國 立 交 通 大 學
生 物 科 技 學 院
生 化 工 程 研 究 所
碩 士 論 文
心房顫動組織與血管加壓素處理對心肌纖維母細胞中
MMPs 和 TIMPs 之表現調節
MMPs and TIMPs Expression in Fibrillating Atria and
Cardiac Fibroblasts Responding to Angiotensin Peptides
研 究 生:陳 文 琪
指導教授:林 志 生 博士
中 華 民 國 九 十 五 年 一 月
謝
誌
寫篇謝誌就像是在寫得獎感言,五味雜處,感觸良多。兩年的研究生涯就在一晃眼 中過去了,兩年的心情,就像是釀酒般的感受,不僅用盡全力在釀出一瓶好酒,對於我, 老師一定也會覺的像是在釀瓶女兒紅般的感受,百般滋味,盡待此刻。 如同一般人,我要感謝的人也是相當多,在這成立的高雄硬幫幫成員,目前已經先 畢業了一位,徐小翔,初到此,也許是高雄人的個性吧,直言直語,很快的讓我融入大 家的生活圈,還有那段唱雙簧的日子裡,默氣十足的沒話說;而尚待打拼中的建龍學長, 也是成員之一,學長的浪人個性及厚重的煙味,是我對這個家族的第一印象;靜怡,是 個漂亮的女孩,也是生科院之花,對她的第一印象就是有吃不完的早餐,在這的兩年生 活裡,因為有她的相互扶持,我才能有個舒發的心靈管道,也因為她,讓我在這繁忙的 研究生活中,仍能保持住我的美貌;瑞萍學姐,她是個相當有實驗經驗的人,因為她, 讓我解決許多在養細胞上的問題,及其它相關的細節。當然,她也是我的婚姻咨詢師, 對於許多婚前要注意的問題,都是她給我指點迷津;俊旭學長,是個相當虔誠的天主教 徒,不過他的幽默風趣及聰明才智,常常因他的一句穿心箭般的冷笑話,讓大家全都笑 到不支倒地,此外,他也是我們的實驗室電腦管理者,諸多電腦上的問題全靠他,對大 家而言,他算是我們大家的心靈窗口吧! 在這的生活,多虧了黃金美指的元平及越挫越勇的豐鵬,讓大家的生活增添了許多 樂趣,更帶給了我們大家青春活力及豐富的流行資訊。而欣怡和思豪兩個新成員,讓這 個剛失血完的實驗室又增添了新血。 對我而言,我相當感謝的就是我的指導教授 林志生老師,在這兩年的生活中,雖 然我是個不太聽話又相當衝動的學生,和老師的相處衝突多過平穩交談,但是,老師還 是相當有耐心的告誡我許多實驗上該注意的小細節及問題,讓我能順利完成實驗,在這 的實驗生活裡,我學會了勤儉忍耐,讓實驗以少少錢做多多事,遺憾的事,雖然在此兩 年,卻未能將老師的優點盡收袋底。 最後,我最想要感謝的是我的家人,雖然我的家人不在我身邊,大家各分異地,但 是大家的心還是彼此的互相鼓勵,也因為他們的鼓勵,才能讓我順利的取得大學及研究 所的學位,我相當慶幸能和他們成為家人,這是我人生的最大收獲。另外,有個不能不 提的人, 翁茂文博士,我非常感謝 柯俊良老師讓我們兩人相識,因為這樣,我才能在 生活及學業上多了另一個良師,我的衝動及任性常讓我在實驗上出問題,但他總是耐心 又細心的為我解答,不只讓我在感情上有所寄託,連在學業上也是我的心靈導師。非常 感謝柯俊良老師、陳春榮老師及楊進木老師,因為老師們的高抬貴手及相當有價值的建 議,讓我能在最後一刻,順利的取得學位。真的非常謝謝大家,為我做的一切,因為有 你們,我才能如願取得學位。心房顫動組織與血管加壓素處理對心肌纖維母細胞中 MMPs 和
TIMPs 之表現調節
研究生:陳文琪
指導教授:林志生博士
國 立 交 通 大 學
生 物 科 技 學 院
生 化 工 程 研 究 所 碩 士 班
中文摘要
心房顫動(atrial fibrillation; AF)為常見的心律不整疾病。在 AF 病程中,常見因細 胞外基質(ECM)的重塑(remodeling)造成的心房組織結構的病變,此種因 ECM 代 謝失常而堆積於細胞間隙的病灶,稱之為纖維化(fibrosis)。已知心臟纖維母細胞中的 基質金屬蛋白酶(matrix metalloproteinases; MMPs)與其組織抑制因子(tissue inhibitor of metalloproteinases; TIMPs)的表現調節,是心臟纖維化過程中調節膠原蛋白(collagen) 分解的重要因子。然而,在持續性(sustained)AF 病程中,MMPs 與 TIMPs 對於 ECM 代謝調節機轉至今仍未清楚。因此,本實驗主旨在探討於持續性 AF 組織與經 angiotensin II(Ang II)和 angiotensin-(1-7)(Ang-(1-7))處理的心臟纖維母細胞(cardiac fibroblast) 中,collagen type I、MMP-2、MMP-9、TIMP-1 及 TIMP-2 的表現調節情形。
在本研究中所使用的動物與細胞分別為經節律器激搏誘發(pacing-induced)持續性
AF的Yorkshire-Landrace豬隻與大鼠心臟纖維母細胞株H9c2。我們一共由14頭AF與12頭 正常竇律(sinus rhythm; SR:即sham control)豬隻取得心房組織。實驗方法則分別以 Masson’s trichrome染色與西方墨點法來檢測心房組織中ECM與collagen type I蛋白表現 量;利用gelatin zymography分析MMP-2與MMP-9酵素活性;以半定量RT-PCR方法測定 collagen type I、MMP-2、MMP-9、TIMP-1及TIMP-2的mRNA表現量。
實驗結果顯示,在AF心房的細胞間隙中有顯著組織纖維化病灶(AF, 21.6 ± 3.3%; SR,
5.2 ± 1.7%, p < 0.01);在AF心房組織中的collagen type I(1.79 ± 0.14)也顯著高於SR組
織中者(1.17 ± 0.26)(p < 0.01)。在AF心房組織中MMP-9的酵素活性與mRNA表現量分 別較SR組織高出7.1倍(p < 0.01)及4.3倍(p < 0.01);但是MMP-2活性,MMP-2、TIMP-1 及TIMP-2的mRNA表現量在AF和SR組織中則相似。在心臟纖維母細胞的實驗結果顯 示,經Ang II和Ang-(1-7)(10-9 , 10-7及10-5 M)處理的H9c2細胞之MMP-2活性皆下降(p < 0.05);另外在10-7
M的Ang-(1-7)處理下,可顯著誘發collagen type I 與MMP-9的mRNA 表現量(p < 0.05)。
本研究中,我們發現在持續性 AF 組織中有高度纖維化現象,並伴隨著高量 collagen type I 與 MMP-9 的表現,但是並不影響 MMP-2、TIMP-1 及 TIMP-2 的基因表現。我們 認為在 AF 發生組織結構性重塑病程中,可能與心臟纖維母細胞調控 MMP-9 表現的機 制有關;而當 MMPs 與 TIMPs 的平衡關係被破壞時,其會影響 ECM 代謝的異常,可能 造成持續性 AF 組織的纖維化病變。
MMPs and TIMPs expression in fibrillating atria and cardiac
fibroblasts responding to angiotensin peptides
Graduate Student: Wen-Chi Cheng Advisor: Chich-Sheng Lin Ph. D.
