以二倍頻顯微術觀測熱致膠蛋白構造改變
46
0
0
全文
(2) 誌謝 感謝指導老師 高甫仁教授兩年來的細心指導,使我在碩士班兩 年的時間裡能夠完成我的論文。感謝中山大學材料所教授 徐瑞鴻教 授在我的實驗上的指導,並且在論文口試時,非常仔細地指正出論文 中的錯誤。 感謝我的口試委員中山光電所教授 鄭木海教授、中山材料所教 授 沈博彥教授與中山光電所教授 李晁逵教授在口試時對實驗結果 所提出種種寶貴的意見。 最後要感謝我的父母,謝謝他們為我付出的一切,讓我能夠在這 兩年時間裡放心的學習。以及實驗室全體同仁的互相幫助,讓我的論 文可以順利的完成。. I.
(3) 中文摘要 膠原蛋白為生物體的結構型蛋白質,例如:肌肉組織、骨骼和韌 帶等,在動物體中所扮演的角色為連接細胞與組織。而「溫度」這個 與我們生活習習相關的天然環境變數,則是深深地影響到膠原蛋白的 功能與型態。在生物學的研究中,溫度引起之膠原蛋白結構改變是一 重要主題。傳統上,研究的方式大多係使用差分掃瞄熱量儀 DSC 與非 線性光學偵測型變(conformational change)的產生。DSC 可以量測 出膠原蛋白的相變點、反應熱與熱穩定性等熱力學性質,但缺點則是 無法產生局部化之資訊。使用非線性倍頻光學的方式,則可以藉由倍 頻光的強度推測得膠原蛋白結構的變化,並且可以產生倍頻影像,進 行局部變化的分析。在之前的研究成果中發現到,環境中的水分子濃 度關係到膠原蛋白結構對溫度變化的穩定性。相對地,本研究使用一 真空變溫平台以控制環境,藉以隔絕環境中的水分子、氧氣與雜質。 配合以非線性掃瞄式倍頻光學顯微鏡的使用,便可以即時變溫的方式 觀察真空中膠原蛋白的變化,並藉由倍頻顯微影像的強度,推測出膠 原蛋白結構的改變。. II.
(4) Abstract Collagen is an important structural protein in living organisms and plays an indispensable role in connecting cells and tissues, such as in musculature, bone, and ligament. The stability and conformation of collagen are, however, strongly influenced by ambient temperature and constitutes an interesting subject of study. Thermally induced conformation change of collagen has been investigated by techniques such as differential scanning calorimetry (DSC) and second harmonic generation. DSC is a powerful method in uncovered important thermal dynamics properties including phase change, enthalpy, and thermal stability of the collagen. However, due to its collective nature, no localized information can be found. For comparison, second harmonic generation, which reflects structural symmetry, can be combined with laser scanning microscopy to investigate localized variation. It has been shown in previous studies that the thermal stability of collagen is strongly influenced by the water content within collagen. For comparison, we are investigating the conformational change of collagen under a vacuum stat with second harmonic microscopy so as to isolate environmental effects, particularly those from water and oxygen. In this way, we have found the conformational change of collagen takes place at a much higher temperature and activation energy. Additionally, the high spatial resolution achieved also allows many further possibilities.. III.
(5) 論文名詞中英對照表 amino acids:氨基酸 collagen:膠原蛋白 conformation change:結構改變 carboxyl group:羧基群 hemoglobin:血紅素 keratin:角素 nucleic acids:核酸 second harmonic generation:二倍頻 polysaccharides:多醣 polypetide:多胜肽. IV.
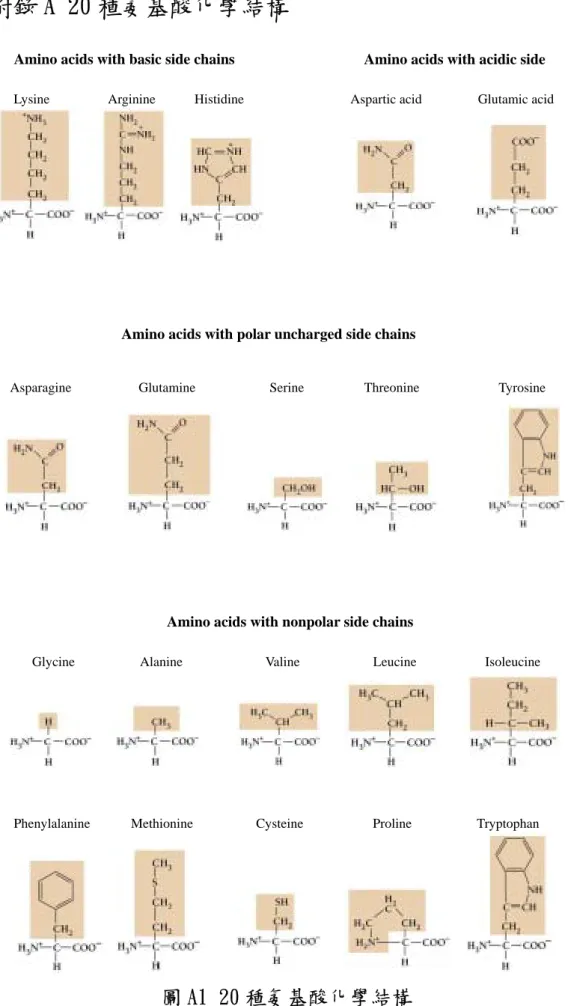
(6) 目. 錄. 第一章. 緒論.......................01. 第二章. 二倍頻顯微術理論與膠原蛋白結構. 2.1 非線性光學倍頻簡介................04 2.2 蛋白質與膠原蛋白之結構..............06 第三章. 研究方法. 3.1 樣本選取.....................13 3.2 研究架構.....................13 3.3 實驗步驟.....................15 3.4 資料處理.....................16 第四章. 結果與討論. 4.1 倍頻訊號與溫度關係................18 4.2 不同溫度下,倍頻訊號隨時間的衰減.........19 第五章. 結論.......................23. 參考文獻.........................24 附錄 A. 20 種氨基酸化學結構................27. 附錄 B 雷射光源系統...................28 附錄 C 電子偵測系統...................31 附錄 D 真空變溫系統...................34 附錄 E 光學元件.....................35 附錄 F 掃描式共焦顯微系統................37. V.
(7) 圖表目錄 圖 2.1 二倍頻產生....................06 圖 2.2 氨基酸基本組成結構圖...............08 圖 2.3 氨基酸脫水結合示意圖...............08 圖 2.4 蛋白質鏈的首要結構................09 圖 2.5 第二類蛋白質鏈結構................10 圖 2.6 第三類蛋白質鏈結構與第四類蛋白質鏈結構......11 圖 2.7 膠原蛋白基本結構.................12 圖 2.8 膠原蛋白顯微影像.................12 圖 3.1 反射式倍頻真空變溫顯微系統架構..........14 圖 3.2 反射式倍頻真空變溫顯微系統實體架設圖.......14 圖 3.3 膠原蛋白樣品放置完成圖..............15 圖 3.4 膠原蛋白之二倍頻影像...............17 圖 4.1 倍頻強度與溫度關係................18 圖 4.2 不同溫度條件,倍頻強度隨時間衰減的速率......20 圖 4.3 將圖 4.2 做數學的一階指數擬合(420K 和 440K 除外)..21 圖 4.4 在一般環境下,膠原蛋白倍頻訊號強度與溫度關係圖..22 圖 4.5 環境中的水分子濃度與熱穩定點 Td(○)關係圖.....22 圖 A1 20 種氨基酸化學結構.................27 圖 B1 Verdi 內部結構圖..................28 圖 B2 Verdi 激發 Mira 900 輸出功率隨波長調整曲線圖.....29 圖 B3 Mira 900 內部結構圖.................29 圖 C1 頭端入射型光電倍增管結構..............31. VI.
(8) 圖 C2 低噪音前置電壓放大器前視圖............32 圖 C3 高壓電源供應器前視圖...............33 圖 D1 真空變溫平台結構圖................34 圖 E1 隔離器內部光路結構圖...............36 圖 F1 FV-300 雷射光束掃瞄系統..............37 表 2.1 膠原蛋白在生醫材料上的應用............11. VII.
