HMJ-53A加速Kv通道慢性鈍化之機轉; Mechanisms for the acceleration of Kv channel slow inactivation by HMJ-53A
99
0
0
全文
(2) 誌謝辭 回想兩年前,剛剛進入研究所的我,對於研究有著很大的抱負,但 卻不知道如何做起時,很幸運的進入到充滿歡樂氣氛的梁育民老師實 驗室,並成為老師在台灣任教的第一位研究生。 兩年前,對電生理一知半解的我,一開始很惶恐但又很期待,又擔 心自己做不好時,而我的指導教授-梁育民老師,很有耐心及細心的 指導我,是一位很肯花時間在學生身上的好老師,使我在研究所中接 受到一個完整的訓練過程。並從他的身上感受到研究及教學的熱誠, 而讓我覺得做學問及研究是如此的有趣,對於思考問題更加縝密及有 邏輯性,並影響我對於整個人生價值觀有著不同的想法,也因此讓我 有繼續深造的動力;而對這位亦師亦友的梁老師之感謝,是無法用三 言兩語形容-但真得很謝謝您,這段時間的用心栽培及指導。 感謝我的口試委員鄭瑞棠老師及鄭志鴻老師在口試過程中的專業 指導,及建議我往後實驗方向,並引導我對於研究思考範圍更加廣闊, 讓我受益良多。 也要謝謝這兩年來醫研所及生理科所有老師的指導,讓我對於口頭 報告、思考問題及研究問題的批判,有很大的進步。另外,還要謝謝 惠華姐在生理實驗課上的各項幫忙,也要謝謝醫研所麗如姐,協助處 理我們研究生各項業務。.
(3) 在實驗室一開始起步時,因林靜茹及許明志老師實驗室的全體成員 幫忙,才讓我們能順利運作,謝謝你們。也要謝謝醫研所二年級的全 體同學,大家的互相扶持及幫忙,才能開心及順利完成這兩年學業。 並要感謝實驗室所有成員,因大家相處融洽及互相幫忙,才讓整個實 驗室的運作順利。 再來我要謝謝我的家人及未來的家人,特別是我的母親,在背後一 直默默的支持我,不讓我有任何顧慮的專心研究,要不是你們的支持, 我也不可能順利畢業,謝謝你們! 這段研究生生涯,過程中雖有苦有笑有甜,但現在回想起來,全 都是美妙的回憶也嘗到甜美的果實,不會忘記這裡的點點滴滴,我將 會帶著這些果實,繼續充實我自己,並往上邁進。最後,再次感謝這 一路所有幫助過也曾關心過我的人,謝謝你們。. 研究生. 趙家佳 謹識. 民國九十七年六月二十三日 於台中.
(4) 總目錄 頁次 總目錄 ..................................................................................................... 1. 中文摘要 ................................................................................................. 8. 英文摘要 ................................................................................................. 9. 第一章:前言 ....................................................................................... 11. 第一節:鉀離子通道的種類及特性................................................ 11 1. 鈣離子活化鉀離子通道[ Ca2+-activated K+ (KCa) channels ] .. 11 2. 漏鉀通道[ Leak K+ channel ] .................................................... 12 3. 內流型鉀離子通道[ Inward rectifier K+ (Kir) channels ] ........ 12 4. ATP 敏 感 性 型 鉀 離 子 通 道 [ Adenosine 5′- triphosphate sensitive K+ (KATP) channels ] ................................................... 13 5. 電壓調控鉀離子通道[ Voltage - gated K+ (Kv) channels ] ..... 13. 1.
(5) 第二節:電壓調控鉀離子通道[Voltage-gated K+ (Kv) channels] 結構 .................................................................................... 14. 第三節:電壓調控鉀離子通道之鈍化............................................ 20. 第四節:電壓調控鉀離子通道中 N 型鈍化................................... 20. 第五節:電壓調控鉀離子通道中 C 型鈍化 ................................... 22. 第二章:研究方法 ............................................................................... 28 第一節:細胞培養 ............................................................................ 28. 第二節:藥劑 .................................................................................... 29. 第三節:轉殖過程 ............................................................................ 31. 第四節:電生理(Electrophysiology)紀錄方法 .......................... 32 一、溶液準備 ................................................................................ 32 二、電生理記錄過程 .................................................................... 32 三、濃度反應曲線 ........................................................................ 33. 2.
(6) 四、活化曲線圖及鈍化曲線圖 .................................................... 34 五、穩定狀態之鈍化( Steady-state inactivation )曲線 ................ 34. 第五節:電生理( Electrophysiology)記錄流程 .............................. 35. 第六節:統計方法 ............................................................................ 35. 第三章:研究結果 ............................................................................... 36 第一節:HMJ-53A 在 N2A 細胞外側上作用,並加速慢性鈍化 現象 .................................................................................... 36. 第二節:HMJ-53A 作用,是否與蛋白質激酶之磷酸化有關呢? .............................................................................................. 38. 第三節:HMJ-53A 作用在 Kv 通道之關閉階段 ........................... 39. 第四節:HMJ-53A 並非為通道外孔之直接阻斷劑 ...................... 39. 第五節:HMJ-53A 作用在 Kv 通道之鈍化閥門 ........................... 41. 第六節:HMJ-53A 不會影響 Kv 通道上之活化閥門 ................... 43. 3.
(7) 第七節:HMJ 化合物其結構與產生加速鈍化閥門關閉之相關性 ............................................................................................ 44. 第四章:討論 ....................................................................................... 45. 第五章:結果圖表與說明 ................................................................... 51. 第六章:参考文獻 ............................................................................... 75. 附錄一:SCI 期刊之文章發表............................................................ 86. 附錄二:獎狀 ....................................................................................... 87. 4.
(8) 圖表目錄 頁次 圖 1:Kv 通道上之單個 α-膜蛋白次單元 ( α- subunits ) ................. 17. 圖 2:Kv 通道上,單個 α- subunit 之六條穿膜蛋白( S1-S6 ) ......... 18. 圖 3:Kv 通道之選擇性過濾器( selectivity filter ) ............................ 19. 圖 4:鉀離子通道上之 N 型鈍化 ....................................................... 22. 圖 5:鉀離子通道上之 C 型鈍化 ....................................................... 23. 圖 6:Kv1.2 通道上,Glu370 與 Asp379 會相互牽引, 產生 C 型鈍化 .......................................................................... 25. 圖 7:Kv1.2 通道上,產生 C 型鈍化過程 ........................................ 26. 圖 A:HMJ-53A 結構式與合成程序.................................................. 29 5.
(9) 圖 B:HMJ 系列之藥物結構式 .......................................................... 30. 結果圖表與說明 ............................................................................... 51 Fig. 1:HMJ-53A 加速 N2A 細胞 Kv 電流慢性鈍化現象 ............. 51. Fig. 2:HMJ-53A 抑制 Kv 電流濃度依賴曲線 ............................... 53. Fig. 3:HMJ-53A 在轉殖 Kv2.1 後之 N2A 細胞上,加速 Kv 電流 慢性鈍化現象 ......................................................................... 55. Fig. 4:從 N2A 細胞內加入 HMJ-53A 並不影響 Kv 電流之慢性鈍 化 ............................................................................................. 57. Fig. 5:HMJ-53A 作用於 N2A 細胞之 Kv 通道之關閉階段 ......... 59. Fig. 6:HMJ-53A 加劇電流之衰退,沒有電壓依賴性 .................. 61. Fig. 7:HMJ-53A 阻斷 Kv 電流之強度不受電壓與細胞內鉀離子濃 度影響 ....................................................................................... 63 6.
(10) Fig.8:TEA 對電流衰退之影響與 HMJ-53A 不同 ......................... 66. Fig. 9:HMJ-53A 使穩定狀態之鈍化曲線左移但不影響活化閥門 ................................................................................................. 68. Fig. 10:HMJ-53A 不影響 Kv 電流之恢復 ..................................... 71. Fig. 11:HMJ 衍生物抑制 Kv 電流之濃度依賴性曲線 ................. 73. 7.
(11) 中文摘要 在可興奮性細胞,例如,神經細胞和內分泌細胞,利用電壓調 控鉀離子(Kv)通道,使其膜電位於去極化後可復極化。某些Kv通道 在去極化之過程中,會表現慢性鈍化之現象,此鈍化作用會使Kv 通道產生某種形式的關閉,而其發生是因通道裡離子選擇性過濾器 壓縮。至今,尚未有一個針對此慢性鈍化之特異性的探測藥物。我 們發現一個新的藥物,HMJ-53A (30 μM)在小鼠神經瘤細胞株 (N2A) 上 , 有 選 擇 性 的 加 速 Kv 通 道 之 慢 性 鈍 化 ( 沒 加 與 有 加 HMJ-53A之衰退 τ,分別為1677 ± 120 ms 和 85.6 ± 7.7 ms)。 HMJ-53A把Kv通道穩定鈍化曲線左移12mV,且不影響其活化階段。 此外,我們在微細玻璃管電極內(細胞內)加入HMJ-53A,並不會 產生加速Kv通道之慢性鈍化現象;而從此數據顯示,HMJ-53A作用 在細胞外而非細胞內。HMJ-53A阻斷Kv通道上鉀離子外流,而所測 得之電流,不需要Kv通道開啟。此外,HMJ-53A之阻斷Kv通道電 流及鈍化之時間常數,不會受到去極化程度及細胞內鉀離子濃度的 影響。所以,HMJ-53A不會直接堵塞Kv通道之孔洞,而是讓圍著通 道孔之選擇性過濾器加快壓縮。綜合以上的結果可以得知, HMJ-53A可有選擇性的加速Kv通道之慢性鈍化,可作為探測Kv通 道鈍化閥門之對抗藥。 8.
(12) Abstract Voltage-gated K+ (Kv) channels are important in repolarization of excitable cells such as neurons and endocrine cells. Kv channel gating exhibits slow inactivation (slow current decay) during continuous depolarization. The molecular mechanism involved in such slow inactivation is not completely understood, but evidence has suggested that it involves a restriction of the outer channel pore surrounding the selectivity filter. Pharmacological tools probing this slow inactivation process are scarce. In this work we reported that bath application of HMJ-53A (30 μM), a novel compound, could drastically speed up the slow decay (decay τ=1677±120 ms and 85.6 ±7.7 ms, respectively, in the absence and presence of HMJ-53A) of Kv currents in neuroblastoma N2A cells. HMJ-53A also significantly left-shifted the steady-state inactivation curve by 12 mV. HMJ-53A, however, did not affect voltage-dependence of activation and the kinetics of channel activation. Intracellular application of this drug through patch pipette dialysis was ineffective at all in accelerating the slow current decay, suggesting that HMJ-53A acted extracellularly. Blockade of currents by HMJ-53A did not require an open state of channels. In addition, the 9.
