態(如: 葉 的型態、葉開展的方式及葉的顏色等)或生長勢(如:發根能力、長根的數目、莖延 長的能力及長側枝的數目等)上,以漸進式的轉換呈現與年輕樹一致之年輕化的苗,
並以生長素GA3 或 BA 測試,並非這二個生長素的存在而導致相的轉換(Huang et al., 1992)。因嫁接而使老樹年輕化的植株,簡稱為 Rejuvenated Sequoia(RS)。經由五次 嫁接而產生與年輕樹完全一致的生長勢,是什麼因子誘使老樹年輕化,尚不明瞭。目 前推測粒線體的基因可能參與在這樣的一個轉變過程(Huang et al., 1995;Huang et al., 2003)。由 Esser 等人在 1980 提出,真菌 Podospora anserine 在老化時,粒腺體質體 DNA 會伴隨著改變;而 Yen 等人在 1991 年也提出,人類肝臟細胞粒腺體 DNA 結合 年齡的增長約有5Kb 大小的片段會被刪除,由此證據更深一層驗證粒腺體 DNA 可能 參與紅杉相的轉變。
一些研究顯示,蛋白質酪氨酸磷酸化(protein tyrosine phosphorylation)(Huang et
al., 2003)或一些生化的標記,如同功異構酶 isoesterase 及 isoperoxidases(Brand et al., 1992;Huang et al., 1996),也都可以藉由磷酸化形式的不同作為相轉變的研究(Kuo et al., 1995)。在生理方面的研究,前已論及既非植物荷爾蒙GA3也非BA 存在而導致老 樹年輕化,而且已知處理GA 或 cytokinin 會誘導植物年輕化,而處理離層酸會使植物 停留在年老的時期,因此,Huang 等人於 2003 年也試著去分析另一植物荷爾蒙乙烯 是否在相的轉變中扮演角色。結果顯示,乙烯生成量與生長勢強弱成正比,但是,乙 烯產生量之高低是否影響植物生長快慢,則不能直接證明(Huang et al., 2000)。若以 光合作用的的角度去探測相之間的差異,不論在光合作用及呼吸作用率、含氮量及葉
綠素 a 含量,幼年莖皆比成齡老株來得高,但在光合系統的電子傳遞效率並無差異
(Huang et al., 2003)。
由以上看來,在生理層次的分析,都僅是一種現象而已,若真要了解隱含的機制,
仍須以生化或分子生物學的角度去深入了解相轉變的過程。蛋白質酪氨酸磷酸化扮演 一個關鍵的角色在調節細胞的機制(Cans et al., 2000),專一性在蛋白質酪氨酸殘基上 進行磷酸化可以闡明此蛋白質的功能,而這一類的蛋白質在訊息傳遞路徑對外在環 境,例如細胞週期的調控或細胞的生長與發育扮演一個重要的角色。例如,蛋白質酪 氨酸磷酸化形式的不同參與在胚的形成已在蘿蔔(carrot)(Barizza et al., 1999)及可 可果(coconut)(Islas-Flores et al., 1998)上被研究,因此,Huang 等人於 2003 年,
證明植物體伴隨著年齡的增長在蛋白質酪氨酸磷酸化上可能扮演一重要的角色。
在動物系統研究上,Dr. Holbrook 之研究室利用從年輕老鼠(4-6 month)及老齡 老鼠(24-32 month)來研究,發現肝的再生能力及肝細胞受生長因子刺激後之分裂潛 力會隨著年齡的生長而逐漸降低。在他們的實驗中發現 MAPK 的活性會隨著年齡生 長而遞減(Liu et al., 1996)。最近他們發現到以H2O2處理細胞將造成從老的鼠取出之 細胞較年輕鼠取出之細胞容易死。並且H2O2所誘導之MAPK 活性在老的鼠取出之細 胞較弱 (Ikeyama et al., 2002)。而 Dr. Majumdar 之研究則指出老鼠胃黏膜受傷後之再 生能力,將隨年齡生長而遞減,同時,酪胺酸蛋白質激酶(Tyrosine Kinase)的活性也隨 著年齡生長而有差別,因此我們可以了解生物將隨著年齡增長,逐漸喪失細胞內某些
訊息傳遞能力,導致受傷後之再生修補能力降低 (Fligiel et al., 1994) ,作者也觀察到 老化將造成ERKs(p42/p44)和 JNK1 的活性增高。然而 p38 MAPK 之活性則會降低 (Xiao et al., 2000)。
訊息傳遞調節細胞對外在或內在刺激的反應過程決定了所有生物的生長及發育
(Luan et al., 2001)。這些訊息傳遞包含細胞的分裂與分化、細胞週期的調節、代謝、
對植物荷爾蒙的反應、以及細胞對生物與非生物逆境刺激的反應。這些反應的進行主 要是藉由蛋白質磷酸化來快速放大細胞外的刺激物所要傳遞的訊息。MAP kinase cascade 便是其中一個特殊的訊號傳遞機制,從酵母菌、經由黏菌、果蠅、線蟲至哺 乳動物,甚至植物,在這些真核生物中扮演一個重要的角色。而近來的研究結果更加 突顯訊息傳遞途徑的重要性和複雜度,尤以模式生物—酵母菌及哺乳動物對 MAP kinase 路徑的研究最為徹底與最好了解;研究訊息傳遞在植物領域至今也有一些文獻 的報導(Hirt, 1997;Hirt, 2000;Ichimura et al., 2000a;Jonak et al., 1994; Meskiene &
Hirt, 2000;Mizoguchi et al., 1997),但其廣及性及透徹性卻不及動物的領域。
MAPK cascades 主要由三個激酶所組成,分別為最上游的 MAPK Kinase Kinases
(MAPKKKs)、中游的 MAPK Kinases(MAPKKs)及下游的 MAP kinases(MAPKs),
其間作用模式為一個磷酸化連級(phosphorylation cascade)的作用模式,藉由磷酸化 的作用互相調控其活性,在植物研究領域上歸屬於轉譯後(post-translational)具有活 性之蛋白質激酶。當外界的逆境訊息被細胞膜上的受器(receptor)接收後,細胞會 啟動與細胞膜相結合之蛋白質,如small G-protein(如 Rho 1)或 protein kinase C 等,
經一連串的反應後,將訊號以蛋白質磷酸化的型式傳遞給MAPKKKs,使得 MAPKKKs 因自我磷酸化而具有活性,被活化之 MAPKKKs 再去活化其下游 MAPKKs 上 S/TXXXXXS/T motif(在哺乳動物及酵母菌為 SXXXS/T),將serine 及 serine/threonine 殘基加以磷酸化而使其具有活性;被活化的MAPKKs 又會在其下游的 MAPKs 之 TXY motif 上,將 threonine 及 tyrosine 殘基磷酸化而活化 MAPKs(Mizoguchi et al., 1997)。