(9) 第一章 1-1. 緒論. 前言與研究動機 將高強度的雷射光束聚焦於物質上,可以讓我們研究物質內部的. 光學非線性效應。這種主要的非線性效應是由物質在電場中的二階極 化向量(polarization)所產生的,並導致二倍頻(second harmonic generation 簡稱 SHG)的效應。這種現象首在 1961 年時,由 Franken 和他的同事所發現(Franken et al., 1961)。之後,Bloembergen 以方程式的方式將非線性光學和另外一些二倍頻的基本原理表示出 來(Bloembergen, 1965) 。 由於二倍頻的特性,讓我們可以使用於局部的非等向性 (anisotropic)探測、分子超極化(hyperpolarizabilities)的分 佈情形、相位匹配(phase-matching)的特性和表面電漿增強的交互 作用 (surface plasmon enhanced interaction) 。在 1973 年時,Dolino 使 用 一 個 Q-switched 脈 衝 雷 射 得 到 硫 酸 三 甘 鈦 ( triglycine sulfate)晶體鐵電域的實際二倍頻影像(Dolino et al., 1973)。不 久之後,Hellwarth 和 Christensen 使用雷射光束的聚焦點來當成微 探針,以二倍頻的效應來檢查硒化鋅 ZnSe 複晶(polycrystalline) 結構,且其觀測到的是一般使用線性光學顯微鏡所無法觀得到的結果 (Hellwarth and Christiansen, 1974)。此結果導致二倍頻應用於 非線性光學顯微鏡的觀念(Hellwarth and Christiansen, 1975)。 在 1978 年,Gannaway 和 Sheppard 在掃瞄式顯微鏡上實現了光學二 倍頻(Gannaway and Sheppard, 1978)。他們應用此種技術到非線性 的顯微研究上,像是 KDP 和 LiNbO3。二倍頻量測與線性光效應相比之 下,只有在輸入較高的雷射能量時其優勢才能顯現出來,但是較高的 雷射能量並不適合做為脆弱樣品的研究。因此,在掃瞄式顯微鏡上二 倍頻影像(SHGI)的應用空間就被侷限了,直到有一種加強非線性效 應的方法被發現,才改善了先前的問題。在八十年代末期,鎖模 (mode-locked)雷射的出現提供了一個滿足產生非線性光學現象所 需條件的光源。在相同的平均雷射光功率之下,超快雷射的脈衝其高 1.
(10) 瞬間峰值功率大大的增加了二倍頻訊號的強度。透過二倍頻的測量方 式 ,可量 測的樣 品範圍 包括複 晶系的 分子膜 (Jentsch et al., 1996)、薄膜(Bottomley et al., 1996; Vydra and Eich, 1998)、 鋁的表面(Janz et al., 1991)和金的表面(Moustaizis et al., 1995)。在 LBO-based 樣品上已發表,使用飛秒(femtosecond)雷 射脈衝,可以得到高解析三維二倍頻影像的技術(Gauderon et al., 1998)。二倍頻的應用不僅止於材料科學,它也已經成功的應用在生 物和醫學的領域。例如:使用二倍頻來繪製老鼠尾巴肌腱的影像 (Freund et al., 1996)、探測細胞膜電位(membrane potentials) (Bouevitch et al., 1993; Lewis et al., 1997)和動物組織的影 像(Guo et al., 1996;Guo et al., 1997)。以上內容為參考 Gauderon 等人相關文獻(Gauderon, 2001) 「溫度」是與我們生活習習相關的天然環境變數,也深深影響膠 原蛋白的穩定性,例如:肉類食品保存、皮膚燒燙傷等,均是膠原蛋 白與溫度的關係。而本研究是以二倍頻理論為基礎,利用非線性光學 的方法來量測膠原蛋白(collagen)在結構上的變化。膠原蛋白佔哺 乳動物身體體重百分之六左右。其分子由三個氨基酸鏈旋繞成一個棒 狀的三重螺旋(triple helix)結構,這是由 Fraser 等人透過 X 光 繞射圖案所發現的(Fraser et al., 1979)。至於膠原蛋白的結構 我們會在之後的章節加以詳細的描述。透過之前的研究已經證實,經 過加熱能使得膠原蛋白中的螺旋狀(helix-coil)結構發生改變 ( Miles et al., 1995; Tiktopulo et al., 1998; Shnyrov et al.,1992; Rochdi et al., 1999)。由微差掃描熱量儀(differential scanning calorimetry 簡稱 DSC)來進行實驗,其大部份的特點在老 鼠尾巴組織的膠原蛋白纖維上,分子結構改變皆在相近的溫度 62°C, 這是經由特殊的熱容量測方式所顯示出來的(Miles et al., 1995)。 以 Type I 型式的膠原蛋白之熱轉變再一次的利用微差掃描熱量儀 DSC 和分光光譜議(spectrophotometry)來進行研究,其結果指出一個 不可逆過程的中心溫度大約在 52°C。利用二倍頻效應結合掃瞄式顯 微鏡,便能以即時的方式量測膠原蛋白型變的二倍頻影像。而膠原蛋 2.
(11) 白會有非線性光學的二倍頻效應是由於其分子結構所造成的。二倍頻 強度根據推測是與微晶(microcrystalline)結構的三重螺旋膠原蛋 白分子百分比有關。二倍頻強度的強弱能顯出結構改變率,此即為型 態改變後的膠原蛋白纖維部分,因缺少結晶結構和其附屬的非線性特 性所致。換句話說,透過二倍頻的方式可以很直接的量測到分子結構 改變後的纖維比例(Theodossiou et al.,2002)。在 Rochdi 等人的 研究顯示,膠原蛋白所處環境的水分子濃度會影響到膠原蛋白對溫度 的存活能力,當水分子的濃度越高,膠原蛋白的熱穩定性就會越低, 這是由於水分子會影響到膠原蛋白分子內部鏈結能力(Rochdi et al.,1999)。 本論文係延續 Theodossiou 等人所做的膠原蛋白變溫實驗之基 本概念(Theodossiou et al.,2002),並且加入 Rochdi 等人的研究 結果 (Rochdi et al.,1999) 修改其實驗架構與設備。由於 Theodossiou 等人的實驗並無法得到完整的倍頻影像,因此在經由實驗架構與設備 的改良之後,我們能夠以即時的方式得到二倍頻影像。以短脈衝雷射 做為量測光源,透過掃瞄式共焦顯微鏡後,便可得到二倍頻影像,再 以真空變溫平台做為樣品台,消除水分子的影響,並改變不同的實驗 溫度。由此方式,便可得到在真空下,膠原蛋白隨溫度變化之摺疊影 像。. 1-2. 論文架構概要 在第二章中,將詳述本論文的量測原理,即為二倍頻之基本原. 理,緊接著再介紹蛋白質的基本結構與膠原蛋白的相關資訊。 第三章繫針對在實驗過程中,二倍頻顯微鏡的基本架構及實驗步 驟之完整的說明。同時,將使用的實驗設備和光學元件做一簡單的介 紹。 第四章對二倍頻顯微鏡所得之實驗結果做探討與分析,並比較之 前相關研究數據,以提供合理、完整的解釋。 第五章為本論文之結論與建議。 3.
(12) 第二章 二倍頻顯微術理論與膠原蛋白結構 光學倍頻顯微術為近幾十年來所發展出來的顯微技術,其特點為 對結構變化的敏感性極佳。對於同一個樣品而言,不同區域之結構所 產生的倍頻特性也不盡相同,如二倍頻與三倍頻,其利用不同的接收 方式,便可將這些結構資訊組合起來,而得到樣品完整的結構資訊。 由於倍頻效應只在雷射聚焦點處激發,所以倍頻效應在空間上有著較 佳的三維解析度,因此組合而成的資料便得以以三維的影像來表現, 這是以往使用傳統光學顯微鏡所無法達成的技術。更因為倍頻效應無 涉及電子能階躍遷的問題,不會將雷射光部份能量留在物質上,才不 至於導致對物質破壞。這項量測技術目前也已經應用在生物醫學與材 料科學領域。以下將開始介紹二倍頻的基本理論。. 2.1 非線性光學二倍頻簡介 由於非線光學的內容太為廣泛,所以我們只針對此研究所採用的 光學二倍頻原理做簡單的介紹。對於一個線性介質而言,當一個電場 通過該介質時,其介質受到電場感應所產生的極化向量 (Polarization)可寫成. P = ε0χ E. (2.1). 其中 χ 為介質之線性電化率(Linear susceptibility),而 ε 0 是指真 空的介電常數(Vacuum permittivity)。 但是,若將此介質通過一道很強的電場時,由於介質受到強電場 的影響,會使得此介質的極化向量與電場強度之間的關係成為了一個 非線性的關係,將此關係以 power series 的型式展開. P = ε 0 ( χ (1) + χ (2) E + χ (3) E 2 + ⋅ ⋅ ⋅) E = ε 0 χ (1) E + ε 0 χ (2) E 2 + ε 0 χ (3) E 2 + ⋅ ⋅ ⋅. (2.2). = P1 + ( P2 + P3 + ⋅ ⋅ ⋅) 其中 χ (2) 和 χ (3) 分別為非線性項的二階與三階非線性光學電化率。 4.
