Isolates with Reduced Susceptibility to Fluconazole
Yun-Liang Yang1,2, Chih-Chao Lin3, Te-Pin Chang3, Tsai-Ling Lauderdale3, Hui-Ting Chen2,3, Ching-Fu Lee4, Chih-Wen Hsieh4, Pei-Chen Chen3, Hsiu-Jung Lo3,5*1 Department of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan, 2 Institute of Molecular Medicine and Bioengineering, National Chiao Tung University, Hsinchu, Taiwan,3 National Institute of Infectious Diseases and Vaccinology, National Health Research Institutes, Miaoli, Taiwan, 4 Department of Applied Science, National Hsinchu University of Education, Hsinchu, Taiwan,5 School of Dentistry, China of Medical University, Taichung, Taiwan
Abstract
Infections caused by treatment-resistant non-albicans Candida species, such as C. tropicalis, has increased, which is an emerging challenge in the management of fungal infections. Genetically related diploid sequence type (DST) strains of C. tropicalis exhibiting reduced susceptibility to fluconazole circulated widely in Taiwan. To identify the potential source of these wildly distributed DST strains, we investigated the possibility of the presence in soil of such C. tropicalis strains by pulsed field gel electrophoresis (PFGE) and DST typing methods. A total of 56 C. tropicalis isolates were recovered from 26 out of 477 soil samples. Among the 18 isolates with reduced susceptibility to fluconazole, 9 belonged to DST149 and 3 belonged to DST140. Both DSTs have been recovered from our previous studies on clinical isolates from the Taiwan Surveillance of Antimicrobial Resistance of Yeasts (TSARY) program. Furthermore, these isolates were more resistant to agricultural azoles. We have found genetically related C. tropicalis exhibiting reduced susceptibility to fluconazole from the human hosts and environmental samples. Therefore, to prevent patients from acquiring C. tropicalis with reduced susceptibility to azoles, prudent use of azoles in both clinical and agricultural settings is advocated.
Citation: Yang Y-L, Lin C-C, Chang T-P, Lauderdale T-L, Chen H-T, et al. (2012) Comparison of Human and Soil Candida tropicalis Isolates with Reduced Susceptibility to Fluconazole. PLoS ONE 7(4): e34609. doi:10.1371/journal.pone.0034609
Editor: Kirsten Nielsen, University of Minnesota, United States of America Received December 26, 2011; Accepted March 6, 2012; Published April 5, 2012
Copyright: ß 2012 Yang et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This study was supported by 99A1-ID-PP-04-014 and 00A 1-ID-PP-04-014 from National Health Research Institutes and NSC 99-2320-B-400-006-MY3 and NSC 99-2320-B-009-001-MY3 from National Science Council in Taiwan. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist. * E-mail: [email protected]
Introduction
Due to the elevated number of risk populations, the prevalence of fungal infections has increased significantly in past two decades. Candida species are the most frequently isolated fungal pathogens causing morbidity and mortality in seriously immunocompromised hosts. Although Candida albicans is the most prevalent species in hospitalized individuals and in nosocomial infections, there has been a shift toward the more treatment-resistant non-albicans Candida species [1,2,3,4,5]. This has become an emerging issue in the management of fungal infection. The prevalence of these species differed significantly in various geographic areas [4,5,6,7,8,9,10]. Candida glabrata was the most frequently isolated species in Western countries [4,11,12], whereas C. tropicalis predominated in Asia [5,13,14,15]. For the treatment, azoles, echinocandins, polyenes, and 5-flucytosine are the four major classes of antifungal drugs. Due to low cost and less side effects, fluconazole has become one of the most commonly prescribed drugs.
Among the phenomena associated with azole resistance, ‘trailing’ describes the reduced but persistent growth that some isolates exhibit at drug concentrations above the minimum inhibitory concentrations (MICs) in broth dilution tests [16]. When the MIC of an isolate measured after 48 hours (h) incubation is approximately 4-fold higher than that at the 24 h point [17], the isolate is defined to have trailing growth. Thus, in
the present study, isolates with fluconazole MICs$64 mg/L or voriconazole MICs$4 mg/L after 48 h incubation were consid-ered to have reduced susceptibility to azole drugs.
In order to monitor the trends of species distribution and drug susceptibilities of yeast pathogens, the Taiwan Surveillance of Antimicrobial Resistance of Yeasts (TSARY) program was initiated in 1999 [18]. Subsequently, two more rounds of TSARY were conducted in 2002 and 2006. Previously, we found 23 of the 162 C. tropicalis isolates collected from TSARY in 1999 with fluconazole MICs$64 mg/L to be closely related despite being collected from different hospitals throughout Taiwan [19,20]. Furthermore, 5 of the 23 isolates exhibiting reduced susceptibility to fluconazole were from hospital N4 and all belonged to the same diploid sequence type (DST), DST140 (allele combination, 1, 3, 3, 17, 54, and 3 for ICL1, MDR1, SAPT2, SAPT4, XYR1, and ZWF1a, respectively). However, we did not find any evidence to suggest that these five isolates were transmitted horizontally from person to person [21]. In addition, 2 genetically closely related DST C. tropicalis strains, DST140 and DST98 (allele combination, 1, 3, 3, 17, 9, and 3), exhibited reduced susceptibility to fluconazole were identified in TSARY 1999 and 2006. Among them, 18 DST140 isolates were recovered from 10 different hospitals located in all 4 geographic regions in Taiwan and 7 DST98 ones were recovered from 4 different hospitals, two each in northern and southern Taiwan. There were also 3 DST149 (allele combination, 1, 44, 3, 7, 58, and 3) isolates exhibiting reduced
susceptibility to fluconazole and from 3 hospitals located in northern Taiwan were identified. These results indicated that those DST strains exhibiting reduced susceptibility to fluconazole circulated widely in Taiwan from 1999 to 2006 and their presence was not a result of outbreaks in certain hospitals or geographic regions [21].