(13) inactivation time constants and percentage block of Kv currents in the presence of HMJ-53A were independent of the (i) degree of depolarization and (ii) intracellular K+ concentration. Therefore, this drug did not appear to directly occlude the outer channel pore during stimulation (depolarization). Taken together, our results suggest that HMJ-53A selectively affected (accelerated) the slow inactivation gating process of Kv channels, and could thus be a selective and novel probe for the inactivation gate.. 10.
(14) 第一章:前言 在細胞膜上有許多的離子通道,扮演各種不同功能,而控制離子 通道的開關因子包括:神經傳送素、荷爾蒙、膜電位、壓力、光及 代謝產物等。可興奮性細胞受到刺激時,鈉與鉀離子通道隨即打開, 例如:神經細胞因受到藥物及細胞膜電位改變等刺激;而上述因素 導致,鈉離子通道開啟,鈉離子進入細胞膜內,造成膜電位去極化 (depolarization) ( Berne et al., 1998 )。接著鈉離子通道開始關閉,鈉 離子無法進入細胞內,此時,鉀離子通道開啟,鉀離子開始往細胞 外移動,使細胞膜漸漸恢復其負電性,此稱為復極化 (repolarization),此時膜電位稍微超過靜止膜電位,而形成過極化 (hyperpolarization),進而漸漸達到鉀離子通道平衡電位,鉀離子通 道關閉,回到靜止膜電位;這就是在興奮性細胞上的電性活動階 段。. 第一節:鉀離子通道的種類及特性 目前已知鉀離子通道的種類很多,最多被研究者包括下列幾種 (Hille, 2001): 1. 鈣 離 子 活 化 鉀 離 子 通 道 channels ]: 11. [ Ca2+-activated K+ (KCa).
(15) 當細胞膜電位發生改變,細胞內鈣離子濃度增加 1–10 μM, 並使細胞呈現去極化現象,此時鉀離子通道開啟,鉀離子從 細胞內流出;再依傳導性的大小,可分為 big K(Ca) 【BK】、 intermediate K(Ca)【IK】及 small K(Ca)【SK】通道。而目前 已 知 , 此 通 道 存 在 於神 經、心臟及骨骼肌肉細胞…等 中 (Poolos and Johnston, 1999; Wang et al., 2008)。 2. 漏鉀通道 [ Leak K+ channel ]: 此通道結構是由 4 個穿膜片段 (S1-S4) 連接及 2 個 P-loop 所構成的。而其一直呈現開啟狀態,主要是受膜電位及細胞 內外鉀離子濃度影響。當漏鉀離子通道開啟時,為了要抗衡 化學梯度,而在此通道中,必須產生膜內相對於膜外較負的 電位差,而膜內的負電荷會拉住鉀離子,使鉀離子無法往膜 外流動,使鉀電流動達到平衡。所以,此通道的主要功能是 維持靜止膜電位 (Goldstein et al., 1996)。而此通道會受到細胞 外 pH 值變化及麻醉藥(例如:Bupivacaine)影響 (Bayliss et al., 2001; Kindler et al., 1999)。 3. 內流型鉀離子通道 [ Inward rectifier K+ (Kir) channels ]: 調控主要是受膜電位的改變而開啟;當細胞膜電位低於平 衡電位時,會使此通道開啟,並使鉀離子往細胞內流入;而 12.
(16) 其特別的是,並不會因為隨著膜電位增加,而使鉀離子往細 胞外流動 (Hille, 2001);最主要調控此通道者,是靠著細胞內 的陽離子:Mg2+及 putrescine、spermidine、spermine,去阻止 鉀離子往細胞外移動 (Loussouarn et al., 2002)。Kir 通道多存 於心臟及神經細胞…等。 4. ATP 敏感性型鉀離子通道 [ Adenosine 5′- triphosphate sensitive K+ (KATP) channels ]: 結構是由Kir6.x(6.1或6.2)及sulfonylurea接受器(SUR1或 SUR2)所構成。此通道的開啟與關閉是受細胞內ATP和ADP 的濃度影響,也就是當細胞內[ATP]/[ADP]下降,會使此通道 開啟,而現今在神經保護機制及內分泌的調控…等,占有很重 要的角色。例如:當人體處於飽食狀態,胰島上的β-細胞內 [ATP]/[ADP]比例上升,讓此通道關閉,鉀離子不往細胞外流 動,使β-細胞去極化並興奮,再使Ca通道開啟,而使胰島素釋 放,進而降低血糖。目前已知此通道存在於神經、心臟及胰島 細胞…等中 (Leung et al., 2006; Sun et al., 2007);而現今對於 KATP 通 道 對 神 經 保 護 之 作 用 已 被 證 實 但 機 制 卻 不 清 楚 (Jahangir and Terzic, 2005; Yamada and Inagaki, 2005)。 5. 電壓調控鉀離子通道 [ Voltage - gated K+ (Kv) channels ]: 13.
(17) Kv 通道是鉀離子通道種類中,目前最多被廣泛研究的一種, 而其在興奮性細胞之動作電位中,占有重要的角色。 而 Kv 通道在動作電位中之復極化,控制鉀離子往細胞外流 動上扮演很重要的角色;尤其在興奮性細胞:內分泌、肌肉、 神經及心肌細胞…等。而另一方面,Kv 通道會受到閥門影響, 去控制其開關;當 Kv 通道開啟的過程稱為活化 (activation), 在通道開啟後隨之漸漸失去通透鉀離子能力稱為鈍化 (inactivation),而當復極化之後,Kv 通道會回到關閉的狀態, 此過程稱為去活化 (deactivation)。這就是整個 Kv 通道從開 啟到關閉的整個過程。. 第二節:電壓調控鉀離子通道[Voltage-gated K+ (Kv) channels]結構 Kv通道構造是由四個α-膜蛋白次單元 (α-subunits)(圖1)而環繞 著中心孔所構成,而每個α-subunit是由六條穿膜片段 (S1-S6)連接 而成(圖2) ,每個α-subunit包含C端 (C-terminus)和N端 (N-terminus), 皆位於細胞內側 (Choe et al., 1999)。 S4因帶有規則排列的七個正電鹼性胺基酸,可以感應電場的變化, 而S4最靠近外側的四個鹼性胺基酸為主要的感應電荷位置,而在通 14.
(18) 道關閉狀態時是位在近細胞膜內側內,當通道受電場變化而開啟時, 四個鹼性胺基酸是位在近細胞膜外側上。S4運動方式主要有兩種: 第一種:S4利用旋轉並往細胞外位移的方式運動。 第二種:S4位於通道蛋白質的外圍與細胞膜接觸,利用擺動的方 式移動。 2003年,Mackinnon團隊利用Aeropyrum pernix的嗜熱性古細菌 ( thermophilic archaebacterium ) 分離出鉀離子通道蛋白KvAP,而其 胺基酸序列與真核細胞的Kv通道蛋白非常相近,並以結晶X-ray繞 射技術做出KvAP電壓關閉性的鉀離子通道結晶結構,其感應電荷 (gating charges) 位 置 在 離 子 通 道 最 外 圍 , 是 位 在 S3b-S4 、 helix-turn-helix結構中,當細胞去極化時,整個電壓感測器擺動至細 胞外,進而牽動孔洞上S6使通道開啟,而此種模式稱之為paddle model (Jiang et al., 2003)。 鉀 離 子 通 道 上 S5 和 S6 間 的 P-loop 是 由 -TXXTXGYGD( -thr-X-X-thr-X-gly-tyr-gly-glu- )此特徵序列組成,而由膜外側伸入 膜內後,做一個轉折再往膜外側伸出,形成一個圈環且是整個通道 最狹窄的孔洞。Kv通道中,其中-TVGYG- (-thr-V-gly-tyr-gly-)構成 離子選擇最關鍵的部位稱之為選擇性過濾器 (selectivity filter)(圖3) (Choe, 2002; Yellen, 2002);因為選擇性過濾器相當狹窄,依凡得瓦 15.
(19) 力定律 (van der Waal's law),鉀離子在通過時必須脫去外側層水合 分 子 (hydration waters) , 在 選 擇 性 過 濾 通 道 上 羰 基 的 氧 原 子 (carbonyl oxygens)則取代鉀離子外側層水分子,與鉀離子結合。由 於鉀離子的體積大小能精確的配合選擇性過濾通道大小與羰基的 氧原子皆都可碰到,而能量得失取之平衡;雖鈉離子的體積較鉀離 子小卻無法精確的配合此過濾通道大小,無法平衡,所以,這也就 是鉀離子通道能選擇性的只供鉀離子通過之原因 (Choe et al., 1999; Doyle et al., 1998)。 Kv通道的整個結構:由S1-S6六個穿膜片段 (six transmembrane domain, 6TM),構成一個次單位,其間S5與S6連接一條P-loop (P) 組成,稱為6TM/P (Hille, 2001)。 Kv通道的功能:可將膜電位拉回 至靜止膜電位,而使得動作電位在適當的時間內形成,且可結束一 連串的放電,並在重複性的極化模式中、調控動作電位之輪廓與間 隔時間 (Nerbonne, 2000)。. 16.
(20) 圖 1:Kv 通道上之單個 α-膜蛋白次單元 (α- subunit)立體結構: 由六個( S1-S6)穿膜蛋白連接而成一個 α- subunit,而一個 Kv 通道由四個 α- subunit 所組成,其 N 端與 C 端皆位於細 胞內側。modify (Choe et al., 1999)。. 17.
(21) 圖 2:Kv 通道上,每個 α- subunit 是由六條穿膜蛋白( S1-S6 )連接 而成,包含 C-terminus 和 N-terminus 皆位於細胞內側, S4 為電壓感應器。. 18.
(22) 圖 3:Kv 通道之選擇性過濾器( selectivity filter ), 鉀離子經過此過濾器時,須經過脫水的過程後,與過 濾器中的氧原子結合後,往細胞膜外流出 (Choe, 2002)。. 19.