(13) 上述的展開式之中, P1 為線性項,其極化向量正比於電場強度, 所以此結果正滿足於傳統的光學理論,例如:重疊定理 (Superposition principle)、反射(Reflection)、折射(Refraction) 等。 P2 之後的項為非線性項,其極化向量與電場之間為非線性關係, 由於其前方的高階項係數之數量級很小,所以必需將電場強度提高, 非線性高階項的效應才會明顯表現出。以二階非線性項為例, χ (2) 和. χ (1) 數量級相差約為108 數量級,若要讓二階非線性的效果顯現出來, 則電場強度至少要為104 等級,因此必須使用特殊的雷射光源系統, 此 雷 射 光 源 系 統 可 聚 焦 至 1µ m 的 小 點 , 其 產 生 的 電 場 強 度 可 達. 1010V / m ,這使得高階的非線性部分得以顯現出來。以下將介紹二倍 頻的產生理論,其產生原理可由非線性部分的二次項 P2 = ε 0 χ 2 E 2 來解 釋,假設入射到非線性材料的雷射光其頻率為 ω ,其電場表示式為 (Boyd, 2003): E (t ) = Re { E0e − iωt } = E0 cos ωt. (2.3). 代入非線性項 P2 表示式中. 1 P2 (t ) = ε 0 χ (2) E02 cos 2 ωt = ε 0 χ (2) E02 (1 + cos 2ωt ) 2 1 1 P2 (t ) = ε 0 χ (2) E02 + ε 0 χ (2) E02 cos 2ωt 2 2 1 其數學推導結果中, ε 0 χ (2) E02 cos 2ωt 項即表示著二倍頻的存在。 2 如圖 2.1 所示圖(a)為二倍頻的示意圖,圖(b)則為二倍頻的能階簡 圖,一個處於基態(Ground State)的電子吸收了兩個頻率相同的光子 之後,到達了虛能態(Virtual State),然後再由虛能態放出了一個 倍頻的光子。非線性極化強度如下:. P(r , t ) = χ (2) E 2 (r , t ). (2.4). E (r , t ) = E (r )cos ωt. (2.5). 假設輸入的電場形式為:. 5.
(14) ω. χ (2). ω. ω. 2ω. ω. (a). 2ω (b). 圖 2.1 (a)SHG 示意圖 (b)SHG 能階簡圖. 此時假設材料有反對稱性(Inversion symmetry),在經由坐標轉換後. r → − r ,極化強度與電場則為 − P 及 − E 。將改變後的(2.5)式代入 (2.4)式便可得到以下的式子:. − P(r , t ) = χ (2) [ − E (r , t ) ]. 2. (2.6). 將(2.6)式簡化之後與(2.4),可以得到以下結果. χ (2) = 0. (2.7). 根據此結果可以得知,若材料存有中心對稱(Centrosymmetry),便無 法有 χ (2) 的產生。所以一個材料如要產生二倍頻訊號,非中心對稱即 為主要要求。然而結構與晶格的完整性,亦會影響到二倍頻訊號之強 弱;因結構與晶格受到外力所產生的改變,也會使得二倍頻轉換效率 跟著改變。因此如利用二倍頻來做為快速光學檢測方式是極為便利的 (Boyd, 2003)。. 2.2 蛋白質與膠原蛋白之結構 結構蛋白質有上千種,它們能將身體聚集在一起、給予骨頭支撐 的力量、在細胞內部或外部傳送分子和離子等等。每個有機體身上, 所包含蛋白質種類從數百種到數千種,而人類身上的蛋白質至少也包 含了 80,000 種以上。 6.
(15) 膠原蛋白則是在人體上最普遍的蛋白質,給與肌腱、韌帶、骨骼、 肌肉和皮膚等組織上強大的連結力量。再者,在人體上的膠原蛋白卻 有如鋼鐵般的強壯。強力的蛋白質讓皮膚富於彈性,多纖維的角素 (Keratin)使得頭髮、角質、指甲變得堅韌。另外某些蛋白質則是 用來運送物質,例如血紅素(Hemoglobin),它藉由血液運送方式攜 帶氧氣到肺部。如荷爾蒙、抗體或毒素(例如響尾蛇的毒液)均為蛋 白質的另一種。以下即為蛋白質的組成和種類詳細的介紹。. 2.2.1 蛋白質之組成與種類 蛋白質是由一個或多個氨基酸(amino acids)鏈所形成的,而 每個鏈稱之為”多胜肽(Polypeptide)”。一個蛋白質可能由一個 很簡單的鏈(多胜肽)或者數個複雜不同的鏈所組成,一個鏈包含數 百個甚至幾千個氨基酸。不管這個鏈有多長,或是鏈有多少個,每次 細胞只能建造一個蛋白質氨基酸元素。 氨基酸的種類共有 20 種,在這 20 種氨基酸中,每一種氨基酸都 有著相同的基本結構。每個結構包含著一個羧基群(carboxyl group) 和一個氨基群(amino group) ,如圖 2.2 所示。羧基群和氨基群透過 一個碳原子連接起來,我們稱這個碳原子為”alpha carbon”。 氨基酸的種類會不同,在於它有著不同種類的側鏈,我們稱這個 側鏈為“R Group”,此側鏈也與 alpha carbon 原子連接在一起。除 了“Proline”此種型式不同之外,其餘的 19 種均由 Alpha carbon、 羧基群和氨基群所組成。由於每一種氨基酸由不同的 R group 連接, 所以也產生了不同的特性,氨基酸類型如附錄 A 所示。透過這 20 種 不同的氨基酸單元就能形成不同種類的蛋白質。. 7.
(16) α (alpha) carbon. Side Chain. Amino group. Carboxyl group. 圖 2.2 氨基酸基本組成結構圖. 多胜肽的形成是經由氨基酸的脫水濃縮反應而成的,就像多醣 (polysaccharides)和核酸(nucleic acids)等。連接兩個氨基酸 元 素 而 形 成 多 胜 肽 的 特 別 鍵 結 被 稱 之 為 “ 胜 肽 鍵 ( peptide bond)”。當一個氨基群連接下一個羧基群,則兩個氨基酸就會連接 在一起,如圖 2.3 所示。 Amino acid. Amino acid. 1 water molecule. Peptide bond. 圖 2.3 氨基酸脫水結合示意圖 每一個氨基酸所形成鏈狀的方法是相同的,均是氨基群連接羧基 群,羧基群再連接氨基群的方式,如此便可形成多胜肽的基本結構。 而側鏈 R groups 則懸掛在主要結構的側邊,並且與同一環境下的其 他分子側鏈產生交互作用,所以導致蛋白質有不同的組成結構與特 8.
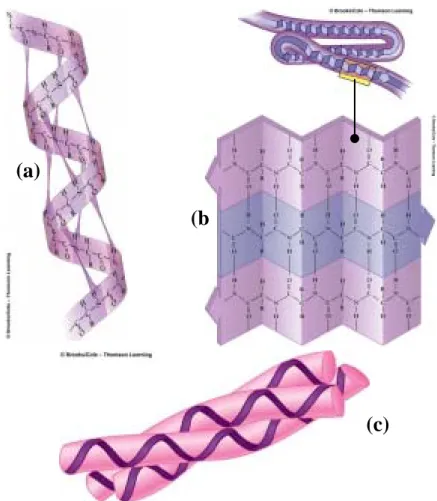
(17) 性。 儘管蛋白質有多種不同的結構,生物學家卻依然找出了分類它們 的方法。大多數密集且圓型的分子稱之為“球狀蛋白質(globular proteins)”,例如血紅素(hemoglobin),在血液之中此種蛋白質 主要是用來攜帶氧氣。除了球狀蛋白質之外還有“纖維狀蛋白質 (fibrous proteins)”,其特徵為長條且薄的分子,如形成頭髮的 角質,或者組成骨頭和皮膚的膠原蛋白。此種纖維狀的蛋白質為植 物、動物和其他有機體提供了結構支撐的能力。 生物學家將蛋白質分類為四種結構級別,蛋白質的首要結構 (primary structure)為一連串的氨基酸序列,如圖 2.4 所示,此 為最簡單的種類。第二類的結構(secondary structure)為盤繞、 彎曲或多胜肽鏈個體區域的摺疊。例如,α helix 結構,其形狀就如 同電話電線的盤繞狀,如圖 2.5(a)所示。. 圖 2.4 蛋白質鏈的首要結構 第二類結構還包括 β pleated sheet 和膠原蛋白螺旋(collagen helix) 。β pleated sheet 為一長條的鏈,並且前後重覆摺疊,有如 一條扭曲的蛇,或者像是一長串在機場排隊等候的人,如圖 2.5(b) 所示。這類摺疊的結構是由於氫氧鍵所結合在一起,就像是被水分子 吸住一樣。而膠原蛋白螺旋不像 α helix 只有包含一個多胜肽鏈,它 是由三個多胜肽鏈緊密的相互捲曲而成,如圖 2.5(c)所示。. 9.