Institute of Biochemical Engineering College of Biological Science and Technology
National Chiao Tung University
Abstract
Atrial fibrillation (AF) is the most common cardiac arrhythmia. Structural abnormality is usually accompanied with extracellular matrix (ECM) remodeling process of AF. During structural remodeling of AF, abnormal ECM metabolism in the atrial tissues yields extensive interstitial matrix accumulation, a process termed fibrosis. Matrix metalloproteinases (MMPs) and their natural inhibitor of tissue inhibitor of metalloproteinases (TIMPs) are expressed in the cardiac fibroblasts have been shown to play important role in the regulation of collagen degradation in cardiac fibrosis. However, it is still unclear that mechanisms of MMPs and TIMPs in ECM metabolism during sustained AF. Therefore, the aims of this study was to explore altered collagen type I, MMP-2, MMP-9, TIMP-1 and TIMP-2 expression in the atrial tissues with sustained AF and in the cardiac fibroblasts treated with angiotensin II (Ang II) and angiotensin-(1-7) (Ang-(1-7)).
Yorkshire-Landrace pigs with pacing-induced sustained AF and sinus rhythm (SR; i.e., sham control) and a cardiac fibroblast H9c2 cell line of rat were used in this study. Atrial tissues were obtained from 14 pigs with sustained AF and 12 pigs with SR. The ECM was
investigated by Masson’s trichrome stain. Western blot was used to analyze the protein expression of collagen type I and gelatin zymography was used to assay the enzyme activity of MMP-2 and MMP-9. The gene expression of collagen type I, MMP-2, MMP-9, TIMP-1, and TIMP-2 were assayed by quantitative RT-PCR.
The myocardial fibrosis markedly accumulated in the interstitial cardiomyocytes of atrial tissues with AF (AF, 21.6 ± 3.3%; SR, 5.2 ± 1.7%, p < 0.01). The content of collagen
typeⅠin the AF subjects (1.79 ± 0.14) was also increased as compared with that in the SR subjects (1.17 ± 0.26) (p < 0.01). MMP-9 activity and expressed MMP-9 mRNA in the AF was higher than that in the SR by 7.1 folds (p < 0.01) and 4.3 folds (p < 0.01), respectively. However, MMP-2 activity and expressed MMP-2, TIMP-1 as well as TIMP-2 mRNA in the AF and SR subjects were insignificantly different. In the cardiac fibroblast H9c2 cells, the MMP-2 activity was decreasing for both Ang II and Ang-(1-7) (10-9, 10-7and 10-5M) treatments (p < 0.05). The mRNA levels of collagen type I and MMP-9 in the cells treated with 10-7M Ang-(1-7) could be significantly induced (p < 0.05).
In this study, we demonstrated the up-regulation of MMP-9 and collagen type I in the AF atria with fibrosis, but no significantly different on the mRNA levels of MMP-2, TIMP-1 and TIMP-2. We supposed that a role of structure remodeling in the AF is associated with MMP-9 regulated in cardiac fibroblasts. Moreover, imbalance of MMPs and TIMPs influences the ECM metabolism that may contribute in the process of sustained AF.
Keywords
AF atrial fibrillation
ECM extracellular matrix
MMPs matrix metalloproteinases
TIMPs tissue inhibitor of metalloproteinases
Ang II angiotensin II
Ang-(1-7) antiotensin-(1-7)
SR sinus rthym
CHF congestive heart failure
BK bradykinin
ACE angiotensin-converting enzyme
RAS renin-angiotensin system
Content
Acknowledgement … … … .i
Chinese Abstract… … … iii
English Abstract … … … ...v
Keywords… … … ...vi
Content… … … ...vii
Content of Tables… … … ..… … … .xi
Content of Figures… … … .… ...xii
Content of Annexes… … … ...x
. Research Background and Significance Ⅰ … … … 1
1-1. The relationship between atrial fibrillation with fibrosis… … … ...1
1-2. Relationship of cardiac remodeling and ECM… … … ...2
1-3. Relationship of cardiac remodeling and MMPs… … … 3
1-4. MMPs: structures and functions… ..… … … ...4
1-4-1. Gelatinase A (MMP-2, Type II collagenase)… … … ...6
1-4-2. Gelatinase B (MMP-9, Type V collagenase)… … … ...6
1-5. TIMPs: structures and functions… ..… … … ...7
1-5-1. TIMP-1… … … .… … … ...8
1-5-2. TIMP-2… … … .… … … ...8
1-6. Activation of the renin-angiotensin-aldosterone system in heart failure… ...9
1-6-1. Angiotensin II (Ang II)… … … ..… ...9
1-6-2. Angiotensin-(1-7) (Ang-(1-7))… … … ...10
II. Material and Methods… … … ..… … … 12
2-1. Reagents… … … ...12
2-2. Experimental model… … … ...12
2-3. Tissue sampling and treatments in AF… … … .… … … .13
2-4. Cell culture of cardiac myofibroblasts… … … 13
2-5. Isolated dufferent tissues from normal rat… … … .… ...14
2-6. Protein extraction and electrophoresis… … … .… ...14
2-7. MMPs activity by gelatin zymography… … … .… … … … ...14
2-8. RNA isolation… … … .… … … ...15
2-9. RT-PCR… … … .… … … .16
2-10. Western blot assay… … … ...17
2-11. Statistical analysis… … … .… … … ...17
III. Results… … … .… … … ...19
3-1 Induction MMP-9 in the atrial with AF… … … .… … … ...19
3-1-1. Histological findings in the atria with AF… … … .… … … ...19
3-1-2. Collagen type I protein in the atria with AF… … … … ..… … … ..20
