G PROTEIN AND ADENYLATE CYCLASE COMPLEX-MEDIATED SIGNAL
TRANSDUCTION IN THE RAT HEART DURING SEPSIS
Li-Ling Wu,* Shaw-Lang Yang,
†Rei-Cheng Yang,
†Hseng-Kuang Hsu,
†Chin Hsu,
†Lin-Wang Dong,
‡and Maw-Shung Liu
‡*Department of Physiology and Pathophysiology, Peking University Health Science Center, China;
†Department of Physiology, Kaohsiung Medical University, Kaohsiung, Taiwan; and
‡Department of
Pharmacological and Physiological Science, Saint Louis University School of Medicine,
St. Louis, Missouri
Received 20 Sep 2002; first review completed 18 Oct 2002; accepted in final form 19 Dec 2002
ABSTRACT—Changes in the protein level of various subunits of GTP-binding protein and the activity of adenylate cyclase in the rat heart during different phases of sepsis were studied. Sepsis was induced by cecal ligation and puncture (CLP). Experiments were divided into three groups: control, early sepsis, and late sepsis. Early and late sepsis refers to those animals sacrificed at 9 and 18 h, respectively, after CLP. The protein levels of various subunits of GTP-binding protein were determined by Western blot analysis. The activity of adenylate cyclase was measured based on the rate of formation of cAMP from [␣-32P]ATP. The results show that protein levels of G␣s and Gremained stable during the early and the
late phases of sepsis. The protein levels of G␣i-2 and G␣i-3 remained relatively unaltered during the early phase of sepsis, but they were increased by 46.5% (P < 0.05) and 61.3% (P < 0.01), respectively, during the late phase of sepsis. The basal adenylate cyclase activity remained unchanged during the early phase while it was decreased by 25.7% (P < 0.05) during the late phase of sepsis. The isoproterenol-stimulated adenylate cyclase activity was unchanged during early sepsis while it was decreased by 44.6% (P < 0.01) during late sepsis. These data demonstrate that during the late hypodynamic phase of sepsis, myocardial G␣i-2 and G␣i-3 protein levels were increased and the increases were coupled with a reduction in adenylate cyclase activity. Because GTP-binding proteins mediate sympathetic control of cardiac function, the present findings may have a pathophysiological significance in contributing to the understanding of the pathogenesis of cardiac dysfunction during the late stage of sepsis.
KEYWORDS—GTP-binding proteins, signal transduction, septic shock, myocardial dysfunction, heart failure
INTRODUCTION
Sympathetic control of cardiac function is mediated through
GTP-binding proteins. GTP-binding proteins are composed of
a large family of highly homologous proteins that couple more
than 100 different receptors with different effector enzymes
(1–3).

-adrenergic receptor (

AR;

1AR and

2AR) and
␣
-adrenergic receptor (
␣
AR;
␣
1AR) are coupled to G
␣
s and
Gq subunits, respectively, whereas
␣
2AR is coupled to G
␣
i
subunit of GTP-binding proteins (1, 4–6). There is direct
evidence that organ-specific alterations in GTP-binding protein
expression contribute to the pathogenesis of heart diseases in
human and animal models. Changes in G
␣
s and G
␣
i protein
levels and the gene transcripts encoding them are found to be
associated with heart failure, ischemic cardiomyopathy,
hyper-trophy, and hypertension (7–13). In shock and sepsis, G
␣
i
levels are found to be elevated in the heart of
catecholamine-refractory septic shock patients (14) and in the neonatal rat
heart myocytes upon exposure with the plasma of
noradrena-line-treated septic shock patients (15). Because the available
data in the literature regarding changes in myocardial
GTP-binding proteins are limited to the hypodynamic phase and
because sepsis is a two-phase process in which patients initially
go through a hyperdynamic phase and, subsequently, the
hypo-dynamic phase, it is important that an animal model capable of
exhibiting a biphasic septic syndrome be adapted for the
inves-tigation of the role of GTP-binding proteins in the pathogenesis
of heart dysfunction in septic shock. Furthermore, because
previous work from this laboratory has revealed that