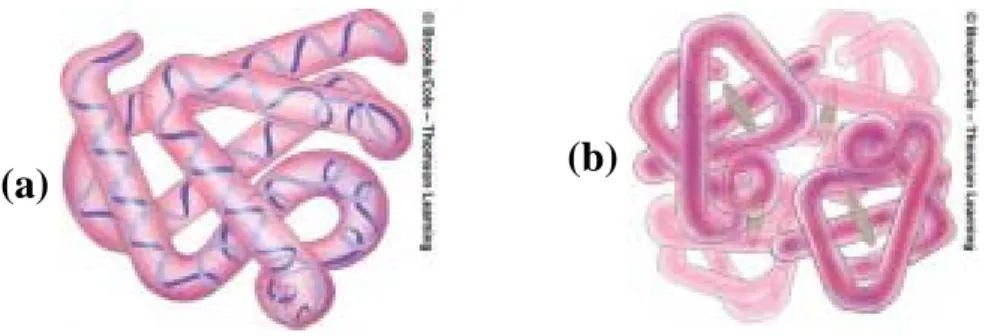
(18) (a) (b. (c). 圖 2.5 第二類蛋白質鏈結構 (a) α helices (b) β pleated sheet (c)Collagen helices 球型蛋白質和纖維型蛋白質均包含著 α helices 和 β pleated sheet 這兩種結構。但是纖維型蛋白質通常有較多的 α helices,而 球狀蛋白是由很多種第二類結構所混合而成。 蛋白質的第三類結構是一個連續的多胜肽鏈作三維的摺疊,如圖 2.6(a)所示。假如你捲曲一段緞帶成為螺旋型,你就可以得到第二類 結構。假如你將這個捲曲的緞帶捆在生日禮物上,這個緞帶就成為了 第三類的結構。 大多數的蛋白質也有第四類的結構,其結構是由兩種或多種的摺 疊鏈相稱結合在一起,如圖 2.6(b)所示,這就類似將不同的緞帶拼 湊成一精巧的蝴蝶結一樣。緞帶可以是同一種或者是不同種顏色的混 合,蛋白質也是可以由同一個或不同的多胜肽鏈組合而成。. 10.
(19) (b). (a). 圖 2.6 (a)第三類蛋白質鏈結構 (b)第四類蛋白質鏈結構. 2.2.2 膠原蛋白結構 膠原蛋白是位於細胞外部纖維狀的蛋白質,並且用來連接兩個組 織。事實上,它在動物體上是以單一種氨基酸型態組合而成的蛋白 質。光是它的種類就有 16 多種,但在動物體上 80%~90%所包含均為 Type I、Type II 和 Type III 這類型態。而這些膠原蛋白的分子都 捆在一起形成一個長薄形的纖維。雖然有著不同種類的蛋白質與結 構,但它們目的卻都是相同的,那就是要幫助組織抵擋組織受力所產 生的扭曲,而膠原蛋白在醫學上的應用如表 2.1 所示 表 2.1 膠原蛋白在生醫材料上的應用 醫. 學. 應. 用. 一般外科. 止血劑、人造皮膚、傷口敷料、尿失禁治療劑. 皮膚科. 皮膚整容及修補、化妝保養品. 骨科. 骨、軟骨及韌帶修補及再生. 口腔外科. 口腔組織修補及再生. 神經外科. 周圍神經修補及再生. 心臟血管外科 血管及心瓣修補 整型外科. 器官組織整形及修補. 藥物遞送. 藥物的局部性遞送 11.
(20) 膠原蛋白的基本結構組成單位是由長 300nm、厚度直徑為 1.5nm 的蛋白質所構成,這個蛋白質包含著三個子單位,分別為兩個 α1( I ) 鏈 和一個 α 2( I ) 鏈。而每個鏈又有著 1050 個氨基酸纏繞在四周,並且形 成右手三重螺旋(Triple helix),如圖 2.7(a)所示。所有的膠原蛋 白最終會顯示成類似三串螺旋片斷結構,而每一種類型的膠原蛋白有 著獨一無二的特性,主要是由於 Triple helix 的中斷節點和三維結 構互相交疊所導致的。 (a). (b). 圖 2.7 膠原蛋白基本結構(a)膠原蛋白之基本結構單元,其型態為一 個 Triple helix 的分子,每個分子長度為 300nm (b)纖維狀的 膠原蛋白是由分子的端點和端點連接之後,再包裹在一起的。 眾多由三個 Type I 組成的膠原蛋白由端點和端點連接之後,再 包裏在一起,其構成的纖維直徑約為 50-200nm。在此纖維之中,鄰 近的膠原蛋白分子與另一個分子移開 67nm,約為本身分子長度的四 分之一,如圖 2.7(b)所示,其顯微影像如圖 2.8 所示。由以上的介 紹可以讓我們了解到,膠原蛋白在結構上有著重複的性質,並且不為 中心對稱結構,所以能夠產生二倍頻的訊號,正因為如此,其二倍頻 訊號的轉換效率很高,因此只要使用較大的瞬間雷射功率,即可輕易 得到其二倍頻訊號。. 圖 2.8 膠原蛋白顯微影像. 12.
(21) 第三章. 研究方法. 在這個章節中,將會開始逐一地介紹本實驗,其介紹的內容依序 為樣本選取、研究架構、研究步驟、研究工具以及資料處理等。 3.1 樣本選取 本實驗所採用的膠原蛋白為 Sigma 公司所生產製造,由於膠原蛋 白的類型有很多種,所以本實驗採用的是 Type I(C-9879, Sigma)型 式,此型式的膠原蛋白是由牛的阿基里斯腱所粹取出來的。而且,此 型式與皮膚組織、骨頭、肌腱的組成結構相同,所以我們採用這類型 式的膠原蛋白來做為本實驗的樣本,讓我們能對平常所知道知識做聯 繫。 由於以往顯微鏡所使用的載玻片是玻璃材料;然而,玻璃的導熱 性不佳,所以我們使用剪裁後的銅片來取代載玻片,使實驗進行時能 夠準確地控制膠原蛋白的溫度。在樣本製作方法方面,我們先將純化 的膠原蛋白浸泡在水中,然後利用微量滴管將浸泡在水中的膠原蛋白 吸出10 µ L ,將其滴於真空變溫系統的加熱銅片上,並且使用無塵紙 將水份吸乾。此時要注意的是,在加熱銅片上的膠原蛋白量不能太多 (只需少許量),如此才不會影響二倍頻訊號的反射。 由於本實驗需要對膠原蛋白加溫,在經歷過一次實驗之後,該膠 原蛋白結構已造成破壞,不適合再進行下一次的實驗,所以要進行下 次實驗時要將樣本換掉,並且清洗已加熱的銅片表面,再換上新的膠 原蛋白樣本。. 3.2 研究架構 在此實驗之中,我們使用正立反射式光學顯微鏡結合一真空變溫 平台來進行二倍頻量測,如圖 3.1 所示。利用超短脈衝雷射做為產生 二倍頻之光源,並搭配自行製作的不鏽鋼支架來重組顯微鏡架構,使 得真空變溫平台(MicrostatHe, Oxford)可以與顯微鏡做結合。. 13.