(23) 第三節:電壓調控鉀離子通道之鈍化 (inactivation) 當細胞膜去極化時,S4能感測電場變化而使S6移動,進而使得鉀 離子通道活化閥門 (activation gate)開啟,而允許細胞內的鉀離子往 細胞外流出 (Choe, 2002; Yellen, 2002)。而當活化閥門開啟後,鉀離 子通道接著走向鈍化 (inactivation)階段,而此階段必須是通道開啟 後產生。復極化後,通道走向去活化 (deactivation)階段,使通道關 閉,而鈍化之通道亦慢慢回復 (recovery) (Hille, 2001)。 在電壓調控鉀離子通道上,鈍化主要分成快速及慢速兩大類: 第一類:N型鈍化 ( N - type inactivation ),因發生鈍化速度快約為 100至200ms,又稱為快速型鈍化 ( fast inactivation ),見於 Kv1.4 、 Kv3.1 、 Kv3.4 和 Kv4.2 通 道 上 (Kukuljan et al., 1995)。 第二類:C型鈍化 ( C - type inactivation ),大部分Kv通道上皆可見, 因其鈍化速度長達一秒至幾秒鐘,速度很慢,又稱為慢速 型鈍化 ( slow inactivation ) (Kurata and Fedida, 2006),可見 於Kv1.5和 Kv2.1通道上 (Andalib et al., 2004; Kurata et al., 2001)。. 第四節:電壓調控鉀離子通道中N型鈍化 20.
(24) 在Kv通道中,N型鈍化發生在六個穿膜片段所組成的每個次單位 上的N端,而此端是位於細胞內連接著20個胺基酸所組成的球 (ball) 與21至80號胺基酸所形成的鏈 (chain),稱之為“ball and chain”。當 細胞膜因膜電位去極化的刺激,使Kv通道活化時,導致通道蛋白結 構變化,閥門開啟,細胞膜內之通道孔洞內露出鈍化球接合的位置, 此時Kv通道上N端的“ball and chain”由下往上堵住孔洞而形成快速 鈍化,並防止鉀離子通過此孔洞 (Gulbis et al., 2000),再當細胞膜 復極化後,因電場改變而產生回復作用,並使球離開孔洞,再產生 去活化,使通道回到休息的狀態(圖4)。 若快速給予過極化時,球無法及時在通道關閉前掉落,此時在通 道內會產生“卡住” (trapped)之現象 (Holmgren et al., 1996)。若將鉀 離子通道上的N端(20個胺基酸,ball)除掉,可去除N-型鈍化,另 外,假若切除N端後,合成20個胺基酸序列於細胞內,發現可以回 復N型鈍化反應,且發生速率與球之濃度成正比的現象 (Zagotta et al., 1990)。 所以,Kv通道內的四個鈍化球受電位改變移動後在細胞內卡上孔 洞,形成N型鈍化,而這四個球皆有相同機率產生這樣的結果,只 要其中一個球即可,速度很快的達成N型鈍化,所以,四個鈍化球 是一個鈍化球達成N型鈍化的四倍速率 (MacKinnon et al., 1993)。 21.
(25) 而TEA (tetraethylammonium chloride)已是現今普遍用來探討阻斷N 型鈍化的鉀離子阻斷劑,其阻斷方式乃是經由細胞內往上堵住通道 內孔,而使鉀離子通道上之N端之“ball and chain”無法堵住內孔,使 N型鈍化無法達成 (Baukrowitz and Yellen, 1996; Choi et al., 1991; Choi et al., 1993)。. 圖 4 :N-type inactivation 鉀離子通道上之 N 型鈍化. 第五節:電壓調控鉀離子通道中 C 型鈍化 慢性鈍化幾乎發生在所有的Kv通道上,不論有無N型鈍化,而此 種慢性鈍化現象常常會持續幾秒鐘,且發生在通道持續不斷的去極 化期間 (Kurata and Fedida, 2006)。慢性鈍化現象又稱為C型鈍化 22.
(26) (C-type inactivation)且發生在許多不同種類的Kv通道上,例如: Kv1.5和Kv2.1通道 (Andalib et al., 2004; Kurata et al., 2001)。 現今來說,這種複雜的C型鈍化機制,並不是完全清楚了解,但 現今有研究顯示:C型鈍化現象是通道外孔的選擇性過濾器產生倒 塌或壓縮而造成結構改變 (Kurata and Fedida, 2006)。而至今已知其 作用機制:當細胞膜因膜電位去極化的刺激,使Kv通道活化時,導 致通道蛋白結構變化,閥門開啟,進而使通道外孔的選擇性過濾器 產生收縮及壓縮而造成結構不穩定,形成慢速鈍化,而細胞膜處於 復極化狀態後,因後電場改變而產生回復作用,並使通道孔洞內的 選擇性過濾器慢慢的往外回復,通道繼而再產生去活化,使通道回 到休息的狀態,並準備好下一次的活化及放電(圖5)。. 圖5:C-type inactivation 鉀離子通道上之 C 型鈍化 23.
(27) 哪些胺基酸影響C型鈍化?最早是在Shaker 通道(Kv1家族)外 孔上的threonine(T)位置449的改變可以加快(T449E,T449A, T449K) 或減慢(T449Y, T449V)C型鈍化速度,也就是改變T449:arginine (R)、lysine(K)、alanine(A)或 glutamate(E)這些胺基酸, 會加快其速度;另一方面,改變為valine(V)或tyrosine(Y)會減 慢C型鈍化速度。若切斷N端只存有C端後,並不會停止其鈍化現象, 而定義為C型鈍化 (Hoshi et al., 1990; Lopez-Barneo et al., 1993)。 1998年,學者發現,在Kv1.3及1.5通道上,細胞外加入低濃度 (30-50 μM)鉀離子,而鉀離子與通道上的選擇性過濾器親和力增加, 並大於鈉離子,而抑制C型鈍化 (Kiss et al., 1998)。Shaker通道上, 增加細胞外鉀離子的濃度及加入TEA,皆會減慢C型鈍化的速度。 另 外 在 Kv1.3 中 , 發 現 細 胞 內 酪 胺 酸 磷 酸 化 (tyrosine kinase phosphorylation)可造成C型鈍化速度很輕微的加快,但作者卻無法 定義出磷酸化會影響C型鈍化的相關性或影響Kv1.3哪個胺基酸 (Bowlby et al., 1997)。 2007年有學者研究出:利用能夠穩定二級結構的氫鍵 (hydrogen bonds)去探討選擇性過濾器上的殘基與鄰近在通道外孔上的helix之 間的相關性及重要性,而去證實C型鈍化機制 (Cordero-Morales et al., 2007)。因此在Kv1.2上, 把Val370改變成Glu370(圖6),在活 24.
(28) 化Kv1.2後,選擇性過濾器上的Glu370可與Asp379作用更強之相互 牽引,使過濾器不穩定(或倒塌),隨之產生C型鈍化。當回復時, Glu370與Asp379會互相彈開,隨之去活化後,通道回到休息狀態 (Cordero-Morales et al., 2007)(圖7)。. 圖 6:在 Kv1.2 通道上,把原本 Val370 突變成 Glu370,發 現 Glu370 與 Asp379 會更強地相互牽引,產生 C 型鈍化 (Cordero-Morales et al., 2007)。. 25.
(29) 圖7:Kv1.2通道(A)通道處於休息狀態(B)選擇性過濾器 上的Glu370與Asp379(兩個紅點表示)會相互牽引(C)隨之 產生C型鈍化(D)此過程即是選擇性過濾器倒塌,而使鉀離 子無法順利通過此通道(E)回復,Glu370與Asp379互相彈開 (F)去活化後,通道回到休息狀態 (Cordero-Morales et al., 2007) 。. 26.
(30) 鉀離子通道的鈍化作用影響動作電位的輪廓及頻率,因此對於興 奮細胞的調節是相當重要,例如:神經細胞 ( Hille, 2001 )。而藥物 改變Kv通道的鈍化閥門也就是能改變神經細胞的興奮性,例如: 4-acetamido-4'-isothiocyano-2,2'-disulphonic. stilbene. (SITS) 和. N-bromoacetamide (NBA),已在腦垂下體細胞GH3中證實,能抑制 Kv通道的N型鈍化 (Oxford and Wagoner, 1989)。 然而,目前尚無專一及選擇性的藥物去作為探測藥物進而探討Kv 通道之C型鈍化。進而與侯曼貞老師合作新藥開發,篩選HMJ系列 合成物過程中,發現此系列合成物中之HMJ-53A能影響Kv通道,而 先前已發表其抗凝血作用藥效比Aspirin強,可以抑制花生四稀酸所 造成的血小板凝集疾病 (IC50:1.60 μM) ( Hour et al., 2000 ),這也 讓我們懷疑所產生的抗凝血作用,是否因為阻斷離子通道所產生的 呢? 所以,在此研究工作中,我們利用已知表達Kv通道的小鼠神經瘤 N2A( mouse neuroblastoma )細胞,並著手探討HMJ-53A是如何影響 Kv通道。結果發現此細胞上之Kv通道存有C型鈍化現象,且經由我 們強而有力的結果顯示,HMJ-53A作用在細胞外而非細胞內,並大 幅度的加速C型鈍化卻不影響Kv通道上的活化閥門。這些結果顯示 HMJ-53A能當作一個新穎的合成物,去探測C型鈍化閥門。 27.
(31) 第二章:研究方法 第一節:細胞培養 N2A (mouse neuroblastoma)為小鼠神經瘤細胞株,生長於37℃、5 % CO2 培 養 箱 , 培 養 基 為 Dulbemlo's modified Eagle's medium (DMEM) (Gibco)、10% fetal bovine serum (FBS) (Invitrogen, Carlsbad, CA) 及 1 % penicillin-streptomycin (100 units/ml, 100 μg/ml) (Invitrogen),為黏附型細胞,繼代培養方式:抽掉上清液,再用 Dulbemlo's Phosphate Buffered Saline(PBS) (Sigma-Aldrich)沖兩次後, 再加入1ml trypsin (0.25%, Invitrogen),等待一分半鐘,再加入培養 基6ml,然後放入離心管離心(900rpm、5分鐘)。之後,抽掉上清 液,再加入1ml培養基,混合均勻,再加入少許的混合液至已有培 養基的培養皿中,再放置培養箱中,三至四天後繼代。 PC-12 為 大 鼠 腎 上 腺 嗜 鉻 細 胞 瘤 細 胞 株 (Rat adrenal pheochromocytoma cells),生長於37℃、5%CO2培養箱,培養基為 Dulbemlo's modified Eagle's medium (DMEM) (Gibco)、10% fetal bovine. serum(FBS). (Invitrogen,. Carlsbad,. CA) 及 1 %. penicillin-streptomycin (100 units/ml, 100 μg/ml) (Invitrogen),為半懸 浮型細胞,而其繼代培養方式:由原本培養皿中的培養基沖下細胞, 再加入離心管離心(900rpm、3分鐘),抽掉上清液,再加入1ml培 28.
(32) 養基,混合均勻,再加入少許的混合液至已有培養基的培養皿中, 再放置培養箱中,約四至五天後繼代。. 第二節:藥劑 1. HMJ-53A於中國醫藥大學藥學院藥學系侯曼貞老師實驗室取得。 儲存藥物溶液濃度為100 mM,溶化於Dimethyl Sulfoxide(DMSO)。 其結構式與合成程序如圖A。HMJ-53A對細胞沒有毒性,因加完 HMJ-53A ,20分鐘後,利用trypan blue(10倍稀釋)測試N2A 細胞存活率,而其存活率>95%。. 圖A:HMJ-53A結構式與合成程序. 29.