MAPKs 在植物中的研究至今已超過近十年了,舉凡從紫花苜蓿(Duerr et al., 1993)、阿拉伯芥(Mizoguchi et al., 1993)、碗豆(Stafstrom et al., 1993)、菸草(Wilson
et al., 1993)、矮牽牛(Decroocq-Ferrant et al., 1995)至芫荽(Ligterink et al., 1997)等 這些雙子葉植物都被發現有 MAPKs 基因存在,並分別扮演與植物生長及發育的調 控。將植物這些 MAPKs 與動物作序列分析比對,發現與典型動物 MAPKs(ERK)
具有高的同源性,分子量大約都為42 kDa 左右;在分類上,依其氨基酸序列同源性,
可分成四大族群,除了D 族群受上游磷酸化磷酸化位點為 TDY motif 外,其餘 A、B 及C 族群的磷酸化位點都為 TEY motif。
我們在過去實驗中利用試管內(in vitro)磷酸化技術發現到有一 32kDa 的磷酸化 蛋白質僅表現在老樹,而一31kDa 的磷酸化蛋白質則只在年輕樹中表現 (kuo et al., 1995),最近本實驗室更偵測到酪胺酸磷酸化蛋白質會隨著植物生長相轉變而有變化 (Huang et al, 2003)。因此我們認為訊息傳遞蛋白質必在年輕化上扮演一定之角色。若 以動物系統研究之結果參考,我們認為或許老樹喪失長根能力,可能是因為部份訊息 傳遞蛋白在植物成熟過程改變了。
由先期結果已指出植物生長相改變將會調控一個蛋白質激酶之基因表現,因此,
我們以此為基礎一方面深入了解此一蛋白質激酶,並將找尋其它受生長相轉變而調控 的訊息傳遞蛋白質。因此,本實驗的目的:
1、利用 RACE(5’-and 3’-rapid amplification of cDNA ends)的方法得到受生長相轉變 調控的蛋白質激酶基因的全長。
2、研究此蛋白質激酶在相轉變植株中基因的表現情形。
3、找尋其它受生長相調控的訊息傳遞蛋白質激酶。
我們將以此基因為出發點對此類基因群進行研究,將可增進對植物發育過程的了 解。另外,因為研究目標為訊息傳遞分子,從研究結果亦可了解植物如何隨著發育時 期的轉變而改變其對環境刺激的反應。雖然目前在植物訊息傳遞方面的研究進展相當 快速,但是在生長相之間的差異卻付之闕如,而不同的生長相對相同環境刺激的差異 反應,卻是存在的事實。因此,我們將從訊息傳遞的角度對紅杉在年輕與年老植株中 差異表現之訊息傳遞基因與蛋白質進行深入的探討,以提供對植物生長相轉變機制有 更深入的了解。
二、材料與方法
1、紅杉組培苗之培養(Huang et al., 1992)
2、紅杉組培苗五次稼接過程(Huang et al., 1992)
3、紅杉基因選殖
(1)退化性引子(degenerate primers)之設計及製備
(2)RNA 的萃取(RNeasy Plant Mini Kits, QIAGEN, Hilden, Germany)
(3)反轉錄聚合酶鏈鎖反應(RT-PCR, Acess RT-PCR Kit, Promega, USA)
(4)瓊脂膠體回收 DNA 片段
(5)黏合反應(Ligation reaction, pGEM-T Easy Vector, Promega, USA)
(6)DNA 轉型(Transformation)
(7)萃取質體 DNA
(8)分析質體 DNA 樣本及 DNA 定序
4、快速增幅 cDNA 末端(RACE, 5’-and 3’-rapid amplification of cDNA ends; BD Biosciences Clontech, USA)
5、南方墨點轉漬法(Sorthern blot analysis; Sambrook et al., 1989)
三、結果
1. 紅杉 SsMAPKA 基因之選殖及序列分析
搜尋資料庫中植物之 MAPK 序列,利用共有的保留性序列設計退化性引子 (degenerate primers),並分別萃取紅杉老樹與年輕樹組培苗頂端~1.5cm 之總體 RNA 為 模板,進行RT-PCR 基因選殖(RT-PCR cloning),可偵測到符合預測大小之 cDNA 片段 (圖片未顯示),經載體黏合、E. coli 轉型篩選及定序等步驟,在老樹與年輕樹共選殖 出 1 個似 MAPK 基因之部分片段,經由資料庫比對結果顯示應屬於 Group A2 之 MAPK。
將此一紅杉 Group A2 之 MAPK 命名為 SsMAPKA,並利用 RACE-PCR (5’-and
3’-rapid amplification of cDNA ends)取得基因全長共 1843bp,其中 5’端非譯碼區為 147bp,3’端非譯碼區為 526bp,譯碼區共 1170bp,可轉譯出 389 個氨基酸(圖 1),與 紫花苜蓿之 MsSIMK 大小相同,顯示此一基因之大小在不同物種間具有相當高度保留 性。
此SsMAPKA 具有 11 個典型 Ser/Thr 激酶所共有的激酶區域,並於第Ⅶ及Ⅷ激酶 區之間具有可被上游MEK 所磷酸化之 TEY motif,亦為 Group A MAPK 之重要活化 位點。將SsMAPKA 之氨基酸序列與同類基因家族成員(Group A2 MAPK)進行排序比 對,發現這些MAPK 在各個激酶區均具有高度保留之氨基酸結構(圖 2)。利用分子演 化分析程式,構築 MAPK 基因族系之氨基酸親緣演化樹(圖 3),分析結果顯示 SsMAPKA 確實屬於 Group A2 之 MAPK。其氨基酸序列與 Group A2 之 MAPK 具有 78%以上之相似度,其中與芫荽 PsD5 具有 82%之相同度、與水稻 OsSIPK 具有 79%
之相似度。而Group A2 之 MAPK 如 ZmMPK5、OsSIPK、AtMPK6、NtNTF4 及 NtSIPK 均已被廣泛報導,主要與多種生物性及非生物性逆境之訊息傳遞有關,且其調控方式 兼具有mRNA 轉錄及轉譯後修飾等作用,其中水稻 OsSIPK,為 Group 2 的新成員(未 發表),相似於菸草NtSIPK,受生物性及逆境而活化。另一單子葉植物玉米 ZmMPK5,