Exposure to azole compounds paves the way for the selection and enrichment of fungal isolates exhibiting reduced susceptibility to drugs, which may occur in patients receiving azole treatments. However, emergence of person to person transmission of isolates exhibiting reduced susceptibility to fluconazole during medical treatment is unlikely. Alternatively, the use of azole compounds in the environment may select organisms exhibiting reduced susceptibility to drugs, which may find their ways to human. Candida tropicalis is prevalent in organically enriched soil and aquatic environments [22] as well as in wild birds [23]. Thus, in the present study, we investigated the presence in soil of these C. tropicalis strains exhibiting reduced susceptibility to fluconazole, especially DST98, DST140, and DST149.
Results
Candida tropicalis isolates
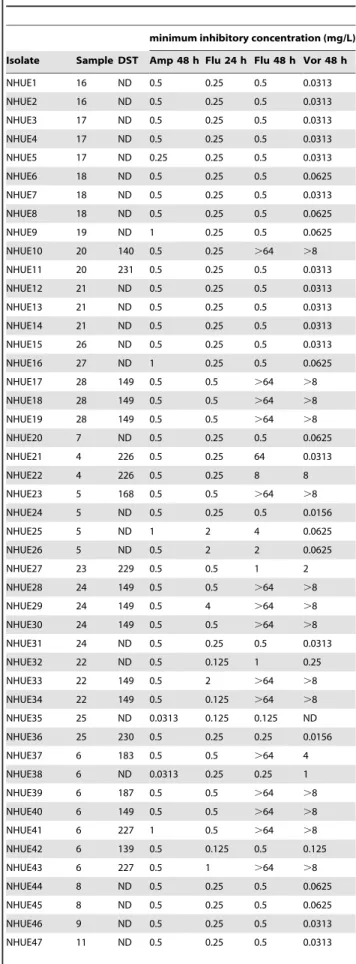
A total of 56 C. tropicalis isolates (Table 1) analyzed in the present study were recovered from 26 of the 477 soil samples collected from 2006 to 2008 around Taiwan. The positive culture rates of C. tropicalis from various types of soil differed significantly. It was 85.7% (6/7) for petroleum-contaminated soil samplings, 44.8% (13/29) for agricultural fields, 38.5% (5/13) for sludge soil, and 0.5% (2/427) for forest soil. To identify whether multiple strains were present in a sample, three individual colonies (if present) of each representative morphotype in every soil sample were picked for subsequent workup. In the present study, 10 samples had only one C. tropicalis isolate recovered, 8 had 2, 5 had 3, 2 had 4, and 1 had 7(Table 1). All isolates were susceptible to amphotericin B. Among the 18 isolates with fluconazole MICs$64 mg/L after 48 h incubation, 17 had voriconazole MICs$4 mg/L as well. Nevertheless, they all exhibited trailing growth phenotype.
Molecular typing
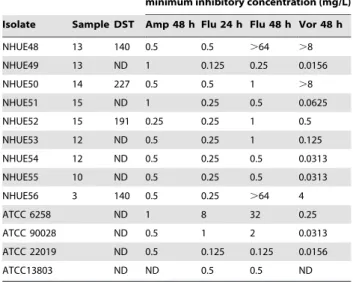
To determine whether the C. tropicalis isolates with reduced susceptibility to fluconazole recovered from soil samples were genetically related to those from human, we applied pulsed field gel electrophoresis (PFGE) method to analyze their genetic relatedness. The 18 isolates that were investigated included 11 soil and 7 human isolates. The soil isolates comprised 6 isolates exhibiting reduced susceptibility to fluconazole (NHUE10, 28, 33, 40, 48, and 56) and 5 susceptible ones (NHUE11, 27, 36, 42, and 52). The 7 clinical isolates (YM990275, 490, 537, 579, 592, 649, and 659) all exhibited reduced susceptibility to fluconazole. As expected, all 6 DST140 isolates from human shared an indistinguishable PFGE pattern (Fig. 1). In addition, 2 isolates exhibiting reduced susceptibility to fluconazole from soil, NHUE48 and NHUE56, shared the same PFGE pattern with those human isolates. Among the remaining 4 soil isolates exhibiting reduced susceptibility to fluconazole, 3 (NHUE28, NHUE33, NHUE40), from different samples, shared at least 97% similarity. Furthermore, fluconazole susceptible isolates recovered from soil were not closely related to the ones exhibiting reduced susceptibility to fluconazole from the same areas, i.e. NHUE 11 vs. NHUE 10, NHUE 36 vs. NHUE 33, and NHUE 42 vs. NHUE 40. The fact that YM990579, a DST149 human isolate, also shared the indistinguishable PFGE pattern with the six DST140 human isolates, suggests that MLST has a higher discriminatory
Table 1. Characteristics of Candida tropicalis isolates from soils (N = 56).