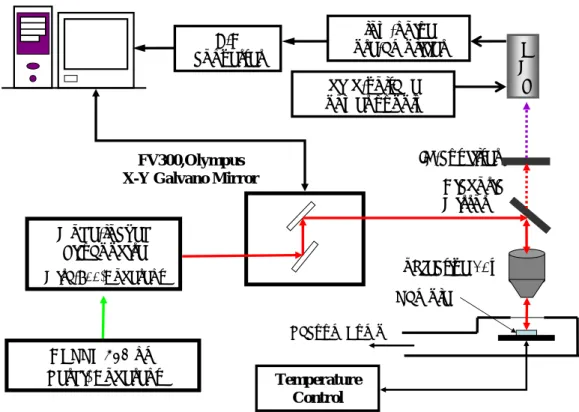
(22) low-noise pre-amplifier. A/D converter. P M T. high-voltage power supply IR-cut filter. FV300,Olympus X-Y Galvano Mirror. Dichoric Mirror. Mode-locked Ti:sapphire. objective 10x. (Mira-900,Coherent). Sample Vacuum Pump. DPSSL 532 nm (Verdi, Coherent). Temperature Control. 圖 3.1 反射式倍頻真空變溫顯微系統架構 在光路部分,以 DPSSL 雷射(Verdi-10, Coherent)激發鎖模鈦藍 寶石(Ti:Sapphire)雷射(Mira-900F, Coherent)之後,產出超短的脈 衝光源,並將此光源導入共焦顯微鏡的掃瞄器內,再經由一面双色鏡 (Dichoric Mirror)將光束經由 10X 物鏡(x10, Olympus)導入樣本之 中,而此樣本必須放置於一個真空變溫系統之內。由樣品產出的訊號 FV300. 真空變溫平台. 光電倍增管. 圖 3.2 反射式倍頻真空變溫顯微系統實體架設圖. 14.
(23) 光反射出真空變溫平台之後,經由二色鏡入射到光電倍增管(PMT) 內,必須在光電倍增管之前加上一塊濾波片,來濾除原本的雷射光 束,進而保留所需的二倍頻訊號。之後,將光電倍增管所收到的二倍 頻訊號經由前置電流放大器放大,最後此倍頻訊號會呈現在電腦螢幕 上,圖 3.2 為實驗實體架設圖。 3.3 實驗步驟 雖然對此實驗設計了兩種不同的量測方式,不過量測的原則均是 相同的。首先,先前的準備工作是:脈衝雷射的熱機與波長的調整, 我們所使用的波長為 800nm 的近紅外光,架設之後並且調整雷射光 路,使得雷射光得以導入到掃瞄器之中。安裝適當的濾波片與光電倍 增管,完成以上步驟之後,要記得將雷射光束由發光處擋住。 著手進行時,先將有放置樣本的加溫銅片安置於真空變溫的平台 之上,並且在加熱銅片上放置一片玻璃的蓋玻片,目的是為了避免此 銅片上的膠原蛋白因為受熱而產生變型,使得對膠原蛋白加熱不均 勻,並且也可以防止所收集到的影像產生變化,如圖 3.3 示。 圖 3.3 膠原蛋白樣品放置完成圖. Pure Collagen Temperature control. Vacuum Chamber. Glass cover. 在完成樣本銅片的放置之後,蓋上真空變溫平台的封蓋,並對此 平台進行抽真空的動作,在經過粗抽和細抽之後,可由真空計讀出真 15.
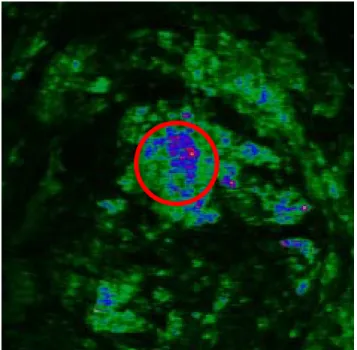
(24) 空度到達約為 4 × 10−5 mbar 時,便可開始進行量測。 由於使用變溫平台的緣故,我們採用長工作距離的特殊 10X 物鏡 (NA=0.25, Olympus),如此可以防止平台玻璃窗因平台移動與物鏡碰 撞而毀損,並且能夠順利的完成量測。此時需要調整三維平移台的高 度,讓物鏡的位置移動到所需的對焦位置,便可將已待機完成的超短 脈衝雷射光束入射到掃瞄器之中,同時啟動儀器開始掃瞄。本實驗所 使用的儀器參數如下,雷射入射光平均功率為 143mW,光電倍增管電 壓 850V,前置電流放大器放大率為 20 倍。 由於使用目鏡對樣本對焦的焦平面和使用光電倍增管收取二倍 頻訊號的焦平面位置有少許的誤差,所以在使用光電倍增管收取訊號 時,還需些許的調整焦平面的位置,直到電腦顯示理想的位置。在掃 瞄時,可以改變變溫平台的溫度參數,以完成所希望的量測結果,此 時所要注意的是:當改變溫度時,要等到整個溫度穩定下來,以確保 樣本已到達所需溫度。. 3.4 資料處理 如圖 3.4 示為一張1024 × 1024 的膠原蛋白二倍頻影像,其頻色的 深淺表示二倍強訊號的強弱,其滿刻度為 4096。利用 Fluoview 程式 中的分析功能,來分析此數據。在此程式中,我們可以設定特定的範 圍做為軟體數據統計的區域,如圖 3.4 的紅色圓圈表示,在此域的強 度可由分析軟體而計算出平均值、總數等。 將相同的樣本,以不同溫度和時間所量測到的二倍頻做量化的動 作,將數值統計出來。再透過下式. I SHGnorm =. I SHG (T ) I SHG (300 K ). 做歸一化的動作。其中 I SHG (T ) 為在 T 溫度下所分析到的強度, 例如: I SHG (300 K ) 為在 300K 時的強度,我們以此強度為一個基準來 進行強度比較, I SHGnorm 為以 300K 當基準後規一化的強度, 接著對這 些強度做圖,可得到第四章所顯示的結果。 16.
(25) 圖 3.4 膠原蛋白之二倍頻影像. 17.
(26) 第四章. 結果與討論. 由於膠原蛋白在低溫時,其二倍頻訊號強度沒有發生什麼變化, 由此証明了一件事,膠原蛋白在低溫的條件之下其結構不會產生變 化。基於這個理由,之後的實驗便沒有再對低溫的部分做研究,只對 高溫的部分做相關的研究。 4.1 倍頻訊號與溫度關係 此測量的方式是將樣本,在不同溫度下條件的量測,我們以 10K 為一個溫度間隔量測,直到二倍頻訊號衰減到無法測量時即結束,經 過數據分析之後,完成的結果如圖 4.1 所示。. 340K. 1.0 Intensity Normalization. 370K. 0.5 420K. 460K. 0.0 300. 350. 400. 450. 500. Temperature(K). 圖 4.1 倍頻強度與溫度關係 由分析圖表可以看出,膠原蛋白在溫度升高超過 340K 時,二倍 頻訊號強度已開始產生變化;當溫度上升到達 370K 時,二倍頻訊號 強度有著較明顯的衰減變化;在溫度超過 420K 時,二倍頻訊號強度 衰減到原始強度的百分之 30,意味著溫度已破壞了多數的結構。在 溫度超過 460K 以後,二倍頻訊號強度就己極微弱,幾乎無法量測。 在完成溫度 500K 的量測後,將樣本的溫度控制調回室溫 300K 時,二 18.
(27) 倍頻訊號並未恢復,此証明了一件事,膠原蛋白的二倍頻訊號是由結 構的對稱性所產生的,因此在結構破壞後,二倍頻不會再次產生。. 4.2 不同溫度下,倍頻訊號隨時間的衰減 由上方的分析數據,使得我們設計了下一個實驗,來討論膠原蛋 白在不同溫度的條件下,其倍頻訊號的衰減速率為何,若它們之間的 衰減速率是不同的,之間又存在什麼關係。 本量測方式是一組樣本只使用一個固定的溫度,以五分鐘為一個 間隔,來量測同一個溫度下,二倍頻訊號的衰減速率。以相同的方式, 改變不同的溫度條件之方式來量測。將不同溫度之下所得到結果與之 前方式做分析之後,其結果如圖 4.2 所示。 其中分析圖表內溫度 420K 與 440K 兩條曲線之可信度不高,此乃 實驗誤差所產生。會產生如此大的誤差是因,升高樣本溫度時,加熱 銅片產生膨脹使得樣本的焦距偏移所至。本量測是由 420K 開始量 測,以每 10K 為一個間隔,由於在 420K 之前的溫度,其倍頻訊號衰 減的速率過慢,也就是說衰減無太大的變化,所以才由 420K 開始量 測,根據此分析圖依然可看出一些相對關係。溫度越高,其二倍頻訊 號隨時間衰減的速率越快,由此即可說明溫度的上升,的確會對膠原 蛋白造成快速的破壞。由此分析圖表所得資訊還無法完全建立一個很 明確的關係,因此使用數學軟體來分析圖表之每一條曲線而做一階指 數擬合,其數學式如(4.1)所示:. y = y0 + A1e − t / t1. (4.1). 其中 y0 、 A1 、t1 為一擬合係數,t 為時間,擬合的結果如圖 4.3 所示。 經過擬合後的曲線讓我們更容易看出每條曲線之間的變化率,不 過由此方法還是無法確實地了解之間的關係,因此我們使用 Arrhenius Equation 來估計此數據。 (上述方程式常被用來分析此相 關問題). 19.