(33) 2. 其他HMJ系列之藥物(HMJ-1、HMJ-29A及HMJ-73A)皆從中國 醫藥大學藥學院藥學系侯曼貞老師實驗室取得,儲存藥物溶液濃 度為50-100 mM,溶化於DMSO,而它們的結構式如圖B。. 圖 B:HMJ 系列之藥物結構式 30.
(34) 3. TEA Cl(tetraethylammonium chloride)鉀離子通道阻斷劑,購買 於Sigma-Aldrich (St. Louis, MO,USA),儲存藥物溶液濃度為1 M, 溶解於millipore water。. 4. Staurosporine為非選擇性蛋白酶抑制劑,購買於Tocris (USA),儲 存藥物溶液濃度為100 μM,溶解於DMSO。. 第三節:轉殖過程 PcDNA3-Kv2.1. 及 PEGFP 之 cDNA , 從 加 拿 大 多 倫 多 大 學 的. Professor Gaisano實驗室取得。再從裝有PcDNA3-Kv2.1及PEGFP之 cDNA的微量離心管中,分別取出 3 μg及 1 μg加至無菌微量離心管 中,並與Opti-MEM (Invitrogen, USA) (44 μl)混合均勻,等待20分鐘; 而在等待過程中,把Lipofectamine TM 2000 (Invitrogen, USA)10 μl與 40 μl的Opti-MEM,混合均勻並等待5分鐘。 之後,將兩個部分混合均勻,並再等待20分鐘,把有N2A細胞的 3.5 cm 培養皿,用PBS沖洗三次,加入無抗生素的culture medium (DMEM內含有10%FBS) 2 ml;把上述已過了20分鐘之混合物,共 100 μl加入至培養皿中,放至CO2培養箱過一晚後,用螢光顯微鏡 ( Excitation:484nm,Emission:510nm )觀察細胞是否有轉殖成功; 31.
(35) 假如成功會見到綠色的細胞;因為PEGFP (maker)已順利送至細胞核 內,並已分別表達在細胞質及膜上,再經由顯微鏡上藍光激發,可 見綠細胞,這就代表PcDNA3-Kv2.1也應該進入細胞了。如此,細胞 可以繼代分盤,隔日即可做電生理記錄。. 第四節:電生理(Electrophysiology)紀錄方法 一、溶液準備 (Leung et al., 2003): 1. 細胞外液 Bath solution (mM):140 NaCl , 4 KCl , 1 MgCl2 , 2 CaCl2 , 10 HEPES(用NaOH調整至pH 7.4)。 2. 細胞內液 Intracellular solution (mM):140 KCl , 1 MgCl2 , 1 EGTA , 10 HEPES , 5 MgATP(用KOH調整至pH 7.25)。 3. 微細玻璃管電極 (thin-walled borosilicate glass tubes)(OD 1.5 mm , ID 1.10 mm , Sutter Instrument , Novato , CA),用 puller (P-87 , Sutter Instrument)加熱拉開成兩根,再用 microforge (Narishige Instruments,Inc., Sarasota, FL)加熱打磨。 二、電生理記錄過程: 繼代細胞後隔日,拿出種有細胞的3.5 cm 細胞培養皿至室溫,用 細胞外液沖洗五次,再加入2 ml的細胞外液,放置顯微鏡下,並在 室溫( ~22℃ )下進行全細胞電壓箝制( whole-cell patch clamp )電生 32.
(36) 理之測量。 首先,加入細胞內液至微細玻璃管電極(電阻為4-7 MΩ)內,再 以微細玻璃管電極附上細胞膜,進行封口,並進一步產生全細胞電 流 測 量 模 式 。 利 用 電 壓 箝 制 放 大 器 ( EPC-10 amplifier, HEKA Electronik, Lambrecht, Germany )和Pulse 8.60來記錄全細胞電流變 化;而過濾是2 kHz,收集實驗結果頻率是10 kHz,再用Pulsefit 8.60( HEKA Electronik, Lambrecht, Germany )軟體分析結果。因電極 同時負責偵測細胞膜電位,並使電壓可箝制在設定之數值中,而 N2A細胞及PC-12 細胞,其膜電位被固定在-70 mV,再以不同流程 去測量,得到實驗結果(請詳見結果之圖例部分)。 三、濃度反應曲線: 以連續+30mV電壓刺激(持續3.5秒,間隔20秒)N2A細胞,再加 入不同濃度之HMJ-53A (1、3、6、10、18、30 μM)至N2A細胞,再 以 連 續 +30mV 電 壓 刺 激 , 再 記 錄 受 HMJ-53A 抑 制 後 的 穩 定 (steady-state)Kv電流,並以Hill方程式得到曲線。Hill方程式: Idrug/Icontrol = 1/{1+([HMJ-53A]/Kd)n},Idrug表示加入HMJ-53A後, 記錄到的Kv電流;Icontrol 表示未加入HMJ-53A,獲到的Kv電流, [HMJ-53A]表示加入細胞外液中HMJ-53A的濃度,Kd表分離常數,n 表示Hill係數。 33.
(37) 四、活化曲線圖及鈍化曲線圖: Kv通道活化之電壓依賴性,通常使用尾部電流 (tail current)去評 估,但是在此實驗中,因加入HMJ-53A後,鈍化速率非常快,所以, 用尾部電流去評估是不準確的。因此,用去極化的刺激而得到的Kv 電流和傳導性(G),然後用G = I/V-Vr 評估並得到曲線,而Vr = (RT/zF)ln(Ko/Ki),V表示實際設定的電壓,Vr表鉀離子反轉電位 ( reversal potential ),I是電流,R是氣體常數,T是溫度,z是離子電 荷數,而鉀離子為+1 ,F表法拉第常數 (Faraday constant),Ko和 Ki分別代表為細胞外液及微細玻璃管電極內之細胞內液的鉀離子 濃度。活化之電壓依賴性結果,可用波茲曼方程式 (Boltzmann equation)得到活化曲線,G/Gmax = 1/{1+exp[(V1/2 - V)/k]}來描述通 道開或關的機率與細胞膜電位之間的關係。V1/2代表全部通道打開 一半時的細胞膜電位,k值是斜率 (slope factor),代表通道之開關 對於細胞膜電位變化之依賴程度。 五、穩定狀態之鈍化 ( Steady-state inactivation )曲線: 由 I/Imax = 1/{1+exp[(V - V1/2)/k]} 得之,V1/2 表示全部通道打開 後,呈現鈍化狀態一半時的細胞膜電位, k 是斜率 (slope factor), 即通道之鈍化對於細胞膜電位變化之依賴程度。. 34.
(38) 第五節:電生理( Electrophysiology)記錄流程 因每個實驗流程不相同,為了方便閱讀,詳細的流程放置在每個 結果之圖例解說中。. 第六節:統計方法 結果數據顯示皆以means ± SEM;視實驗情況,用unpaired或paired student’s t test 去比較兩組差異,而 p<0.05為顯著差異。. 35.
(39) 第三章:研究結果 第一節:HMJ-53A在N2A細胞外側上作用,並加速慢性鈍 化現象 當N2A細胞被去極化,會產生衰退(decay)速度極慢的外電流 (Fig.1 A,B),而這樣的慢性衰退現象可稱之為C型鈍化 (Kurata and Fedida, 2006)。而在此實驗中,給予N2A細胞10分鐘的連續(每一 次刺激,間隔20秒)+30mV去極化,測得其電流衰退速度,並無因 時間增加而加快衰退速度 (Fig.1 A,B) (衰退 τ 在0秒和600秒,分 別為 1440 ± 170 ms 和 1376 ± 164 ms, n = 5)。 在Fig.1C,D中,N2A細胞連續去極化時,在細胞外加入30 μM的 HMJ-53A後,意外發現整個衰退速度大幅度的加快,而HMJ-53A影 響衰退速度至平衡時,衰退 τ 甚至小於 0.1秒(加入HMJ-53A之衰 退 τ = 85.6 ± 7.7 ms,n = 16;未加HMJ-53A之衰退 τ 在0 分鐘 = 1677 ± 120 ms,n = 19;p < 0.05)。 而這樣的影響是可逆的,也就是在產生加速衰退後,再以灌流沖 洗方式,洗走HMJ-53A,發現經過10至12分鐘的沖洗後,回復到未 加HMJ-53A的衰退速度(Fig.1E,F;恢復95.3 ± 16.5 %, n = 4)。 Fig.2中,表示HMJ-53A抑制穩定狀態電流之濃度依賴性。分別以 不同濃度1、3、6、10、18 及30 μM的HMJ-53A,看其抑制穩定狀 36.
(40) 態電流,其IC50為9.2 μM,並從Hill plot獲得Hill係數為3.5;因此數 值非常的高;所以,可得知HMJ-53A作用在N2A細胞之Kv通道,有 數個結合位置。 此外,我們在另一細胞株PC-12細胞,去證實HMJ-53A是否在另 一株細胞上,也會影響Kv電流之衰退速度;在PC-12細胞,給予連 續(每一次間隔20秒)+30 mV去極化,發現未加HMJ-53A (30 μM), 衰退 τ 為762 ± 272 ms,而加HMJ-53A (30 μM)等至作用平衡時, 其衰退 τ 加速至77 ± 13 ms (p<0.05, n = 6)。 更進一步,我們想探討出,到底HMJ-53A針對哪一種Kv通道作用 呢?而在N2A細胞轉殖入Kv2.1之cDNA,當測量到已轉殖入Kv2.1 cDNA的N2A細胞之電流(可>3 nA)後,給予連續(每一次間隔 20秒)+30 mV去極化,發現衰退 τ 為4600 ± 130 ms,而加入 HMJ-53A (30 μM)等至作用平衡時,其衰退 τ 加速至300 ± 70 ms (p<0.05, n = 3)。從此結果可得知, HMJ-53A會針對Kv2.1產生加速 慢性鈍化現象 (Fig.3)。 從以上結果,讓我們想到究竟HMJ-53A作用在細胞何處呢?而主 要作用位置,分為細胞內及細胞外兩大部分;首先探討細胞內作用, 在N2A細胞中,在微細玻璃管電極之細胞內液中加入HMJ-53A (30 μM)(表示細胞內加入)後,給予細胞連續(每一次間隔20秒)+30 37.