AR and
␣
AR undergo a biphasic expression in the rat heart during the
two distinct cardiodynamic phases of sepsis (16–18), the
present work dealing with altered G protein/adenylate cyclase
system was undertaken to clarify changes in postreceptor
events in an attempt to understand the pathogenesis of
myocar-dial dysfunction during the progression of sepsis.
MATERIALS AND METHODS
Animal model
All animal experiments in this study were performed with the approval of the Animal Care Committee of Saint Louis University School of Medicine and in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Male Sprague-Dawley rats weighing from 270–320 g were used. All animals were fasted overnight with free access to water before the induc-tion of sepsis. Sepsis was induced by cecal ligainduc-tion and puncture (CLP) as described by Wichterman et al. (19) with minor modifications (20). Under halothane anes-thesia, a laparotomy was performed (the size of the incision was 2.5 cm), and the cecum was ligated with a 3-0 silk ligature and punctured twice with an 18-gauge needle. The cecum was then returned to the peritoneal cavity and the abdomen was closed in two layers. Control rats were sham operated (a laparotomy was performed and the cecum was manipulated but neither ligated nor punctured). It should be mentioned that the values of the biochemical and the molecular biological indexes (GTP-binding protein levels and adenylate cyclase activities) that we reported in this study were indistinguishable between the sham-CLP and the nonsurgical controls. All animals were resuscitated subcutaneously with 4 mL of 0.9% NaC1/100 g body weight at the completion of surgery and also at 7 h postsurgery. Animals were fasted but had free access to water after operative procedure. Experiments were divided into three groups: control, early sepsis, and late sepsis. Early and late sepsis refers to those animals sacrificed at 9 and 18 h, respectively, after CLP. Previous experi-Address reprint requests to Dr. Rei-Cheng Yang, Department of Physiology,
Kaohsiung Medical University, Kaohsiung, Taiwan.
This work was supported by the National Heart, Lung and Blood Institute (grant no. HL-30080) and the National Institute of General Medical Sciences (grant no. GM-31664), the National Institutes of Health.
DOI: 10.1097/01.shk.0000055816.40894.cd
ments have indicated that myocardium was in the hyperdynamic state during the early phase, whereas it was in the hypodynamic state during the late phase of sepsis (18). The mortality rates were 0% for control, 4% for early sepsis, and 27% for late sepsis.
Preparation of cardiac sarcolemmal membranes
Rat heart sarcolemmal membranes were prepared according to a procedure previously used in this laboratory (16, 17). Hearts removed from control and septic rats were weighed, freeze-clamped with aluminum clamps precooled in liquid nitro-gen, and pulverized with a mortar and pestle precooled in liquid nitrogen. The powdered tissues were thawed and homogenized with Tekmar Tissumizer (Model SDT) in 5 vol of buffer A (50 mM NaH2PO4/Na4P2O7, pH 7.4; 10 mM EDTA; 25 mM NaF; 1g/mL soybean trypsin inhibitor; 1 g/mL aprotinin; 0.75 g/mL pepstatin A; and 2g/mL leupeptin). The homogenates were centrifuged at 14,000 g for 20 min. The resulting pellets were suspended in buffer B (0.6 M NaCl, 50 mM NaH2PO4/Na4P2O7, pH 7.4; 10 mM EDTA; and 25 mM NaF), rehomogenized, and recentrifuged at 14,000 g for 20 min. The pellets from the second centrifugation were suspended in buffer A, homogenized three times (each time for 30 s), and centrifuged at 2300 g for 15 min. The resulting pellets were resuspended, rehomog-enized, and recentrifuged at 2300 g for 15 min. All homogenizations throughout were performed with Tekmar Tissumizer. The 2300-g supernatants were combined and centrifuged at 79,700 g for 40 min. The 79,700-g pellets were suspended in 1 M sucrose dissolved in buffer C (0.3 M NaCl, 50 mM NaH2PO4/Na4P2O7, and 100 mM Tris-HCl, pH 7.4). Ten milliliters of this suspension were layered at the bottom of a discontinuous sucrose gradient consisting of 9 mL of 0.6 M sucrose (dissolved in buffer C) and 9 mL of 0.25 M sucrose (dissolved in 10 mM histidine, pH 7.4). The gradients were centrifuged at 254,000 g for 70 min. Fractions at 0.6:0.25 M sucrose interfaces were collected, diluted with buffer C, and then centrifuged at 162,600 g for 40 min. The final pellets were suspended in a buffer containing 0.25 M sucrose and 30 mM histidine (pH 7.4), stored at −80°C, and then used as sarcolemmal membranes. The entire procedure was performed at 4°C unless other-wise stated.
Determination of GTP-binding protein levels by Western
blot analysis
Western blot analysis was performed according to the method of Ausubel et al. (21) with minor modification (22). Sarcolemmal membranes containing various amounts of protein (100g for G␣s, 100 g for G␣i-2, 100 g for G␣i-3, and 40 g for -subunit) were denatured and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (10–20% sulfate-polyacrylamide gradient gel). Using the Bio-Rad Mini Trans-Blot Electrophoretic Transfer Cell, proteins separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis were transferred to a polyvinylidene fluoride membrane (Bio-Rad) at 8 mA (constant current) for 14–16 h in transfer buffer (25 mM Tris base, pH 8.3; 192 mM glycine; 5% [vol/vol] methanol). Nonspecific binding sites were blocked with 10% (wt/vol) of nonfat dry milk in Tris-buffered saline (TBS: 20 mM Tris-HC1, pH 7.4; 137 mM NaC1) for 4 h at 4°C. Blots were washed three times (10 min each) with TBST (TBS containing 0.5% Tween-20) followed by incubation with a 1:1000 dilution of a polyclonal antibody recognizing G␣s, G␣i-1 and