(28) 430K 450K 460K 470K 480K 490K. Intensty Normalization. 1.0. 0.8. 0.6. 0.4. 0.2. 0.0. 0. 50. 100. 150. 200. 250. 300. Time(Min). 圖 4.2 不同溫度條件,倍頻強度隨時間衰減的速率 此乃 Arrhenius 在 1899 年所提出,在化學領域中,最小能量要 歸咎於化學反應。兩個物質之間化學反應發生只有存在於一物質的原 子、離子或分子碰撞到另一物質之原子、離子或分子的情形下才會發 生。在一次反應之中,碰撞的只是占全部的一小部份,這是由於物質 產生互相作用時只有一小部分的百分比存在著最小動能,此動能又為 一分子要產生反應所需之能量。當這種反應物受到碰撞,它便會形成 一過渡產物,這項產物的化學能會比反應物結合後的化學能高。為了 讓此項轉變態能達到進而產生反應,因此必需加入一些反應物之外的 能量,如此反應才會產生,這種能量被稱之為 Activation Energy。 再者,Arrhenius 更提出方程式以計算出 Activation Energy ∆E 。 計算方法如下: 1 t1. =e. −∆E. 20. KT. (4.2).
(29) 430K 450K 460K 470K 480K 490K. Intensity Normalization. 1.0. 0.5. 0.0. 0. 50. 100. 150. 200. 250. 300. Time(Min). 圖 4.3 將圖 4.2 做數學的一階指數擬合(420K 和 440K 除外) 其中 K 為波茲曼常數,T 為各曲線所使用之溫度,將不用曲線擬合出 的 t1 代入(4.2)式之中,便可得到個別曲線之 ∆E ,再將此平均之後得 到的數值為: ∆E = 0.16 ± 0.04eV 故由此實驗可以計算出膠原蛋白的活化能。由此活化能,讓我們更了 解膠原蛋白的化學特性。 接下來我們來回顧一些之前對膠原蛋白的實驗結果, Theodossiou 等人利用倍頻光學方式所得量測結果如圖 4.4 所示,其 倍頻訊號於 40°C 時便開始衰減(Theodossiou et al.,2002)。而 Rochdi 等人利用差分掃瞄熱量儀 DSC 所得結果,也顯示出以 40°C 進 行預熱的程序之後,其膠原蛋白熱穩定性有降低趨勢(Rochdi et al.,1999)。將本實驗結果,與先前之結果比較,可以發現在真空環 境下,膠原蛋白結構熱穩定點約為 67°C(340K) ,而在較高水分子濃 21.
(30) 度環境之下,其熱穩定點為 40°C,此相差大約 27°C,其結果很容易 說明。. 圖 4.4 在一般環境下,膠原蛋白倍頻訊號強度與溫度關係圖 在 Rochdi 等人研究結果顯示,環境中的水分子濃度越高,膠原 蛋白結構的熱穩定性(Td)也會相對地下降,如圖 4.5 所示(Rochdi et al.,1999)。此乃由於組成膠原蛋白的基本結構三重螺旋(Triple helix)中的 α helix 內部氫氧鍵結被打斷,導致整個三重螺旋結構受 到破壞,而受到溫度破壞的膠原蛋白便無法產生非線性光學倍頻的條 件。至於影響膠原蛋白結構的熱穩定性不僅只有環境中水分子濃度這 個變數,還包括了環境介質中的酸鹼度和結構交互結合程度等(Alina 2005)。. 圖 4.5. 環境中的水分子濃度與熱穩定點 Td(○)關係圖 22.
(31) 第五章. 結論. 藉由此光學倍頻顯微術的架設,讓我們更進一步地了解到膠原蛋 白之特性。透過溫度和時間的改變,使我們能夠以即時的方式來監測 結構上的改變。再經由更進一步地量測方式,量測在不同溫度之下, 倍頻光強度隨時間的衰減速度,讓我們發現在什麼溫度條件下,結構 上破壞的速度會加快。不僅如此,再經過一些數學上的曲線擬合和 Activation Energy 的計算,將此一結果與化學相關領域連上了關 係。由於本實驗所採用的膠原蛋白為 Type I 型式,是在動物體上最 為常見的,讓此研究的結果更具普遍性。 比較以往的實驗結果,其結果與本研究結果不同之處在於,本實 驗將膠原蛋白放置於真空的環境,而之前研究進行方式是將膠原蛋白 直接放置於水中加熱一段時間,因此膠原蛋白結構的熱穩定點也不相 同,水分子濃度與結構的熱穩定性有著很緊密的關係也再一次被証實 出。. 23.
(32) 參考文獻 Allan J. Tobin, Jennie Dusheck, 2004, Asking about life, Third edition. Bloembergen, N., 1965. Nonlinear Optics. New York. Bottomley, D.J., Mito, A., Niki, S., Yamada, A., 1996. Second-harmonic generation from chalcopyrite-structure semiconductor thin films. Optics Letters 21, 254-257. Bouevitch, O., Lewis, A., Pinevsky, I., Wuskell, J.P., Loew, L.M., 1993. Probing membrane potential with nonlinear optics. Biophysical Journal 65, 672-679. Boyd Robert W., 2003, Nonlinear Optics, second edition. Dolino, G., 1973. Direct observation of ferroelectric domains in TGS with second-harmonic light. Applied Physics Letters 22, 123-124. Dolino, G., Bachheimer, J.P., Vallade, M., 1973. Direct observation of Dauphine twins in quartz with second-harmonic light. Applied Physics Letters 22,623-625. Franken, P.A., Hill, A.E., Peter, C.W., Weinreich, G., 1961. Generation of optical harmonics. Physics Letters 22, 123-124. Fraser RDB, Macrae TP. 1979, The crystalline structure of collagen fibrils in tendon. Journal Mol Biol 127, 129–33. Freund, I., Deutsch, M., Sprecher, A., 1986. Connective tissue polarity optical second harmonic microscopy, crossed-beam summation and small-angle scattering in rat-tail tendon. Biophysical Journal 50, 693-712. Gannaway, J., Sheppard, C.J.R., 1978. Second-harmonic imaging in the scanning optical microscope. Optical and Quantum Electronics 10, 435-439. Gauderon, R., Lukins P.B., Sheppard C.J.R., 1998, Second-hamonic generation imaging using femtosecond laser pulses. Optics Letters 23, 1209-1212. Gauderon, R., Lukins P.B., Sheppard C.J.R., 2001, Optimization of second-harmonic generation microscopy. Micron 32, 691-700. Guo, Y., Ho, P.P., Savage, H., Harris, D., Sacks, P., Schantz, S., Liu, F., Zhadin, N., Alfano, R.R., 1997. Second-harmonic tomography of tissues. Optics Letters 22, 1323-1325. Guo, Y., Ho, P.P., Tirksliunas, A., Liu, F., Alfano, R.R., 1996. Optical harmonic generation from animal tissues by the use of picosecond and femtosecond laser pulses. Applied Optics 35, 6810-6813. Harvey Lodish, Arnold Berk, S. Lawrence Zipursky, Paul Matsudaira, David 24.