3-1-3. Enzyme activities of MMP-2 and MMP-9 in the fibrillating atria… … … ..20
3-1-4. MMP-2 and MMP-9 mRNA levels in AF… … … … ...… … … ...21
3-1-5. TIMP-1 and TIMP-2 mRNA levels in AF… … … .… ..21
3-2 MMP-2 expression in cardiac fibroblasts treated with Ang II and Ang-(1-7)… … … 22
3-2-1. Collagen type I mRNA in the H9c2 treated with Ang II and Ang-(1-7)...22
3-2-2. MMP-2 activity in H9c2 cells treated with Ang II and Ang-(1-7)… … … .23
3-2-3. MMPs and TIMPs mRNA levels in the H9c2 cells treated with Ang II and Ang-(1-7)… … … .23
3-3 MMP-2 expression in the different tissues … … … ...24
3-4 Ratios of MMP2/TIMP-2 and MMP-9/TIMP-2… … … .25
IV. Discussion… … … ...26 V. References… … … ...31 Tables… … … .41 Figures… … … ...47
Annexes… … … .61
Content of Tables
Table 1. Classification and nomenclature of the MMPs… … … .41 Table 2. Common and unique features of TIMPs… … … ...43 Table 3. Review articles of matrix metalloproteinases on different diseases… … ...44 Table 4. Sequence of the primers specific for MMP-2, MMP-9, TIMP-1, TIMP-2,
collagen type I and GAPDH RT-PCR… … … ..… … … .46
Content of Figures
Figure 1. Masson’s trichrome stain atrial tissues in the SR and in the sustained atrial
fibrillation … … … ...47
Figure 2. Collagen type I in the atrial tissues with AF and SR subjects… … … 48
Figure 3. The induction of MMP-9 activity in the atria with AF… … … ...49
Figure 4. The induction of MMP-9 mRNA in the atria with AF… … … 50
Figure 5. Expression of TIMP-1 and TIMP-2 mRNA in the atria with AF and SR...51
Figure 6. The ratios of MMP-2/TIMP-2 and MMP-9/TIMP-1 mRNA in the atria with AF and SR… … … ..… … … .52
Figure 7. The effects of Ang II and Ang-(1-7) on collagen type I mRNA in the H9c2 Cells… … … .53
Figure 8. MMP-2 activation in the H9c2 cells induced by Ang II and Ang-(1-7)… … … … .54
Figure 9. The effects of Ang II and Ang-(1-7) on MMP-2 and MMP-9 mRNA in the H9c2 cells… … … ..… … … ...55
Figure 10. The effects of Ang II and Ang-(1-7) on TIMP-1 and TIMP-2 mRNA in the H9c2 cells… … … ...56
Figure 11. The ratios of MMP-2/TIMP-2 and MMP-9/TIMP-1 mRNA in the Ang II and Ang-(1-7)-treated H9c2 cells… … … ..… … ...57
Figure 12. Standard curve of MMP-2 activity … … .… … … ...58
Figure 13. MMP-2 activity in different tissues of rat… .. … … … .59
Figure 14. MMP-2 mRNA level in different tissues of rat...… … … .… … … ....60
Content of Annexes
Annex-1. Three hypothesis of AF mechanisms..… … … ...61 Annex-2. Schematic relationship between profibrotic factors and anti-fibrotic factors
in the regulation of ECM metabolism … … … ...70
Annex-3. Relationship of angiotensin II and fibrosis… … … 71
I. Research Background and Significance
1-1. The relationship between atrial fibrillation with fibrosis
Atrial fibrillation (AF), one of the most common arrhythmias in clinical practice, is related to increase disability and mortality. Recent research is focusing increasingly on the atrial structural remodeling, which underlies the development of AF in different pathological conditions[Dilaveris et al., 2005]. Typically, AF is characterized as a storm of electrical energy that travels in spinning wavelets across both atria, causing these upper chambers to quiver or to fibrillate at 300 to 600 times per minute waves can occur; the detailed mechanism behind this arrhythmia was still unclear[Goette and Lendeckel, 2004]. During the
development of AF in dilated cardiomyopathy and congestive heart failure (CHF), obvious structural changes in atrial myocytes may occur, including increase in cell size, perinuclear accumulation of glycogen, myolysis, alterations in connexin expression, changes in
mitochondrial shape, and homogeneous distribution of nuclear chromatin, which may facilitate AF recurrence and maintenance[Xu et al., 2004].
Cardiac remodeling is manifested clinically as changes in the size, shape, and function of the heart[Cohn et al., 2000]. Histopathologically, it is characterized by a structural
rearrangement of components of the normal chamber wall that involved cardiomyocyte hypertrophy, cardiac fibroblast proliferation, fibrosis, and cell death[Swynghedauw, 1999]. Fibrosis, which is a disproportionate accumulation of fibrillar collagen, is an integral feature of the remodeling characteristic of the failing heart[Kostin et al., 2000]. Accumulation of type I collagen, the main fibrillar collagen found in cardiac fibrosis, stiffens the ventricles and impedes both contraction and relaxation[Sun and Weber, 2005; Zannad and Radauceanu, 2005]. Fibrosis can also impair the electrical coupling of cardiomyocytes by separating myocytes with extracellular matrix (ECM) proteins[Swynghedauw, 1999]. Furthermore, fibrosis results in reduced capillary density and an increased oxygen diffusion distance that
can lead to hypoxia of myocytes[Sabbah et al., 1995]. Thus, fibrosis profoundly affectes myocyte metabolism and performance and ultimately ventricular function[Manabe et al., 2002; Schnee and Hsueh, 2000].