minimum inhibitory concentration (mg/L) Isolate Sample DST Amp 48 h Flu 24 h Flu 48 h Vor 48 h NHUE1 16 ND 0.5 0.25 0.5 0.0313 NHUE2 16 ND 0.5 0.25 0.5 0.0313 NHUE3 17 ND 0.5 0.25 0.5 0.0313 NHUE4 17 ND 0.5 0.25 0.5 0.0313 NHUE5 17 ND 0.25 0.25 0.5 0.0313 NHUE6 18 ND 0.5 0.25 0.5 0.0625 NHUE7 18 ND 0.5 0.25 0.5 0.0313 NHUE8 18 ND 0.5 0.25 0.5 0.0625 NHUE9 19 ND 1 0.25 0.5 0.0625 NHUE10 20 140 0.5 0.25 .64 .8 NHUE11 20 231 0.5 0.25 0.5 0.0313 NHUE12 21 ND 0.5 0.25 0.5 0.0313 NHUE13 21 ND 0.5 0.25 0.5 0.0313 NHUE14 21 ND 0.5 0.25 0.5 0.0313 NHUE15 26 ND 0.5 0.25 0.5 0.0313 NHUE16 27 ND 1 0.25 0.5 0.0625 NHUE17 28 149 0.5 0.5 .64 .8 NHUE18 28 149 0.5 0.5 .64 .8 NHUE19 28 149 0.5 0.5 .64 .8 NHUE20 7 ND 0.5 0.25 0.5 0.0625 NHUE21 4 226 0.5 0.25 64 0.0313 NHUE22 4 226 0.5 0.25 8 8 NHUE23 5 168 0.5 0.5 .64 .8 NHUE24 5 ND 0.5 0.25 0.5 0.0156 NHUE25 5 ND 1 2 4 0.0625 NHUE26 5 ND 0.5 2 2 0.0625 NHUE27 23 229 0.5 0.5 1 2 NHUE28 24 149 0.5 0.5 .64 .8 NHUE29 24 149 0.5 4 .64 .8 NHUE30 24 149 0.5 0.5 .64 .8 NHUE31 24 ND 0.5 0.25 0.5 0.0313 NHUE32 22 ND 0.5 0.125 1 0.25 NHUE33 22 149 0.5 2 .64 .8 NHUE34 22 149 0.5 0.125 .64 .8 NHUE35 25 ND 0.0313 0.125 0.125 ND NHUE36 25 230 0.5 0.25 0.25 0.0156 NHUE37 6 183 0.5 0.5 .64 4 NHUE38 6 ND 0.0313 0.25 0.25 1 NHUE39 6 187 0.5 0.5 .64 .8 NHUE40 6 149 0.5 0.5 .64 .8 NHUE41 6 227 1 0.5 .64 .8 NHUE42 6 139 0.5 0.125 0.5 0.125 NHUE43 6 227 0.5 1 .64 .8 NHUE44 8 ND 0.5 0.25 0.5 0.0625 NHUE45 8 ND 0.5 0.25 0.5 0.0625 NHUE46 9 ND 0.5 0.25 0.5 0.0313 NHUE47 11 ND 0.5 0.25 0.5 0.0313
power than PFGE among C. tropicalis isolates. Therefore we further employed MLST to compare the genetic relatedness among all 18 soil isolates exhibiting reduced susceptibility to fluconazole. Among them, 9 from 4 soil samples (#6, 22, 24, and 28) were DST149 and 3 from 2 soil samples (#3 and 20) were DST140 (Table 1). In addition to DST140 and DST149, 10 additional DST patterns were identified from 13 isolates in the present study. They were DST139 (allele combination, 1, 4, 22, 23, 36, and 9), DST168 (allele combination, 1, 3, 3, 17, 57, and 3), DST183 (allele combination, 1, 20, 12, 17, 68, and 3), DST187 (allele combination, 1, 63, 2, 30, 69, and 25), DST191 (allele combination, 9, 54, 2, 10, 70, and 9), DST226 (allele combination, 1, 80, 27, 10, 44, and 9), DST227 (allele combination, 5, 54, 1, 47, 48, and 22), DST229 (allele combination, 1, 82, 3, 17, 57, and 3), DST230 (allele combination, 1, 4, 12, 10, 3, and 9), and DST231 (allele combination, 27, 9, 1, 43, 23, and 7). DST140 and DT168 are closely related. They are single locus variants and differ by only one SNP (nt 242) in the XYR1 gene. Among the 6 tested fluconazole susceptible isolates, NHUE22 (MIC = 8 mg/L) was less susceptible than the other 5 (MIC ,–– 1 mg/L) and they all had their own unique DST. Interestingly, NHUE22 and NHUE21 were both from sample 4 and belonged to DST226.
