行政院國家科學委員會專題研究計畫 期中進度報告
猪脂肪細胞結合素及其受體功能表現調控機制之探討(2/3)
期中進度報告(精簡版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2313-B-002-024- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學動物科學技術學系暨研究所 計 畫 主 持 人 : 丁詩同 報 告 附 件 : 國外研究心得報告 出席國際會議研究心得報告及發表論文 處 理 方 式 : 期中報告不提供公開查詢中 華 民 國 96 年 06 月 22 日
Running Head: PORCINE ADIPONECTIN AND ADIPONECTIN RECEPTORS
1
Key Words: Adiponectin, Adiponectin receptor, Insulin, Pigs.
2
Insulin regulates the expression of adiponectin and adiponectin receptors
3in mature porcine adipocytes
45
Bing Hsien Liu1, Ya Chin Wang1, Shinn Chih Wu1,2, Harry John Mersmann1, Winston
6
Teng Kuei Cheng1,2 and Shih Torng Ding1,2
7
1Department of Animal Science and Technology, National Taiwan University, Taipei
8
106, Taiwan 9
2Institute of Biotechnology, National Taiwan University, Taipei 106, Taiwan
10
Corresponding Author : 11
Shih-Torng Ding, Department of Animal Science and Technology / Institute of 12
Biotechnology, National Taiwan University 13
50, Lane 155, Kee-Long Rd. Sec. 3, Taipei 106, Taiwan 14 Phone: +886953610078 15 Fax: +886227324070 16 Email = [email protected] 17 18
Abstract
19
Adiponectin is an adipocyte-derived hormone that can improve 20
insulin-sensitivity. Its functions in regulating glucose utilization and fatty acid 21
metabolism in mammals are mediated by two subtypes of adiponectin receptors 22
(AdipoR1 and AdipoR2). This study was conducted to determine the effect of 23
insulin on the expression of adiponectin and its receptors. The expression of both 24
AdipoR mRNAs was increased in the liver and s. c. adipose tissue of fasted pigs 25
compared with fed pigs. The data also showed that the expression of either AdipoR1 26
and AdipoR2 mRNA in muscle and visceral adipose tissue was not different between 27
the fasted and fed conditions. We demonstrated that in the presence of 10 nM insulin, 28
addition of 1 uM of insulin or rosiglitazone [a peroxisome proliferator - activated 29
receptor γ (PPARγ) agonist] had no effect on the expression of adiponectin and 30
AdipoR genes in differentiated porcine adipocytes. However, the addition of 1 μM 31
insulin plus 1 μM rosiglitazone significantly increased the AdipoR2 mRNA in 32
well-differentiated porcine adipocytes. Using the phosphatidylinositol 3 - kinase 33
inhibitor (PI3K inhibitor, LY294002), we found that insulin inhibited the expression 34
of AdipoR2 through the PI3K pathway and this inhibition can be blocked by addition 35
of rosiglitazone. When porcine adipocytes were cultured without insulin, 36
supplementation with 10 nM insulin inhibited the expression of AdipoR2 and this 37
inhibition effect can also be blocked by addition of rosiglitazone. Therefore, these 38
data suggest that a PPARγ agonist increases expression of AdipoR2 and that insulin 39
inhibits the expression of AdipoR2 through the PI3K pathway. 40
Introduction
41
Adiponectin is an adipocyte-produced protein hormone circulating in the blood 42
(1-4). Administration of adiponectin to mice decreases plasma glucose, free fatty 43
acids, and triglycerides, but increases muscle fatty acid oxidation and induces weight 44
loss (5). The function of adiponectin is carried out through the activation of 45
AMP-activated protein kinase (4). Adiponectin-deficient mice are mildly insulin 46
resistant and glucose intolerant when fed a standard diet (6, 7). Furthermore, 47
decreased circulating adiponectin concentrations are associated with insulin resistance, 48
obesity, and type diabetes Ⅱ (8, 9). Therefore, adiponectin may modify the function 49
of insulin in mammals. 50
Thiazolidinediones (a class of type diabetes drugs)Ⅱ , ligands for peroxisome 51
proliferator-activated receptor γ (PPARγ) increase adiponectin expression and plasma 52
adiponectin concentration in rodents (8, 10). Thiazolidinediones also increase the 53
expression of adiponectin in type Ⅱ diabetes mellitus and obese patients (11, 12). 54
Dual activation of PPARα and γ increases serum adiponectin concentration in adipose 55
tissue of obese diabetic KKAy mice (13). Furthermore, there is a functional 56
PPAR-response element (PPRE) in the promoter region of the adiponectin gene (14). 57
Thus, adiponectin should be regulated by PPARγ and its ligands. 58
Yamauchi et al. (15) first cloned the cDNA encoding adiponectin receptors 1 59
(AdipoR1) and 2 of human and mouse. These two adiponectin receptors contain 7 60
transmembrane domains, but they are structurally and functionally distinct from 61
G-protein-coupled receptors (15). Both AdipoR1 and AdipoR2 can mediate the 62
function of adiponectin and the expression of these two receptors is regulated by 63
PPARγ ligands or a combination of ligands for PPARα and PPARγ in obese patients 64
and mice(11-13). In order to clarify the regulation of gene expression of adiponectin 65
receptors in pigs, we have cloned the full length cDNA from porcine AdipoR genes 66
(16); and in this study, we determined the insulin regulation of AdipoR1 and AdipoR2 67
expression in adipocytes. We also investigated the interaction of a PPARγ ligand 68
and insulin on the expression of adiponectin and AdipoR genes in porcine adipocytes. 69
70
Materials and Methods
71
Fasting and feeding animals. The animal protocol was approved by the 72
Experimental Animal Care and Use Committee at National Taiwan University. Four 73
male and four female crossbred pigs (Sus domesticus; sows were predominantly 74
Landrace-Yorkshire crossbreds mated to a Duroc boar) were weaned at 28 d of age 75
and fed a commercial diet (a corn, soy-based diet containing 18% crude protein and 76
4% fat) ad libitum and raised to 60 d of age for the experiment. In order to 77
determine the effect of nutritional conditions on the expression of adiponectin and its 78
receptors, fed pigs were euthanized by electrical stunning coupled with 79
exsanguination at 10:00 h after feeding at 8:00 h, whereas the fasted group was killed 80
after a 24 h fast. Liver, longissimus muscle, visceral adipose tissue of the belly and 81
dorsal subcutaneous (s.c.) adipose tissue were dissected and frozen in liquid N2 and
82
stored at -70℃ for RNA extraction. The average body weight of pigs for both 83
treatments was 20.4 + 0.66 kg when euthanized. Total RNA was extracted for 84
detecting AdipoR1, AdipoR2, and β-actin gene expression by Northern analysis 85
Isolation of porcine stromal vascular (S/V) cells. Porcine adipose tissue 86
samples were digested and S/V cells were isolated and cultured as previously 87
described (17, 18). In brief, adipose tissue from 9 d old crossbred pigs was removed 88
from the dorsal s.c. depot in the neck, shoulder, and back regions. The slices of 89
adipose tissue were digested with collagenase (Sigma C6885; Sigma, St Louis, MO, 90
USA) in sterile Krebs Ringer bicarbonate buffer at 37 for 90 min℃ . The S/V cell 91
fraction was isolated by centrifugation at 800 x g for 10 min and the pellet was 92
washed three times by resuspension with DMEM/F12 medium (Sigma D8900) 93
supplemented with NaHCO3, 100 U penicillin/mL, 100 mg streptomycin/mL, 1.5
94
μg/mL amphotericin B and 10% fetal bovine serum. Before the last washing step, 95
the S/V cell fraction was treated with red blood cell lysing buffer (155 mM NH4Cl,
96
5.7 mM K2HPO4, 0.1 mM EDTA at pH 7.3) to remove red blood cells which may
97
reduce adhesion of S/V cells. The washed S/V cells were resuspended in 98
DMEM/F12 containing 10% fetal bovine serum and plated at a concentration of 5 x 99
104 cells/cm2. The S/V cells were then cultured at 37 in air containing 5℃ % CO2 for
100
48 h to let the cells fully attach to the dish. 101
Cell culture and differentiation of porcine adipocytes. After 48 h of initial 102
incubation for proliferation (defined as day 0), the medium was removed and replaced 103
by serum-free, hormone-supplemented differentiation medium (DMEM/F12 104
containing NaHCO3, 25 mM glucose, 1 μM bovine insulin, 10 µg transferrin/mL, 2
105
mM L-glutamine, 33 μM biotin, 17 μM pantothenate, 100 nM dexamethasone, 1 nM 106
triiodothyronine, 100 U penicillin/mL, 100 mg streptomycin/mL, 1.5 μg/mL 107
amphotericin B and 1 μM rosiglitazone) for 3 d to induce adipogenesis. The 108
medium was then changed to differentiation medium without rosiglitazone. The 109
medium was replaced every 3 d. After 9 d, up to 90-95% of the attached cells were 110
differentiated to cells with visible lipid droplets. For studying the expression of 111
genes during porcine adipocyte differentiation, porcine S/V cells were induced to 112
differentiation and on the indicated days (0, 3, 6, 9), total RNA was extracted for 113
detecting adiponectin, AdipoR1, AdipoR2, and β-actin mRNA by Northern analysis. 114
The results are the means of 3 independent experiments with S/V cells isolated from 3 115
different pigs. 116
Effect of rosiglitazone and insulin on AdipoRs in porcine adipocytes. To study 117
the effect of insulin on the expression of adiponectin and AdipoRs, differentiated 118
adipocytes were washed with phosphate-buffered saline and then cultured in low 119
glucose DMEM/F12 (DMEM, Sigma D5523 : nutrient mixture F12, Sigma N6760 = 120
1 : 1, with a final glucose concentration of 7.78 mM) with 10 nM insulin for 6 hrs, and 121