Symptom of AF has three remodeling: (1) electrical remodeling; (2) contractile
remodeling; (3) structural remodeling[Veenhuyzen et al., 2004] (Annex-1). The frequent coexistence of AF and heart failure has expanded the focus to the atrial myocardium, where profibrillatory include conduction slowing associated with scarring and fibrosis[Sanders et al., 2003]. In a cow heart failure model, despite recovery of atrial electrical and contractile remodelling, AF remained inducible and appeared to be related to persistent atrial fibrosis
[Cha et al., 2004]. In animals with long-standing AF, atiral fibrosis can be prevented by inhibition of the rennin-angiotensin system (RAS), which appears to significant reduce the duration of AF[Kumagai et al., 2003].
1-2. Relationship of cardiac remodeling and ECM
Cardiac fibroblasts play a central role in the maintenance of ECM in the normal heart and as mediators of inflammatory and fibrotic myocardial remodeling in the injured and failing heart[Brown et al., 2005]. In the myocardium, ECM proteins are mainly produced by fibroblasts that also produce matrix metalloproteinases (MMPs), growth factors, and cytokines, all of which are involved in the maintenance of myocardial structure, and in diseased hearts play pivotal roles in remodeling[Libby and Lee, 2000] (Annex-2).
The ECM is a dynamic structure, with continuous changes in the amount and proportions of its structural proteins that include different types of collagen, elastin, proteoglycans, and glycoproteins[Dollery et al., 1995].
Different enzymes such as the MMPs family help with the degradation of extracellular components[Li et al., 2000]. The myocardial ECM is made up of a fibrillar collagens
network, a basement membrane, proteoglycans, and glycosamioglycans and contains a diverse array of bioactive signaling molecules[Ito et al., 2005; Janicki and Brower, 2002]. The fibrillar collagen network ensures the structural integrity of the adjoining myocytes, provides the means by which myocyte shortening is translated into ventricular pump function, and is essential for maintaining alignment of the myofibrils within the myocytes through with a collagen-integrin-cytoskeletal myofibril relation[Janicki and Brower, 2002].
Myocardial fibrosis due to maladaptive ECM remodeling contributes to dysfunction of the failing heart[Li et al., 2000]. Fibrosis has been classified into two groups : reparative and reactive fibrosis. Reparative (replacement) fibrosis or scarring accompanies myocyte death. Reactive fibrosis appears as “interstitial” or “perivascular” fibrosis and does not directly associate with myocyte death. In interstitial fibrosis, fibrillar collagen appears in intermuscular spaces[Kai et al., 2005]. Perivascular firosis refers to the accumulation of collagen within the adventitia of intramyocardial coronary arteries and arterioles. Although there are a number of apparent differences between reparative and reactive fibrosis (eg, cells involved and the time course of fibrotic change), many factors likely work in common to control fibroblasts function[Kai et al., 2005].
1-3. Relationship of cardiac remodeling and MMPs
MMPs and their inhibitors, mainly tissue inhibitor of metalloproteinase-1 (TIMP-1), have been related to cardiovascular disease. Indeed, the MMP/TIMP system seems to be crucial to the ECM degradation seen in cardiovascular disease. For example, an increase in MMP activity has been noted in the shoulders of atherosclerotic plaques prone to rupture
[Galis et al., 1994; Johnson et al., 1998]or during remodeling after acute myocardial infarction[Thomas et al., 1998]. Descreased serum concentration of MMP-1 (the most important enzyme in the extracellular degradation of collagen) with raised levels of TIMP-1 in
patients with essential hypertension. Recently, plasma TIMP-1, the molecule that inhibits collagen degradation, has been proposed as a noninvasive marker of interstitial fibrosis
[Lindsay et al., 2002].
Structural abnormalities have been described in the atria from patients with AF
[Mary-Rabine et al., 1983], which are most severe in patients with chronic permanent AF. Recently, increased expression of ADAMs (A Disintegrin and Metalloproteinases), a new family of proteases that regulates cell-matrix interaction, has been reported in AF, suggesting that their increased activity could contribute to atrial remodeling[Arndt et al., 2002].
The expressions of ECMs and MMPs change dynamically during the developmental process of heart failure[Moshal et al., 2005]. In the present study, patients with AF have evidence of impaired matrix degradation, but this finding was not independently associated with the presence of AF on multivariate analysis[Goette et al., 2004]. In addition, after adjustment for confounding variables, there were also statistical correlations between the MMP/TIMPS system and echocardiographic indexes of left ventricular hypertrophy (left ventricular mass index) and left ventricular remodeling (end-diastolic ventricular diameter) but no relationship to left atrial size or systolic function parameters[Ke et al., 2005; Lombard et al., 2005].
1-4. MMPs : structures and functions
The MMPs are a family of at least eighteen secreted and membrane-bound
zincendopeptidases[Lombard et al., 2005; Ravanti and Kahari, 2000]. Collectively, these enzymes can degrade all the components of the ECM, including fibrillar and non-fibrillar collagens, fibronectin, laminin and basement membrane glycoproteins[Fedarko et al., 2004; Tschesche et al., 2000]. In general, a signal peptide, a propeptide, and a catalytic domain containing the highly conserved zinc-binding site characterizes the structure of the MMPs
[Nagase, 1994]. In addition, fibronectin-like repeats, a hinge region, and a C-terminal hemopexin-like domain allow categorization of MMPs into the collagenase, gelatinase, stomelysin and membrane-type MMP subfamilies[Nagase, 1994]. All MMPs are synthesized as proenzymes, and most of them are secreted from the cells as proenzymes. Thus, the activation of these proenzymes is a critical step that leads to extracellular matrix breakdown[Carmeli et al., 2004]. MMPs are considered to play an important role in wound healing, apoptosis, bone elongation, embryo development, uterine involution, angiogenesis and tissue remodeling, and in diseases such as multiple sclerosis, Alzheimer's, malignant gliomas, lupus, arthritis, periodontis, glumerulonephritis, atherosclerosis, tissue ulceration, and in cancer cell invasion and metastasis[Jones et al., 2003; Roeb and Matern, 2001]. Classification and nomenclature of all the types of MMPs were listed inTable 1.