(33) Baltimore, JamesDarnell, 2002, Molecular Cell Biology, Fourth edition. Hellwarth, R., Christiansen, P., 1974. Nonlinear optical microscopic examination of structure in polycrystalline ZnSe. Optics Communications 12, 318-322. Janz, S., Pedersen, K., van Driel, H.M., 1991. Dispersion and anisotropy of the optical second-harmonic response of single-crystal Al surfaces. Physical Review B 44, 3943-3946. Jentsch, T., Jupner, H.J., Ashworth, S.H., Elsaesser, T., 1996. Second-order nonlinearities of polycrystalline molecular films studied on a 20-fs time scale. Optics Letters 21, 492-494. Lewis, A., Khatchatouriants, A., Treinin, M., Chen, Z., Peleg, G., Friedman, N., Bouevitch, O., Rothman, Z., Loew, L. Sheres, M., 1999. Second-harmonic generation. of. biological. interfaces:. probing. the. membrane. protein. bacteriorhodopsin and imaging membrane potential around GFP molecules at specific sites in neuronal cells of C. elegans. Chemical Physics 245, 133-144. Miles CA, Burjanadez TV, Bailey AJ., 1995, The kinetics of the thermal denaturation of collagen in unrestrained rat tail tendon determined by differential scanning calorimetry. Journal Mol Biol 245, 437–46. Moustaizis, S.D., Papadogiannis, N.A., Fotakis, C., Farkas, G., Toth, C., 1995. Generation and tuning of second-harmonic radiation produced by ultrashort dye laser pulses from a gold surface. Applied Physics Letters 67, 3239-3242. Rochdi A, Foucat L, Renou JP., 1999, Effect of thermal denaturation on water collagen interactions: NMR relaxation and differential scanning calorimetry analysis. Biopolymers 50, 690–696. Shnyrov VL, Lubsandirzhieva VC, Zhadan GG, Permyakov EA., 1992, Multi-stage nature of the thermal denaturation process in collagen. Biochem Int 26, 211–17. Theodossiou T., Rapti G.S., Hovhannisyan V., Georgiou E., Politopoulos K., D. Yova, 2002, Thermally induced irreversible conformational changes in collagen probed by optical second harmonic generation and laser-induced fluorescence. Lasers Med Sci 17, 34-41. Tiktopulo EI, Kajava AV.,1998, Denaturation of type I collagen fibrils is an. 25.
(34) endothermic process accompanied by a noticeable change in the partial heat capacity. Biochemistry 37, 8147–52. Vydra, J., Eich, M., 1998. Mapping of the lateral polar orientation distribution in second-order nonlinear thin films by scanning second-harmonic microscopy. Applied Physics Letters 72, 275-277.. 26.
(35) 附錄 A 20 種氨基酸化學結構 Amino acids with basic side chains Lysine. Arginine. Amino acids with acidic side. Histidine. Aspartic acid. Glutamic acid. Amino acids with polar uncharged side chains. Asparagine. Glutamine. Serine. Threonine. Tyrosine. Amino acids with nonpolar side chains Glycine. Alanine. Valine. Leucine. Phenylalanine. Methionine. Cysteine. Proline. 圖 A1 20 種氨基酸化學結構 27. Isoleucine. Tryptophan.
(36) 附錄 B 雷射光源系統 在雷射光源方面主要由兩部分構成,第一部分是激發光源,使用 由 Coherent 公司生產的倍頻半導體激發固態雷射(Diode pump solid state laser 簡稱 DPSSL),Verdi。第二部份是超快光源,同樣使用 由 Coherent 公司生產的 Mira 900 鈦藍寶石雷射﹙Ti︰sapphire Laser﹚ ,藉由此兩種雷射的搭配以產生超短脈衝的雷射光束。 B1 激發雷射 我們所使用的雷射 Verdi,是由一個波長 807.5nm 的半導體雷射 ,產生波長 陣列激發超快雷射晶體 Neodymium Vanadate﹙Nd︰YVO4﹚ 為 1064nm 的近紅外光雷射,再經過 LBO 倍頻晶體,倍頻為波長 532 的綠光雷射。其輸出功率最高為 10W,光點大小為 2.25mm。內部構造 如下圖 B1 所示。. 圖 B1 Verdi 內部結構圖 B2 超短脈衝雷射 我們使用 Verdi-10V 做為鎖模超快雷射的激發光源,藉由此光源 來激發鎖模鈦藍寶石雷射(Mira-900F, Coherent),而所激發出的雷 射光束為一個超短雷射脈衝,其脈衝寬度約為 150fs,雷射中心波長 可調範圍為 710nm∼1000nm 之間如圖 B2 所示。但由於實際波長的可 調範圍侷限於雷射共振腔內的鏡片組,使得雷射實際可調範圍約在 28.
(37) 720nm∼920nm 之間。其雷射內部共振腔的結構如圖 B2 所示。. 圖 B2 Verdi 激發 Mira 900 輸出功率隨波長調整曲線圖,此脈衝雷射 的重複頻率為 76MHz,所謂的重複頻率是指單位時間的脈衝數目. 圖 B3 Mira 900 內部結構圖 一般鎖模的方法分為主動式鎖模(Active mode-locking)以及被 動式鎖模(Passive mode-locking)兩種。而我們所使用的鈦藍寶石超 短脈衝雷射為使用被動式克爾透鏡鎖模(Kerr lens mode-locking) 的方式,此技術是近幾年所新開發出來的,它是利用材料非線性克爾 29.
(38) 效應來產生自聚焦的效果,在以針孔保留中間較強的部份,去除周圍 弱光的部份,因而達到波型整形的作用,便可產生超短脈衝雷射。 所謂的克爾效應,指的是介質的折射率隨光強度而作非線性的改 變,如下式所示 n = n0 + n2 I 其中 I 是光的強度, n0 是無入射光時的折射率, n2 為非線性係數。由 上式可知,光的強度愈強時,所產生的折射率也會變高,所以雷射光 束強度為高斯分佈時,則光束將會往中心集中,此種效應稱之為自我 聚焦。 當激發光源 Verdi-10V 使用平均功率 8W 作為激發光時,此時的 鎖模鈦藍寶石雷射所產生的脈衝雷射其平均功率約為 1.1W,再經由 光學量測系統的衰減之後,在物鏡處所量測到的平均光功率為 143mW,換算之後每個雷射脈衝的瞬間功率約為 1.88nJ/puls 左右, 若脈衝寬度為 150fs,則瞬間功率可高達 12.5KW,如此高能量的脈衝 能量再經由透鏡聚焦之後,不但足夠產生非線性效應,還要小心樣品 和光學元件被雷射光束破壞。. 30.
(39) 附錄 C 電子偵測系統 本實驗所使用顯微鏡為正立反射式結構,由於 Phase Matching 的先天限制,所以二倍頻的訊號以穿透式的方式較為容易接收,但是 因為結合了變溫平台的緣故,必須改用反射式的結構來接收倍頻訊 號。由於接收膠原蛋白所產生的倍頻訊號在加熱銅片之反射光,造成 光電倍增管所收到的訊號大小會比穿射式的少很多。不過由於脈衝雷 射光束的能量很高,所以產生的二倍頻訊號強度也很高,才得以讓使 用光電倍增管所收到的訊號不至於太弱。 C.1 光電倍增管 光電倍增管(PMT)是一種最常被使用的基本光學偵測元件,其偵 測範圍包括紫外(Ultraviolet)、可見光(Visible)、近紅外線(Near Infrared)。光電倍增管(PMT)具有很高的電流放大率和低雜訊,可說 是極靈敏偵測器之一。在適當操作的修件下,可偵測低達10−19 瓦的功 率。 Focusing electrode Photocathode. Vacuum envelope. Dynodes. D3. D1. D5. D7 Anode. D2. D4. iA. A. D6. A. Faceplate V1 R1. R2. V2 R3. V3 R4. V4 R5. V5 R6. V6 R7. +V A. V7 R8. 圖 C1 頭端入射型光電倍增管結構 本實驗所使用的 PMT 為 Hamamatsu 公司所生產,其型號為 R376, 屬於頭端入射型,如圖 C1 所示。其玻璃內部為一真空環境,目的是 為了防子電子運動時,受到空氣粒子的阻礙。當光線入射到光陰極 31.
(40) (Photocathode)後,光陰極射出電子到真空之中。這些由光所產生的 電子藉由電壓聚集之後,直接入射到電子放大的區域以產生二次電 子,經過幾次電子數量的放大之後,由陽極接收所產生出的電子,並 且輸出訊號。因為是利用二次電子的倍增,使光電倍增管獨特的靈敏 度得以顯露,因此它成為在測量紫外光、可見光和紅外光區域常用的 光電感測元件。而光電倍增管的另一項特色為有著快速的時間響應和 低雜訊。 在倍增管內部電子倍增的部份包括了各種設計,以滿足不同的需 求,例如:快速的時間響應、受光位置的均勻性、大電流脈衝的輸出、 防 磁 場 干 擾 等 。 而 我 們 所 使 用 的 R376 其 子 電 倍 增 部 份 是 使 用 Box-and-grid 這種形態。此種形態是由一連串四分之一圓柱殼所構 成的電極板,一般是常用於頭端入射型的光電倍增管內,因為他的電 極板設計容易,並且改善了受光處激發光電子的均勻性;然而,卻在 某些應用上,時間響應可能會太慢。 在光陰極材質的部份,R376 所使用的是 Multialkali (Na-K-Sb-Cs)。 此種材料的光陰極有著很高並且廣闊的光譜響應,其範圍為紫外光到 近紅外光之間的光譜,並且廣泛地使用在寬頻帶的分光光度計 (Spectrophotometers)和光子計數的應用。利用此特殊的光陰極處 理,可將長波長的響應延伸至 930nm。. C.2 低噪音前置電壓放大器(Low-Noise Voltage Preamplifier) 本實驗所使用的低噪音前置電壓放大器為 Stanford Research Systems 公司所生產的 SR560。二倍頻訊號藉由 PMT 接收到後,轉換. 圖 C2 低噪音前置電壓放大器前視圖 32.