Growth curves
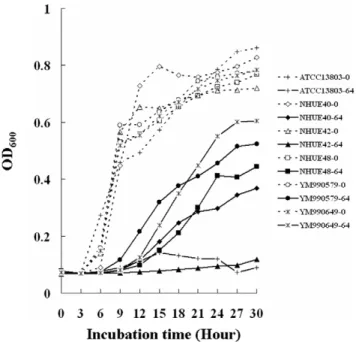
All 18 soil isolates with fluconazole MICs$64 mg/L after 48 h incubation, such as NHUE40, NHUE48, YM9900579, and YM990649, were able to grow in the presence of 64 mg/L fluconazole (Fig. 2). In contrast, the growth of the susceptible ones Table 1. Cont.
minimum inhibitory concentration (mg/L) Isolate Sample DST Amp 48 h Flu 24 h Flu 48 h Vor 48 h NHUE48 13 140 0.5 0.5 .64 .8 NHUE49 13 ND 1 0.125 0.25 0.0156 NHUE50 14 227 0.5 0.5 1 .8 NHUE51 15 ND 1 0.25 0.5 0.0625 NHUE52 15 191 0.25 0.25 1 0.5 NHUE53 12 ND 0.5 0.25 1 0.125 NHUE54 12 ND 0.5 0.25 0.5 0.0313 NHUE55 10 ND 0.5 0.25 0.5 0.0313 NHUE56 3 140 0.5 0.25 .64 4 ATCC 6258 ND 1 8 32 0.25 ATCC 90028 ND 0.5 1 2 0.0313 ATCC 22019 ND 0.5 0.125 0.125 0.0156 ATCC13803 ND ND 0.5 0.5 ND DST, diploid sequence type; h, hours; ND, not determined; amp, amphotericin B; flu, fluconazole; vor, voriconazole;
doi:10.1371/journal.pone.0034609.t001
Figure 1. Dendrogram of 18Candida tropicalisisolates from human and soil. The Dice coefficient was used to analyze the similarities between the PFGE band patterns. UPGMA was used for the cluster analysis. The position tolerance and optimization were set at 1%. A total of 7 isolates were recovered from human (YMs) and 11 from soil (NHUEs). MIC refers to fluconazole MIC (in mg/L). An MIC of 64 indicates isolates with reduced susceptibility.
was inhibited, such as NHUE42 (Fig. 2, triangles solid line) and ATCC13803 (Fig. 2, crosses solid line). Hence, those isolates with fluconazole MICs$64 mg/L definitely had reduced susceptibility to fluconazole than those isolates with lower fluconazole MICs. Susceptibilities to drugs used in agriculture
Since 17 of the 18 soil isolates exhibiting reduced susceptibility to fluconazole also had high voriconazole MICs, we selected 6 isolates to investigate whether isolates with reduced susceptibility to fluconazole from human and soil also had reduced susceptibility to other azole drugs used in agriculture. The relative growth of 6 isolates after incubation for 48 h is showed in Table 2. In the presence of 64 mg/L fluconazole, isolates exhibiting fluconazole reduced susceptibility, NHEU40, NHUE48, YM990579, and YM990649 grew better (66–100% of growth) than the fluconazole
susceptible ones, ATCC13803 and NHUE42 (27–32% of growth), confirming our previous results obtained from broth microdilution method. All tested isolates grew poorly (10–11% of growth) in the presence of 64 mg/L penconazole and tebuconazole. Interesting-ly, the 4 isolates exhibiting fluconazole reduced susceptibility grew better than fluconazole susceptible ones in the presence of 4 mg/L penconazole (36–60% vs. 23–24% of growth), 4 mg/L tebucona-zole (54–83% vs. 12–15% growth), 16 mg/L fluquinconatebucona-zole (71– 87% vs. 41–59% of growth), or 64 mg/L tridimenol (24–35% vs. 15–17% growth). Hence, isolates exhibiting fluconazole reduced susceptibility were also less susceptible than the fluconazole susceptible ones to other azole drugs used in agriculture. Discussion
Candida tropicalis is one of the most frequently isolated non-albicans Candida species from patients [8,10,24,25]. Furthermore, C. tropicalis develops drug resistance in the presence of fluconazole much more rapidly than other Candida species [26,27]. Our findings in the present study suggest that soil may be a potential source of C. tropicalis with reduced susceptibility to azole type of antifungal drugs.
The observation that 3 C. tropicalis isolates (NHUE28-30) with reduced susceptibility to fluconazole recovered from the same soil sample, #24, belonged to the same DST strain (DST149) suggests that they may be the progenies of the same strain. Nevertheless, one isolate (NHUE31) recovered from the same #24 soil sample was susceptible to fluconazole. On the other hand, of the 7 isolates (NHUE37-43) recovered from another soil sample, #6, 5 exhibited reduced susceptibility to fluconazole and belonged to 4 different DSTs (DST183, 187, 149, and 227). These results indicated that multiple strains of C. tropicalis exist in a single soil sample.