MMP roles in normal and pathophysiological processes have been demonstrated. MMPs are involved in several cardiovascular disease processes, including coronary artery remodeling, particularly as it relates to plaque and aneurysm formation and rupture [Pyo et al., 2000; Silence et al., 2002]; Left ventricular remodeling follows pressure and/or volume overload or genetic alteration[Cox et al., 2002; Stroud et al., 2002]; and during all stages of CHF progression[Lindsey and Lee, 2000; Spinale, 2002]. All cell types found in the myocardium, either under basal conditions (myocytes, fibroblasts, endothelial cells) or response to an inflammatory stimulus (neutrophils and macrophages), express one or more types of MMP species. For example, cardiac fibroblasts can synthesize several groups of MMPs, including collagenases, gelatinases, stromelysins, and membrane type MMPs, as well as TIMPs 1-3[Birkedal-Hansen et al., 1976; Leicht et al., 2001; Wang et al., 2002]. MMP-9 increases during vessel remodeling in arteria, and MMP-9 null mice display decrease intimal hyperplasia and significant collagen accumulation in a carotid artery flow cessation model, indicating a role for MMP-9 in vessel matrix remodeling[Galis et al., 2002]. In a pig rapid
pacing model, MMP-1 levels increased and correlated with functional changes. An MMP inhibitor preserved LV size and function[Clair et al., 1998; Spinale et al., 1999]. When MMP-1 was overexpressed in mice (adult mice do not express MMP-1 protein), they
developed cardiac dysfunction at 12 months of age[Kim et al., 2000]. Together, these data implicate MMPs in vessel and myocardial matrix remodeling.
MMPs are a large family of enzymes that degrade the ECM. Of those enzymes, MMP-2 and MMP-9 are the most commonly proteolytic enzymes, also called type IV collagenases or gelatinases, are related enzymes that break down type IV collagen
[Morgunova et al., 1999].
1-4-1. Gelatinase A (MMP-2, Type II collagenase):
In 1978, Sellers et al. were the first to separate a gelatinase activity from collagenase and stromelysin in culture medium from rabbit bone[Sellers et al., 1978]. A similar enzyme, acting on basement membrane type IV collagen was reported by Liotta et al. the following year[Liotta et al., 1979]. Gelatinase was purified from human skin, mouse tumor cells, rabbit bone, and human gingival. The completed sequence of the human enzyme except for the signal peptide was reported by Collier et al[Collier et al., 2001]. Gelainase A had a triple repeat of fibronectin type I domains inserted in the catalytic domain; these participate in binding to the gelatin substrates of the enzyme[Lee et al., 1997; Libson et al., 1995].
1-4-2. Gelatinase B (MMP-9, Type V collagenase):
Harrwas and Krane in 1972 detected a gelatinase activity in rheumatoid synovial fluid. Sopata et al. described a gelatinase from human polymorphonuclear leukocytes[Sopata and Wize, 1979]. Rabbit macrophages produce a very similar enzyme which was able to digest type V collagen[Horwitz et al., 1977]. The neutrophil collagenase and gelatinase were
resolved in 1980[Murphy et al., 1980]. Purification was achieved in 1983 and sequencing of the cDNA in 1989. An interesting phenomenon, still not fully understood, is the binding of TIMP-1 to proMMP-9 to form a complex[Sakyo et al., 1983; Stetler-Stevenson et al., 1989]. Human neutrophil MMP-9 commonly occurs as a complex with lipocalin[Fernandez et al., 2005]. A series of papers concerned a 95 kDa protein in plasma that binds to gelatin culminated in the identification of this protein as MMP-9[Makowski and Ramsby, 1998].
1-5. TIMPs: structures and functions
The family of TIMPs presently numbers four distinct gene products that are specific inhibitors of the MMPs[Cook et al., 1994; Greene et al., 1996; Okada et al., 1994; Silbiger et al., 1994]. These secreted proteins are thought to regulate MMPs activity during tisssue remodeling[Baker et al., 2002]. All four mammalian TIMPs have many basic similarities, but they exhibit distinctively structural features, biochemical properties and expression patterns (Table 2). This suggests that each TIMP has specific roles in vivo. The local balance between MMPs and TIMPs is believed to play a major role in ECM remodeling during process of diseases such as cancer and arthritis[Anand-Apte et al., 1996]. The TIMPs have molecular weights of ~21 kDa and are variably glycosylated (Table 2)[Baker et al., 2002]. They have six disulphide bonds and comprise a three-loop N-terminal domain and an interacting three-loop C-subdomain. Most of the biological functions of these proteins discovere thus far are attributable sequences within the N-terminal domain, although the C-subdomain mediated interaction with the catalytic domains of some MMPs[Li et al., 1999]and with the hemopexin domains of MMP-2 and MMP-9[Brew et al., 2000]. The TIMPs are secreted proteins, but may be found at the cell surface in association with
membrane-bound proteins; for example, TIMP-2, TIMP-3 and TIMP-4 can bind MMP-14, a membrane-type (MT) MMP. All four TIMPs inhibit active forms of all MMPs studies to
date, their binding constants being in the low picomolar range, although TIMP-1 is a poor inhibitor of MMP-19 and a number of the MT-MMPs (Table 2)[Baker et al., 2002].
1-5-1. TIMP-1 :
TIMP-1 is one representative of the natural MMP inhibitor family, encompassing four members. It inhibits all MMPs, excepts several MT-MMPs. Unexpectedly, its
upregulation was associated to poor clinical outcome for several cancer varieties[Hornebeck et al., 2005]. TIMP-1 is a 28 kDa glycoprotein that appears to play a major role in
modulating the activity of interstitial collagenase as well as a number of connective tissue metalloendoproteases[Okada et al., 1994]. TIMP-1 functions through the formation of a tight 1:1 complex with active collagenase. TIMP-1 is a secretory product of platelets and alveolar macrophages. Previous studies indicate that regulation of MMP action by TIMP-1 is a critical event during the reproductive cycle[Nothnick, 2000; Nothnick, 2001a; Nothnick, 2001b].
1-5-2. TIMP-2 :
This gene is a member of the TIMP gene family. The proteins encode by this gene family are natural inhibitors of the MMPs, a group of peptidases involve in degradation of the EMC[Yu et al., 1996]. In addition to an inhibitory role against MMPs, the encoded protein has a unique role among TIMPs family in its ability to directly suppress the proliferation of endothelial cells[Stetler-Stevenson and Seo, 2005]. As a result, the encoded protein may be critical to the maintenance of tissue homeostasis by suppressing the proliferation of quiescent tissues in response to angiogenic factors, and by inhibiting protease activity in tissues
undergoing remodeling of the ECM. However, recent findings demonstrate that an
MMP-independent effect of TIMP-2 inhibits the mitogenic response of human microvascular
endothelial cells to growth factors[Stetler-Stevenson and Seo, 2005].