then 1 μM insulin or 1 μM rosiglitazone or insulin + rosiglitazone were added for 2 , 122
12, or 24 hrs. Total RNA was extracted to detect adiponectin, AdipoR1, AdipoR2, 123
and β-actin mRNA by Northern analysis. The results are the means of 4 independent 124
experiments using cells isolated from 4 different pigs. 125
The involvement insulin signal pathways on regulation expression of AdipoRs 126
in porcine adipocytes. Porcine S/V cells were differentiated for 9 days and then 127
differentiated adipocytes were cultured in low glucose (7.78 mM) DMEM/F12 with 128
10 nM insulin for 6 h. After 6 h, 1 μM insulin[concentration was reported to be 129
effective, (19)] + 1 μM rosiglitazone (20) were added to some cells and incubation 130
was continued for 24 h. Also at 6 h, the mitogen-activated protein kinase (MAPK) 131
inhibitor, PD98059 at 25 μM (21) or the phosphatidylinositol 3 - kinase (PI3K) 132
inhibitor, LY294002 at 10 μM (19) were added along with 1 μM insulin + 1 μM 133
rosiglitazone; incubation was continued for 24 h. Total RNA was extracted for 134
determining AdipoR1, AdipoR2, and β-actin gene expression by Northern analysis. 135
The results are the means of 4 independent experiments using cells isolated from 4 136
different pigs. 137
Insulin, rosiglitazone, and PI3K inhibitor treatments in porcine adipocytes. 138
Porcine S/V cells were differentiated for 9 days and then differentiated adipocytes 139
were cultured in low glucose (7.78 mM) DMEM/F12 without 10 nM insulin for 6 hrs, 140
then 10 nM insulin, 1 μM insulin, 1 μM rosiglitazone, or 10 μM LY294002 were 141
added to study the effect of low insulin, high insulin concentrations and insulin 142
sensitizer on the expression of AdopoRs. Total RNA was extracted for determining 143
adiponectin, AdipoR2, and β-actin gene expression by Northern analysis. The 144
results are the means of 4 independent experiments using cells isolated from 4 145
different pigs. 146
Northern analysis. Total RNA was extracted by the guanidinium - phenol - 147
chloroform extraction method (22). The integrity of RNA was determined by 148
examination of the 18S and 28S ribosomal RNA bands after electrophoresis. The 149
RNA was quantified by spectrophotometry at 260 nm and stored at -70 .℃ Total 150
RNA (10 µg of each sample) was electrophoresed and transferred to nylon membranes 151
for Northern analysis following the procedure described by Liu et al (23). The 152
porcine adiponectin, AdipoR1, AdipoR2 and β-actin probe sequences were previously 153
described (16, 18) and labeled by P32dC with PCR amplification. Hybridization 154
blotting images were quantified using a Typhon 9200 phosphorimage scanner and 155
ImageQuant TL v2005 software (GE). The densitometric value for an individual 156
transcript in a sample lane was normalized to the densitometric value for the β-actin 157
mRNA in the same lane. 158
Statistical analysis. Data were presented as mean ± S.E.M. Statistical 159
analysis were using an ANOVA procedure to determine the major effects of insulin, 160
rosiglitazone, and insulin signaling pathway blockers. Duncan’s new multiple-range 161
test was used to evaluate differences among means (SAS Inst., Inc., Cary, NC). A 162
significant difference was indicated at P ≤ 0.05. 163
164
Results and Discussion
165
Fasting increased AdipoR genes expression in s.c. adipose tissue and liver. In 166
porcine s.c. adipose tissue and liver, the AdipoR1 and AdipoR2 mRNAs increased 167
(P<0.05) after a 24 h fast, but there was no effect of fasting on the expression of 168
either receptor in the visceral adipose tissue and muscle (Fig. 1A and 1B). The data 169
suggest differential regulation of AdipoR1 and AdipoR2 by nutritional status in 170
various tissues. Previously, we indicated that the expression of AdipoR2, but 171
AdipoR1 and adiponectin was increased in the s.c. adipose tissue after an 8 h fast 172
(16). In rodents, similar to the current study in pigs, both AdipoR1 and AdipoR2 173
gene expression are up-regulated after 48 h fasting in the liver (24). The reason 174
that fasting increased the expression of AdipoR genes in mouse muscle (24), but not 175
in pig muscle is not known. We speculate that the 48 h fast in the mouse is more 176
extreme than the 24 h fast in the pig; this difference may also contribute to the 177
observed species difference. Others have showed that serum adiponectin remained 178
stable concentrations during 72 h of fasting in normal- and over-weight humans (25). 179
Thus, hormonal regulation of the feeding / fasting status on adiponectin function 180
may mainly act through regulating the expression of AdipoRs. In summary, hepatic 181
and s.c. adipose tissue AdipoR genes, but not visceral adipose tissue and muscle 182
AdipoR genes, were responsive to the feeding / fasting status in pigs. 183
Expression of adiponectin and AdipoR genes during porcine adipocyte 184
differentiation. Expression of both adiponectin and AdipoR2 increased during 185
adipocyte differentiation, whereas expression of AdipoR1 increased during the initial 186
3 d with no further increase during the latter stages of differentiation (Fig. 2). The 187
data suggest that either AdipoR2 is more sensitive to the hormones present in the 188
adipocyte cell culture medium than AdipoR1 or the AdipoR2 is a more important 189
receptor mediating adiponectin function in the differentiated adipocyte. A recent 190
study indicates that adiponectin increases PPARγ2 expression in porcine adipocytes 191
(26), suggesting that adiponectin may involve in regulating the PPARγ2 function 192
during adipogenesis. Furthermore, overexpression of adiponectin not only 193
promotes adipogenesis by prolonging and enhancing the key adipogenic 194
transcription factors, ie., PPARγ, CCAAT/enhancer binding protein α (C/EBPα), and 195
adipocyte determination and differentiation factor 1/sterol-regulatory element 196
binding protein 1c (ADD1/SREBP1c), but also stimulates glucose uptake through 197
increasing glucose transporter 4 gene expression in 3T3-L1 adipocytes (27). 198
Therefore, a greater expression of the adiponectin gene in the latter stages of porcine 199
adipocyte differentiation may function to enhance or maintain adipocyte morphology 200
through the regulation of PPARγ and other adiponectin target genes. 201
Fu et al. (27) found that the expression of both AdipoR genes were 202
down-regulated in adiponectin-overexpressing adipocytes, suggesting that 203
adiponectinmay act in an autocrine or paracrine fashion to regulate the function of 204
its receptors in adipose tissue. A similar study also found that adiponectin 205
downregulated its own production and the expression of its AdipoR2 receptor in 206
transgenic mice (28). The receptor down-regulation may decrease the adiponectin 207
responses to slow the adipogenic progression; however, perhaps receptor 208
down-regulation only occurs when adiponectin concentration reaches extreme levels, 209
as in the overexpressing cells and mice. 210
Effects of insulin and rosiglitazone on expression of adiponectin and AdipoRs. 211
Treatment with a high concentration of insulin for 2 to 24 hr did not change the 212
expression of adiponectin or the AdipoRs (Fig. 3). In contrast, several studies 213
indicate that insulin inhibits the expression of adiponectin and AdipoR genes in 214
adipocytes of humans and rodents (13, 19, 29). We cannot exclude the possibility 215
that the 10 nM insulin concentration used in the pre-incubation medium was great 216
enough to suppress the expression of adiponectin and AdipoR genes so that the 217
higher insulin concentration (1 μM) used to test for insulin effects had no additional 218
effect. 219
The PPARγ agonist, rosiglitazone alone did not increase the expression of 220
adiponectin or the AdipoRs (Fig. 3). However, the combination of a high insulin 221
concentration (1 μM) plus rosiglitazone (1 μM) increased expression of adiponectin 222
and AdipoR2 in porcine adipocytes. In 3T3-L1 adipocytes cultured with DMEM + 223
10% FBS, rosiglitazone does not affect the expression of adiponectin, even though 224
there is a PPAR response element (PPRE) in the adiponectin promoter region (30). 225
Rosiglitazone does increase plasma adiponectin concentration in humans and mice in 226
vivo (30, 31). The authors suggest that an adequate insulin concentration may be 227
necessary to observe the rosiglitazone-stimulated increase in the expression of the 228
adiponectin gene (30). Recent studies also showed that AdipoR2, but not AdipoR1, 229
was increased by rosiglitazone treatment in mouse primary adipocytes, and HepG2 230
hepatocytes cultured with FBS (13, 20). 231
In order to clarify the relationship between insulin and the PPARγ agonist, we 232
used insulin signal pathway inhibitors to block phosphatidylinositol 3-kinase (PI3K) 233
or mitogen-activated protein kinase (MAPK). Expression of AdipoR2, but not 234
AdipoR1 mRNA was increased in the presence of 1 μM insulin + 1 μM rosiglitazone 235
(Fig. 4), confirming data in Fig. 3B, C. Addition of the MAPK inhibitor, PD98059 236
had no effect on expression of AdipoR1 or AdipR2 mRNA. However, the AdipoR2 237