(41) mV去極化,經過15分鐘後,衰退速度並未改變(衰退 τ:0分鐘和 15分鐘分別為1270 ± 151 ms 和 1418 ± 128 ms, n = 4)。隨之於細 胞外液中加入(表示細胞外)HMJ-53A (30 μM)後,發現電流衰退 速度大幅度增加(於作用平衡,衰退 τ 為 58 ± 6 ms, n = 3) (Fig.4A,B)。 綜合以上結果可確定,HMJ-53A產生加速慢性鈍化現象,是作用 於細胞外並非細胞內。. 第二節:HMJ-53A作用,是否與蛋白質激酶之磷酸化有關 呢? 用 產 生 磷 酸 化 作 用 的 非 選 擇 性 蛋 白 酶 抑 制 劑 : Staurosporine (Cogolludo et al., 2003),去確定HMJ-53A是否因產生磷酸化作用而 加速Kv電流衰退。加入Staurosporine (100 nM)於培養皿中,等待30 分鐘後,給予N2A細胞連續(每一次間隔20秒)+30 mV去極化,發 現並未改變衰退速度;再加入HMJ-53A (30 μM)後,整個大幅度加 快其衰退速度。比較已加入HMJ-53A之Staurosporine (已加及未加) 兩組比較,發現加入Staurosporine不會改變HMJ-53A對衰退速度之 影響 [已加和未加Staurosporine (100 nM)分別為96±7 ms 和 105±9 ms,. p>0.05, n = 3 ]。 38.
(42) 從上述結果可知,HMJ-53A產生加速慢性鈍化現象,與蛋白質激 酶之磷酸化似乎無相關性。. 第三節:HMJ-53A作用在Kv通道之關閉階段 接下來要探討HMJ-53A作用在Kv通道之關閉或開啟階段呢?當 N2A細胞於全細胞電流測量後,先給予第一次+30 mV去極化並記錄 其電流 (Fig.5A),隨後在細胞外加入 HMJ-53A (30 μM),並等待4 分鐘(4分鐘內,並未給予任何去極化);而4分鐘後,再給予第二 次+30 mV去極化並記錄其電流 (Fig.5B),再比較這兩次的結果,發 現第二次比第一次的高峰電流下降至61.9 ± 5.3 %. (p<0.05, n = 4)。. 從此結果得知,HMJ-53A阻斷作用於Kv通道之關閉階段,進而以灌 流方式將HMJ-53A沖洗走,而其回復至未加HMJ-53A之電流(102.7 ± 3.8 %) (Fig.5C)。. 第四節:HMJ-53A並非為通道外孔之直接阻斷劑 HMJ-53A是否為通道外孔之直接阻斷劑呢?已知HMJ-53A作用 在細胞外,而作用在細胞外有兩種主要原因,造成Kv通道電流加速 衰退,第一個是直接堵住通道外孔上,第二個是使選擇性過濾器倒 塌所造成的。 39.
(43) 首先要探討的是,當Kv通道開啟時,是否HMJ-53A與通道上的孔 洞有親和力呢?假如此假設是正確的,HMJ-53A與通道之間的相關 性:在加入HMJ-53A後,會隨著去極化刺激上升,而增加電流衰退 速度。於是,我們著手實驗,給予不同程度去極化刺激,在N2A細 胞,加入HMJ-53A後,是否會影響電流衰退之速率 (Fig6)。在未加 HMJ-53A此組中,給予不同去極化刺激,發現電流衰退速度一致, 因給予極端的去極化 (> +70 mV)時,所以會有輕微電流衰退速度減 慢現象 (Fig.6A,C);而在加入HMJ-53A (30 μM)此組中,給予寬廣 範圍的去極化(0至+100 mV),得到的結果顯示,增加衰退的速度幾 乎一樣,不會因隨著去極化增加而衰退速度加快 (Fig.6B,C)。從此 結果可以反駁以下之假設:加入HMJ-53A後,因增加去極化刺激, 而增加HMJ-53A與通道親和力,使電流衰退速度更快。 Fig. 7A表示,在N2A細胞中,加入HMJ-53A (10 μM)(選擇此濃 度是因為其可大約阻斷Kv電流一半,見Fig.2)後,穩定狀態之Kv 電流的阻斷百分比,不會受不同程度電壓刺激而改變。不禁讓人聯 想到,藥物和通道間的親和力與通道開啟程度無相關性。 所以,更進一步去證實HMJ-53A並非直接堵住通道外孔,而造成 加速衰退現象。於是,改變細胞內鉀離子濃度去探查是否會影響 HMJ-53A作用?假如HMJ-53A直接堵住通道外孔,HMJ-53A與鉀離 40.
(44) 子會彼此在孔道上產生衝突。所以,降低細胞內的鉀離子濃度,將 會有助HMJ-53A阻斷電流之能力。因此,在此實驗中,降低一半細 胞內的鉀離子濃度(從140 mM降至70 mM)【為了維持細胞內滲透 壓,所以,加入140 mM 蔗糖(sucrose)】,是否會增加HMJ-53A作 用?而從結果得知,在N2A細胞上,對HMJ-53A (10 μM)阻斷Kv電 流百分比或是加快C型鈍化現象無影響 (Fig. 7B, C)。 為了更進一步確定HMJ-53A並不是堵住通道外孔而造成加速衰 退現象;用TEA來證實,而TEA是普遍使用的Kv通道阻斷劑,其作 用是直接堵住通道外孔而阻斷Kv電流 (Hille, 2001)。在此實驗中, 從細胞外中加入TEA (3 mM)後,立即抑制Kv電流,但是卻無改變 電流衰退速率 (Fig. 8)(衰退 τ,未加TEA之0分鐘和加入TEA後10 分鐘,分別為1727 ± 373 ms 和1430 ± 350 ms, n = 3),此結果與 HMJ-53A對比出衰退速度完全不同。 所以,綜合以上結果,強烈顯示,HMJ-53A並不是直接堵住通道 外孔的。. 第五節:HMJ-53A作用在Kv通道之鈍化閥門 由以上結果可知HMJ-53A並非直接堵塞通道外孔作用。因此, HMJ-53A加快電流衰退,有可能是作用在鈍化閥門關閉階段上,並 41.
(45) 讓人懷疑是否作用在選擇性過濾器本身上而加快其壓縮,增加C型 鈍化速度;換句話說,或許HMJ-53A作用是直接經由增加C型鈍化 速度。 更仔細去探討,在N2A細胞上,到底HMJ-53A如何影響Kv通道之 鈍化作用;所以,加入HMJ-53A後,觀察Kv電流的穩定狀態之鈍化 是否有受影響呢 (Fig. 9A)?在加入HMJ-53A此組,比未加HMJ-53A 此組,其穩定狀態之鈍化曲線左移12 mV(未加及已加HMJ-53A, 分別為V1/2 = -21.2 ± 2.5 mV和-33.0 ± 2.3 mV; p<0.05);不管如何, HMJ-53A並未改變其斜率(未加及已加HMJ-53A,分別為6.9 ± 1.3 and 5.9 ± 1.2;p>0.05)。而另一點引人注意,在未加HMJ-53A組別 中,鈍化現象在較強烈的去極化下 (> +50 mV)逐漸減慢,此結果稱 之為U型鈍化 (U-type inactivation)(Klemic, 2001; Kurata, 2002, 2005),而此鈍化作用,為何在極端的去極化中,會減弱其鈍化作 用,現今對於其主要的機制尚未明白。在已加HMJ-53A此組中,已 看不到U型鈍化,顯示對鈍化之抗拒作用,已被HMJ-53A克服。這 樣的結果可進一步支持HMJ-53A作用在鈍化閥門之假說。 另一方面,HMJ-53A是否會影響鈍化閥門的恢復呢?用dual-pulse 方式去檢視有或沒有加入HMJ-53A的Kv電流恢復的情況 (Fig.10)。 讓人意外的發現,有否加入HMJ-53A對Kv電流之恢復的速率沒有影 42.
(46) 響,兩者皆須要好幾秒才能完全恢復,而恢復速率最佳的fit方式為 double-exponential;快速恢復中,未加及已加HMJ-53A τ 分別為187 ± 55 和 142 ± 38 ms (p > 0.05; n= 6-7),而在慢速恢復中,未加及 已加HMJ-53A, τ 分別為3115 ± 982和1542± 810 ms(p > 0.05; n = 6-7)。從這些結果中得知,HMJ-53A加快鈍化閥門關閉的速度, 但卻不阻礙其恢復情形。. 第六節:HMJ-53A不會影響Kv 通道上之活化閥門 此實驗中,再深入研究,在N2A細胞中,是否HMJ-53A會影響Kv 通道中的活化閥門?從電壓依賴曲線圖 (-70mV至+70mV)中,可知 Kv通道之活化閥門並無受到HMJ-53A影響 (Fig.9B)(未加及已加 HMJ-53A,分別為V1/2 = 12.8 ± 3.2 mV和15.2 ± 1.6 mV;p>0.05), 並且HMJ-53A也不影響斜率(未加及已加HMJ-53A,分別為11.1 ± 0.5 和 13.9 ± 1.4; p>0.05)。 在Fig.9C中,比較未加及已加HMJ-53A組之活化kinetics,得知並 無分別。從Fig.9D的量化曲線圖顯示,Kv電流被更強之去極化刺激 時,隨之增加活化速率;已加HMJ-53A組別中,可得知在不同的去 極化刺激下,與未加HMJ-53A組別比較並不會改變其活化速率。由 以上結果可得知,HMJ-53A不會影響Kv通道之活化閥門。 43.
(47) 第七節:HMJ化合物其結構與產生加速鈍化閥門關閉之 相關性 綜合以上的結果,得知 HMJ-53A 有選擇性的作用在 Kv 通道上之 鈍化閥門,而加速其關閉。在 Fig11 表示數個 HMJ 衍生物,抑制穩 定狀態電流之濃度依賴性 (concentration-dependence)。而到底 HMJ 化合物的結構中,到底是哪一個部分(moiety)會造成這樣的作用呢? 在此實驗中,假如增加或減短 HMJ-53A 其中的 CH2CH3,是否會影 響作用的強度?而依 CH2CH3 的短長,分別有 HMJ-1、HMJ-29A、 HMJ-53A 及 HMJ-73A(見圖 A、B)。 在記錄 N2A 電流時,給予連續去極化+30mV,加入 HMJ-1 (30 μM), 並無抑制穩定狀態之電流。再進一步證實,在 N2A 細胞上,給予 連續去極化+30mV,再加入不同濃度之其它 HMJ 化合物,而看其 對穩定狀態電流之影響。得之 HMJ-29A 之 IC50 為 17.5 μM,並從 Hill plot 獲得 Hill 係數為 2.3。HMJ-53A 結果見 Fig2。HMJ-73A 之 IC50 為 6.5 μM,Hill 係數為 1.9。 從以上結果得知 HMJ-1 不影響 Kv 電流。而 HMJ-29A 為中度影 響 Kv 電流,HMJ-53A 及 HMJ-73A 為強度影響 Kv 電流 (Fig. 11)。 所以,綜合以上結果,與我們的假設符合,可以得知 HMJ-53A 的 結構中,CH2CH3 為主要影響鈍化閥門的關閉加速之結構。 44.