G␣i-2, G␣i-3, or G subunit (Calbiochem-Novabiochem International, San Diego, CA) for 5 h at room temperature. Subse-quently, the blots were washed three times (10 min each) with TBS and then incubated with a 1:2,000 dilution of an antirabbit immunoglobulin, peroxidase-linked species-specific whole antibody (Amersham Life Science) for 1 h at room temperature. The blots were then washed four times (10 min each) with TBST, followed by incubation with enhanced chemiluminescent (ECL) Western blotting detection reagent (Amersham Life Science), and finally exposed to Hyperfilm-ECL (Amersham Life Science) for 40 s. Autoradiographs were scanned with Hewlett-Packard ScanJet 4C Scanner, and the relative densities were quantified by Jandel Scientific Software program (Sigma Gel). It should be mentioned that the quanti-tation of immunoblots was performed by comparing each individual subunit of GTP-binding protein to the control only for that particular blot. For each subunit, six separate blots were performed. Each blot consisted of control, early septic, and late septic samples in equal amount of protein. Preliminary experiments have indicated that the control levels (six control samples prepared from six different animals and performed on the same blot) for each subunit were almost identical.
Assay of adenylate cyclase activity
Adenylate cyclase activity was determined by a method previously used in this laboratory (22) with modification. The reaction mixture in a final vol of 200L contained 50 mM Tris-HC1, pH 7.5; 5 mM MgCl2; 5 mM creatine phosphate; 0.4 mg/mL creatine phosphokinase; 1 mM cAMP; 1 mM dithiothreitol; 1 mM [␣-32P]ATP (2 × 106counts/min). The reaction was initiated by the addition of sarcolemmal membranes containing 100g of protein and was allowed to proceed for 8 min at 37°C. The reaction was terminated by the addition of 0.2 mL of a solution containing 2% sodium dodecyl sulfate; 40 mM ATP; 50 mM Tris-HCL, pH 7.5; 1.4 mM [2,8-3H]cAMP (20,000 counts/min). The reaction mixture was then diluted with 0.6 mL of deionized water and was decanted into Dowex columns. The eluate from this and two subsequent 1-mL washes was discarded. Three milliliters of water were then added to each column and the eluate was collected in a test tube containing 0.2 mL of 1.5 M imidazole (pH 7.5). After mixing, the contents of each tube were poured into a second column containing neutral alumina pre-equilibrated with 0.1 M imidazole HC1 (pH 7.5). The eluate of the alumina column was collected directly into a scintillation vial and then counted by a liquid scintillation counter. It should be mentioned that [2,8-3
H]cAMP was included in the stop solution to deter-mine the recovery of cAMP from the columns.
Other assays
(Na+-K+)-ATPase, Ca2+-ATPase, and glucose-6-phosphatase activities were determined as described previously by us (22). The protein concentration of sarco-lemmal membranes was determined by the method of Lowry et al. (23).
Statistical analysis
The statistical analysis of the data was performed using one-way ANOVA followed by Student-Newman-Keuls tests. A P value of less than 0.05 was accepted as statistically significant.
Materials
[␣-32P]ATP (650 Ci/mmol) and [2,8-3H]adenosine-3⬘,5⬘-cyclic monophosphate (38 Ci/mmol) were purchased from ICN Pharmaceuticals, Inc. Horseradish peroxi-dase-labeled second antibody conjugate (donkey anti-rabbit), ECL Western blotting detection reagent, and Hyperfilm were obtained from Amersham Life Science. ATP, cyclic AMP, creatine phosphate, and creatine phosphokinase were purchased from Sigma Chemical Co. Purified primary antibodies, including rabbit anti-G␣s subunit, C-terminal (385-394); rabbit anti-G␣i-1 and G␣i-2 subunits, C-terminal (345-354); rabbit anti-G␣i-3 subunit, C-terminal (345-354); and rabbit anti-G-protein  subunit, internal (127–139) were supplied by Calbiochem-Novabiochem Interna-tional. Other chemicals and reagents were of analytical grade.
RESULTS
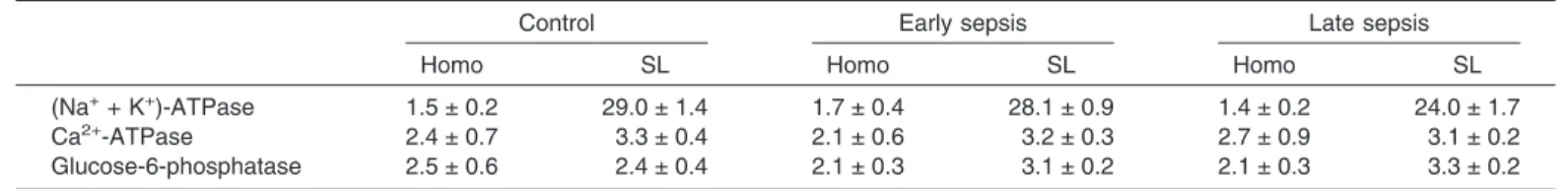
Table 1 shows marker enzyme activities of heart
sarcolem-mal membranes isolated from control and septic rats. (Na
+-K
+)-ATPase served as marker for sarcolemmal membranes,
while Ca
2+-ATPase and glucose-6-phosphatase served as
markers for sarcoplasmic reticulum. In sarcolemmal
membranes, (Na
+-K
+)-ATPase activities were enriched 17- to
19-fold whereas Ca
2+-ATPase and glucose-6-phosphatase
activities were not enriched as compared with the homogenates
within each experimental group. The pattern of changes in the
enzymatic marker in the sarcolemmal membrane fraction was
essentially identical among the three experimental groups:
control, early sepsis, and late sepsis. These data indicate that
sarcolemmal membrane preparations were highly purified and
minimally contaminated with sarcoplasmic reticulum.
Further-more, the degrees of purity of sarcolemmal membranes and the
extents of cross contamination with sarcoplasmic reticulum
TABLE1. Marker enzyme activities of cardiac sarcolemmal membranes isolated from control and septic ratsControl Early sepsis Late sepsis
Homo SL Homo SL Homo SL
(Na++ K+)-ATPase 1.5 ± 0.2 29.0 ± 1.4 1.7 ± 0.4 28.1 ± 0.9 1.4 ± 0.2 24.0 ± 1.7
Ca2+-ATPase 2.4 ± 0.7 3.3 ± 0.4 2.1 ± 0.6 3.2 ± 0.3 2.7 ± 0.9 3.1 ± 0.2
Glucose-6-phosphatase 2.5 ± 0.6 2.4 ± 0.4 2.1 ± 0.3 3.1 ± 0.2 2.1 ± 0.3 3.3 ± 0.2 Values are Mean ± SE in µmol/mg/h. Each value represents a mean of six experiments. Homo, homogenate; SL, sarcolemmal membrane.
among the control, the early septic and the late septic groups
were the same.
Preliminary experiments were conducted to correlate
autora-diographic intensities of the immunoblots with various amounts
of GTP-binding protein (G
␣
s, G
␣
i-2, G
␣
i-3, and G