在葉子老化過程中而活化之一個蛋白激酶,顯示植物體在老化後,這些蛋白激酶也參 與訊號調控(Berberich et al., 1999)。
值得注意的是,Group A1 與 Group A2 之 MAPK 彼此之間的氨基酸同源度相當 高,顯示其所扮演的生化角色可能非常接近,前人研究指出AtMPK6 與 NtSIPK 之激 酶活性,亦會受到多種生物性與非生物性逆境之短暫活化(Kovtun et al., 2000),但 Group A1 與 Group A2 之 MAPK 最大的差別,在於兩者受活化後的調控方式不同,
Group A2 MAPK 之 mRNA 並不會受到環境訊號之刺激而有所變化,其調控方式主要 透過磷酸化與否,亦即偏重於轉譯後修飾作用的方式,因此,我們也曾以In-gel kinase activity 分析及以抗體 ERK1(anti-ERK1)及磷酸化在酪氨酸 202 及 204 殘基上之 ERK
(anti-phosphoERK)蛋白質電泳免疫法,試著去分析 SsMAPKA 是否在創傷後
(wounding)具有活性的變化。
2. 紅杉 SsMAPKA 基因族系組成
將紅杉基因組 DNA 以特定酵素切割並由電泳分離後,進行南方墨點轉印
(southern blotting)實驗,再以 SsMAPKA 基因之 3’端非譯碼區探針進行雜合(圖4),
發現以限制酵素切割後都只出現了一條雜合反應條帶,故推測 SsMAPKA 基因在紅杉 基因組中為單一拷貝(single copy)基因。
3. 紅杉 SsMAPKA 之基因表現情形探討
以 SsMAPKA 基因之 3’端非譯碼區探針,透過北方墨點分析結果,SsMAPKA 基因 分別在紅杉年輕樹與老樹頂端(apical)與基部(basal)均出現單一雜合反應條帶(圖 5) ; 以 在 老 樹 基 部 之 SsMAPKA 表 現 量 略 高 於 年 輕 樹 基 部 之 基 礎 轉 錄 量 (basal transcription level),符合最初 RT-PCR 基因選殖過程中之表現情形,由此結果顯示 SsMAPKA 具有 age-related 表現的差異。以紅杉核醣體 RNA (rRNA)之表現作為內控制
組(internal control),可看出每一處理均有一致的表現量,顯示每一處理所使用之 RNA 量均相同,故不同部位表現上的差異,均可直接反應在 SsMAPKA 基因之表現上。
由 SsMAPKA 基因在紅杉老樹與年輕樹頂端(apical)與基部(basal)表現上的差 異,我們也常式著去看 SsMAPKA 基因在五次嫁接植株的表現分析。分別抽取老樹、
1x~5x 嫁接的植株及年輕樹頂端及基部總體 RNA,以 SsMAPKA 基因之 3’端非譯碼區 為探針,進行北方墨點分析,由結果顯示,SsMAPKA 基因在不同植株均出現單一雜 合反應條帶(圖 6),且以基部表現量高於頂端的表現量,若以個體間相比較,不管是 在頂端或基部,老樹及1x~3x 嫁接的植株的表現量均高於 4x~5x 嫁接的植株及年輕樹 的表現量。同樣地以紅杉核醣體RNA (rRNA)之表現作為內控制組(internal control),
每一植株均有一致的表現量,顯示每一植株所使用之RNA 量均相同。
4. 紅杉 SsMAPKA 之融合蛋白分析
MAPK需透過轉譯後修飾作用來調控自身的活性,其TXY motif上之tyrosine及
threonine殘基的磷酸化被認為是MAPK具備活性的指標(Charest et al., 1993)。前人研究 指 出 , 將MAPK 與GST (glutathione S-transferase)連 接,使 之表 現 於E.coli中 產生 GST-MAPK融合蛋白,會使得GST-MAPK融合蛋白於tyrosine上發生自動磷酸化 (autophosphorylation),而自動磷酸化後的MAPK即具有激酶活性(Wilson et al., 1995)。本實驗亦利用GST-SsMAPKA融合蛋白,並以anti-phosphotyrosine抗體(RC-20) 進行蛋白質電泳免疫轉印法,偵測SsMAPKA是否發生自動磷酸化而偵測其蛋白活性。
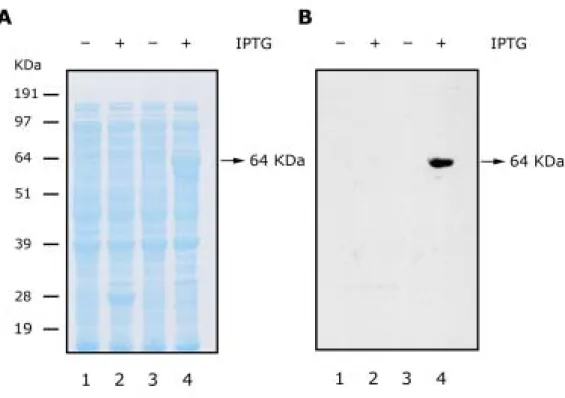
圖7A結果顯示,將GST-SsMAPKA融合蛋白於E. coli之BL21(DE3)品系中表現,
並以IPTG (Isopropylthio-β-galactoside)誘導GST-SsMAPKA融合蛋白之大量表現,發現 在 IPTG 誘 導 後 , 僅 有 GST-SsMAPKA 融 合 蛋 白 會 被 大 量 表 現 ; 而 若 以 anti-phosphotyrosine抗體偵測融合蛋白tyrosine之磷酸化,亦顯示僅有GST-SsMAPKA 之融合蛋白tyrosine會被磷酸化 (圖7B)。由前人研究也顯示,tyrosine若被磷酸化程度 愈高,其磷酸化下游受質之能力愈強,亦即其激酶活性愈高(Wilson et al., 1995),故推 測SsMAPKA是一個具有激酶活性之蛋白。
四、討論
可逆轉蛋白質的磷酸化(reversible protein phosphorylation)對生物體活性的調節 提供了一個多方面及精確的調控,因此,蛋白質磷酸化的修飾作用調節生物體的活性 也許是一種重要的形式。例如,約有2000 個這一類的蛋白質激酶的基因發生在哺乳 動物的細胞(Hunter et al., 1995)。MAPKs 即是一群需經由後轉譯修飾
(post-translational modification)作用而具有活性的蛋白質激酶,屬於一群保留性很 高的Ser/Thr 的蛋白質激酶,廣泛存在酵母菌至哺乳動物等真核生物中。其特性為具 有11 個保留性的區域,其中第Ⅶ及Ⅷ催化區具有 TXY motif 接受上游蛋白質激酶磷 酸化的磷酸化位點。哺乳動物MAPKs 首度的發現即是以上皮生長因子刺激細胞,進 而調節細胞的去分化及細胞週期的調控,因而將其命名為受外在刺激而活化之蛋白質 激酶,稱之為ERKs(extracellular signal-regulated kinases),也是現今我們所稱的典型 MAPKs。現今,一些研究顯示,MAPKs 也參與在植物的生長,例如雙子葉模式植物