Long duration of drug exposure and high numbers of reproducing microorganisms contribute to the selection and/or enrichment of drug-resistant individuals. In fungi, drug resistance is more likely to be the outcome of sequential accumulation of adaptive mutations in the chromosomes [28,29]. Azole-resistant isolates have been shown to emerge following microbial exposure to the drugs in patients and in agricultural settings. The observation that azole-resistant Aspergillus fumigatus isolates recov-ered from soil and compost were genetically related to clinical resistant isolates suggests the existence of an environmental route for developing drug resistance in fungi [30,31,32]. Similar to some antibiotic resistance in bacteria [33,34,35], fluconazole resistance in fungi also involves step-wise mutations affecting more than one
Figure 2. Growth curves of differentC. tropicalisisolates with different fluconazole susceptibilities. Six isolates, ATCC13803 (rectangles), NHUE40 (diamonds), NHUE42 (triangles), NHUE48 (circles), YM990579 (crosses), and YM990649 (stars) were grown in the RPMI medium 1640 (Gibco BRL31800-022) in the absence (dot lines) or in the presence of 64 mg/L (solid lines) fluconazole.
doi:10.1371/journal.pone.0034609.g002
Table 2. Relative growth of six C. tropicalis strains in the presence of different drugs.
Drug none Fluconazole Penconazole Tebuconazole Tridimenol Fluquinconazole Concentration (mg/L) 4 64 4 64 4 64 4 64 4 16* ATCC13803 100# 341 27 23 11 15 10 49 15 55 41 NHUE40 100 71 71 36 10 58 10 71 34 68 77 NHUE42 100 30 32 24 11 12 10 60 17 75 59 NHUE48 100 87 100 54 11 83 11 78 35 94 87 Ym990579 100 71 66 40 10 59 10 75 35 83 72 Ym990649 100 92 81 60 11 54 11 65 24 100 87 *Due to the solubility of fluquinconazole, the highest concentration of this drug in the present study was 16 mg/L instead of 64 mg/L.
#The growth of each isolate in the absence of drug was defined as 100% and the relative growth of each isolate in the presence of drugs was normalized accordingly. 1Drugs capable of reducing the growth of cells by more than 50% are in bold, considered as susceptible to the concentrations of drugs.
genes [36,37,38]. Therefore, the C. tropicalis isolates from soil exhibiting trailing growth phenomena in the present study are prone to be resistant.
In the present study, we have found that C. tropicalis isolates with fluconazole MICs$64 mg/L recovered from human and from soil, albeit from different geographic regions of Taiwan, shared indistinguishable PFGE patterns and belonged to the same DSTs. Whether the original development of C. tropicalis exhibiting reduced susceptibility to fluconazole occurred in human hosts or in the environments needs further investigation. However, our observation is consistent with the idea that medically important drug resistant isolates, not only bacteria but also fungi, exist in the environments.
Previous study has shown that in human immunodeficiency virus infected patients, C. albicans and other yeast species were cross-resistant to medical and agricultural azole drugs [39]. However, no C. tropicalis was recovered in that study. The observation that isolates with reduced fluconazole susceptibility were also more resistant than the susceptible ones to fluquinco-nazole and tridimenol but not to pencofluquinco-nazole and tebucofluquinco-nazole (Table 2) may explain why the later two but not the former two are still used in agriculture in Taiwan. According to the data of ‘‘Domestic Manufacturers Production & Sale of Pesticides ‘‘ published by the Taiwan Crop Protection Industry Association, there is an increased amount of azole-type compounds used in the agriculture in Taiwan, from approximately 100 tons in 2005 to 145 tons in 2009 (about 45% increase). Since both efficient reproduction and spreading of resistant fungi in the environment are to be anticipated, we are confronted with the major challenge of drug-resistance development in environmental setting on a global scale. Hence, to reduce and prevent patients from acquiring azole-resistant C. tropicalis and other fungal species, it is advisable to take precaution against unnecessary use of azoles in not only clinical but also agricultural settings. The mechanisms contribut-ing to the reduced susceptibility to azole drugs are under investigation. Further study should be performed to compare C. tropicalis isolates recovered from soils in the fields as well as those from farmers for genetic relatedness and susceptibility to various antifungal drugs.
Materials and Methods Strain isolation
To isolate yeasts from the soil, we transferred approximately one gram of soil from each sample into a tube containing nine ml of sterile water and vortex-mixed. Diluted 0.2 ml of suspensions were plated onto acidified YMA (1% glucose, 0.5% peptone, 0.3% yeast extract, 0.3% malt extract, 1.5% agar, pH3.5) or Dichloran Rose Bengal Chloramphenicol agar (Merck, Darmstadt, Ger-many). After incubation at 24uC for 3 days, three (if present) representative colonies of each morphotype were picked for subsequent workup. All yeast isolates were identified by the sequences of the D1/D2 domain of the LSU rRNA genes as described by Lee et al. [40].