(48) 第四章:討論 Kv 通道之 C 型(慢性)鈍化現象的分子機制作用,目前並不是 完全了解,而現今已知其作用機制為:Kv 通道上的選擇性過濾器 或是其附近位置被壓縮,而導致孔洞變小,而使鉀離子無法順利通 過孔洞 (Kurata and Fedida, 2006;Hille, 2001)。而在藥理學中,作 用於 C 型(慢性)鈍化閥門之探針藥是很重要的。C 型(慢性)鈍 化現象,調節神經細胞的興奮性並產生動作電位之輪廓及頻率,是 非常重要;但是,至目前為止,特定針對於 C 型(慢性)鈍化現象 之機制探討的藥物是很缺乏的。 所以,在此研究中發現在N2A (neuroblastoma)細胞上,HMJ-53A 能加速Kv電流慢性鈍化現象。我們證實 HMJ-53A在N2A細胞中Kv 通道產生此作用,是在細胞外並非細胞內 (Fig.4)。而Fig.5之結果, 可以是由於兩種可能性:第一個,HMJ-53A之作用是在Kv通道關閉 階段;第二個,HMJ-53A之作用是在Kv通道開啟階段。假設作用於 開啟階段時,當HMJ-53A作用在Kv通道上時,會造成活化速率變慢, 且足以阻礙防止電流至最高峰。但從結果上可知,不可能為後者的 假設,因為從Fig.9C,D可知,活化kinetics並不會受到HMJ-53A的影 響。 在Fig.5的結果上說明,假設當通道剛開啟,HMJ-53A直接堵住通 45.
(49) 道上外孔,並部分重疊正在活化的電流;對於這樣的想法,需提出 的一個假設為:第一階段,HMJ-53A快速地直接堵住孔洞,且在一 開始活化後非常快速的堵住孔洞,而之後第二階段中,在細胞持續 極化情況下,相對以緩慢的速度繼續接近其結合位置,隨之產生越 明顯的電流下降情形。假如前述的第二個階段存在,很有可能表示 有一個drug-channel結合的位置,而此位置是靠去極化(或通道打開) 才暴露出來的。 所以,在我們的研究中去進一步探討,HMJ-53A與通道相互作用, 是否與各種不同程度的去極化有關呢?然而,當有HMJ-53A作用 時,在不同程度的去極化下,電流衰退的速度是不變的 (Fig. 6)。 再者,HMJ-53A抑制Kv電流的百分比也是非電壓依賴性的 (Fig.7A)。 還有,假如HMJ-53A直接堵住通道上孔洞,則將會與鉀離子在通道 之孔洞內相遇;假如降低細胞內鉀離子濃度,HMJ-53A將會更強地 阻礙鉀離子的流出。然而,從結果中得知,HMJ-53A (10 μM)對於 抑制Kv電流的百分比或加速C型鈍化,皆不因為大幅度降低細胞內 鉀離子濃度而受影響 (Fig.7B,C)。 由以上我們的結果可知,HMJ-53A的作用不太可能是直接堵塞通 道洞孔所造成的;這樣的結果更能顯示HMJ-53A作用在鈍化閥門。 可讓我們想像,當Kv通道在關閉狀態,HMJ-53A已經鈍化通道,因 46.
(50) 此讓我們得知,HMJ-53A阻斷Kv通道是在關閉狀態,當Kv通道打 開後,HMJ-53A加速鈍化閥門的關閉。 在Shaker (Kv1)通道及一些哺乳類動物相似Shaker之通道中,提升 細胞外鉀離子濃度和從細胞外加入TEA,會阻礙C型鈍化 (Choi et al., 1991; Fedida et al., 1999; Lopez-Barneo et al., 1993; Molina et al., 1997)。無論如何,在N2A細胞上,Kv通道中的C型鈍化速率,並不 會受到TEA (3 mM)的影響 (Fig. 8)。這樣的結果,或許是因為在N2A 細胞中,並無哺乳類動物相似Shaker之通道存在。而在我們實驗中, 則顯示HMJ-53A能作用於Kv2.1,而加速C型鈍化 (Fig. 3)。 但是有趣的是,HMJ-53A不會使Kv通道回復的速率減慢 (Fig. 10), 這也表示,在復極化階段,HMJ-53A非常快速的離開Kv通道。基於 HMJ-53A抑制電流的Hill 係數3.5 (Fig.2),表示HMJ-53A在Kv通道 上,有數個結合位置;因此我們提出一個觀點,在Kv通道上, HMJ-53A作用於某一個結合位置,而產生closed channel block,而 HMJ-53A作用於其他結合位置,而加速慢性鈍化。所以,在復極化 期間,HMJ-53A離開後者之結合位置比較快,而不減慢及影響通道 的恢復時間。 關於HMJ-53A與鈍化閥門之間的相互作用,HMJ-53A導致穩定狀 態之鈍化曲線左移12 mV。在未有HMJ-53A的N2A細胞上,可以記 47.
(51) 錄一個明顯的U型鈍化之Kv電流。因此,在較強的去極化中 (> +50 mV),鈍化閥門不容易關閉。有文獻指出,Kv1.5、Kv2.1和Kv3.1, 會表現U型鈍化 (Klemic et al., 2001; Kurata et al., 2005; Kurata et al., 2002);這或許表示N2A細胞中,存有一個或著一些這樣的通道,才 產生這樣的U型鈍化。 在極端的去極化下,Kv通道難以鈍化,而因此表現之U型鈍化, 其分子機制至今還不清楚。而HMJ-53A能克服此U型鈍化 (Fig. 9A), 更能證明HMJ-53A能加劇鈍化閥門的關閉。 HMJ-53A是否能影響細胞質內的活化閥門作用呢?在此我們提 供了證據,去顯示HMJ-53A對於Kv通道上活化的電壓依賴性和 kinetics並不影響 (Fig. 9B – D)。綜合以上結果,作用於Kv通道的鈍 化閥門的HMJ-53A是一個新穎的藥物。 目前已有幾篇報導指出,有幾種化合物可以增加鈍化閥門速度。 KN-93是目前已知作用在細胞外,而能提升Kv通道上慢性鈍化的速 度 (Ledoux et al., 1999; Rezazadeh et al., 2006);但此藥物的作用及 影響,沒有完全之可逆性且會作用在活化閥門:活化曲線有輕微的 左移現象,並且Boltzmann斜率有明顯的下降。 (Rezazadeh et al., 2006)。而亞麻油酸 (Linoleic acid)也能在Kv1.5和Kv2.1中,加速慢 性鈍化,而其作用在細胞外而非細胞內 (McKay and Worley, 2001); 48.
(52) 況且,亞麻油酸會影響活化閥門,使活化曲線左移和加速活化 kinetics (McKay and Worley, 2001)。因此,在我們的研究中發現, HMJ-53A的作用是完全可逆的和有選擇性的作用在鈍化閥門上的。 故在往後的研究中,HMJ-53A將可以作為深入探討Kv通道之鈍化閥 門之理想藥物。 Quinidine目前已知能導致C型鈍化加速 (Wang et al., 2003),但 quinidine此作用須經由細胞內allosteric的動作且須通道先開啟而才 能把電流阻斷。此外,4-aminopyridine是一個作用在細胞質內的Kv 通道阻斷劑,但卻被證實出能抑制Shaker通道之C型鈍化,但其機 轉不甚清楚 (Claydon et al., 2007)。 最後,我們要問的,是否HMJ-53A可作用在細胞膜上或細胞內而 激發蛋白質激酶,進而影響鈍化閥門?從我們的結果得知,加入 staurosporine至N2A細胞後,不會改變細胞本身的電流衰退,或者也 不影響加入HMJ-53A後的C型鈍化速度。而這些否定的結果可以讓 我們確定HMJ-53A並不是作用在細胞膜上而再經由某些磷酸化訊 息傳遞路徑,去調控C型鈍化。而從HMJ-53A不能在細胞內影響Kv 電流的實驗中,意味著HMJ-53A的作用,並不是經由調控細胞內的 因子或是擾亂細胞膜上脂質(lipid)的平衡而造成的 (Oliver et al., 2004)。 49.
(53) 往後之研究方向,我們想繼續探討在 HMJ 系列衍生物的結構上, 大幅度地加長 CH2CH3,看其能否加倍增強鈍化之能力?我們也想 深入探討 HMJ 衍生物,對於其它已知存在神經細胞上之 Kv 通道, 例如:Kv1.1 或 Kv1.2…等之影響 (Sheng et al., 1993; Sheng et al., 1994; Shimada et al., 2007)。另一方面,當 HMJ-53A 在高濃度情況 下(例如:100μM),是否會對 Kv 通道上之其它動作有影響,而 在 HEK 細胞株上表達 Kv4.3,去深入探討是否高濃度的 HMJ-53A 會影響 Kv 通道上之 N 型鈍化呢? 我們想再深入研究一個有趣的問題,在生理層面,當神經細胞受 損時,例如:神經退化疾病;此 HMJ 衍生物其是否有神經保護之 功能呢?而作為一種神經保護嶄新之用藥呢?這是往後我們想再 深入探討之方向。. 50.
(54) 第五章:結果圖表與說明 Fig. 1:HMJ-53A 加速 N2A 細胞 Kv 電流慢性鈍化現象 N2A細胞的holding potential是-70mV,連續給予+30 mV去極化 刺激(持續3.5秒,間隔時間為20秒)。從結果中每一次刺激的電流 最高往最低點fit位置,而得到衰退 τ。 (A)典型的K+ 電流外流曲線,被+30mV去極化引起,分別於0秒和 經過600秒後,且未加任何藥物。 (B) 衰退時間常數從(A)圖得之,每個時間點被plot出 。 (C)典型的K+ 電流外流曲線,被+30 mV引起,加入HMJ-53A (30 μM) 之前後之記錄。 (D)衰退時間常數從(C)圖得之,於未加及已加HMJ-53A(30 μM)後之 每個時間點plot出。 (E)典型的K+ 電流外流曲線,被+30 mV引起,記錄於未加及已加 HMJ-53A(30 μM),和從細胞外沖洗走HMJ-53A後,得之的曲 線。 (F)衰退時間常數從(E)圖得之,於未加、已加及沖洗走HMJ-53A (30 μM)之每個時間點plot出。 以上實驗得到的結果,可重複次數皆多於三個細胞以上。. 51.