阿拉伯芥約有20 個 MAPKs 參與在各種生物性、非生物性、荷爾蒙的調節、細胞的 分裂及發育等過程(Ichimura et al., 2002),。
蛋白質激酶參與在木本植物的研究微乎其微,至今也只有紅杉(Sequoia
semphermius)(Huang et al., 2003;kuo et al., 1995;Huttyl and Phillips, 1995)、白楊樹
(Populus nigra var. italica)(Nishiguchi et al., 2002)及蘋果樹(Malus x
bomesticaBorkh.)(Komjanc et al., 1999)有關於這方面的探討。以訊息傳遞路徑方式 在木本植物上的研究在白楊樹方面,Nishiguchi 等人在 2002 年找到一個與受器相似的 蛋白質激酶(receptor-like protein kinase, PnLPK),擴及的層面也僅只於基因的表現及 磷酸化活性的分析,並提出在創傷處理下,PnLPK 基因在年輕葉的表現會增加。在蘋 果樹則是探討一個與受器相似的蛋白質激酶編碼有leucine-rich repeat(LRPKm1)的 序列,已知基因若譯有這樣的一段序列可以作為植物在遭到病原菌的侵襲下,接受病 原菌的訊號以啟動防禦的機制,同時,並對此基因作了一些特性的分析。然而,關於 基因的表現在植物成熟後或相轉變的研究上僅知一點,例如常春藤在老樹與年輕樹二 個時期,在枝頂端RNA 的表現並沒有量上的不同(Rogler and Dahmus, 1974);而Woo 等人在1994 年在常春藤相轉變的研究上也是以長不定根及光合作用的能力作為老樹 與年輕樹在相轉換上的判斷依據,在組織特異性基因表現的分析上顯示,HW101 基 因在HW101 的二個基因,分別為 chlorophyll a/b binding protein(HW101)及 proline rich protein(HW103),的基因在針葉樹落葉松探討一個與 light-harvesting chlorophyll a/b binding protein 同源的基因在年輕樹有 30%的表現量高於老樹的表現(Hutchison et al., 1990)
本實驗室在過去實驗中利用試管內(in vitro)磷酸化技術,發現有一 32KDa 的 磷酸化蛋白質僅表現在老樹,一31KDa 的磷酸化蛋白質則只在年輕樹中表現(kuo et al., 1995);再者 Huang 等人(2003)由蛋白質酪氨酸磷酸化(protein tyrosine
phosphorylation)的實驗指示,植物體將隨著年齡的增長(age-related)及生長相的轉 變(phase-related),在蛋白質酪氨酸磷酸化上的程度大不相同。例如,有較多 36、44 及46 KDa 及較少 29 KDa 大小的蛋白質在較年輕的組織,較多 34 及 36 KDa 蛋白質
在年老的組織;若以蛋白酪氨酸磷酸化抗體偵測,有較多39 及 54 KDa 大小的蛋白質 在較年輕的組織,較多25 及 34 KDa 蛋白質在年老的組織。因此我們認為訊息傳遞蛋 白質必在年輕化上扮演一定之角色。由此角度,我們嘗試著去克隆(cloned)這些存 在生長相轉變間的不同蛋白質激酶,並以典型的MAPKs 這一類為我們的目標。
經搜尋資料庫中植物之 group A1 MAPK 序列,利用共有的保留性序列設計退化 性引子(degenerate primers),得到一個分子量大小約為 44KDa 的 SIPK,並將其命名為 SsMAPKA。SIPK(salicylic acid-induced protein kinase)現已廣泛被研究參與在各種 環境的逆境,且根據一些研究顯示,水楊酸(SA)可以做為一個內生性的訊號,活 化一些防禦基因的表現,如PR(pathogenesis-related)proteins 的合成(Rayls et al., 1994, 1996;Dempsey and Klessig, 1995),其他尚有如感受病源菌所釋放的誘發因子
(elicitors)、菸草在受到鑲嵌病毒(TMV)的感染後、植物體受到機械性的傷害後以 及在高滲透的逆境下均會活化SIPK 訊息傳遞路徑,藉以調控不同逆境下的防禦機制
(Zhang and Klessig, 2000)。
已知Group A1 及 A2 在功能上是很相近的一群 MAPKs。Zhang 及 Klessig 在 2000 年提出,在菸草中的二個MAPK,分別為 SIPK(屬於 Group A2)及 WIPK(屬於 Group A1),結合不同病源菌的刺激及各種逆境,其基因的表現及酵素活性皆會受影響而改 變,並且提出SIPK 同時也會在創傷的情形下,酵素的活性會提升,並提出酵素活性 的改變可能參與很多訊息傳遞路徑。SIPK 的活性廣泛的被認為是在酪氨酸及酥胺酸/
絲氨酸殘基上進行後轉譯修飾作用後,才呈現出酵素的活性;而WIPK 酵素活性改變 不僅只有經由後轉譯修飾作用,其mRNA 的基因表現層次也會提升,並且重新合成 WIPK 的蛋白。故當給於相同的刺激時,會優先啟動 SIPK 的活性,而 WIPK 的活化 尚需進行轉譯、轉錄後及蛋白質的重新合成,所以,在酵素活性的表現上會比SIPK 還要慢(Zhang and Klessig, 2000)。而由我們的實驗結果顯示,紅杉SIPK 基因之表現 在老樹較多,我們認為成熟植株中存在有如此多的SIPK,可幫助木本植物對抗病原 菌的感染。
五、參考文獻
1、Barizza E., Lo Schiavo F., Terzi M., and Filippini F.(1999)Evidence suggesting protein tyrosine phosphorylation in plants depend on the developmental conditions.
FEBS Lett. 447: 191-194
2、Bon M.C. and Monteuuis O.(1991)Rejuvenation of a 100-year-old Sequoiadendeon giganteum through in vitro meristem culture. II. Biochemical arguments. Physiol. Plant.