Drug susceptibility testing
The MICs of amphotericin B (0.0313–16 mg/L), fluconazole (0.125–64 mg/L), and voriconazole (0.0156–8 mg/L) were deter-mined by the same in vitro antifungal susceptibility testing established in our laboratory [41,42] according to the guidelines of M27-A3 recommended by the Clinical and Laboratory Standards Institute [43]. RPMI medium 1640 (31800-022, Gibco BRL) was used for the dilution and growth of the yeast culture. Strains from American Type Culture Collection (ATCC),
including C. albicans (ATCC 90028), C. krusei (ATCC 6258), and C. parapsilosis (ATCC 22019), were used as the standard controls. Growth of each isolate was measured by the Biotrak II plate spectrophotometric reader (Amersham Biosciences, Biochrom Ltd., Cambridge England) after incubation at 35uC for 24 h and 48 h.
The MIC of amphotericin B was defined as the minimum inhibitory concentrations of the drug capable of inhibiting cell growth. For susceptibility to amphotericin B, isolates with MIC§2 mg/L were considered to be resistant and those with MIC,––1 mg/L were susceptible. The MIC for fluconazole and voriconazole was defined as the lowest concentration capable of reducing the turbidity of cells by more than 50%. Isolates with fluconazole MICs$64 mg/L were considered as to have reduced susceptibility and #8 mg/L susceptible. The isolates with MICs in the range of 16–32 mg/L were referred to as susceptible-dose dependent (SDD). Isolates with voriconazole MICs$4 mg/L were considered as with reduced susceptibility and #1 mg/L susceptible. The isolates with MIC 2 mg/L were referred to as SDD.
Pulsed field gel electrophoresis (PFGE)
A total of 18 isolates were examined by PFGE, including 11 soil and 7 human isolates. The soil isolates comprised 6 (NHUE10, 28, 33, 40, 48, and 56) exhibiting fluconazole reduced susceptibility and 5 best matched susceptible ones isolated either from the same sample or from the same type of soil in the same area, NHUE11, 27, 36, 42, and 52 (vs. NHUE10, 28, 33, 40, and 48, respectively). The 7 human isolates, YM990275, 490, 537, 579, 592, 649, and 659, all exhibited reduced susceptibility to fluconazole. Pulsed field gel electrophoresis (PFGE) was performed based on our previous report [44] with some modifications. Briefly, organisms were grown on sabouraud dextrose agar (SDA) plate at 30uC overnight, after which the organisms were embedded in 1% agarose gel (SeaKem Gold agarose, FMC BioProducts). The plugs were subjected to cell lysis by lyticase at 37uC for 2 h, and then treated with proteinase K in 50 mM EDTA buffer at 50uC for 16 h. The genomic DNA was digested with 4 units of BssHII at 37uC for 16 h. Electrophoresis was then performed with a CHEF MAPPER system (Bio-Rad, Hercules, Calif., USA) in 0.56TBE buffer at 6 V/cm at 14uC with alternating pulses at an angle of 120 degrees in a 5–50 s pulse-time gradient for 20 h. A ladder of Salmonella choleraesuis subsp. choleraesuis (Smith) Weldin serotype Braenderup (ATCC BAA664) was used as the molecular weight marker. The PFGE pattern was analyzed by BioNumerics software (Version 4.5, Applied Maths, Kortrijk, Belgium). The position tolerance was set at 1% and optimization was set at 1%. The Dice coefficient was used to analyze the similarities of the band patterns. The unweighted pair group method using arithmetic averages (UPGMA) was used for cluster analysis.
Multilocus sequence typing
A total of 18 isolates exhibiting reduced susceptibility to fluconazole and 5 best matched susceptible ones (either from the same sample or from the same type of soil in the same area), NHUE11, 27, 36, 42, and 52 (vs. NHUE10, 28, 33, 40, and 48, respectively) as well as NHUE 22 with MIC of fluconazole 8 mg/L, were analyzed. Based on our previous report [19], the DNA fragments of six genes: ICL1, MDR1, SAPT2, SAPT4, XYR1, and ZWF1a were sequenced for the analyses. The resulted sequences were aligned with BioNumerics 3.0 (Applied Maths, Kortrijk, Belgium) and compared with those in the database of C. tropicalis (http://pubmlst.org/website) to obtain the identifiers (DST).
Growth curves
The Bioscreen C analyzer (Oy Growth Curves AB Ltd, Helsinki, Finland) was used to re-assess the growth of different isolates. Approximately 16104cells in 200ml of RPMI medium 1640 (Gibco BRL31800-022) were grown in the absence or in the presence of different drugs, including fluconazole, fluquincona-zole, penconafluquincona-zole, tebuconafluquincona-zole, and tridimenol, at 35uC. According to the record from the Taiwan Corp Protection Industry Association, penconazole and tebuconazole but not fluquinconazole and tridimenol were currently in use for agriculture in Taiwan. The growths of cells were determined every 15 minutes. Due to the solubility of fluquinconazole, the highest concentration of this drug in the present study was 16 mg/L instead of 64 mg/L. The experiments were repeated twice with similar results.
Acknowledgments
We would like to thank Pfizer for supplying fluconazole and Mission Biotech for performing sequencing. We would like to express our gratitude toward Dr. Tsung-Chain Chang for providing soil samples from petroleum-contaminated areas. We would like to thank Drs. Ferric Fang and Lawrence C. McDonald for their helpful suggestions on the manuscript.