) in the
control rats before the full-scale Western blot analysis. The values
of signal intensity obtained were proportional to the amounts of
protein in the putative GTP-binding protein subspecies bands
(data not shown). Based on these observations, appropriate
concentrations of sarcolemmal membrane proteins within linear
range were then selected (100
g for G
␣
s, 100
g for G
␣
i-2, 100
g for G
␣
i-3, and 40
g for

subunit) for Western blot analysis
for the comparison between control and septic experiments.
Figure 1 shows the Western blot analysis of G
␣
s protein
levels in rat heart sarcolemmal membranes during different
phases of sepsis. Two putative bands for G
␣
s, with molecular
weights of 45 and 42 kDa, were detected. This finding is
consistent with that reported in the literature (24). Quantitative
analysis of the data reveals that the protein levels for 45-kDa
and 42-kDa forms of G
␣
s were unaltered during both early and
late phases of sepsis. These findings demonstrate that G
␣
s
protein levels remained unaffected in rat heart sarcolemmal
membranes during the progression of sepsis.
Figures 2 and 3 depict changes in G
␣
i-2 and G
␣
i-3 protein
levels in rat heart sarcolemmal membranes during the early and
the late phases of sepsis. Western blot analysis reveals that
G
␣
i-2 protein levels were not significantly affected (+17.3%; P
> 0.05) during the early phase but were increased by 46.5% (P
< 0.05) during the late phase of sepsis (Fig. 2). It should be
noted that the anti-G
␣
i-2 antibody used in this study cross
reacts with G
␣
i-l species. The fact that G
␣
i-l signal was not
detected in our study indicates that G
␣
i-l was absent from rat
heart sarcolemmal membranes (25). As shown in Figure 3,
FIG. 1. Western blot analysis of G␣s protein level in the sarcolemmal
membranes of control and septic rat hearts. Western blot analysis was
performed as described under the Materials and Methods section using poly-clonal antibody raised specifically against the G␣s subunits. Vertical bars indicate SEM. Number of experiments is shown in the parenthesis within each column. C, Control; ES, early sepsis; LS, late sepsis.
FIG. 2. Western blot analysis of G␣i-2 protein level in the
sarcolem-mal membranes of control and septic rat hearts. Western blot analysis
was carried out as described under the Materials and Methods section using polyclonal antibodies raised specifically against the G␣i-1 and G␣i-2 subunits of G-protein. The arrow shows the molecular weight of G␣i-2 subunit. Vertical bars indicate SEM. Number of experiments is shown in the parenthesis within each column. C, Control; ES, early sepsis; LS, late sepsis.
FIG. 3. Western blot analysis of G␣i-3 protein level in the
sarcolem-mal membranes of control and septic rat hearts. Western blot analysis
was performed as described under the Materials and Methods section using polyclonal antibody raised specifically against the G␣i-3 subunit of G-protein. The arrow indicates the molecular weight of G␣i-3 subunit. Vertical bars indicated SEM. Number of experiments is shown in the parenthesis of each column. C, control; ES, early sepsis, LS, late sepsis.
G
␣
i-3 protein levels remained relatively stable (+11.4%, P >
0.05) during early sepsis whereas they were increased by
61.3% (P < 0.01) during late sepsis (Fig. 3). The data presented
in Figures 2 and 3 demonstrate that G
␣
i-2 and G
␣
i-3 protein
levels were increased during the late stage of sepsis.
Figure 4 shows the Western blot analysis of G