81: 116-120
3、Bouriquet R., Tsogas M., and Blaselle A.(1985)Essais de rajeunissement de l’epicea par les cytokinines. Ann. AFOCEL 1984: 173-185
4、Brand M.H. and Lineberger R.D.(1992)In vitro rejuvenation of Betula (Betulaceae):
Biochemical evaluation. Amer. J. Bot. 79: 626-635
5、Cans C., Mangano R., Barila D., Neubauer G., and Superti-Furga G.(2000)Nuclear tyrosine phosphorylation:The beginning of a map. Biochem Pharmacol. 60: 1203-15 6、Charest D.L., Mordret G., Harder K.W., Jirik F., and Pelech S.L. (1993) Molecular
cloning, expression and characterization of the human mitogen-activated protein kinase p44erk1. Mol. Cell. Biol. 13: 4679-4690
7、Decroocq-Ferrant V., Decroocq S., vanWent J., Schmidt E., and Kreis M.(1995)A homology of the MAP/ERK family of protein kinase gene is expressed in vegetative and in female reproductive organs of Petunia hybrda. Plant Mol. Biol. 27: 339-350
8、Dempsey D. A., and Klessig D. F.(1995) Signals in plant disease resistance. Bull. Inst.
Pasteur. 93: 167-186
9 、 Duerr B., Gawienowski M., Ropp T., and Jacobs T. ( 1993 ) MsERK1 : a mitogen-activated protein kinase from a flowering plant. Plant Cell 5: 87-96
10、Ewald D. and Kretzschmar U.(1996)The influence of micrografting in vitro on tissue culture behavior and vegetative propagation of old European larch trees. Plant Cell
Tissue Organ Cult. 4: 249-252
11、Fligiel S.E.G., Relan N.K., Dutta S., Tureaud J., Hatfield J., and Majumdar A.P.N.
(1994) Aging diminishes gastric mucosal regeneration: relationship to tyrosine kinases.
Laboratory Investigation 70: 764-774
12、Fouret Y., Arnaud Y., and Larrieu C.(1985)Rajeunissement in vitro du Sequoia sempervirens. Ann. AFOCEL 1984: 111-137
13、Hirt H.(1997)Multiple roles of MAP kinase in plant signal transduction. Trends in Plant Science 2: 11-15
14、Hirt H.(2000)Connecting oxidative stress, auxin, and cell regulation through a plant mitogen-activated protein kinase pathway. Proc. Natl. Acad. Sci. USA. 97: 2405-2407 15、Huttyl A.K., and Phillips A.L.(1995)Gibberellin-regulated expression in oat aleurone
cells of two kinases that show homology to MAP kinase and ribosomal protein kinase, Plant Mol. Biol. 27:1043-1052
16、Huang H.J., Chen Y., Kuo J.L., Kuo T.T., Tzeng C.C., Huang B.L., Chen C.M., and Huang L.C.(1996)Rejuvenation of Sequoia sempervirens in Vitro:Changes in
Isoesterases and Isoperoxidases. Plant Cell Physiol. 37: 77-80
17、Huang H.J., Lin Y.M., Huang D.D., Takahashi T., and Sugiyama M.(2003)Protein Tyrosine Phosphorylation during Phytohormone-Stimulates Cell Proliferation in Arabidopsis Hypocotyls. Plant Cell Physiol. 44: 770-775
18、Huang L.C., Hsiao C.K., Lee S.H., Huang B.L., and Murashige T.(1992a)
Restoration of vigor and rooting competence in stem tissues of mature citrus by repeated grafting of their shoot apices onto freshly germinated seedlings in vitro. In Vitro Cell.
Dev. Biol. 28: 30-32
19、Huang L.C., Kuo C.I., Wang C.H., Murashige T., and Huang T.C.(2000)Ethylene
evolution by juvenile and adult developmental phases of Sequoia sempervirens shoots cultured in vitro. Bot. Bull. Acad. Sin. 41:263-266
20、Huang L.C., Lin L.Y., Chen C.M., Chen L.J., Huang B. L., and Murashige T.(1995)
Phase reversal in Sequoia sempervirens in relation to mtDNA. Physiol. Plant. 94:
379-383
21、Huang L.C., Lius S., Huang B.L., Murashige T., Mahdi E.F.M. and Van Gundy R.
(1992b)Rejuvenation of Sequoia sempervirens by repeated grafting of shoot tips onto juvenile rootstocks in vitro. Plant Physiol. 98: 166-173
22、Hutchison K. W., Sherman C. D., Smith S. S., Singer P. B., and Greenwood M. S.
(1994)Maturation in larch. II. Effects of age on photosynthesis and gene expression in developing foliage. Plant. Physiol. 94: 1308-1315
23、Ichimura K., Mizoguchi T., Yoshida R., Yuasa T., and Shinozaki K.(2000a)Protein phosphorylation and dephosphorylation in environmental stress responses in plants.
Advances in Botanical Research 32: 355-377
24、Ichimura K., Shinozaki K., and Tena G.(2002)Mitogen-activated protein kinase cascades in plants: a new nomenclature. Trends in Plant Science 7: 301-308
25、Ikeyama S., Kokkonen G., Shack S., Wang X.T., and Holbrook N.J. (2002) Loss in oxidative stress tolerance with aging linked to reduced extracellular signal-regulated kinase and Akt kinase activities. The FASEB J. 16: 114-116
26 、 Islas-Flores I., Oropeza C., Hernanadez-Soromayor, S.M. ( 1998 ) Protein phosphorylation during coconut zygotic embryo development. Plant Physiol. 118:
257-263
27、Jonak C., Heberle-Bors E., and Hirt H.(1994)MAP kinases: universal multipurpose signalling tools. Plant Mol. Biol. 24: 407-416
28、Komjanc M., Festi S., Rizzotti L., Cattivelli L., Cervone F., and De Lorenzo G.
(1999)A leucine-rich repeat receptor-like protein kinase (LRPKm1) gene is induced in Malus x domestica by Venturia inaequalis infection and salicylic acid treatment. Plant
Mol. Biol. 40: 945–957
29、Kovtun Y., Chiu W.L., Zeng W., and Sheen J. (1998) Suppression of auxin signal transduction by a MAPK cascade in higher plants. Nature 395: 716-720
30、Kuo J. L., Huang H.J., Cheng C.M., Chen L.J., Huang B.L., Huang L.C., and Kuo T.T.(1995)Rejuvenation in vitro:Modulation of Protein Phosphorylation in Sequoia
sempervirens. J. Plant Physiol. 146: 333-336
31 、 Ligterink W., Kroj T., zur Nieden U., Hirt H., and Scheel D. ( 1997 ) Receptor-mediated activation of a MAP kinase in pathogen defense of plants. Science 276: 2054-2057
32、Liu Y, Guyton K.Z., Gorospe M., Xu Q., Kokkonen G.C., Mock Y.D., Roth G.S., and Holbrook N.J. (1996) Age-related decline in mitogen-activated protein kinase activity
in epidermal growth factor-stimulated rat hepatocytes. J. Biological chemistry 271:
3604-3607
33、Meskiene I., and Hirt H.(2000) MAP kinase pathways: molecular plug-and-play chips for the cell. Plant Mol. Biol. 42: 791-806
34、Mizoguchi T., Hayashida N., Yamaguchi-Shinozaki K., Kamada H., and Shinozaki K.