Author Contributions
Conceived and designed the experiments: YLY HJL. Performed the experiments: CCL TPC HTC CWH PCC. Analyzed the data: CCL TPC YLY TLL CFL HJL. Wrote the paper: YLY TLL HJL.
References
1. Iatta R, Caggiano G, Cuna T, Montagna MT (2011) Antifungal Susceptibility Testing of a 10-Year Collection of Candida spp. Isolated from Patients with Candidemia. J Chemother 23: 92–96.
2. Pfaller MA, Diekema DJ (2007) Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev 20: 133–163.
3. Pfaller MA, Diekema DJ, Mendez M, Kibbler C, Erzsebet P, et al. (2006) Candida guilliermondii , an opportunistic fungal pathogen with decreased susceptibility to fluconazole: geographic and temporal trends from the ARTEMIS DISK antifungal surveillance program. J Clin Microbiol 44: 3551–3556.
4. Warnock DW (2007) Trends in the epidemiology of invasive fungal infections. Nippon Ishinkin Gakkai Zasshi 48: 1–12.
5. Yang YL, Cheng MF, Wang CW, Wang AH, Cheng WT, et al. (2010) The distribution of species and susceptibility of amphotericin B and fluconazole of yeast pathogens isolated from sterile sites in Taiwan. Med Mycol 48: 328–334. 6. Bard M, Bruner DA, Pierson CA, Lees ND, Biermann B, et al. (1996) Cloning and characterization of ERG25 , the Saccharomyces cerevisiae gene encoding C-4 sterol methyl oxidase. Proc Natl Acad Sci U S A 93: 186–190.
7. Beck-Sague C, Jarvis WR (1993) Secular trends in the epidemiology of nosocomial fungal infections in the United States, 1980–1990. National Nosocomial Infections Surveillance System. J Infect Dis 167: 1247–1251. 8. Cheng MF, Yu KW, Tang RB, Fan YH, Yang YL, et al. (2004) Distribution and
antifungal susceptibility of Candida species causing candidemia from 1996 to 1999. Diagn Microbiol Infect Dis 48: 33–37.
9. Pfaller MA, Boyken L, Hollis RJ, Kroeger J, Messer SA, et al. (2007) In Vitro Susceptibility of Invasive Isolates of Candida spp. to Anidulafungin, Caspofungin, and Micafungin: Six Years of Global Surveillance. J Clin Microbiol 46: 150–156.
10. Pfaller MA, Jones RN, Doern GV, Sader HS, Messer SA, et al. (2000) Bloodstream infections due to Candida species: SENTRY antimicrobial surveillance program in North America and Latin America, 1997–1998. Antimicrob Agents Chemother 44: 747–751.
11. Pfaller MA, Diekema DJ (2010) Epidemiology of invasive mycoses in North America. Crit Rev Microbiol 36: 1–53.
12. Pfaller MA, Moet GJ, Messer SA, Jones RN, Castanheira M (2011) Geographic variations in species distribution and echinocandin and azole antifungal resistance rates among Candida bloodstream infection isolates: report from the SENTRY Antimicrobial Surveillance Program (2008 to 2009). J Clin Microbiol 49: 396–399.
13. Ann Chai LY, Denning DW, Warn P (2010) Candida tropicalis in human disease. Crit Rev Microbiol 36: 282–298.
14. Xess I, Jain N, Hasan F, Mandal P, Banerjee U (2007) Epidemiology of candidemia in a tertiary care centre of north India: 5-year study. Infection 35: 256–259.
15. Yang YL, Ho YA, Cheng HH, Ho M, Lo HJ (2004) Susceptibilities of Candida species to amphotericin B and fluconazole: the emergence of fluconazole resistance in Candida tropicalis. Infect Control Hosp Epidemiol 25: 60–64. 16. Lee MK, Williams LE, Warnock DW, Arthington-Skaggs BA (2004) Drug
resistance genes and trailing growth in Candida albicans isolates. J Antimicrob Chemother 53: 217–224.
17. Arthington-Skaggs BA, Lee-Yang W, Ciblak MA, Frade JP, Brandt ME, et al. (2002) Comparison of visual and spectrophotometric methods of broth microdilution MIC end point determination and evaluation of a sterol quantitation method for in vitro susceptibility testing of fluconazole and itraconazole against trailing and nontrailing Candida isolates. Antimicrob Agents Chemother 46: 2477–2481.
18. Lo HJ, Ho AH, Ho M (2001) Factors accounting for mis-identification of Candida species. J Microbiol Immunol Infect 34: 171–177.
19. Chou HH, Lo HJ, Chen KW, Liao MH, Li SY (2007) Multilocus sequence typing of Candida tropicalis shows clonal cluster enriched in isolates with resistance or trailing growth of fluconazole. Diagn Microbiol Infect Dis 58: 427–433.
20. Wang JS, Li SY, Yang YL, Chou HH, Lo HJ (2007) Association between fluconazole susceptibility and genetic relatedness among Candida tropicalis isolates in Taiwan. J Med Microbiol 56: 650–653.