mRNA was increased when the PI3K signal pathway was inhibited by addition of 238
LY294002, suggesting that insulin may decrease the expression of AdipoR2 through 239
the PI3K pathway. Tsuchida et al. (24) recently demonstrated that insulin 240
suppressed the expression of AdipoRs via the PI3K/Foxo1-dependent pathway in 241
rodent hepatocytes and myocytes. Although we did not observe that insulin alone 242
decreased AdipoR2 mRNA expression, the increase in AdipoR2 mRNA when the 243
PI3K pathway was inhibited by addition of LY294002, suggests that insulin has at 244
least a partial suppressive effect via the PI3K pathway. In addition, studies also 245
showed evidence of a direct insulin-induced inhibition of AMPK activity through 246
PI3K/Akt pathway in the heart (32), which is a possible mechanism by which the 247
expression of AdipoRs are inhibited. 248
In order to exclude the possibility that 10 nM insulin, used in most experiments 249
to maintain cell viability, suppressed adiponectin or AdipoRs gene expression, we 250
designed another experiment to clarify the interaction of insulin and a PPARγ agonist 251
on adiponectin and AdipoRs gene expression. After treating with insulin free 252
medium for 6 h, addition of 10 nM insulin reduced both adiponectin and AdipoR2 253
(Fig. 5A) mRNAs, indicating a minute amount of insulin suppresses the expression 254
of adiponectin and AdipoR2 genes. In pigs, the normal concentration of insulin is 255
maintained at 106 ± 12.5 pM and increased to 850 ± 137.5 pM 30 mins post-prandial 256
(33). Hence, supplementation with 10 nM or 1 μM insulin in porcine adipocyte 257
culture is much higher than concentrations in vivo. In mammals, the concentration 258
of feeding / fasting insulin could not affect the abundance of adiponectin (16, 25). 259
Insulin may regulate the ratio of high molecular weight adiponectin (active form) or 260
the expression of AdipoRs to modify the adiponectin function. This high insulin 261
concentration is similar to the conditions in type II diabetes or other 262
hyperinsulinemia-induced metabolism syndromes. 263
Adiponectin overexpressed mice have been shown to be able to improve insulin 264
resistance (34), whereas adiponectin deficient mice were shown to be mildly insulin 265
resistant and glucose intolerant when fed a standard diet, and severe insulin resistant 266
triggered by high fat diet (6, 7). Mice lacking adiponectin show decreased hepatic 267
insulin sensitivity and reduced responsiveness to PPARγ agonists (35). In addition, 268
several SNPs on adiponectin and AdipoR genes and other environmental factors such 269
as obesity appears to be hypoadiponectinemic and associated with type II diabetes 270
(36). Hence, regulation of adiponectin function may have therapeutic potential for 271
treating type II diabetes mellitus and obesity. 272
Because insulin decreased the expression of adiponectin and AdipoR2, we 273
cultured well-differentiated adipocytes in medium without insulin for 6 h. Addition 274
of the PPARγ agonist, rosiglitazone to these cells for 24 h significantly increased 275
adiponectin mRNA expression, but not AdipoR2 mRNA (Fig. 5B). It should be 276
noted that in the presence of 10 nM insulin, rosiglitazone had no effect on the 277
expression of AdipoR2 (Fig. 3C). This result confirmed data from our first 278
experiment and was similar to that reported in differentiated human myotubes (37), 279
indicating that the PPARγ agonist may improve insulin sensitivity by increasing the 280
serum adiponectin level, but had no effect on its receptors. Recently research also 281
showed that administrating TZD for type 2 diabetes mellitus to improve insulin 282
sensitivity and increase adiponectin levels did not affect the expression of AdipoR1 283
and AdipoR2 in muscle and adipose tissue (38). Although rosiglitazone may 284
require the presence of insulin to increase the expression of adiponectin and 285
AdipoR2 genes (Fig. 3), the effect of rosiglitazone might be in reversing the 286
inhibitory effect of insulin. It was interesting that LY294002 promoted the 287
expression of both adiponectin and AdipoR2 mRNAs more strongly than insulin 288
alone, rosiglitazone alone or rosiglitazone plus insulin (Fig. 5). Hence, these 289
findings suggest that insulin suppresses the expression of adiponectin and AdipoR2 290
mRNAs through PI3K pathway. When PI3K pathway blocked, both rosiglitazone 291
and insulin could increase the expression of adiponectin and AdipoR2 mRNAs 292
through a yet to be determined mechanism. 293
In conclusion, the expression of the adiponectin and AdipoR2 mRNAs is more 294
strongly inhibited by insulin and stimulated by the PPARγ agonist, rosiglitazone than 295
the AdipoR1 mRNA. The PPARγ agonist effects depend on the presence of insulin, 296
perhaps act in a permissive manner. The insulin effect mimicking the type II 297
diabetes was mediated through the PI3K pathway as indicated by use of the PI3K 298
inhibitor, LY294002. Therefore, understanding of interaction between insulin, 299
PPARγ and expression of adiponectin and its receptors will provide mechanisms that 300
may lead to control of adipose fat deposition and to the treatment of type II diabetes. 301
1. Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF. A novel serum
302
protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 303
1995;270:26746-26749. 304
2. Hu E, Liang P, Spiegelman BM. AdipoQ is a novel adipose-specific gene
305
dysregulated in obesity. J Biol Chem 1996;271:10697-10703. 306
3. Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y,
307
Matsubara K. cDNA cloning and expression of a novel adipose specific
308
collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). 309
Biochem Biophys Res Commun 1996;221:286-289. 310
4. Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors. Endocr
311
Rev 2005;26:439-451. 312
5. Fruebis J, Tsao TS, Javorschi S, Ebbets-Reed D, Erickson MR, Yen FT,
313
Bihain BE, Lodish HF. Proteolytic cleavage product of 30-kDa adipocyte
314
complement-related protein increases fatty acid oxidation in muscle and 315
causes weight loss in mice. Proc Natl Acad Sci U S A 2001;98:2005-2010. 316
6. Kubota N, Terauchi Y, Yamauchi T, Kubota T, Moroi M, Matsui J, Eto K,
317
Yamashita T, Kamon J, Satoh H, Yano W, Froguel P, Nagai R, Kimura S,
318
Kadowaki T, Noda T. Disruption of adiponectin causes insulin resistance and
319
neointimal formation. J Biol Chem 2002;277:25863-25866. 320
7. Maeda N, Shimomura I, Kishida K, Nishizawa H, Matsuda M, Nagaretani
321
H, Furuyama N, Kondo H, Takahashi M, Arita Y, Komuro R, Ouchi N,
322
Kihara S, Tochino Y, Okutomi K, Horie M, Takeda S, Aoyama T,
323
Funahashi T, Matsuzawa Y. Diet-induced insulin resistance in mice lacking
324
adiponectin/ACRP30. Nat Med 2002;8:731-737. 325
8. Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K, Mori Y,
326
Ide T, Murakami K, Tsuboyama-Kasaoka N, Ezaki O, Akanuma Y,
327
Gavrilova O, Vinson C, Reitman ML, Kagechika H, Shudo K, Yoda M,
328
Nakano Y, Tobe K, Nagai R, Kimura S, Tomita M, Froguel P, Kadowaki T.
329
The fat-derived hormone adiponectin reverses insulin resistance associated 330
with both lipoatrophy and obesity. Nat Med 2001;7:941-946. 331
9. Spranger J, Kroke A, Mohlig M, Bergmann MM, Ristow M, Boeing H,
332
Pfeiffer AF. Adiponectin and protection against type 2 diabetes mellitus.
333
Lancet 2003;361:226-228. 334
10. Maeda N, Takahashi M, Funahashi T, Kihara S, Nishizawa H, Kishida K,
335
Nagaretani H, Matsuda M, Komuro R, Ouchi N, Kuriyama H, Hotta K,
336
Nakamura T, Shimomura I, Matsuzawa Y. PPARgamma ligands increase
337
expression and plasma concentrations of adiponectin, an adipose-derived 338
protein. Diabetes 2001;50:2094-2099. 339
11. Yu JG, Javorschi S, Hevener AL, Kruszynska YT, Norman RA, Sinha M,
340
Olefsky JM. The effect of thiazolidinediones on plasma adiponectin levels in
341
normal, obese, and type 2 diabetic subjects. Diabetes 2002;51:2968-2974. 342
12. Chinetti G, Zawadski C, Fruchart JC, Staels B. Expression of adiponectin
343
receptors in human macrophages and regulation by agonists of the nuclear 344
receptors PPARalpha, PPARgamma, and LXR. Biochem Biophys Res 345
Commun 2004;314:151-158. 346
13. Tsuchida A, Yamauchi T, Takekawa S, Hada Y, Ito Y, Maki T, Kadowaki T.
347
Peroxisome proliferator-activated receptor (PPAR)alpha activation increases 348
adiponectin receptors and reduces obesity-related inflammation in adipose 349
tissue: comparison of activation of PPARalpha, PPARgamma, and their 350
combination. Diabetes 2005;54:3358-3370. 351
14. Seo JB, Moon HM, Noh MJ, Lee YS, Jeong HW, Yoo EJ, Kim WS, Park J,
352
Youn BS, Kim JW, Park SD, Kim JB. Adipocyte determination- and
353
differentiation-dependent factor 1/sterol regulatory element-binding protein 1c 354
regulates mouse adiponectin expression. J Biol Chem 2004;279:22108-22117. 355
15. Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, Sugiyama T,
356
Miyagishi M, Hara K, Tsunoda M, Murakami K, Ohteki T, Uchida S,
357
Takekawa S, Waki H, Tsuno NH, Shibata Y, Terauchi Y, Froguel P, Tobe
358
K, Koyasu S, Taira K, Kitamura T, Shimizu T, Nagai R, Kadowaki T.