(55) Fig. 1. 52.
(56) Fig. 2:HMJ-53A 抑制 Kv 電流濃度依賴曲線 細胞的 holding potential 為-70mV,連續給予+30mV 去極化刺(持 續 3.5 秒,間隔時間為 20 秒)。再加入不同濃度的 HMJ-53A (3、 6、10、18 及 30 μM),並給予同樣的刺激,記錄穩定狀態之 Kv 電流。 把標準化之穩定狀態電流(已加藥/未加藥),作 Y-軸,並把 HMJ-53A 之濃度作 X-軸。 曲線用 Hill 方程式 fit 之。結果為 mean ± SEM,從 3 至 6 個細胞 得之。. 53.
(57) Fig. 2. 54.
(58) Fig. 3:HMJ-53A 在轉殖 Kv2.1 後之 N2A 細胞上,加速 Kv 電流慢性鈍化現象 在已轉殖 Kv2.1 之 N2A 細胞,holding potential 為-70 mV,給予 連續+30 mV 去極化刺激(持續 3.5 秒,間隔時間為 20 秒)。得到 K v2.1 電流,再加入 HMJ-53A (30 μM)。曲線分別為未加藥及加藥 後 12 分鐘。 以上實驗得到的結果,可重複於三個細胞上。. 55.
(59) Fig. 3. 56.
(60) Fig. 4:從 N2A 細胞內加入 HMJ-53A 並不影響 Kv 電流之 慢性鈍化 (A) 在微細玻璃管電極之細胞內液中(細胞內)加入HMJ-53A (30 μM)。細胞的 holding potential是-70 mV,給予連續+30 mV去極 化刺激(持續3.5秒,間隔時間為20秒)。得到典型的K+ 電流外 流曲線,從0 分鐘至15 分鐘記錄。 再從細胞外,加入HMJ-53A (30 μM),皆在同一個N2A細胞上進 行,得之曲線。 (B) 從結果中每一次刺激的電流最高往最低點fit,而得到衰退 τ。 衰退時間常數從(A)圖得之,於未加及已加HMJ-53A (30 μM)之每 個時間點plot出。 以上實驗得到的結果,可重複於三個細胞上。. 57.
(61) Fig. 4. 58.
(62) Fig. 5:HMJ-53A作用於N2A細胞之Kv通道之關閉階段: (A) 細胞的holding potential為-70 mV,給予第一次+30mV去極化, 得之曲線。隨後給予此細胞HMJ-53A (30 μM),等待4分鐘,並 此期間未給予任何去極化刺激。 (B) 此細胞給予第二次+30mV去極化,並記錄其曲線。 (C) 以灌流方式,從細胞外沖洗走HMJ-53A,並連續(持續3.5秒, 間隔時間為20秒)給予+30mV去極化並記錄其曲線。 以上實驗得到的結果,可重複在多於三個細胞以上。. 59.
(63) Fig. 5. 60.
(64) Fig. 6:HMJ-53A加劇電流之衰退,沒有電壓依賴性 (A) 細胞的holding potential為-70 mV,開始給予刺激,一次持續10 s, 間隔10秒,每增加10 mV刺激一次,至+100 mV為止,停止刺激。 得到典型的K+ 電流外流曲線,被不同的電壓去極化引起。 (B) 加入HMJ-53A (30 μM),經過15分鐘後;同(A)刺激方式,測量 記錄曲線。得到典型的K+ 電流外流曲線,被不同的電壓去極化 引起。 (C) 從每一次刺激的電流之最高往最低點fit之,而得到衰退 τ。衰 退時間常數根據每個不同的去極化電壓plot出。 每一組結果的mean ± SEM,從4個細胞中得到。. 61.
(65) Fig. 6. 62.
(66) Fig. 7:HMJ-53A 阻斷 Kv 電流之強度不受電壓與細胞 內鉀離子濃度影響 (A)HMJ-53A (10 μM)阻斷穩定狀態 Kv 電流之百分比,根據每個不 同的電壓下 plot 出。 細胞的 holding potential 為-70mV,開始給予刺激(一次持續 10 秒,間隔 10 秒),每增加 10mV 刺激一次,至+100mV 為止, 停止刺激。再比較兩組其每個電壓中抑制的電流百分比(已加 藥/未加藥電流)×100%,所得到此曲線。 (B) HMJ-53A (10 μM)阻斷穩定狀態之Kv電流百分比。 兩組:微細玻璃管電極內(表示細胞內)鉀離子濃度分別為140 mM 和70 mM,細胞的 holding potential為-70mV,給予連續+30 mV去極化刺激(一個刺激持續3.5秒,間隔時間20秒)。抑制的 電流其百分比(已加藥/未加藥電流)×100%,再比較兩組的差 異。 (C)在細胞內鉀離子濃度分別為 70 和 140 mM 時,Kv 電流的衰退時 間常數 [以加及未加 HMJ-53A (10 μM)]。 微細玻璃管電極內(表示細胞內)鉀離子濃度分別為140 mM和 70 mM。細胞的 holding potential是-70mV,給予連續+30 mV去 63.
(67) 極化刺激,(一個刺激持續3.5秒,間隔時間為20秒)。再分為 未加及已加HMJ-53A(10 μM)兩大組,從結果中的電流最高至最 低點fit之,而得到衰退 τ。 每一組結果之mean ± SEM,從4至5個細胞中得到。. 64.
(68) Fig. 7. 65.
(69) Fig.8:TEA對電流衰退之影響與HMJ-53A不同 (A) 細胞的holding potential為-70 mV,給予連續+30 mV去極化刺激, (一個刺激持續3.5 秒,間隔時間為20秒)。得到典型的K+ 電 流外流曲線,再加入TEA (3 mM)(20秒和600秒後)。 (B) 衰退時間常數:比較未加及已加TEA (3 mM)的結果。於電流最 高至最低點fit之,為於每個時間點之衰退 τ。 以上實驗得到的結果,可重複於三個細胞上。. 66.
(70) Fig.8. 67.
(71) Fig. 9:HMJ-53A 使穩定狀態之鈍化曲線左移 但不影響活化閥門 (A)已加及未加HMJ-53A (30 μM)之N2A細胞中,進行Kv通道電流測 量,得之穩定狀態之鈍化曲線。在此實驗使用dual-pulse流程(給 予一次pre-pulse,再給予第二次短的pulse)做測量。 細胞的holding potential是-70 mV,先給予pre-pulse刺激(持續 10 s),再給予一個+70 mV的test pulse。休息10秒後,接著下一 個dual-pulse刺激 (於pre-pulse,每次增加10 mV),刺激至 pre-pulse為+100 mV,停止刺激。 而test pulse電流與最大之test pulse電流作標準化,兩組曲線比 較其差異。因為在對照組產生‘‘U型鈍化’’現象,所以只從-80至 +50mV fit(Boltzmann方程式)。結果之mean ± SEM從每組中4 個細胞中得之。 (B) 活化電壓依賴性:Kv 電流被不同電壓去極化刺激,而這些不同 的電壓,從holding potential為-70mV開始,每增加10mV刺激一 次(一次持續0.5秒,間隔2秒),至+70mV止。 Conductances (G)(conductance計算於材料方法中說明)在對 照 組 和 加 入 HMJ-53A 組 中 , 分 別 與 各 自 最 大 的 conductance 68.
(72) (Gmax)作標準化,在不同的去極化刺激下plot出結果。而此曲線 是以Boltzmann方程式去fit之。結果之mean ± SEM從每組中6個 細胞中得之。 (C)較 早 階 段 的 電 流 traces 代 表 活 化 之 kinetics 。 細 胞 的 holding potential為-70mV,給予連續+30 mV去極化刺激(一個刺激持續 3.5 秒,間隔時間20秒),再加入HMJ-53A。而標準化這兩組並 重疊其電流,來比較未加及已加HMJ-53A這兩組。 (D)同 (C) 之 刺 激 方 式 , 活 化 時 間 常 數 乃 於 電 流 上 升 階 段 作 exponential fit,於不同的去極化電壓,再plot出結果。而得到兩 組(未加及已加HMJ-53A)的活化曲線。 每組結果的mean ± SEM,從6至8個細胞中得到。. 69.
(73) Fig. 9. 70.
(74) Fig. 10:HMJ-53A不影響Kv電流之恢復 電流恢復實驗於N2A細胞中進行,用dual-pulse 流程,細胞的 holding potential為-70 mV,給予第一次pulse(+30 mV,持續5秒), 然後第二次pulse(+30 mV,持續200 ms);第一次pulse與第二次 pulse間隔時間不同。 把第二個電流 (I2)與第一個電流 (I1)的最大值作標準化 (I2/I1) 。 再把此標準化結果根據不同之時間間隔而plot之。而此曲線用 double-exponential功能去fit。 兩組的結果的mean ± SEM,從每組中4個細胞中得到。. 71.
(75) Fig. 10. 72.
(76) Fig. 11:HMJ衍生物抑制Kv電流之濃度依賴性曲線 N2A細胞holding potential為-70 mV,給予連續+30 mV去極化刺激 (持續3.5秒,間隔時間20秒)。再加入HMJ-1 (30 μM),得之電流, 與未加入HMJ-1最大的穩定狀態電流作標準化,再plot出此點。 同上刺激方式,分別加入不同濃度(3、6、10、18 及 30 μM)之 其它 HMJ 衍生物,得到之最大的穩定狀態電流與未加入 HMJ 衍生 物之最大的穩定狀態電流作標準化(Idrug/Icontrol),並依藥物之 濃度作 X 軸而 plot 之,曲線用 Hill 方程式 fit 之。 每組的結果 mean ± SEM,從 3 至 6 個細胞得之。. 73.
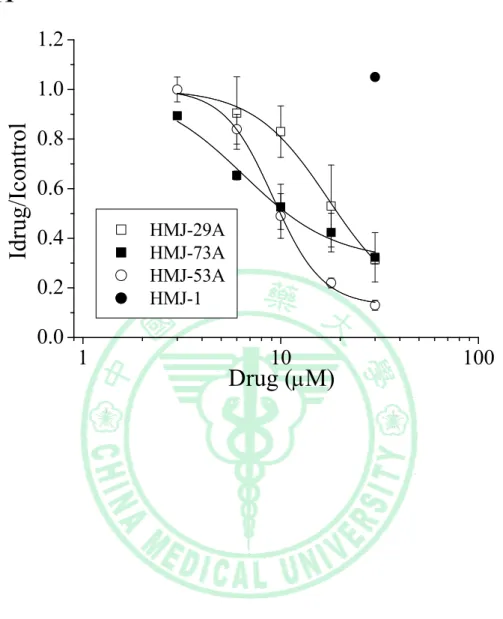
(77) Fig. 11 1.2. Idrug/Icontrol. 1.0 0.8 0.6 HMJ-29A HMJ-73A HMJ-53A HMJ-1. 0.4 0.2 0.0. 1. 10. Drug (μM). 74. 100.