protein
levels in rat heart sarcolemmal membranes during the progress
of sepsis. Quantitative analysis of Western blot indicates that
G

protein levels remained unaltered during the early and late
phases of sepsis. It should be mentioned that the G

antibody
used in this study cross reacts with both

1 and

2 subunits of
GTP-binding protein. In our experiments, only

2 subunit with
a molecular weight of 36 kDa was detected, indicating that

2
is the predominant subtype of G

expressed in the myocardium
(26). These findings indicate that G

protein level was
unaf-fected during the progression of sepsis.
Figure 5 depicts changes of adenylate cyclase activity in rat
heart sarcolemmal membranes during the two
cardiodynami-cally distinct phases of sepsis. Basal adenylate cyclase activity
remained unchanged during the early phase whereas it was
decreased by 25.7% (P < 0.05) during the late phase of sepsis
(Fig. 5A). Similarly, the isoproterenol-stimulated adenylate
cyclase activity was unchanged during early sepsis while it was
decreased by 44.6% (P < 0.01) during late sepsis (Fig. 5B).
These data indicate that the functional endpoint for
␣
-subunit
of GTP-binding protein, the adenylate cyclase activity, was
decreased during the late hypodynamic phase of sepsis.
DISCUSSION
GTP-binding proteins mediate sympathetic control of
cardiac function. The best characterized GTP-binding proteins
consist of two major classes: the stimulatory component, Gs,
which stimulates and the inhibitory component, Gi, which
inhibits adenylate cyclase activity (27). Adenylate cyclase is a
membrane-bond enzyme that catalyzes the hydrolysis of ATP
to cAMP. cAMP, an intracellular messenger, activates protein
kinase A and then initiates a series of enzymatic reactions
leading to a phosphorylation cascade of multiple proteins that
eventually regulate both the rate and the force of cardiac
contraction (27). In the heart, catecholamines released into the
synaptic cleft at sympathetic nerve terminals bind to either
␣
AR or