359
Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. 360
Nature 2003;423:762-769. 361
16. Ding ST, Liu BH, Ko YH. Cloning and expression of porcine adiponectin and
362
adiponectin receptor 1 and 2 genes in pigs. J Anim Sci 2004;82:3162-3174. 363
17. Suryawan A, Hu CY. Effect of serum on differentiation of porcine adipose
364
stromal-vascular cells in primary culture. Comp Biochem Physiol Comp 365
Physiol 1993;105:485-492. 366
18. Liu BH, Kuo CF, Wang YC, Ding ST. Effect of docosahexaenoic acid and
367
arachidonic acid on the expression of adipocyte determination and 368
differentiation-dependent factor 1 in differentiating porcine adipocytes. J Anim 369
Sci 2005;83:1516-1525. 370
19. Fasshauer M, Klein J, Neumann S, Eszlinger M, Paschke R. Hormonal
371
regulation of adiponectin gene expression in 3T3-L1 adipocytes. Biochem 372
Biophys Res Commun 2002;290:1084-1089. 373
20. Sun X, Han R, Wang Z, Chen Y. Regulation of adiponectin receptors in
374
hepatocytes by the peroxisome proliferator-activated receptor-gamma agonist 375
rosiglitazone. Diabetologia 2006;49:1303-1310. 376
21. Pereira RI, Draznin B. Inhibition of the phosphatidylinositol 3'-kinase
signaling pathway leads to decreased insulin-stimulated adiponectin secretion 378
from 3T3-L1 adipocytes. Metabolism: clinical and experimental 379
2005;54:1636-1643. 380
22. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid
381
guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 382
1987;162:156-159. 383
23. Liu BH, Wang YC, Cheng WM, Shen TF, Ding ST. The effects of
384
docosahexaenoic acid oil and soybean oil on the expression of lipid 385
metabolism related mRNA in pigs. Asian-Australasian Journal of Animal 386
Science 2005;18:1451-1456. 387
24. Tsuchida A, Yamauchi T, Ito Y, Hada Y, Maki T, Takekawa S, Kamon J,
388
Kobayashi M, Suzuki R, Hara K, Kubota N, Terauchi Y, Froguel P, Nakae
389
J, Kasuga M, Accili D, Tobe K, Ueki K, Nagai R, Kadowaki T.
390
Insulin/Foxo1 pathway regulates expression levels of adiponectin receptors 391
and adiponectin sensitivity. J Biol Chem 2004;279:30817-30822. 392
25. Merl V, Peters A, Oltmanns KM, Kern W, Born J, Fehm HL, Schultes B.
393
Serum adiponectin concentrations during a 72-hour fast in over- and 394
normal-weight humans. International journal of obesity 2005;29:998-1001. 395
26. Ajuwon KM, Spurlock ME. Adiponectin inhibits LPS-induced NF-kappaB
396
activation and IL-6 production and increases PPARgamma2 expression in 397
adipocytes. Am J Physiol Regul Integr Comp Physiol 2005;288:R1220-1225. 398
27. Fu Y, Luo N, Klein RL, Garvey WT. Adiponectin promotes adipocyte
399
differentiation, insulin sensitivity, and lipid accumulation. J Lipid Res 400
2005;46:1369-1379. 401
28. Bauche IB, Ait El Mkadem S, Rezsohazy R, Funahashi T, Maeda N,
402
Miranda LM, Brichard SM. Adiponectin downregulates its own production
403
and the expression of its AdipoR2 receptor in transgenic mice. Biochem 404
Biophys Res Commun 2006;345:1414-1424. 405
29. Korner A, Wabitsch M, Seidel B, Fischer-Posovszky P, Berthold A,
406
Stumvoll M, Bluher M, Kratzsch J, Kiess W. Adiponectin expression in
407
humans is dependent on differentiation of adipocytes and down-regulated by 408
humoral serum components of high molecular weight. Biochem Biophys Res 409
Commun 2005;337:540-550. 410
30. Combs TP, Wagner JA, Berger J, Doebber T, Wang WJ, Zhang BB, Tanen
411
M, Berg AH, O'Rahilly S, Savage DB, Chatterjee K, Weiss S, Larson PJ,
412
Gottesdiener KM, Gertz BJ, Charron MJ, Scherer PE, Moller DE.
413
Induction of adipocyte complement-related protein of 30 kilodaltons by 414
PPARgamma agonists: a potential mechanism of insulin sensitization. 415
Endocrinology 2002;143:998-1007. 416
31. Kralisch S, Klein J, Bluher M, Paschke R, Stumvoll M, Fasshauer M.
417
Therapeutic perspectives of adipocytokines. Expert Opin Pharmacother 418
2005;6:863-872. 419
32. Kovacic S, Soltys CL, Barr AJ, Shiojima I, Walsh K, Dyck JR. Akt activity
420
negatively regulates phosphorylation of AMP-activated protein kinase in the 421
heart. J Biol Chem 2003;278:39422-39427. 422
33. Rayner DV. The relationships between glucose absorption and insulin
423
secretion and the migrating myoelectric complex in the pig. Exp Physiol 424
1991;76:67-76. 425
34. Combs TP, Pajvani UB, Berg AH, Lin Y, Jelicks LA, Laplante M,
426
Nawrocki AR, Rajala MW, Parlow AF, Cheeseboro L, Ding YY, Russell
427
RG, Lindemann D, Hartley A, Baker GR, Obici S, Deshaies Y, Ludgate M,
428
Rossetti L, Scherer PE. A transgenic mouse with a deletion in the
429
collagenous domain of adiponectin displays elevated circulating adiponectin 430
and improved insulin sensitivity. Endocrinology 2004;145:367-383. 431
35. Nawrocki AR, Rajala MW, Tomas E, Pajvani UB, Saha AK, Trumbauer
432
ME, Pang Z, Chen AS, Ruderman NB, Chen H, Rossetti L, Scherer PE.
433
Mice lacking adiponectin show decreased hepatic insulin sensitivity and 434
reduced responsiveness to peroxisome proliferator-activated receptor gamma 435
agonists. J Biol Chem 2006;281:2654-2660. 436
36. Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K.