(1993)ATMPKs:a gene family of plant MAP kinase in Arabidopsis thaliana. FEBS Lett. 336: 440-444
35、Mizoguchi T., Ichimura K., Shinozaki K.(1997)Environmental stress response in plants: the role of mitogen-activated protein kinase. Trends in Biotechnology 15: 15-19 36、Muzik T.J. and Cruzado H.J.(1958)Transmission of juvenile rooting ability from
seedlings to adults of Hevea brasiliensis. Nature 181: 1288
37、Munnik T., Ligterink W., Meskiene I., Calderini O., Beyerly J., Musgrave A., and
Hirt H. (1999) Distinct osmo-sensing protein kinase pathways are involved in signaling
moderate and severe hyper-osmotic stress. Plant J. 20: 381-388
38、Nishiguchi M., Yoshida K., Sumizono T., and Tazaki K.(2002) A receptor-like protein kinase with a lectin-like domain from lombardy poplar: gene expression in response to wounding and characterization of phosphorylation activity. Mol Genet Genomics. 267:
506-514
39、Revilla M.A., Pacheco J., Casares A., and Rodriguez R.(1996)In vitro reinvigoration of mature olive trees (Olea europaea L.) through micrografting. In Vitro Cell. Dev. Biol.
Plant 32: 257-261
40、Rogler C. E., and Dahmus M. E.(1974)Gibberellic acid-induced phase change in Hedera helix as studied by deoxyribonucleic acid-ribonucleic acid hybridization. Plant.
Ohysiol. 54: 99-94.
41、Ryals J. A., Neuenschwander U. H., Willits M. G., Molina A., Steiner H.-Y., and Hunt M. D.(1996) Systemic Acquired Resistance. The Plant Cell 8: 1809-1819
42、Ryals J., Uknes S., and Ward E.(1994)Systemic Acquired Resistance. Plant Physiol.
104: 1109-1112
43、Stafstrom J. P., Altschuler M., and Anderson D. H.(1993)Molecular cloning and expression of a MAP kinase homologue from pea. Plant Mol. Biol. 22: 83-90
44、Walker N.(1986) Sequoia sempervirens. Rejuvenilisation et culture de meristemes en cascade. Ann. AFOCEL 1985: 25-47
45、Wilson C., Anglmayer R., Vicente O., and Heberle-Bors E. (1995) Molecular cloning, functional expression in Escherichia coli, and characterization of multiple mitogen-activated protein kinase from tobacco. Eur. J. Biochem. 233: 249-257
46、Wilson C., Eller N., Gartner A., Vicente O., and Heberle-Bors E.(1993)Isolation and characterization of a tobacco cDNA clone encoding a putative MAP kinase. Plant Mol.
Biol. 23: 543-551
47、Woo H. H., Hackett W. P., and Das A.(1994)Differential expression of a chlorophyll a/b binding protein gene and a proline rich protein gene in juvenile and mature phase English ivy (Hedera helix). Physiol. Plant. 92: 69-78
48、Zhang S., and Klessig D.F. (1998) The tobacco wounding-activated mitogen-activated protein kinase is encoded by SIPK. Proc. Natl. Acad. Sci. USA. 95: 7225-7230
49、Zhang S., and Klessig D. F.(2000)Pathogen-induced MAP kinase in tobacco. Results Probl Cell Differ. 27: 65-84
ttgtt gctgtactgaagaaaatcccagagactatttacaccggtcaagggttcgaatcccagacttcagttacaaa
tctcttccagatacgtagcggactagttaacaaggagggctaattgattgacgagaaactaatcggatcat ATG GAA GCC GCA GCC GCA TTT CAA TCG ACA GAT ACG CTA ATG TCT GAT GCT CAT 54 1 M E A A A A F Q S T D T L M S D A H
CCT CCC CCT GCA AGC AAC CCG ATG GAT AAC ATC CTC GCA GTG CCC AGC CAC GGT 108 19 P P P A S N P M D N I L A V P S H G
GGC CGC TTT ATG CAG TAT AAT ATT TTC GGC AAC ATT TTT GAA GTG ACT GCC AAG 162 37 G R F M Q Y N I F G N I F E V T A K