AR on the cardiac sarcolemma. Stimulation of

1AR,

2AR, and
␣
1AR activates G
␣
s and leads to increased cAMP
production through adenylate cyclase activation, whereas
stimulation of
␣
2AR activates G
␣
i and leads to decreased
cAMP production through adenylate cyclase inactivation (1,
27). An increase or a decrease in G
␣
s and/or G
␣
i levels
coupled with activation or inactivation of adenylate cyclase,
thus function as a modulating system regulating myocardial
contractility. The present study in cardiac sarcolemma reveals
that during late sepsis, the G
␣
i-2 and G
␣
i-3 protein levels were
increased and the increases were coupled with a decrease in
adenylate cyclase activity, may have a pathophysiological
significance in contributing to the understanding of the
patho-genesis of cardiac dysfunction during the late stage of sepsis.
The exact mechanisms responsible for the increase in G
␣
i-2
and G
␣
i-3 protein levels coupled with a decrease in adenylate
cyclase activity were not completely understood. It has been
reported that in the rat, a prolonged infusion of isoprenaline
decreased myocardial

AR number and increased heart G
␣
i
content (28, 29). In rat heart muscle cells, exposure to
noradrenaline decreased

AR density, increased G
␣
i-2 and
G
␣
i-3 protein levels, and reduced adenylate cyclase sensitivity
(30). The noradrenaline-induced increase in Gi protein
␣
-sub-units and adenylate cyclase desensitization were blocked by a

AR antagonist (30). In human end-stage heart failure,
myocardial

AR density was decreased and the decrease in

AR density was accompanied by an increase in G
␣
i protein
content (7, 12, 31). These reports demonstrate that a decreased

AR density in response to an enhanced sympathetic drive is
an important causative factor leading to the increased G
␣
i
protein level and the decreased adenylate cyclase sensitivity.
FIG. 4. Western blot analysis of G protein level in the sarcolemmalmembranes of control and septic rat hearts. Western blot analysis was
carried out as described under the Materials and Methods section using polyclonal antibody raised specifically against the subunit of G-protein. The arrow shows the molecular weight of G subunit. Vertical bars indicate SEM. Number of experiments is shown in the parenthesis within each column. C, Control; ES, early sepsis; LS, late sepsis.
FIG. 5. Changes in adenylate cyclase activities in the rat heart during
different phases of sepsis. Adenylate cyclase activities were assayed as
described under the Materials and Methods section except that for isoproterenol stimulated adenylate cyclase (B), 1 mM of isoproterenol was present in the reaction mixture. Vertical bars indicate SEM. Number of experiments is shown in the parenthesis within each column. C, Control; ES, early sepsis; LS, late sepsis.
The fact that myocardial

AR and
␣
AR densities have been
found to greatly reduced in response to the elevated circulating
catecholamines during the late phase of sepsis (16–18), it is
conceivable that the observed increase in the G
␣
i-2 and G
␣
i-3
protein levels and the decrease in the adenylate cyclase activity
are the results of underexpression of

AR and
␣
AR. In
addi-tion to the altered

AR and
␣
AR dynamics, changes in
aden-osine A1 receptor may contribute to the increase in G
␣
i protein
level and the decrease in adenylate cyclase activity because
adenosine A1 receptor is known to exist in cardiac myocytes
which coupled to G
␣
i and cyclase inhibition (32). Further
study is required to clarify role of adenosine A1 receptor on the
altered G
␣
i and cyclase inhibition in the heart during sepsis.
Because receptors and their coupling proteins belong to
differ-ent groups of genes that are controlled by distinct mechanisms of
regulation (33), it is possible that the altered expression of various
subspecies of GTP-binding protein is a result of modification of
their gene transcripts, instead of a secondary response to the
altered receptor dynamics. In heart failure, plasma catecholamine
levels were elevated and the elevated plasma catecholamine was
correlated with an increase in the myocardial G
␣
i-2 mRNA
abun-dance and an unchanged G
␣
s mRNA level (7, 12). It is of interest
to note that a consensus sequence of a “cAMP response element”
has been described in the promoter region of the G
␣
i-2 gene (34),
whereas it is lacking in the G
␣
s gene (35). These findings suggest
that an increased sympathetic drive via the cAMP cascade would
lead to an increased transcription rate of the G
␣
i-2, but not that of
the G
␣
s genes. Whether a similar transcriptional regulatory
mechanism is responsible for the increase in G
␣
i-2 and G
␣
i-3,
but not the G
␣
s, protein levels in the rat heart during the late
stage of sepsis, remains to be investigated.
It is of interest to note that lack of change in G