437
Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the 438
metabolic syndrome. J Clin Invest 2006;116:1784-1792. 439
37. Kaltenbach S, Staiger H, Weisser M, Haas C, Stumvoll M, Machicao F,
440
Haring HU. Adiponectin receptor gene expression in human skeletal muscle
441
cells is not regulated by fibrates and thiazolidinediones. Int J Obes (Lond) 442
2005;29:760-765. 443
38. Li W, Tonelli J, Kishore P, Owen R, Goodman E, Scherer PE, Hawkins
444
MA. Insulin-Sensitizing Effects of Thiazolidinediones Are Not Linked to
445
Adiponectin Receptor Expression in Human Fat or Muscle. Am J Physiol 446
Endocrinol Metab 2007;292:E1301-E1307. 447
448 449
Fig 1. The effect of fasting on AdipoR1 and AdipoR2 gene expression in pigs. 450
Samples of liver, longissimus muscle, visceral adipose tissue (VAT) and subcutaneous 451
adipose tissue (AT) were taken 2 hrs after feeding (Fed) or 24 hrs after feeding 452
(Fasted). The total RNA from each tissue of each pig (10 μg) was electrophoresed 453
and transferred to a nylon membrane. The membranes were hybridized with cDNA 454
probes for adiponectin receptor 1 (AdipoR1), adiponectin receptor 2 (AdipoR2), and 455
β-actin. The mRNA abundance was determined by phosphorimage technology and 456
the densitometeric value for each gene was normalized to β-actin. The data 457
represent the means of 4 crossbred pigs (2 males and 2 females) per treatment. Data 458
were analyzed with ANOVA and Duncan’s new multiple range test was used to 459
evaluate the differences from the fed AT value (set to 100). Each bar represents the 460
mean ± S.E.M.; *, P ≤ 0.05; **, P ≤ 0.01. 461
462
Fig 2. Differentiation-dependent adiponectin, and AdipoR genes expression in 463
porcine adipocytes. Porcine S/V cells were differentiated and on the indicated days 464
(0, 3, 6, 9), total RNA was extracted for detecting adiponectin, AdipoR1, AdipoR2, 465
and β-actin gene expression by Northern analysis. The results are the means of 3 466
independent experiments with S/V cells isolated from 3 different pigs. The average 467
of AdipoR1 day 0 data was set to 100 and other data were expressed as relative 468
abundance to this value. Each point represents the mean ± S.E.M. and mRNA 469
values were normalized to β-actin. Means without a common letter differ 470
significantly, P ≤ 0.05. 471
472
Fig 3. The effect of rosiglitazone and insulin on adiponectin, and AdipoR genes 473
expression in porcine adipocytes. Porcine S/V cells were differentiated for 9 days, 474
as indicated in Fig 2. The 9 days differentiated adipocytes were then cultured in low 475
glucose (7.78 mM) DMEM/F12 with 10 nM insulin for 6 hrs, and then 1 μM insulin 476
or 1 μM rosiglitazone (Rosi) or insulin + Rosi were added for 2 , 12or 24 hrs. Total 477
RNA was extracted to detect adiponectin (panel A), AdipoR1 (panel B), AdipoR2 478
(panel C), and β-actin gene expression by Northern analysis, as indicated in Fig 2. 479
The results are the means of 4 independent experiments using cells isolated from 4 480
different pigs. The average of the 2 h control medium data was set to 100 and other 481
data were expressed on relative abundance to the 2 h data. Each point represents the 482
mean ± S.E.M.; *, P ≤ 0.05. 483
484
Fig 4. The effect of an insulin signal pathway inhibitor on rosiglitazone-mediated 485
induction of expression of AdipoR genes in porcine adipocytes. Porcine S/V cells 486
were differentiated for 9 days and then differentiated adipocytes were cultured in low 487
glucose (7.78 mM) DMEM/F12 with 10 nM insulin for 6 h. Control cells were 488
incubated in this medium for an additional 24 h. After 6 h, 1 μM insulin (I) plus 1 489
μM rosiglitazone (R) were added to some cells and incubation was continued for 24 h. 490
Also at 6 h, the MAPK inhibitor, PD98059 at 25 μM (PD) or the PI3K inhibitor, 491
LY294002 at 10 μM (LY) were added along with I + R; incubation was continued for 492
24 h. Total RNA was extracted for detecting AdipoR1, AdipoR2, and β-actin gene 493
expression by Northern analysis as indicated in Fig 2. The results are the means of 4 494
independent experiments using cells isolated from 4 different pigs and the average of 495
the 24 h control medium data was set to 100 with other data expressed as relative 496
abundance to the control value. Each bar represents the mean ± S.E.M. and mRNA 497
values were normalized to β-actin. Means without a common letter differ 498
significantly, P ≤ 0.05. 499
500
Fig 5. The effects of insulin and rosiglitazone on the expression of adiponectin and 501
AdipoR genes in porcine adipocytes. Porcine S/V cells were differentiated for 9 502
days and then differentiated adipocytes were cultured in low glucose (7.78 mM) 503
DMEM/F12 without 10 nM insulin for 6 hrs, then 10 nM insulin (10 nM I), 1 μM 504
insulin (I), 1 μM rosiglitazone (Rosi), or 10 μM LY294002 (LY) were added to 505
separate plates for 24 hrs. Total RNA was extracted for detecting adiponectin, 506
AdipoR2, and β-actin gene expression by Northern analysis as indicated in Fig 2. 507
The results are the means of 4 independent experiments using cells isolated from 4 508
different pigs and the average of 24 hrs control medium data was set to 100 with other 509
data expressed as relative abundance to the control value. Each bar represents the 510
mean ± S.E.M. and mRNA values were normalized to β-actin. Means without a 511
common letter differ significantly, P ≤ 0.05. 512
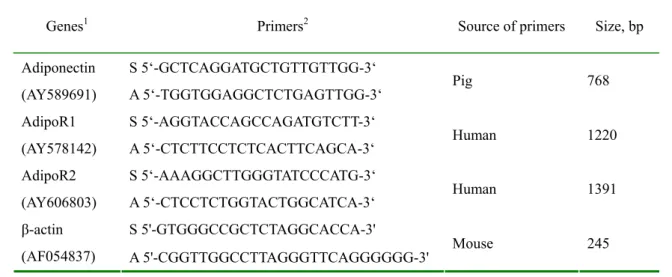
Table 1. List of the Northern analysis probes 513
Genes1 Primers2 Source of primers Size, bp S 5‘-GCTCAGGATGCTGTTGTTGG-3‘
Adiponectin
(AY589691) A 5‘-TGGTGGAGGCTCTGAGTTGG-3‘ Pig 768
S 5‘-AGGTACCAGCCAGATGTCTT-3‘ AdipoR1
(AY578142) A 5‘-CTCTTCCTCTCACTTCAGCA-3‘ Human 1220 S 5‘-AAAGGCTTGGGTATCCCATG-3‘
AdipoR2
(AY606803) A 5‘-CTCCTCTGGTACTGGCATCA-3‘ Human 1391 S 5'-GTGGGCCGCTCTAGGCACCA-3'
β-actin
(AF054837) A 5'-CGGTTGGCCTTAGGGTTCAGGGGGG-3' Mouse 245
514
1 GenBank accession number is indicated parenthetically.
515 516 2 S: sense; A: antisense. 517 518 519
A
520AT VAT Muscle Liver
AdipoR1 mRNA abundance,
arbit ray units 0 20 40 60 80 100 120 140 160 180 Feeding Fasting
*
*
521 AdipoR1 522 β-actin 523 524B
525AT VAT Muscle Liver
AdipoR2 mRNA abundance, arbitray unit s 0 20 40 60 80 100 120 140 160 Feeding Fasting
**
P = 0.06 526 AdipoR2 527 β-actin 528 529 Fig 1. 530531 532 533 534 535 536 537 538 539 540 541 542 543 544 545 546 547 Adiponectin 548 AdipoR1 549 AdipoR2 550 β-actin 551 Fig 2. 552 553 554 555 556 557
0 day
3 day
6 day
9 day
R
e
lative m
R
NA abundanc
e,
arbitray unit
s
0 500 1000 1500 2000 2500 AdipoR1 AdipoR2 Adiponectinb
b
b
b
d
de
e
e
c
a
A
5582h
12h
24h
Adiponectin relative mRNA abundance , arbit ray unit s 60 80 100 120 140 160 180 200Control
Insulin
Rosi
Insulin+Rosi
*
559 560B
5612 h
12 h
24 h
AdipoR1 mRNA abundance, arbitray units
60 80 100 120 140 160 180 200
Control
Insulin
Rosi
Insulin+Rosi
562C
5632 h
12 h
24 h
AdipoR2 mRNA abundanc
e, ar bitray units 40 60 80 100 120 140 160 180 200
Control
Insulin
Rosi
Insulin+Rosi
*
564 Fig 3. 565AdipoR1
AdipoR2
AdipoRs relative mRNA abundance, arbitray units
0 50 100 150 200 250
Control 24h
In + Rosi 24h
In + Rosi + LY 24h
In + Rosi + PD 24h
a
b
b
c
566 AdipoR1 567 AdipoR2 568 β-actin 569 570 Fig 4. 571A d ip o n e c tin A d ip o R 1
A d ip o R 2
Relative mRNA abunda
nce, arbitray units
0 5 0 1 0 0 1 5 0 2 0 0 c o n tr o l 1 0 n M I 1 u M I L y 1 u M I + L y a b b c c a b a c c b
A d ip o n e c tin A d ip o R 1
A d ip o R 2
Relative mRNA abundance, arbitray units
0 5 0 1 0 0 1 5 0 2 0 0 2 5 0 c o n tr o l R o s i R o s i+ L Y I+ R o s i I+ R o s i+ L Y a b b a b a b a d c d b c 572 573 574 575 576 577 578 579 580 581 582 583 584 585 586 587 588 589 Fig 5. 590
A
B
行政院國家科學委員會補助國內專家學者出席國際學術會議報告
96 年 5 月 14 日 報告人姓名 丁詩同 服務機構及職稱 國立台灣大學動物科學技術學系 教授 時間 會議 地點 95 年 4 月 26 日至 5 月 3 日 美國華盛頓首府 本會核定 補助文號94-2313-B-002-024
會議 名稱 (中文)實驗生物學聯合會和美國農部區域討論會(英文)Experimental Biology 2007 and USDA NCC0097 發表
論文 題目
(中文)猪 PPAR d 在脂肪細胞分化的功能
( 英 文 )
The functionality of porcine peroxisomal proliferator
activated receptor delta in adipocyte differentiation
報告內容應包括下列各項:
一、 參加會議經過
April 26 Flew to Washington DC, USA. Stay in Red Roof Inn, 500 H Street NW, Washington DC 20001.
April 27 Attended the meeting for USDA regional meeting NCC0097, Presented a talk on two topics: The function of PPARg and PPARd in pigs and The expression of genes in porcine adipose tissue under the treatment of porcine serum amyloid protein A. The fee for the meeting is 70 US dollars. Went to the dinner function with the scientists with common interests in adipocyte biology. The dinner cost for 67 US dollars. April 28. Attended the Keynote speech by Two scientist both named Tony on the discovery of tyrosine kinases and its function in regulating physiological process and gene expression in ASBMB Biochemistry session. Attended the poster section on lipid metabolism. Attended the evening session on enzyme expression regulation.