(78) 第六章:参考文獻. Andalib, P., Consiglio, J.F., Trapani, J.G., and Korn, S.J. (2004). The external TEA binding site and C-type inactivation in voltage-gated potassium channels. Biophys J 87, 3148-3161.. Baukrowitz, T., and Yellen, G. (1996). Two functionally distinct subsites for the binding of internal blockers to the pore of voltage-activated K+ channels. Proc Natl Acad Sci U S A 93, 13357-13361.. Bayliss, D.A., Talley, E.M., Sirois, J.E., and Lei, Q. (2001). TASK-1 is a highly modulated pH-sensitive 'leak' K(+) channel expressed in brainstem respiratory neurons. Respir Physiol 129, 159-174.. Bowlby, M.R., Fadool, D.A., Holmes, T.C., and Levitan, I.B. (1997). Modulation of the Kv1.3 potassium channel by receptor tyrosine kinases. J Gen Physiol 110, 601-610.. Berne, R.M., Levy, M.N., Koeppen B.M., and Stanton B.A., (1998). Physiology. St. Louis, Missouri press. 75.
(79) Choe, S. (2002). Potassium channel structures. Nat Rev Neurosci 3, 115-121.. Choe, S., Kreusch, A., and Pfaffinger, P.J. (1999). Towards the three-dimensional structure of voltage-gated potassium channels. Trends Biochem Sci 24, 345-349.. Choi, K.L., Aldrich, R.W., and Yellen, G. (1991). Tetraethylammonium blockade. distinguishes. two. inactivation. mechanisms. in. voltage-activated K+ channels. Proc Natl Acad Sci U S A 88, 5092-5095.. Choi, K.L., Mossman, C., Aube, J., and Yellen, G. (1993). The internal quaternary ammonium receptor site of Shaker potassium channels. Neuron 10, 533-541.. Claydon, T.W., Vaid, M., Rezazadeh, S., Kehl, S.J., and Fedida, D. (2007).. 4-aminopyridine. prevents. the. conformational. changes. associated with p/c-type inactivation in shaker channels. J Pharmacol Exp Ther 320, 162-172.. 76.
(80) Cogolludo, A., Moreno, L., Bosca, L., Tamargo, J., and Perez-Vizcaino, F. (2003). Thromboxane A2-induced inhibition of voltage-gated K+ channels and pulmonary vasoconstriction: role of protein kinase Czeta. Circ Res 93, 656-663.. Cordero-Morales, J.F., Jogini, V., Lewis, A., Vasquez, V., Cortes, D.M., Roux, B., and Perozo, E. (2007). Molecular driving forces determining potassium channel slow inactivation. Nat Struct Mol Biol 14, 1062-1069.. Doyle, D.A., Morais Cabral, J., Pfuetzner, R.A., Kuo, A., Gulbis, J.M., Cohen, S.L., Chait, B.T., and MacKinnon, R. (1998). The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science 280, 69-77.. Fedida, D., Maruoka, N.D., and Lin, S. (1999). Modulation of slow inactivation in human cardiac Kv1.5 channels by extra- and intracellular permeant cations. J Physiol 515 ( Pt 2), 315-329.. 77.
(81) Goldstein, S.A., Price, L.A., Rosenthal, D.N., and Pausch, M.H. (1996). ORK1, a potassium-selective leak channel with two pore domains cloned from Drosophila melanogaster by expression in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 93, 13256-13261.. Gulbis, J.M., Zhou, M., Mann, S., and MacKinnon, R. (2000). Structure of the cytoplasmic beta subunit-T1 assembly of voltage-dependent K+ channels. Science 289, 123-127.. Holmgren, M., Jurman, M.E., and Yellen, G. (1996). N-type inactivation and the S4-S5 region of the Shaker K+ channel. J Gen Physiol 108, 195-206.. Hoshi, T., Zagotta, W.N., and Aldrich, R.W. (1990). Biophysical and molecular mechanisms of Shaker potassium channel inactivation. Science 250, 533-538.. Hille, B., (2001). Ion channels of excitable membranes. Sunderland, Mass.: Sinauer press.. 78.
(82) Hour, M.J., Huang, L.J., Teng, C.M., Kuo, S.C., (2000). Synthesis and antiplatelet activity of alkoxy derivatives of 2-phenylquinazolines. Chin. Pharm. J. 52, 167-177.. Jahangir, A., and Terzic, A. (2005). K(ATP) channel therapeutics at the bedside. J Mol Cell Cardiol 39, 99-112.. Jiang, Y., Lee, A., Chen, J., Ruta, V., Cadene, M., Chait, B.T., and MacKinnon, R. (2003). X-ray structure of a voltage-dependent K+ channel. Nature 423, 33-41.. Kindler, C.H., Yost, C.S., and Gray, A.T. (1999). Local anesthetic inhibition of baseline potassium channels with two pore domains in tandem. Anesthesiology 90, 1092-1102.. Kiss, L., Immke, D., LoTurco, J., and Korn, S.J. (1998). The interaction of Na+ and K+ in voltage-gated potassium channels. Evidence for cation binding sites of different affinity. J Gen Physiol 111, 195-206.. Klemic, K.G., Kirsch, G.E., and Jones, S.W. (2001). U-type inactivation of Kv3.1 and Shaker potassium channels. Biophys J 81, 814-826. 79.
(83) Kukuljan, M., Labarca, P., and Latorre, R. (1995). Molecular determinants of ion conduction and inactivation in K+ channels. Am J Physiol 268, C535-556.. Kurata, H.T., Doerksen, K.W., Eldstrom, J.R., Rezazadeh, S., and Fedida, D. (2005). Separation of P/C- and U-type inactivation pathways in Kv1.5 potassium channels. J Physiol 568, 31-46.. Kurata, H.T., and Fedida, D. (2006). A structural interpretation of voltage-gated potassium channel inactivation. Prog Biophys Mol Biol 92, 185-208.. Kurata, H.T., Soon, G.S., Eldstrom, J.R., Lu, G.W., Steele, D.F., and Fedida, D. (2002). Amino-terminal determinants of U-type inactivation of voltage-gated K+ channels. J Biol Chem 277, 29045-29053.. Kurata, H.T., Soon, G.S., and Fedida, D. (2001). Altered state dependence of c-type inactivation in the long and short forms of human Kv1.5. J Gen Physiol 118, 315-332.. 80.
(84) Ledoux, J., Chartier, D., and Leblanc, N. (1999). Inhibitors of calmodulin-dependent protein kinase are nonspecific blockers of voltage-dependent K+ channels in vascular myocytes. J Pharmacol Exp Ther 290, 1165-1174.. Leung, Y.M., Ahmed, I., Sheu, L., Gao, X., Hara, M., Tsushima, R.G., Diamant, N.E., and Gaisano, H.Y. (2006). Insulin regulates islet alpha-cell function by reducing KATP channel sensitivity to adenosine 5'-triphosphate inhibition. Endocrinology 147, 2155-2162.. Leung, Y.M., Kang, Y., Gao, X., Xia, F., Xie, H., Sheu, L., Tsuk, S., Lotan, I., Tsushima, R.G., and Gaisano, H.Y. (2003). Syntaxin 1A binds to the cytoplasmic C terminus of Kv2.1 to regulate channel gating and trafficking. J Biol Chem 278, 17532-17538.. Lopez-Barneo, J., Hoshi, T., Heinemann, S.H., and Aldrich, R.W. (1993). Effects of external cations and mutations in the pore region on C-type inactivation of Shaker potassium channels. Receptors Channels 1, 61-71.. 81.
(85) Loussouarn, G., Rose, T., and Nichols, C.G. (2002). Structural basis of inward rectifying potassium channel gating. Trends Cardiovasc Med 12, 253-258.. MacKinnon, R., Aldrich, R.W., and Lee, A.W. (1993). Functional stoichiometry of Shaker potassium channel inactivation. Science 262, 757-759.. McKay, M.C., and Worley, J.F., 3rd (2001). Linoleic acid both enhances activation and blocks Kv1.5 and Kv2.1 channels by two separate mechanisms. Am J Physiol Cell Physiol 281, C1277-1284.. Molina, A., Castellano, A.G., and Lopez-Barneo, J. (1997). Pore mutations in Shaker K+ channels distinguish between the sites of tetraethylammonium blockade and C-type inactivation. J Physiol 499 ( Pt 2), 361-367.. Nerbonne, J.M. (2000). Molecular basis of functional voltage-gated K+ channel diversity in the mammalian myocardium. J Physiol 525 Pt 2, 285-298.. 82.
(86) Oliver, D., Lien, C.C., Soom, M., Baukrowitz, T., Jonas, P., and Fakler, B. (2004). Functional conversion between A-type and delayed rectifier K+ channels by membrane lipids. Science 304, 265-270.. Oxford, G.S., and Wagoner, P.K. (1989). The inactivating K+ current in GH3 pituitary cells and its modification by chemical reagents. J Physiol 410, 587-612.. Poolos, N.P., and Johnston, D. (1999). Calcium-activated potassium conductances contribute to action potential repolarization at the soma but not the dendrites of hippocampal CA1 pyramidal neurons. J Neurosci 19, 5205-5212.. Rezazadeh, S., Claydon, T.W., and Fedida, D. (2006). KN-93 (2-[N-(2-hydroxyethyl)]-N-(4-methoxybenzenesulfonyl)]amino-N-(4-c hlorocinnamyl)-N-methylbenzylamine),a calcium/calmodulin-dependent protein kinase II inhibitor, is a direct extracellular blocker of voltage-gated potassium channels. J Pharmacol Exp Ther 317, 292-299.. 83.
數據




+7





相關文件
Microphone and 600 ohm line conduits shall be mechanically and electrically connected to receptacle boxes and electrically grounded to the audio system ground point.. Lines in
Vacuum circuit breakers, for a voltage exceeding 1,000 volts but less than 72.5 kv 歐洲商會.
Owing to the pandemic of novel coronavirus pneumonia, Visitor Expenditure Survey was suspended in the 2 nd quarter of 2020, and thus no information is released for this
220V 50 Hz single phase A.C., variable stroke control, electrical components and cabling conformed to the latest B.S.S., earthing through 3 core supply cable.. and 2,300 r.p.m.,
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
The temperature angular power spectrum of the primary CMB from Planck, showing a precise measurement of seven acoustic peaks, that are well fit by a simple six-parameter
前項公司從事交易時,有以不合交 易常規之安排,規避或減少其在中華民