subunit of
GTP-binding protein in the rat heart during sepsis is strikingly
similar to that reported in the liver (22). Furthermore, G

as well
as G
␣
s are found to be quite stable under other pathological
conditions (7, 12, 22). Because chronic infusion of isoprenaline in
rats leads to increases in G
␣
i-2 and G
␣
i-3 mRNA levels whereas
G
␣
s and G

mRNA levels remain unchanged (7), these seem to
suggest that G

and G
␣
s subunits constitute the major
compo-nents of the GTP-binding protein associated signal
transduc-tion pathway that is irresponsive to the external stimuli.
REFERENCES
1. Offernanns S, Schultz G: Complex information processing by the transmem-brane signaling system involving G proteins. Naunyn-Schmiedeberg’s Arch Pharmacol 350:329–338, 1994.
2. Xiao RP, Cheng HP, Zhou YY, Kuschel K, Lakatta EG: Recent advances in cardiac2-adrenergic signal transduction. Circ Res 85:1092–1100, 1999. 3. Susan F: The molecular basis for distinct-adrenergic receptor subtype actions
in cardiomyocytes. Circ Res 85:1101–1111, 1999.
4. Insel PA: Adrenergic receptors, G-proteins, and cell regulation: implications for aging research. Exp Geronto1 28:341–348, 1993.
5. Exton JH: Cell signaling through quanine-nucleotide-binding regulatory proteins (G proteins) and phospholipases. EurJ Biochem 243:10–20, 1997. 6. Fields TA, Casey PJ: Signaling functions and biochemical properties of
pertus-sis toxin-repertus-sistant G-proteins. Biochem J 321:561–571, 1997.
7. Ischenhagen T, Mende U, Nose M, Scbmitz W, Scholz H, Haverich A, Hirt S, Doring V, Kalmar P, Hoppner W, Seitz HI: Increased messenger RNA level of the inhibitory G protein␣ subunit Gi␣-2 in human end-stage heart failure. Circ Res 70:688–696, 1992.
8. Castellano M, Bohm M: The cardiac-adrenoceptor-mediated signaling pathway and its alterations in hypertensive heart disease. Hypertension 29:715–722, 1997.
9. Kaura D, Takeda N, Sethi R, Wang X, Nagano M, Dhalla NS:-adrenoceptor mediated signal transduction in congestive heart failure in cardiomyopathic (UM-X 7.1) hamsters. Mol Cell Biochem 157:191–196, 1996.
10. Schmits W, Boknik P, Linck B, Muller FU: Adrenergic and muscarinic receptor regulation and therapeutic implications in heart failure. Mol Cell Biochem 157:251–258, 1996.
11. Yamamoto J, Ohyanagi O, Morita M, Iwasaki T: -adrenergic-G-protein-adenylate cyclase system complex in rat hearts with ischemic heart failure produced by coronary artery ligation. J Mol Cell Cardiol 26:617–626, 1994. 12. Bohm M: Alterations of-adrenoceptor-G-protein-regulated adenylyl cyclase
in heart. Mol Cell Biochem 147:147–160, 1995.
13. Ping P, Hammond HK: Diverse G protein and-adrenergic receptor mRNA expression in normal and failing porcine hearts. Am J PhysioI 267:H2079– H2085, 1994.
14. Bohm M, Kirchmayr R, Gierschik P, Erdmann E: Increase of myocardial inhibi-tory G-proteins in catecholamine-refracinhibi-tory septic shock or in septic multi-organ failure. Am J Med 98:183–186, 1995.
15. Reithmann C, Ha1lstrom S, Pilz G, Kapsner T, Schlag G, Wardan K: Desen-sitization of rat cardiomyocyte adenylyl cyclase stimulation by plasma of noradrenaline-treated patients with sepsis shock. Circ Shock 41:48–59, 1993. 16. Tang C, Liu MS: Initial externalization followed by internalization of
-adren-ergic receptors in rat heart during sepsis. Am J PhysioI 270:R254–R263, 1996. 17. Tang CS, Yang J, Wu LL, Dong LW, Liu MS: Phosphorylation of-adrenergic receptor leads to its redistribution in rat heart during sepsis. Am J Physiol 274:R1078–R1086, 1998.