TAC AGA CCC CCT CTG CTG CCC ATT GGG AAA GGA GCT TAT GGG ATT GTC TGC TCT 216 55 Y R P P L L P I G K G A Y G I V C S GCA ATG AAT TCG GAG ACA AAT GAG CAT GTT GCC ATC AAG AAG ATT TCT AAT GCC 270 73 A M N S E T N E H V A I K K I S N A TTT GAC AAT CGC ATT GAT GCA AAG CGA ACT CTT CGA GAA ATC AAG TTG TGT CGA 324 91 F D N R I D A K R T L R E I K L C R CAT ATG GAT CAT GAA AAT GTA ATT GCC ATA AGA GAT ATA ATA CCT CCA CCA GAG 378 109 H M D H E N V I A I R D I I P P P E AGA GGA TCT TTT GAT GAT GTA TAC ATT GCA TAT GAA CTG ATG GAT ACT GAT CTC 432 127 R G S F D D V Y I A Y E L M D T D L CAT CAA ATT ATC CGT TCC AAT CAA GCA TTG TCA GAG GAG CAT TGT CAG TAC TTC 486 145 H Q I I R S N Q A L S E E H C Q Y F CTG TAT CAG ATT TTG CGA GGA TTA AAA TAT ATC CAT TCT GCA AAT GTA CTT CAT 540 163 L Y Q I L R G L K Y I H S A N V L H AGG GAT TTG AAG CCC AGC AAT CTT CTT CTG AAT GCA AAC TGT GAT CTA AAG ATA 594 181 R D L K P S N L L L N A N C D L K I TGT GAT TTT GGT CTT GCC CGT ATT ACC TCT GAA ACT GAT TTT ATG ACA GAA TAT 648 199 C D F G L A R I T S E T D F M T E Y GTT GTT ACA AGA TGG TAT AGA GCG CCA GAA TTG CTA CTA AAT TCG TCC GAT TAC 702 217 V V T R W Y R A P E L L L N S S D Y ACC GCA GCA ATT GAT GTG TGG TCT GTA GGT TGT ATT TTT ATG GAA TTA ATG AAC 756 235 T A A I D V W S V G C I F M E L M N AGG ACA CCA TTA TTT CCA GGA AGG GAT CAT GTG CAT CAG CTG CGT TTG TTA ACA 810 253 R T P L F P G R D H V H Q L R L L T GAG TTG ATT GGT ACA CCA ACT GAG GCG GAT CTT GGG TTT GTT CGG AGT GAT AAT 864 271 E L I G T P T E A D L G F V R S D N GCT AGG AGA TTT ATT CGG CAA ATG CAT CAA TAT CCA AGG CAG TCA TTT GCT GAA 918 289 A R R F I R Q M H Q Y P R Q S F A E AAA TTT CCA CAT ATA CAC CCT TCA GCA ATT GAT CTC TGT GAA AAA ATG TTG ACA 972 307 K F P H I H P S A I D L C E K M L T TTT GAT CCA ACT CAG AGA ATC ACA GTG GAA GAT GCC CTT GCT CAT CCA TAC CTG 1026 325 F D P T Q R I T V E D A L A H P Y L GCA ACT TTG CAT GAT GTG AGT GAT AAG CCA GCC TGT TCT ATG CCT TTC AGC TTT 1080 343 A T L H D V S D K P A C S M P F S F GAA TCT GAG TAT AAT AAC CTC ACA GAG GAA CAG ATG AAA GAA CTG ATA TAT AGG 1134 361 E S E Y N N L T E E Q M K E L I Y R
GAG GCT CTG GCA TTT AAT CCA GAG TAT GCA CAA TAA aag tca cct ttg ttc cca 1170 379 E A L A F N P E Y A Q *
gcagtactatacagctttggcaagtataccactgggcagtttcatgtttgtggtaattgtattgtcattat ttattgagttttgaaagtaatgtaaatatgtatattaaaaccgtgttagggcctggagaagggataaggaa ttatattatatttgatttaaaaaatctgcatatatactccatacatataccatcccttttatgaggtgtat catggatatatgggccttggacctaaacatcatgtctcaaaactgcagttttccttattcatccttctcat caaactccaagaggttgtatgagcatttttgctaattatgggagaaaaattcattaagttgacaaggagat tgatggtagtttgtttgtttgtttgaggtagatgccaaaacaataagatttgttgtttggcactgatgtat tggtgttagtctgcatttatatgctgtattccataaatttttagtatcacagtcaaccaaagtttggtcca aaaaaaaaaaaaaaaaaaaaaaaaaaaaa
圖 1、紅杉 SsMAPKA cDNA 序列示意圖
SsMAPKA 之總長為 1843bp,包含 5’端非譯碼區 147bp,3’端非譯碼區 526bp,其中譯碼區共 1170bp,可轉譯出 389 個氨基酸。序列之左邊數字代表氨 基酸序列順序,序列之右邊數字代表核苷酸序列順序。M 代表轉譯起始甲硫氨 酸(methionine),*代表終止密碼子,網底部分為 3’端非譯碼區之探針序列。
SsMAPKA 1 MEAAAAFQSTDTLMSDAHP----PPASN---P--MDNILAVPSHGGRFMQYNIFGN MsSIMK 1 MEGGG-APPADTVMSDAAP---APPQ---MG---IENIPAVLSHGGRFIQYNIFGN PsD5 1 MEGGGGAPAADAVMEDAAPQQ--QEPQQQAAMG---IENIPATLSHGGRFIQYNIFGN AtMPK6 1 MDGGSGQPAADTEMTEAPGGF-PAAAPSPQMPG---IENIPATLSHGGRFIQYNIFGN OsSIPK 1 MDAG--AQPPDTEMAEAGGGQQPPAAAAAAGAGAGAGMMENIQATLSHGGRFIQYNIFGN
Ⅰ Ⅱ Ⅲ SsMAPKA 48 IFEVTAKYRPPLLPIGKGAYGIVCSAMNSETNEHVAIKKISNAFDNRIDAKRTLREIKLC MsSIMK 47 IFEVTAKYKPPIMPIGKGAYGIVCSAHNSETNEHVAVKKIANAFDNKIDAKRTLREIKLL PsD5 54 IFEVTAKYRPPIMPIGKGAYGIVCSAHNSETNEHVAVKKIANAFDNKIDAKRTLREIKLV AtMPK6 55 IFEVTAKYKPPIMPIGKGAYGIVCSAMNSETNESVAIKKIANAFDNKIDAKRTLREIKLL OsSIPK 59 VFEVTAKYKPPILPIGKGAYGIVCSALNSETGEQVAIKKIANAFDNKIDAKRTLREIKLL
Ⅳ Ⅴ
SsMAPKA 108 RHMDHENVIAIRDIIPPPERGSFDDVYIAYELMDTDLHQIIRSNQALSEEHCQYFLYQIL MsSIMK 107 RHMDHENVVAIRDIVPPPQREVFNDVYIAYELMDTDLHQIIRSNQALSEEHCQYFLYQIL PsD5 114 RHMDHENVVAIRDIVPPPQREVFNDVYIAYELMDTDLHQIIRSNQALSEEHCQYFLYQIL AtMPK6 115 RHMDHENIVAIRDIIPPPLRNAFNDVYIAYELMDTDLHQIIRSNQALSEEHCQYFLYQIL OsSIPK 119 RHMDHENIVAIRDIIPPPQRNSFNDVYIAYELMDTDLHQIIRSNQALSEEHCQYFLYQIL
Ⅵ Ⅶ ▼ Ⅷ SsMAPKA 168 RGLKYIHSANVLHRDLKPSNLLLNANCDLKICDFGLARITSETDFMTEYVVTRWYRAPEL