1-6. Activation of the renin-angiotensin-aldosterone system in heart failure
Several studies have demonstrated that a prolonged over-activation of neurohormonal mechanisms contributes to drive structural and functional abnormalities of the cardiovascular system and leads to poor prognosis in patients with CHF[Davila et al., 2005]. In particular, activation of the renin-angiotensin-aldosterone system (RAAS) leads to increased levels of angiotensin II and plasma aldosterone, and promote development of arterial vasoconstriction and remodeling, sodium retention, oxidative process, and cardiac fibrosis[Volpe et al., 2005]. In the past few years the combination of classical physiopharmacological techniques with modern genomics and protein chemistry methods has led to the identification of many novel components of the RAAS : the Ang VI binding site insulin-regulated aminopeptidase
[Albiston et al., 2001], angiotensin-converting enzyme 2 (ACE2)[Donoghue et al., 2000; Tipnis et al., 2000], the renin receptor[Nguyen et al., 1996], and the Ang-(1-7) receptor Mas
[Santos et al., 2003]. ACE inhibitors or Ang II receptor blockers and beta-blockers may modulate this excessive over-activity and improve survival in those patients[Cruden and Newby, 2004; Zeller and Battegay, 2005]. However, high circulating and tissue levels of Ang II and aldosterone may persist and contribute to further progression of CHF[Mahmud and Feely, 2004]. Many aspects of the pathophysiological role of the RAAS in CHF are still debated, and a more thorough comprehension of this fundamental system is needed
(Annex-3).
1-6-1. Angiotensin II (Ang II):
Angiotensin II (Ang II) is an important modulator of vascular homeostasis and an important link in the pathophysiology of cardiovascular disease[Brunner, 2001; Ruiz-Ortega
et al., 2001]. Elevated Ang II and/or increased sensitivity to AngⅡhave been etiologically associated with major vascular diseases[Brunner, 2001; Ruiz-Ortega et al., 2001]. Ang II was primary effector molecule of the rennin-angiotensin system (RAS). It was formed by the action of ACE on the precursor molecule Ang I and was primarily recognized for its role in the regulation of arterial pressure and blood volume[Ruiz-Ortega et al., 2001]. This vasopressor action of Ang II, which could be lifersave, may also lead to hypertension if the RAS was activated inappropriately[Ruiz-Ortega et al., 2001].
1-6-2. Angiotensin-(1-7) (Ang-(1-7)) :
The RAS is a major regulator of renal and cardiovascular function, play a pivotal role in the homeostasis of arterial pressure and of the hydro-electrolyte balance[Burnier and Brunner, 2000; Kim and Iwao, 2000]. Emerging evidence suggests Ang III [Ang-(2–8)], Ang IV [Ang-(3–8)], and Ang-(1–7) may also mediate the actions of the RAS[Ardaillou, 1997]. Ang-(1–7) has become an angiotensin of interest in the past few years, because its
cardiovascular and baroreflex actions counteract those of Ang II[Ferrario et al., 1998; Santos et al., 2000]. Recent reports have indicated heart and blood vessels as the main targets for the action of Ang-(1-7). These actions include biochemical and functional alterations leaded to vasodilation and improved cardiac function[Brosnihan et al., 1996; Ferreira et al., 2001; Loot et al., 2002]. Several studies suggest that the heptapeptide Ang-(1-7) has beneficial cardiovascular effects either directly or indirectly through bradykinin (BK) potentiation or by counter-regulation of the actions of Ang-(1-7). The presence and local generation of
Ang-(1-7) in the myocardium was first reported by Santos et al[Santos et al., 1990]in the canine heart. Immunoreactive Ang-(1-7) was demonstrated in the aortic root, coronary sinus, and right atrium under basal conditions[Ferreira and Santos, 2005]. Ang-(1-7) was
markedly reduced following treatment with the ACE inhibitor CGS-14,831[Santos et al.,
1990]. In isolated rat hearts, Ang I was extensively metabolized during a single pass through the coronary bed, leading to the generation of Ang II, Ang III, Ang IV and Ang-(1-7)
[Mahmood et al., 2002; Neves et al., 1995]. Ang-(1-7) formation in this model was not significantly changed by ACE inhibitors[Neves et al., 1995]. Ang-(1-7) was also formed in the intact human myocardial circulation but, in contrast to the rat heart, ACE inhibitors markedly decreased Ang-(1-7) generation[Zisman et al., 2003].
II. Materials and Methods
2-1. Reagents
Dulbecco’s Modified Eagle’s Medium (DMEM), and Trizol reagent were obtained from GIBCO-Invitrogen (Carlsbad, CA). Oligo-dT, dNTPs (dTTP, dATP, dGTP and dCTP), Pro Taq polymerase were purchased from Protech (Taipei, Taiwan). AngⅡand Ang-(1-7) and all other chemicals and biochemicals were obtained from Sigma (St. Louis, MO) and Merk (Darmstadt, Germany).
2-2. Experimental model
26 female pigs (average weight of 65 kg) were used. Of these, 12 were sham controls that were kept in sinus rhythm (SR), while the remaining 14 pigs were subjected to
experimentally induced AF as described in our previous report. In brief, Yorkshire-Landrace pigs were treated with intravenous anesthesia by thiamylal (2-3 mg/kg) (Kyorin
Pharmaceutical, Tokyo, Japan) and then transvenously implanted with a high-speed pacemaker (Itrel-III, model 7425; Medtronic, Minneapolis, MN). A screw-in atrial lead (Model 4568; Medtronic) was positioned at the right atrial appendage via the left internal jugular vein under fluoroscopy. The pacemaker was set to pace the atria at a rate of 400-600 beats per minute in the AF group. Consistency of the atrial pacing was checked daily in the first week and weekly thereafter with a portable ECG monitor by a programmer turning the atrial pacing on and off. After 4 to 6 weeks of continuous pacing, the pigs exhibited
sustained AF (i.e., AF was maintained after discontinuance of atrial pacing). The AF and SR group pigs were euthanized by high-dose intravenous barbiturate, the atrial pacemaker was removed and the animals were sacrificed for atrial tissue sampling. The experimental protocol conformed to the Guide for the Care and Use of Laboratory Animals (NIH
Publication No. 85-23, revised 1996) and was approved by the animal welfare committees of the National Chiao Tung University and the National Taiwan University. All pigs were provided by the Animal Technology Institute Taiwan (ATIT) and housed at the animal facility at ATIT[Lin et al, 2003].