April 29. Attended the Keynote speech on Phosphoinisitide molecules and the genes involved in making the enzymes. What we learn from yeast model. Also attend Lipid metabolism. Went to Symposium on animal model for human nutrition 1 delivered by DH Baker and 2 delivered by Spurlock on adipocyte models. Also went to a
biochemistry teaching for cultivating future strong biochemist. The speakers were too good. But picked up some points, including bring research into classroom, set the expectation high, recognize the students, enthusiasm in research is contagous, and interactive teaching to enhance student learning. We also posted our poster. The full length of the report is attached at the end of the report.
April 30. Attended the ASBMB Merck Award for the talk on PEPC Kinase by R.W. Hanson. I also attended a section on role of nuclear receptors in metabolic syndrome in the morning. Went to poster section for genetics and metabolic approaches to obesity and proteionics: proteomics and bioinformatics. Went to Symposium on lipid
metabolism and transport and also lipid signaling track.
May 1. I attended the Nutrition section and mostly the poster and two keynote speeches. Two areas are very important, one was on companion animal nutrition and the other was on aging. There were lots of research on the feed limitation on aging and well-being of animals. Two specific experiments on monkey long term
restriction on feeding improve the health condition and longevity.
May 2. Attended the FASEB Excellence in Science Award before we took off to Columbus, Ohio for an international exchange program discussion.
表 Y04
二、 與會心得
This year I started to get the feeling of how well we have done researchwise. We have demonstrated a very specific pathway of regulating nutrition metabolism and such a finding is world-class. We need to collect the most recent progress of the research in order to know where we are and what to do to compete with international scientists. This meeting always has a lot of activities going on. Lots of science to learn, but too little time available. I have also got to interact with lots of scientist in my field of research. That help me develop the sense of where we are and our competitiveness.
三、 考察參觀活動(無是項活動者省略)
四、 建議
The areas of nutritional science research are evolving very fast during the past few years. This Meeting collects a broad spectrum of research progress which is
important for the researchers in Taiwan. I would suggest that we should encourage researchers to go and joint this meeting to get updated research progress report and to improve our research quality.
五、 攜回資料名稱及內容
Experimental Biology 2007, Conference information and scientific program in a CD format. All the station reports from the USDA regional meeting NCR-97.
六、 其他
The full article of our presentation.
Ectopic expression of porcine peroxisome-proliferator-activated receptor delta regulates adipogenesis in myoblasts
Y. H.Yu1 and S. T. Ding1
Department of Animal Science and Technology, National Taiwan University, 50, Lane 155, Kee-Long Rd. Sec. 3, Taipei 106, Taiwan 1
ABSTRACT
It is well known that peroxisome-proliferator-activated receptor γ (PPAR γ) plays a critical role in regulating adipogenesis. In rodents, PPAR δ is expressed before PPAR γ during adipocyte differentiation. Thus, the interaction between PPAR δ and PPAR γ during adipogenesis needs to be elucidated. The current experiment was designed to study the interaction of porcine PPAR δ and PPAR γ in mouse myoblast cells. Inhibition of myogenesis was observed in myoblasts expressing porcine PPAR δ, similar to myoblast expressing PPAR γ. Treatment of myoblasts expressing PPAR δ with ligands for both PPAR δ and γ enhanced lipogenesis to a greater extent than treatment with a PPAR γ ligand alone. The ability to transdifferentiate myoblasts into adipocytes was decreased in myoblasts co-expressing PPAR δ with either wild-type or mutated PPAR γ (serine 112 was mutated to alanine) compared to myoblasts expressing either type of PPAR δ alone. Adipose transdifferentiation in myoblasts co-expressing PPAR δ and mutated PPAR γ was greater than in myoblasts co-expressing PPAR δ and wild-type PPAR γ. Our results suggest that PPAR δ has two different roles in regulating adipogenesis, ie., suppression of myogenesis to enhance transdifferentiation of myoblasts into adipocytes and interaction with PPAR γ to modify adipogenesis. Therefore, PPAR δ may have a significant role in adipogenesis.
Key Words: Adipocyte differentiation, Peroxisome proliferator-activated receptor δ, Peroxisome
proliferator-activated receptor γ.
INTRODUCTION
In rodent, peroxisome-proliferator-activated receptor δ (PPAR δ) is wildly expressed in several tissues, including adipose tissue, intestine, skeletal muscle, lung and heart. The
INTRODUCTION
In rodent, peroxisome-proliferator-activated receptor δ (PPAR δ) is wildly expressed in several tissues, including adipose tissue, intestine, skeletal muscle, lung and heart. The expression of PPAR δ in proliferating preadipocytes is undetectable and increases gradually during adipocyte differentiation (Amri et al., 1995). Preadipocyte overexpressing PPAR δ with long chain fatty acids promotes adipogenesis (Bastie et al., 2000). Ectopic expression of PPAR δ in fibroblasts with long chain fatty acids alone do not induce adipogenesis but stimulation in the presence of PPAR γ ligand (Bastie et al., 1999). Therefore, PPAR δ seems to have a facilitating role in adipogenesis.
The information of porcine PPAR δ is still poorly understood, especially in functional study. In previous studies, we have demonstrated that ectopic expression of porcine PPAR γ induces adipogenesis in myoblasts (Yu et al., 2006). The expression of PPAR δ is earlier than PPAR γ during adipocyte differentiation in rodent adipocytes (Amri et al., 1995). We hypothesize that a relationship between PPAR δ and PPAR γ in regulating adipocyte differentiation. In this study, we created C2C12 myoblasts expressing porcine PPAR δ, or co-expressing PPAR δ with either wild-type or mutated PPAR γ (serine 112 was mutated to alanine). Transfected myoblasts with porcine PPAR δ stimulated adipogenesis after addition of both PPAR δ and PPAR γ ligands, whereas a decreased lipid accumulation was observed in myoblasts co-expressing PPARs compared with expressing PPAR γ alone.
MATERIALS AND METHODS
Stably transformed cells with PPAR δ or PPAR γ and induction of myoblast transdifferentiation
The porcine PPAR δ cDNA was cloned from porcine adipose tissue. The PCR products were cloned into a mammalian expression vector and transfected into C2C12 myoblasts by lipofection. To establish expression of both porcine PPAR δ and PPAR γ cell models, C2C12 myoblasts containing either wild-type PPAR γ or mutated PPAR γ were also transfected with porcine PPAR δ. Myoblasts stably expressing PPAR δ were established by puromycin selection. After drug selection, the cells were cultured without selection medium and allowed to propagate to 80% confluence in DMEM with 10% FBS. Confluent cells were then cultured in adipogenic differentiation medium [DMEM containing 10% fetal bovine serum, 1μM dexamethasone, and 5μg/mL insulin] and with or without 1 μM rosiglitazone, a PPAR γ ligand and 1 μM L165041, a PPAR δ ligand. After 10 days of culture, total RNA was purified to determine gene expression.
Northern blot and statistical analysis
The RNA was separatedby electrophoresis and blotted to nylon membranes. The membrane was prehybridized at 42 ℃ and then hybridized withisotope labeled complementary DNA probes. Hybridization results were quantified by phosphor-image analysis. The densitometric value for an individual transcript in a sample lane was normalized to the densitometric value for the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA in the same lane. The treatment effects were analyzed using an ANOVA procedure to determine the main effects of the form of PPAR δ and PPAR γ in presence or absence of its ligands. Duncan’s new multiple range test was
used to evaluate differences among means (SAS Inst. Inc., Cary, NC). A significant difference indicates that P value is not greater than 0.05.