18. Wu LL, Tang C, Liu MS: Hyper- and hypocardiodynamic states are associated with externalization and internalization, respectively, of␣-adrenergic receptors in rat heart during sepsis. Shock 7:318–323, 1997.
19. Wichterman KA, Baue AB, Chaudry IH: Sepsis and septic shock: a review of laboratory models and a proposal. J Sug Res 29:189–201, 1980.
20. Wu LL, Tang C, Dong LW, Liu MS: Altered phospholamban-calcium ATPase interaction in cardiac sarcoplasmic reticulum during the progression of sepsis. Shock 17:389–393, 2002.
21. Ausubel FM, Brents R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K: Immunoblotting and immunodetection. In Current Protocols in Molecular Biology. Vol 2. New York: John Wiley & Sons, Inc., 1994, pp 10.8.1–10.8.14. 22. Wu LL, Dong LW, Liu MS: Alterations of G-protein and adenylate cyclase signaling in rat liver during the progression of sepsis. Shock 11:39–43, 1999. 23. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ: Protein measurement with
the Folin phenol reagent. J Biol Chem 193:265–275, 1951.
24. Bushfield M, Griffiths SL, Murphy GJ, Pyne NJ, Knowler JT, Milligan G, Parker PJ, Mollner S, Houslay MD: Diabetes-induced alterations in the expres-sion, function and phosphorylation state of the inhibitory guanine nucleotide regulatory protein Gi-2 in hepatocytes. Biochem J 271:365–372, 1990. 25. Garibay JLR, Kozasa T, Itoh H, Tsukamoto T, Matsuoka M, Kaziro Y: Analysis
of mRNA levels of the expression of six G protein␣-subunit genes in mamma-lian cells and tissues. Biochim Biophys Acta 1094:193–199, 1991.
26. Hinsch KD, Tychowiecka I, Gausepohl H, Frank R, Rosenthal W, Schultz G: Tissue distribution of1- and2-subunits of regulatory guanine nucleotide-binding proteins. Biochim Biophys Acta 1013:60–67, 1989.
27. Ishikawa Y, Homcy CJ: The adenylyl cyclases as integrators of transmembrane signal transduction. Circ Res 80:297–304, 1997.
28. Mende U, Eschenhagen T, Geertz B, Schmitz W, Scholz H, Schulte AM, Esch J, Sempell R, Seinfath M: Isoprenaline-induced increase in 40/41 kDa pertussis toxin substrates and functional consequences on contractile response in rat heart. Nauyn-Schmiedeberg’s Arch Pharmacol 345:44–50, 1992.
29. Eschenhagen T, Mende U, Schmitz W, Scholz H, Schulte AM, Esch J, Sempell R, Warnholtz A, Wustel JM: Beta-adrenoceptor stimulation-induced increase in cardiac Gi protein expression and in carbachol sensitivity. Eur Heart J 12:127–131, 1991. 30. Reithmann C, Gierschik P, Muller U, Warden K, Jakobs KH: Pseudomonas
exotoxin A prevents-adrenoceptor-induced up-regulation of Gi protein ␣-sub-units and adenylyl cyclase desensitization in rat heart muscle cells. Mol Phar-macol 37:631–638, 1990.
31. Bunemann M, Lee KB, Pals-Rylarsdam R, Roseberry AF, Hosey MM: Desen-sitization of G-protein-coupled receptors in the cardiovascular system. Annul Rev Physiol 61:169–192, 1999.
32. Feldman AM: Experimental issues in assessment of G protein function in cardiac disease. Circulation 84:1852–1861, 1991.
33. Mondry A, Bourgeois F, Carre F, Swynghedauw B, Moalic JM: Decrease in 1-adrenergic and M2-muscarinic receptor mRNA levels and unchanged accu-mulation of mRNAs coding for G␣i-2 and G␣s proteins in rat cardiac hyper-trophy. J Mol Cell Cardiol 27:2287–2294, 1995.
34. Weinstein LS, Spiegel AM, Carter AD: Cloning and characterization of the human gene of the␣-subunit of Gi2, a GTP-binding signal transducing protein. FEES Lett 232:333–340, 1988.
35. Kozasa T, Itoh H, Tsukamoto T, Kaziro Y: Isolation and characterization of the human Gs␣ gene. Proc Natl Acad Sci USA 85:2081–2085, 1988.