MsSIMK 167 RGLKYIHSANVLHRDLKPSNLLLNANCDLKICDFGLARVTSETDFMTEYVVTRWYRAPEL PsD5 174 RGLKYIHSANVLHRDLKPSNLLLNANCDLKICDFGLARVTSETDFMTEYVVTRWYRAPEL AtMPK6 175 RGLKYIHSANVLHRDLKPSNLLLNANCDLKICDFGLARVTSESDFMTEYVVTRWYRAPEL OsSIPK 179 RGLKYIHSANVLHRDLKPSNLLLNANCDLKICDFGLARTTSETDFMTEYVVTRWYRAPEL
Ⅸ Ⅹ
SsMAPKA 228 LLNSSDYTAAIDVWSVGCIFMELMNRTPLFPGRDHVHQLRLLTELIGTPTEADLGFVRSD MsSIMK 227 LLNSSDYTAAIDVWSVGCIFMELMDRKPLFPGRDHVHQLRLLMELIGTPSEDDLGFLN-E PsD5 234 LLNSSDYTAAIDVWSVGCIFMELMDRKPLFPGRDHVHQLRLLMELIGTPSEADLGFLN-E AtMPK6 235 LLNSSDYTAAIDVWSVGCIFMELMDRKPLFPGRDHVHQLRLLMELIGTPSEEELEFLN-E OsSIPK 239 LLNSSEYTAAIDVWSVGCIFMELMDRKPLFPGRDHVHQLRLLMELIGTPNEADLDFVN-E
XI
SsMAPKA 288 NARRFIRQMHQYPRQSFAEKFPHIHPSAIDLCEKMLTFDPTQRITVEDALAHPYLATLHD MsSIMK 286 NAKRYIRQLPPYRRQSFQEKFPHVHPEAIDLVEKMLTFDPRKRITVEDALAHPYLTSLHD PsD5 293 NAKRYIRQLPLYRRQSFQEKFPHVHPEAIDLVEKMLTFDPRQRITVENALAHPYLTSLHD AtMPK6 294 NAKRYIRQLPPYPRQSITDKFPTVHPLAIDLIEKMLTFDPRRRITVLDALAHPYLNSLHD OsSIPK 298 NARRYIRQLPRHARQSFPEKFPHVHPLAIDLVEKMLTFDPRQRITVEGALAHPYLASLHD
SsMAPKA 348 VSDKPACSMPFSFESEYNNLTEEQMKELIYREALAFNPEYAQ MsSIMK 346 ISDEPVCMTPFSFDFEQHALTEEQMKELIYREALAFNPEYQQ 80%
PsD5 353 ISDEPVCTTPFSFDFEQHALTEEQMKELIYREALAFNPEYQQ 82%
AtMPK6 354 ISDEPECTIPFNFDFENHALSEEQMKELIYREALAFNPEYQQ 78%
OsSIPK 358 ISDEPVCSSPFSFDFEQHALSEEQMKDLIYQEGLAFNPDYQ- 79%
圖 2、紅杉 SsMAPKA 與不同物種 MAPK 氨基酸序列保留性比較
紅杉 SsMAPKA 與不同物種 MAPK 族系成員,於 BioEdit 套
組中以 ClustalW Multiple Alignment 程式進行氨基酸排序比對,黑
色區塊表示相同的氨基酸,灰色區塊表示特性相似的氨基酸。羅
馬數字Ⅰ∼ⅩI 表示典型 Ser/Thr 激酶所共有的激酶區域,
▼為植
物 MAPK 所特有的 TEY 磷酸化位點,即為 MAPK 之活化區。右
邊數字百分比表示與該物種之相似度。
圖 3、紅杉 SsMAPKA 與不同物種 MAPK 所構築之親緣演化樹
各基因之氨基酸序列取自於NCBI (National Center Biotechnology Information)資料庫,註冊編號 如下:NtNTF4 (Q40532), NtSIPK (AAB58396), AtMPK6 (BAA04869), NtWIPK (BAA09600), PsD5 (Q06060), MsSIMK (Q07176), OsSIPK (CAD59793), ZmMPK5 (BAA74734), OsMAPK2 (CAC13967), ZmMPK4 (BAA74733), AtMPK3 (BAA04866), NtWIPK (BAA09600), PsMAPK3 (AAF73236), MsSAMK (T09622), AtMPK4 (BAA04867), MsMMK2 (CAA57719), AtMPK13 (AAF75067), NtNTF6 (CAA58760), AtMPK1 (BAA03535), AtMPK2 (BAA03536), AtMPK7 (BAA04870), AtMPK14 (CAB16812), OsMAPK3 (AAG40581), OsMAPK4 (CAB61889), AtMPK18 (AAG51978), AtMPK19 (NP_188090), AtMPK20 (AAD21721), OsMPKG1 (BAB63546), AtMPK16 (NP_197402), OsMPKG2 (BAB40147), AtMPK8 (AB038693), AtMPK17 (AAC67338), AtMPK9 (AB038694), OsBWMK1 (AAD52659), OsRMAPK2 (AAF23903), AtMPK15 (AAG52072)。各亞群內包含現今已發表或假設性之植物 MAPK (Ichimura, 2002)。
NtNTF4 NtSIPK PsD5 MsSIMK
AtMPK6 OsSIPK
ZmMPK5 SsMAPKA
OsMAPK2 ZmMPK4 AtMPK3
NtWIPK PsMAPK3 MsSAMK AtMPK4
MsMMK2 AtMPK13 NtNTF6
AtMPK7 AtMPK14 AtMPK1
AtMPK2 OsMAPK3
OsMAPK4 OsMPKG1
AtMPK18 AtMPK19
AtMPK20 AtMPK16
OsMPKG2 AtMPK17 OsBWMK1
OsRMAPK2 AtMPK9 AtMPK8
AtMPK15
OsMEK1
0.2
A1
A2
B
C
D
圖 4、紅杉 SsMAPKA 基因族系組成
將紅杉(Sequoia sempervirens)之基因組 DNA 進行限制酶切割作用,
並由電泳分離後使用南方墨點法分析紅杉 SsMAPKA 基因族系組成,以
SsMAPKA 基因之 3’端非譯碼區探針進行雜合。數字 1 及 4、2 及 5、3 及
6 分別代表 EcoRⅠ, HindⅢ及 XbaⅠ限制酶。
圖 5、紅杉 SsMAPKA 基因在老樹(AS)與年輕樹(SS)組織特異性表
現分析
將 生 長一 個 半月紅 杉 老樹 與 年輕樹 組 培苗 分 別取頂 芽 1.5 cm
(apical)及頂芽下之 1.5 cm(basal)組織之總體 RNA 以電泳分離後,
選取 SsMAPKA 之 3’端非譯碼區序列,並以α-
32P-dCTP 標定後作為雜合
探針,進行北方墨點分析 SsMAPKA 基因組織特異性之表現。以紅杉之
rRNA 轉錄表現作為內控制組。
圖 6、紅杉 SsMAPKA 基因在不同嫁接植株中表現分析
分別萃取紅杉老樹(AS)、一次嫁接(1x)、二次嫁接(2x)、三次嫁
接(3x) 、四次嫁接(4x) 、五次嫁接(5x)及年輕樹(SS)之頂端(apical)
(A 圖)及基部(basal) (B 圖)總體 RNA,以電泳分離後,選取 SsMAPKA
之 3’端非譯碼區序列,並以 α-
32P-dCTP 標定後作為雜合探針,進行北方
墨點分析 SsMAPKA 基因在不同嫁接植株中表現。以紅杉之 rRNA 轉錄表
現作為內控制組。
![TraditionalMLCalgorithmsmainlytacklethebatchMLCproblem,wheretheinputdataarepresentedinabatch[24,28].Nevertheless,inmanyMLCapplicationssuchase-mailcategorization[22],multi-labelexamplesarriveasastream.Onlineanalysisistherefore dimensionreducermotivatedbyma](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)