RESULTS AND DISCUSSION
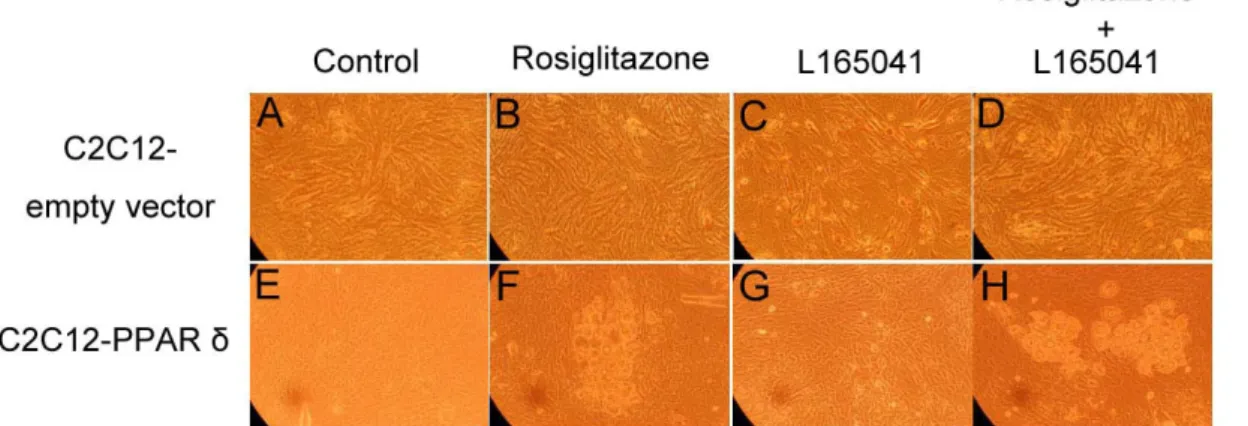
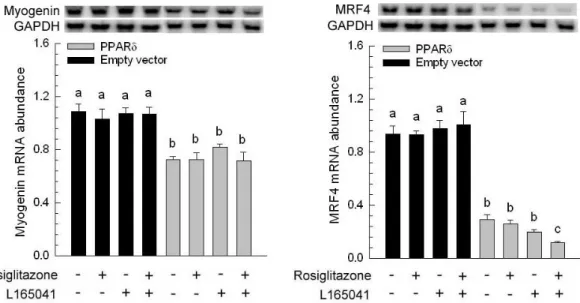
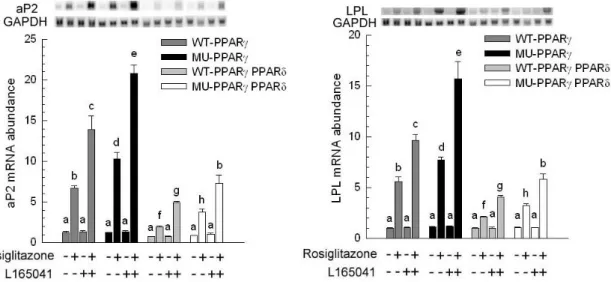
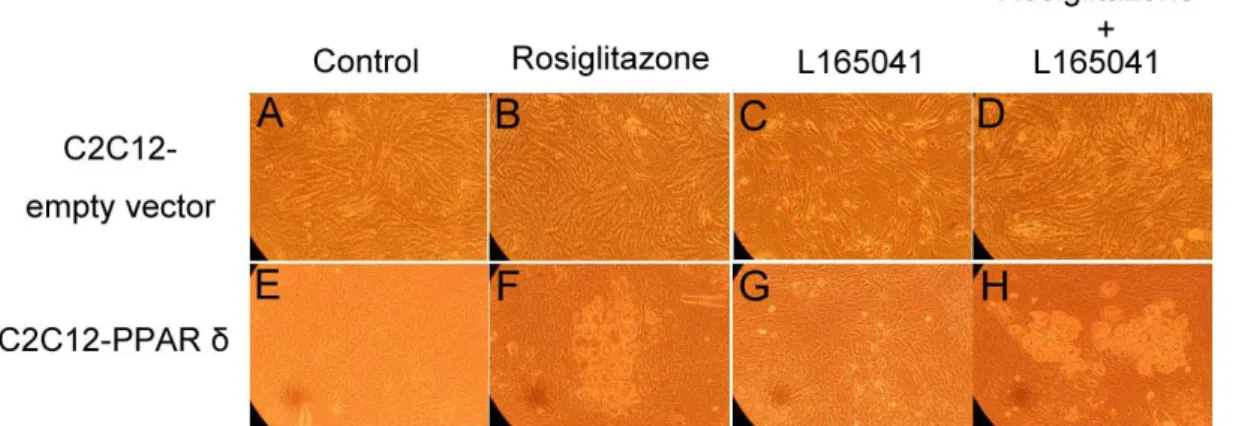
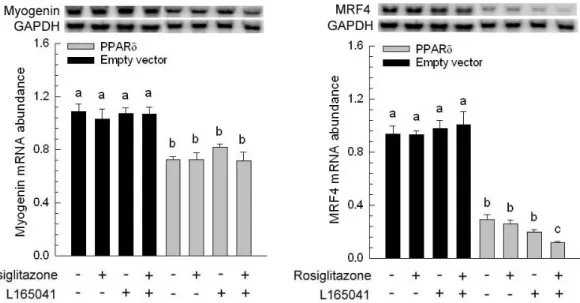
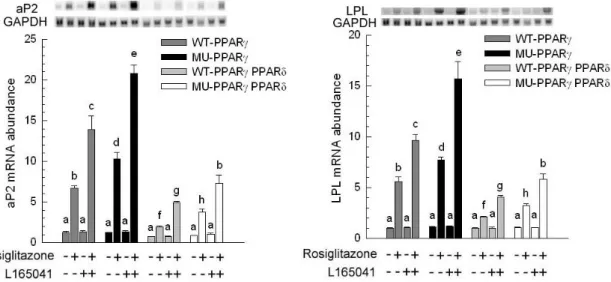
The presence or absence of rosiglitazone and L165041 in adipogenic medium had no effect on myogenesis (Figure 1B, C and D). The myotube formation was inhibited when myoblast expressing PPAR δ compared with transfection of empty vector cells (Figure 1A vs. E). Similar results were observed in our previous studies, myoblasts containing PPAR γ had an ability to interfere in myocyte differentiation (Yu et al., 2006). After exposure of rosiglitazone to the adipogenic differentiation medium for 10 days, lipid-droplets were visualized in myoblasts expressing PPAR δ but absence in addition L165041 in adipogenic differentiation medium (Figure 1). It was well known that ligands for PPARs can activate more than one receptor isoform, hence adipocyte differentiation was increased in medium containing rosiglitazone even if absence of L165041 (Figure 1F). The maximum of lipid accumulation was observed in addition of both PPAR ligands (Figure 1H). This result suggests indirectly that PPAR γ with its ligand has crucial potential in modulating adipocyte differentiation. In loss of function study, it has been demonstrated that lipid accumulation and adipogenic marker genes are decreased in PPAR δ -null adipocytes (Matsusue et al., 2004). In our results, myoblats expressing PPAR δ with PPAR γ and PPAR δ ligands enhanced adipogenesis. It implies that PPAR δ appeared to accelerate adipogenesis. The downstream gene of PPAR γ, adipocyte fatty acid binding protein (aP2) mRNA was highly expressed in myoblasts containing PPAR δ in the presence of rosiglitazone. However, addition of PPAR δ and PPAR γ ligands in adipogenic medium had a greater stimulation of aP2 expression compared with presence of single PPAR ligand. For lipoprotein lipase (LPL) mRNA, it was also increased in the same condition. It has been known that expression of aP2 and LPL are regulated by PPAR γ. Deducing from our results, we hypothesize that high aP2 and LPL transcripts were attributed to PPAR γ function and ectopic PPAR δ modulated PPAR γ expression by binding its peroxisome proliferator response element. A late myogenic marker gene, myogenin was decreased in cells transfected with PPAR δ and both of ligands had no effect on myogenesis (Figure 3). Similar results were observed in another myogenic marker gene, myogenic regulatory factor 4 (MRF4). The suppression of myogenic marker genes in myoblasts expressing PPAR δ was consistent with ectopic expression of PPAR γ in myocytes. These results demonstrated that adipogenesis related transcription factors have the capability of impairing myogenesis. Furthermore, mRNA for aP2 and LPL were expressed at a low level in myoblasts containing either wild-type PPAR γ or mutated PPAR γ and PPAR δ compared with expressing either wild-type PPAR γ or mutated PPAR γ alone (Figure 4). This phenomenon was also found in preadipocyte expressing PPAR δ and PPAR γ. The over-expression of PPAR δ indeed can suppress PPAR γ-mediated adipogenesis (Shi et al., 2002). However, reduction of myogenic genes expression was enhanced in C2C12 myoblasts containing both PPARs (Figure 5). Thus, PPAR δ and PPAR appeared to have a synergic effect in the inhibition of myogenesis.
CONCULSION
In the current study, we demonstrated that PPAR δ has the ability to promote transdifferentiation of myoblasts into adipocytes and interact with PPAR γ to modify adipogenesis. Therefore, PPAR δ may have a significant role in adipogenesis.
REFERENCES
Amri, E., F. Bonino, G. Ailhaud, N. A. Abumrad, and P. A. Grimaldi. 1995. Cloning of a protein that mediates transcriptional effects of fatty acids in preadipocytes. J. Biol. Chem. 270: 2367-2371.
Bastie, C., D. Holst, D. Gaillard, C. Jehl-Pietri, and P. A. Grimaldi. 1999. Expression of peroxisome proliferators-activated receptor PPARδ promotes induction of PPARγ and adipocyte differentiation in 3T3C2 fibroblasts. J. Biol. Chem. 274:21920-21925.
Bastie, C., S. Luquet, D. Holst, C. Jehl-Pietri, and P. A. Grimaldi. 2000. Alterations of peroxisome proliferators-activated receptor δ activity affect fatty acid-controlled adipose differentiation. J. Biol. Chem. 275:38768-38773.
Liu, B. H., C. F. Kuo, Y. C. Wang, and S. T. Ding. 2005. Effect of docosahexaenoic acid and arachidonic acid on the expression of adipocyte determination and differentiation-dependent factor 1 in differentiating porcine adipocytes. J. Anim. Sci. 83:1516-1525.
Matsusue, K., J. M. Peters, and F. J. Gonzalez. 2004. PPARβ/δ potentiates PPARγ-stimulated adipocyte differentiation. FASEB J. 18:1477-1479.