(41) 為電子訊號之後,藉由低噪音的前置放大器將電子訊號放大,並且可 選擇 50Ω 的輸出阻抗與 AD Converter 達到阻抗匹配,以減少訊號的 反射而降低了訊號強度,其前視圖如圖 C2 所示。. C.3 高壓電源供應器(High Voltage Power Supply) 本實驗所使用的高壓電源供應器為 Stanford Research Systems 公司所生產的 PS350。此種的電源供應器可以提供 DC50V~5KV 之間的 電壓,而在此實驗所扮演的角色為提供光電倍增管所需的高電壓,並 且產生的高壓穩定性很高,平均電壓縺波率只有滿刻度的 0.0015%, 其前視圖如圖 C3 所示。. 圖 C3 高壓電源供應器前視圖. 33.
(42) 附錄 D 真空變溫系統 此真空變溫系統(MicrostatHe, Oxford)如圖 D1 所示。將一組低 溫恒溫器放置於一真空的腔體之內,其腔體的真空度可到達約. 4 × 10−5 mbar ,當使用液態氦作為冷卻時,其低溫可降至 2.2 K 的絕 對溫度。此種特別的設計是為了搭配顯微鏡使用,它有著小工作距離 和很薄的窗口玻璃,薄的窗口玻璃可以減少光學上的球面像差。而此 平 台 也 可 應 用 於 下 列 用 途 : 冷 光 的 研 究 、 拉 曼 散 射 (Raman Scatting)、反射和吸收的研究、掃瞄光學顯微鏡和微探針之測定等。 此系統還有另一項優點,就是可以很精準的控制溫度,其內部有 三組溫度感測器來監視溫度的變化,因此可溫度的最小刻度為 0.1K, 而溫度穩定時的變動範圍為+/-0.1K,再者,溫度經調整之後,改變 的速度很快,以減少溫度變化時對樣本特性所造成的改變。. 圖 D1 真空變溫平台結構圖. 34.
(43) 附錄 E 光學元件 E.1 反射鏡、分光鏡和紅外線濾光片 我們將反射鏡、分光鏡和紅外線濾光片製作成下表,在表中我們 將會簡述各種鏡片之功用並列出其應用光譜範圍。 元件名稱. 功用. 特性. 為了將雷射光束. 工作波長為 700nm~950nm,最大的 雷射功率承度為在 1000W / cm 2,在 10nsec 的脈衝雷射之下為 2nJ / cm 2 ,並且其反射 率大於 99%。. 順利的導至我們 所需要的位置, 反射鏡的選擇是. 反射鏡. 必要要主意,要 考慮到反射鏡的 使用波長、入射. 應用光譜範圍. 角度、最大雷射 功率承受率等參 考因素. 分光鏡. 波長在 650nm 以上的. 本實驗所使 用的二色分 光鏡光譜,它 的功用為對 某個波段的 光可以穿 透,剩下的波 段則為反射. 它與一般的分光鏡 (Beam Splitter)的 不同之處在於,一舨 的分光鏡其訊號的損 失太大,因此訊噪比 也會大大的降低,因 此才使用此類型的光 學元件. 100. 光都無法通過。所以. 800nm 的近紅外光濾 除,讓二倍頻 400nm. Transmittance(%). 可以將原本雷射光源. 80. 60. 40. 20. 的光穿過濾波片。. 0 300. 400. 500. 600. 700. 800. 900. 1000. Wavelength(nm). 100. 80. Transmittance(%). 紅外線濾光 片. 使用光電倍 增管接收二 倍頻訊號 時,為了避免 接收到原始 的光源訊號 使得光電倍 增管受損,所 以我們使用 一個帶通濾 波片. 60. 40. 20. 0 200. 300. 400. 500. 600. 700. Wavelength(nm). 35. 800. 900. 1000.
(44) E.2 隔離器(Optical Isolators) 我們所使用的隔離器是由 1845 年法拉第所發現,其利用法拉第 效應(Faraday effect),其偏轉角如下:. φ = VlB 對於給定的介質,偏振面旋轉角 φ 與介質的長度 l 和磁感應強度 B 成 正比,而V 為介質的比例常數。其工作原理如圖 E1 所示,在雷射光 經第一個 BSP 時,其穿透光的極化方向不變,緊接著入射到晶體之 中,此晶體會充當光的極化而做一個旋轉的動作然後輸出。若在這個 光路上有從原光路反射的光,此反射光再一次進入隔離器時,經過晶 體後,極化方向又再一次旋轉到達 BSP 時,其極化方向的反射光無法 通過,所以完全地將反射的雜光隔離掉。藉此可避免回饋光進入電射 共振腔內,造成相位的擾亂,而無法鎖模。. 圖 E1 隔離器內部光路結構圖. 36.
(45) 附錄 F 掃描式共焦顯微系統 由於實驗上的需求,我們使用 Olympus 公司所生產的生物用共焦 掃描顯微系統 FV300,如圖 F1 所示,它與一般工業用的顯微鏡不同 之處,在於生物用顯微鏡著重於生物螢光訊號的量測,所以不論在光 源的入射或螢光的接收,都有著不同型式的濾光片,以滿足生物研究 領域的需求。而共焦顯微鏡的出現,改變了以往傳統光學顯微鏡的缺 點。傳統的光學顯微鏡在觀測較細微結構時,由於受到光波繞射的限 制,使得它無法提供良好的光學解析度,而且其成像方式依舊是停留 在平面成像的階段。雖然使用傳統螢光顯微鏡時,亦會產生相同的問 題,在生物體經由螢光染色之後,無法觀測到生物體的某一斷面,以 致於螢光影像的清晰度不佳,無法確切的分辨出螢光染色部份位於何 處;雖然薄切片的技術也因此發展了出來,但是依然無法做觀測活體 的樣品。這些問題直到 1969 年時,世界上第一部共焦顯微鏡設計完 成,再加上目前雷射技術的進步,以上問題才得於迎刃而解。近十幾 年來,無論是雷射技術或者是電腦都有著驚人的進展,使得共焦顯微 術發展更為完備。. 圖 F1 FV300 雷射光束掃瞄系統 而我們實驗室發展方向是顯術量測技術,利用商用的共焦顯微. 37.
(46) 鏡,配合實驗室自行設計、改裝的顯微鏡基台模組,再加上不同的實 驗架構,使得整個系統不儘具有生物螢光測的功能,更可以成為新的 量測機台,如:光致電流顯微量測技術、光的倍頻顯微量測技術。 我們所用的 Fluoview 影像掃描系統是由 XY galvanomirrors 和步進 馬達組成,XY galvanomirrors 能快速做 XY 平面之掃瞄,平均取一 張1024 × 1024 像素大小的影像,只需數十秒的時間。配合 Z 軸的步進 馬達,其解析度可達 0.1µ m 的 Z 軸移動,因此可對活體樣本做光學切 片。. 38.
(47)
數據
+4


Outline
相關文件
The Centre for Learning Sciences and Technologies (CLST), The Chinese University of Hong Kong (CUHK) launched the!. EduVenture ® learning system, which has been well received by
Income is generated from wages and salaries, interest, rent and profits. In a labour-intensive industry such as tourism, a large part of income comes from wages and salaries earned
However, the PDE of arithmetic MAL has never been derived and the strike based on all past moving-average term is very di¢ cult to value by …nite di¤erence approach.The second, we
A good game helps students to enhance their learning techniques, such as learning by doing, learning from mistakes, goal-oriented learning, discovery learning, task-based
The second part is to the interactions between the brightness, color temperature, and other performance of the bulb and the fabricating parameters such as: the filling volume of
Classifier which uses OpenCV (Open Source Computer Vision Library) was a detector that has been training for face detecting.. The result of the experiment is 2 frames per second, and
ii. Drama as a Second Language: a Practical Handbook for Language Teachers. Cambridge: National Extension College Trust. Drama Techniques in Language Learning: a Resource Book
This kind of algorithm has also been a powerful tool for solving many other optimization problems, including symmetric cone complementarity problems [15, 16, 20–22], symmetric