2-3. Tissue sampling and treatments in AF
The left atrial appendages were excised from the isolated hearts and trimmed for
pathological examination and frozen storage. For RT-PCR and Western blotting, the excised specimens were immediately frozen in liquid nitrogen and stored at80°C until use for RNA or protein extraction. For pathological examination, the tissues were cut into small blocks about 105 mm and immersed in 4% phosphate-buffered formalin for 24 h. After
dehydration, each section was cut into 5m-thick slices. Deparaffinized sections were stained with Masson’s trichrome.
2-4. Cell culture of cardiac myofibroblasts
The H9c2 (2-1) cells (cardiac myofibroblasts) were purchased from BCRC (BCRC number: 60096). The cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Sigma) supplemented with 10% heat-inactived fetal calf serum (FCS). Subconfluent cells (70 - 80%) were subcultured 1:4. The cells were trypsinized, plated (2 x 105cells/well-2 x 106 cells/dish) in 12-well plate for gene expression analysis and MMPs activity or 100-mm dishes for protein analysis. Subconfluent serum starved cells were incubated with Ang II or
Ang-(1-7) (10-9, 10-7and 10-5M) for gene expression in 12h or protein assay and MMPs activity in 24 h.
2-5. Isolated multiple tissues from normal rat
Male Sprague-Dawley rats weighting 250 ~ 300 g were used in these experiments and all animals were maintained on food and water throughout the course of the study. The
experimental protocol conformed to the Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996) and was approved by the animal welfare
committees of the National Chiao Tung University and the National Taiwan University. Animals were anesthetized intraperitoneally with Avertin 0.4-0.6 mg/ body. . We collected the different sources of heart, liver, spleen, lung, kidney, brain, muscle, vessel, intestine and testis. The organs were rained in PBS and incubated in protein extract buffer or Trizol reagent for RT-PCR or protein analysis.
2-6. Protein extraction and electrophoresis
Frozen atrial tissues of pig and different tissues of rat (about 0.2 g) were homogenized in 1 ml of ice-cold lysis buffer containing 20 mM Tris-HCl, 1 mM dithiothreitol, 200 mM sucrose, 1 mM EDTA, 0.1 mM sodium orthovanadate, 10 mM sodium fluoride, 0.5 mM PMSF, and 1% (v/v) Triton X-100. The homogenate was then centrifuged at 12,000g at 4°C for 10 min, and the supernatant was collected for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Protein concentration was determined by the Bio-Rad protein assay kit (Bio-Rad, Hercules, CA) with bovine serum albumin as a standard. Aliquots containing 30g protein were resolved on 10% SDS-PAGE gels.
2-7. MMPs activity by gelatin zymography
For the determination of MMP-2 and MMP-9 activity, H9c2 cells were plated in 12-well culture plates at a density of 2 x 105cells/well and grown for 18 h before Ang II and Ang-(1-7)
treatment. After 24h treatment, the conditioned medium were collected at the indicated time points and centrifuged (6000 rpm, 30 min) then was rapidly frozen, and stored at -20℃. Atrial tissue was trypsinized, washed with ice-cold phosphate-buffered saline and lysed by sonication. Tissue extract were centrifuged (12,000 rpm, 10 min). Supernatant and condition medium were used for Zymography analysis. The MMPs activity analysis was done using total protein (20 μg/lane) or culture medium (20 μl/lane) electrophoresed on a 10% SDS-polyacrylamide gel under nonreducing condition. After electrophoresis, MMPs were refolded in 2.5% (v/v) Triton X-100 to remove the SDS, washed with distilled water then incubated 50 mM Tris pH 7.6, 200 mM NaCl, 5 mM CaCl2for 18 to 20 h at 37°C. The gel
was stained in Comaisse blue for 30 minutes prior to destain with destain buffer (50%
methanol, 10% acetic acid, 40% ddH2O). The presence of enzyme activity was evident by a
clear or unstained region, indicating the action of the enzyme on the gelatin substrate
[Stawowy et al., 2004]
2-8 RNA isolation
Total cellular RNA of the left atrial appendages, rat tissues and H9c2 cells were extracted as recommended by the manufacturer of TRIzol(GIBCO BRL, Rockville, MD). Briefly, the TRIzol method consists of the addition of 1 ml of the TRIzol reagent to each homogenized tissue (about 100 mg) or each well. The mixture was vigorously agitated for 30 sec and incubated at room temperature for 5 min. After this procedure, 200lchloroform was added to the tube, and the solution was centrifuged at 12,000g for 15 min. The aqueous phase was transferred to a clean tube, precipitated with 500lisopropyl alcohol, and
centrifuged at 12,000g for 15 min. The resulting RNA pellet was then washed with 1 ml of 75% cold ethanol and centrifuged at 7,500g at 4C for 5 min. The pellet was dried at
room temperature, resuspended in 20lof diethylpyrocarbonate-treated water, and stored at 80°C. RNA was quantified by measuring absorbance at 260 nm and 280 nm and
electrophoresed on a denaturing 1% agarose gel. The integrity and relative amounts of RNA were evaluated using ultraviolet visualization of ethidium bromide-stained RNA.