Shi, Y., M. Hon, and R. M. Evans. 2002. The peroxisome proliferator-activated receptor δ, an integrator of transcriptional repression and nuclear receptor signaling. Proc. Natl. Acad. Sci. USA 99:2613-2618.
Yu, Y. H., B. H. Liu, H. J. Mersmann, and S. T. Ding. 2006. Porcine peroxisome proliferator-activated receptor gamma induces transdifferentiation of myocytes into adipocytes. J. Anim. Sci. 84:2655-2665.
Figure 1. Ligand-induced morphological alterations and accumulation of lipid droplets.
Microscographs of C2C12 myocyte with empty vector (A-D), C2C12 expressing porcine peroxisome proliferator-activated receptor δ (PPAR δ; E-H) are shown. Cells were maintained in adipogenic medium (Dulbecco’s modified Eagle medium/dexamethasone/insulin/10% fetal bovine serum) ± 1 μM rosiglitazone (B and F), 1 μM L165041 (C and G) and both of ligands (D and H) to
d 10 postconfluence. Magnification was 60.
Figure 2. Expression of adipogenic marker genes. After confluence, C2C12 myocytes with empty
vector (Empty vector) and C2C12 expressing porcine peroxisome proliferator-activated receptor δ (PPAR δ) were cultured for 10 d. The expression of adipocyte-specific genes [adipocyte fatty acid-binding protein (aP2) and lipoprotein lipase (LPL)] was determined and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The bars indicate the means ± SE for cells from 3 independent replicates (n = 3). ND = not detected. a–c Means without a common letter differ, P < 0.05.
Figure 3. Expression of myogenic marker genes. After confluence, C2C12 myocyte with empty
vector (Empty vector) and C2C12 expressing porcine peroxisome proliferator-activated receptor δ (PPAR δ) were cultured for 10 d. The expression of myogenic genes [Myogenin and myogenic regulatory factor-4 (MRF4)] was determined and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The bars indicate the means ± SE for cells from 3 independent replicates
(n = 3). a–c Means without a common letter differ, P < 0.05.
Figure 4. Expression of adipogenic marker genes. After confluence, C2C12 expressing wild-type porcine PPAR γ (WT-PPAR γ), C2C12 expressing mutated porcine PPAR γ (MU-PPAR γ), C2C12 expressing wild-type porcine PPAR γ and PPAR δ (WT-PPAR γ PPAR δ) and C2C12 expressing mutated porcine PPAR γ and PPAR δ (MU-PPAR γ PPAR δ) were cultured for 10 d. The expression of adipocyte-specific genes [adipocyte fatty acid-binding protein (aP2) and lipoprotein lipase (LPL)] was determined and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The bars indicate the means ± SE for cells from 3 independent replicates (n = 3). a–c Means without a common letter differ, P < 0.05.
Figure 5. Expression of myogenic marker genes. After confluence, C2C12 expressing wild-type
porcine PPAR γ (WT-PPAR γ), C2C12 expressing mutated porcine PPAR γ (MU-PPAR γ), C2C12 expressing wild-type porcine PPAR γ and PPAR δ (WT-PPAR γ PPAR δ) and C2C12 expressing mutated porcine PPAR γ and PPAR δ (MU-PPAR γ PPAR δ) were cultured for 10 d. The expression of myogenic genes [Myogenin and myogenic regulatory factor-4 (MRF4)] was determined and
normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The bars indicate the means ± SE for cells from 3 independent replicates (n = 3). ND = not detected. a–c Means without a common letter differ, P < 0.05.
行政院國家科學委員會補助國內專家學者出席國際學術會議報告
96 年 5 月 14 日 報告人姓名 丁詩同 服務機構及職稱 國立台灣大學動物科學技術學系 教授 時間 會議 地點 95 年 4 月 26 日至 5 月 3 日 美國華盛頓首府 本會核定 補助文號94-2313-B-002-024
會議 名稱 (中文)實驗生物學聯合會和美國農部區域討論會(英文)Experimental Biology 2007 and USDA NCC0097 發表
論文 題目
(中文)猪 PPAR d 在脂肪細胞分化的功能
( 英 文 )
The functionality of porcine peroxisomal proliferator
activated receptor delta in adipocyte differentiation
報告內容應包括下列各項:
一、 參加會議經過
April 26 Flew to Washington DC, USA. Stay in Red Roof Inn, 500 H Street NW, Washington DC 20001.
April 27 Attended the meeting for USDA regional meeting NCC0097, Presented a talk on two topics: The function of PPARg and PPARd in pigs and The expression of genes in porcine adipose tissue under the treatment of porcine serum amyloid protein A. The fee for the meeting is 70 US dollars. Went to the dinner function with the scientists with common interests in adipocyte biology. The dinner cost for 67 US dollars. April 28. Attended the Keynote speech by Two scientist both named Tony on the discovery of tyrosine kinases and its function in regulating physiological process and gene expression in ASBMB Biochemistry session. Attended the poster section on lipid metabolism. Attended the evening session on enzyme expression regulation.
April 29. Attended the Keynote speech on Phosphoinisitide molecules and the genes involved in making the enzymes. What we learn from yeast model. Also attend Lipid metabolism. Went to Symposium on animal model for human nutrition 1 delivered by DH Baker and 2 delivered by Spurlock on adipocyte models. Also went to a
biochemistry teaching for cultivating future strong biochemist. The speakers were too good. But picked up some points, including bring research into classroom, set the expectation high, recognize the students, enthusiasm in research is contagous, and interactive teaching to enhance student learning. We also posted our poster. The full length of the report is attached at the end of the report.
April 30. Attended the ASBMB Merck Award for the talk on PEPC Kinase by R.W. Hanson. I also attended a section on role of nuclear receptors in metabolic syndrome in the morning. Went to poster section for genetics and metabolic approaches to obesity and proteionics: proteomics and bioinformatics. Went to Symposium on lipid
metabolism and transport and also lipid signaling track.
May 1. I attended the Nutrition section and mostly the poster and two keynote speeches. Two areas are very important, one was on companion animal nutrition and the other was on aging. There were lots of research on the feed limitation on aging and well-being of animals. Two specific experiments on monkey long term
restriction on feeding improve the health condition and longevity.
May 2. Attended the FASEB Excellence in Science Award before we took off to Columbus, Ohio for an international exchange program discussion.
表 Y04
二、 與會心得
This year I started to get the feeling of how well we have done researchwise. We have demonstrated a very specific pathway of regulating nutrition metabolism and such a finding is world-class. We need to collect the most recent progress of the research in order to know where we are and what to do to compete with international scientists. This meeting always has a lot of activities going on. Lots of science to learn, but too little time available. I have also got to interact with lots of scientist in my field of research. That help me develop the sense of where we are and our competitiveness.
三、 考察參觀活動(無是項活動者省略)
四、 建議
The areas of nutritional science research are evolving very fast during the past few years. This Meeting collects a broad spectrum of research progress which is
important for the researchers in Taiwan. I would suggest that we should encourage researchers to go and joint this meeting to get updated research progress report and to improve our research quality.
五、 攜回資料名稱及內容
Experimental Biology 2007, Conference information and scientific program in a CD format. All the station reports from the USDA regional meeting NCR-97.
六、 其他
The full article of our presentation.
Ectopic expression of porcine peroxisome-proliferator-activated receptor delta regulates adipogenesis in myoblasts
Y. H.Yu1 and S. T. Ding1
Department of Animal Science and Technology, National Taiwan University, 50, Lane 155, Kee-Long Rd. Sec. 3, Taipei 106, Taiwan 1
ABSTRACT
It is well known that peroxisome-proliferator-activated receptor γ (PPAR γ) plays a critical role in regulating adipogenesis. In rodents, PPAR δ is expressed before PPAR γ during adipocyte differentiation. Thus, the interaction between PPAR δ and PPAR γ during adipogenesis needs to be elucidated. The current experiment was designed to study the interaction of porcine PPAR δ and PPAR γ in mouse myoblast cells. Inhibition of myogenesis was observed in myoblasts expressing porcine PPAR δ, similar to myoblast expressing PPAR γ. Treatment of myoblasts expressing PPAR δ with ligands for both PPAR δ and γ enhanced lipogenesis to a greater extent than treatment with a PPAR γ ligand alone. The ability to transdifferentiate myoblasts into adipocytes was decreased in myoblasts co-expressing PPAR δ with either wild-type or mutated PPAR γ (serine 112 was mutated to alanine) compared to myoblasts expressing either type of PPAR δ alone. Adipose transdifferentiation in myoblasts co-expressing PPAR δ and mutated PPAR γ was greater than in myoblasts co-expressing PPAR δ and wild-type PPAR γ. Our results suggest that PPAR δ has two different roles in regulating adipogenesis, ie., suppression of myogenesis to enhance transdifferentiation of myoblasts into adipocytes and interaction with PPAR γ to modify adipogenesis. Therefore, PPAR δ may have a significant role in adipogenesis.
Key Words: Adipocyte differentiation, Peroxisome proliferator-activated receptor δ, Peroxisome
proliferator-activated receptor γ.
INTRODUCTION
In rodent, peroxisome-proliferator-activated receptor δ (PPAR δ) is wildly expressed in several tissues, including adipose tissue, intestine, skeletal muscle, lung and heart. The