21. Li SY, Yang YL, Lin YH, Ko HC, Wang AH, et al. (2009) Two Closely Related Fluconazole-Resistant Candida tropicalis Clones Circulating in Taiwan from 1999 to 2006. Microb Drug Resist 15: 205–210.
22. Vogel C, Rogerson A, Schatz S, Laubach H, Tallman A, et al. (2007) Prevalence of yeasts in beach sand at three bathing beaches in South Florida. Water Res 41: 1915–1920.
23. Lord AT, Mohandas K, Somanath S, Ambu S (2010) Multidrug resistant yeasts in synanthropic wild birds. Ann Clin Microbiol Antimicrob 9: 11.
24. Hung CC, Yang YL, Lauderdale TL, McDonald LC, Hsiao CF, et al. (2005) Colonization of human immunodeficiency virus-infected outpatients in Taiwan with Candida species. J Clin Microbiol 43: 1600–1603.
25. Prasad KN, Agarwal J, Dixit AK, Tiwari DP, Dhole TN, et al. (1999) Role of yeasts as nosocomial pathogens & their susceptibility to fluconazole & amphotericin B. The Indian Journal of Medical Research 110: 11–17. 26. Barchiesi F, Calabrese D, Sanglard D, Falconi DF, Caselli F, et al. (2000)
Experimental induction of fluconazole resistance in Candida tropicalis ATCC 750. Antimicrob Agents Chemother 44: 1578–1584.
27. Calvet HM, Yeaman MR, Filler SG (1997) Reversible fluconazole resistance in Candida albicans : a potential in vitro model. Antimicrob Agents Chemother 41: 535–539.
28. Cowen LE (2001) Predicting the emergence of resistance to antifungal drugs. FEMS Microbiol Lett 204: 1–7.
29. Cowen LE, Sanglard D, Calabrese D, Sirjusingh C, Anderson JB, et al. (2000) Evolution of drug resistance in experimental populations of Candida albicans. J Bacteriol 182: 1515–1522.
30. Snelders E, Huis In ‘t Veld RA, Rijs AJ, Kema GH, Melchers WJ, et al. (2009) Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl Environ Microbiol 75: 4053–4057.
31. Snelders E, van der Lee HA, Kuijpers J, Rijs AJ, Varga J, et al. (2008) Emergence of azole resistance in Aspergillus fumigatus and spread of a single resistance mechanism. PLoS Med 5: e219.
32. Verweij PE, Snelders E, Kema GH, Mellado E, Melchers WJ (2009) Azole resistance in Aspergillus fumigatus: a side-effect of environmental fungicide use? Lancet Infect Dis 9: 789–795.
33. Chen FJ, Lo HJ (2003) Molecular mechanisms of fluoroquinolone resistance. J Microbiol Immunol Infect 36: 1–9.
34. McDonald LC, Chen FJ, Lo HJ, Yin HC, Lu PL, et al. (2001) Emergence of reduced susceptibility and resistance to fluoroquinolones in Escherichia coli in Taiwan and contributions of distinct selective pressures. Antimicrob Agents Chemother 45: 3084–3091.
35. Yang YL, Lauderdale TL, Lo HJ (2004) Molecular mechanisms of fluoroquin-olone resistance in Klebsiella. Curr Drug Targets Infect Disord 4: 295–302. 36. Akins RA (2005) An update on antifungal targets and mechanisms of resistance
in Candida albicans. Med Mycol 43: 285–318.
37. White TC, Marr KA, Bowden RA (1998) Clinical, cellular, and molecular factors that contribute to antifungal drug resistance. Clin Microbiol Rev 11: 382–402.
38. Yang YL, Lo HJ (2001) Mechanisms of antifungal agent resistance. J Microbiol Immunol Infect 34: 79–86.
39. Muller FM, Staudigel A, Salvenmoser S, Tredup A, Miltenberger R, et al. (2007) Cross-resistance to medical and agricultural azole drugs in yeasts from the oropharynx of human immunodeficiency virus patients and from environmental Bavarian vine grapes. Antimicrob Agents Chemother 51: 3014–3016. 40. Lee CF, Yao CH, Liu YR, Young SS, Chang KS (2009) Kazachstania wufongensis
sp. nov., an ascosporogenous yeast isolated from soil in Taiwan. Antonie Van Leeuwenhoek 95: 335–341.
41. Yang YL, Cheng HH, Lo HJ (2004) In vitro activity of voriconazole against Candida species isolated in Taiwan. Int J Antimicrob Agents 24: 294–296.
42. Yang YL, Wang AH, Wang CW, Cheng WT, Li SY, et al. (2008) Susceptibilities to amphotericin B and fluconazole of Candida species in TSARY 2006. Diagn Microbiol Infect Dis 61: 175–180.
43. Clinical Laboratory Standards Institute CLSI (2008) Reference method for broth dilution antifungal susceptibility testing of yeasts; approved standard-third edition. CLSI document M27-A3. Wayne, PA.
44. Chen KW, Lo HJ, Lin YH, Li SY (2005) Comparison of four molecular typing methods to assess genetic relatedness of Candida albicans clinical isolates in Taiwan. J Med Microbiol 54: 249–258.