2-9. RT-PCR
For cDNA synthesis, 3g RNA was supplemented in a total reaction volume of 20 l with 1reverse trascriptase (RT) buffer, 0.5 mM dNTPs, 2.5 M oligo-dT (Invitrogen, Carlsbad, CA), 40 U/lRNase inhibitor (Invitrogen), and 20 U/lSuperscript IIreverse trascriptase (Invitrogen). After incubation for 60 min at 50°C, the mixture was incubated for 15 min at 70°C to denature the products. The mixture was then chilled on ice. PCR
primers for RT-PCR analysis are shown inTable 4. PCR reactions contained 2lcDNA, 1 leach primer (10 M), 5 l10 PCR buffer, 2 l10 mM dNTP, 1 lof 5 U/lTaq
polymerase (Violet Bioscience, Hsinchu, Taiwan) and 38ldistilled water in a total volume of 50 l. Thermal cycler (MiniCycler; MJ Research, Waltham, MA) conditions were as follows: 1 cycle of 5 min at 94°C, 21~36 cycles of denaturation at 94°C for 30~60 sec, annealing at 55°C for 30~60 sec, and elongation at 72°C for 45~120 sec, and 1 cycle of 5~10 min at 72°C. The resulting PCR products were visualized on 2% agarose gels stained with ethidium bromide. The stained image was recorded by an image analyzer (Kodak DC290 Digital camera System; Eastman Kodak, Rochester, NY), and the band intensity was
quantified using densitometric analysis by Scion image. The relative mRNA expression of the MMP-2, MMP-9, TIMP-1, TIMP-2 and collagen type I were calculated as ratios to
glyceraldehyde-3-phophate dehydrogenase (GAPDH) or 18S ribosomal RNA (18S rRNA) expression.
2-10. Western blot assay
Protein extracts of the left atrial appendages and rat tissues separated by SDS-PAGE were electrophoretically transferred to PVDF membranes (Immobilon-P; Millipore, Bedford, MA) by semi-dry electroblotting (HoeferTM77, Amersham Biosciences) (90 mA, 1 hr). Briefly, nonspecific binding sites were blocked by incubating membranes in 3% non-fat milk of Tris-buffered saline (0.1% Tween-20) and the membranes were incubated overnight with antibody against collagen type I (1:1000 dilution of anti-collagen type I mouse mAb, cat. no. Ab6308, Abcam), MMP-2 (1:1000 dilution of anti-MMP-2 (Ab-3) mouse mAb, cat. no.
IM33L, Calbiochem), MMP-9 (1:1000 dilution of anti-MMP-9 (626-644) (Ab-3) mouse mAb, cat. no. IM37L, Calbiochem) and anti--actin (1:10,000 dilution of -actin mouse mAb, cat. no. AC-15, Abcam). The probed blots were washed several times with 3% nonfat powdered milk in Tris-buffered saline containing 0.1% Tween-20. Antibody binding of incubated horseradish peroxidase-conjugated goat antimouse IgG (1:5,000 - 1:20,000 dilution) was visualized by Enhanced Luminol Chemiluminescence (ECL) Reagent (NEN, Boston, USA) and by exposing the membranes to X-Ray film (Super Rx Medical X-Ray Film; Fujifilm, Kanagawa, Japan). The bands were detected at the expected size. The band intensity was quantified using densitometric analysis by imaging software (Scion image; Scion, Frederick, MD). The amounts of collagen type I, MMP-2 and MMP-9 are expressed relative to the amount of-actin in respective samples[Weng et al., 2005a].
2-11. Statistical analysis
Statistical comparisons were performed using SPSS software (SPSS Inc.). Correlation between MMPs and TIMPs expression in RT-PCR analysis was performed using the paired sample t test. Densitometric analyses of the zymograms, Western blots and RT-PCR were
analyzed by on-way ANOVA with a LSD post-hoc test. Data are means ± standard deviation (SD), and statistical significance was considered if p < 0.05.
III. Results:
3-1. Induction MMP-9 in the atrial with AF
Sustained AF model in pigs has been established successfully in our laboratory[Lin et al., 2003; Hsu, 2003]. The electrophysiological and pathological findings in the porcine atria with AF showed the characteristics resembling those in human. Therefore, we have recognized a suitable animal model of sustained AF in adult pigs for studying human AF. AF is characterized by structural remodeling on cytoskeleton and ECM proteins[Lai et al., 2004; Lin et al., 2005]; however, the underlying molecular mechanisms involved ECM remodeling are largely unknown. To get a better insight into the ECM remodeling at molecular level, we analyzed the mRNA and activity changes in ECM metabolic enzymes, MMPs and TIMPs, in the fibrillating atria. Moreover, we attempt to test hypothesis whether the abnormal expressed MMPs are associated with the disease.
3-1-1. Histological findings in the atria with AF
Histological studies were conducted to identify the potential pathological substrate underlying the abnormalities in sustained AF. Atrial tissues from the AF subjects revealed that disordered or denatured cardiomyocytes were associated with extensive myocytolysis, thinning bundles and fragmentation of myofibrils, and large space within myofibrils[Hsu, 2003]. Masson’s trichrome staining showed that myocardial fibrosis markedly accumulated in interstitial cardiomyocytes in the AF atria(Figure 1A and 1B). The extent of fibrosis (%F) was measured and the result showed that %F of the atrial appendage in the AF group (21.6 ± 3.3%) was significantly greaten than that in the SR group (5.2 ± 1.7%, p < 0.01) (Figure 1C).
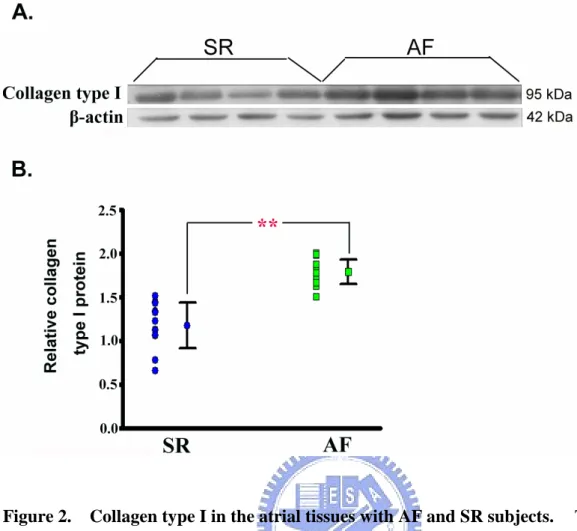
3-1-2. Collagen type I protein in the atria with AF
According the pathological results of Masson’s trichrome staining, ECM was significantly accumulated interstitial space in the atria with AF. Therefore, we were interesting to measure the collagen level in the tissue, because collagen was a major
component of ECM. The collagen type I in the AF (n = 14) and SR (n = 12) subjects was determined to Western blotting assay used the-actin as internal control(Figure 2A). The content of collagen typeⅠ(collagen/-actin) in the AF group (1.79 ± 0.14, ranging from 1.51 to 2.03) was significantly increased as compared with that in the SR group (1.17 ± 0.26, ranging from 0.66 to 1.51) ( p < 0.01) (Figure 2B).
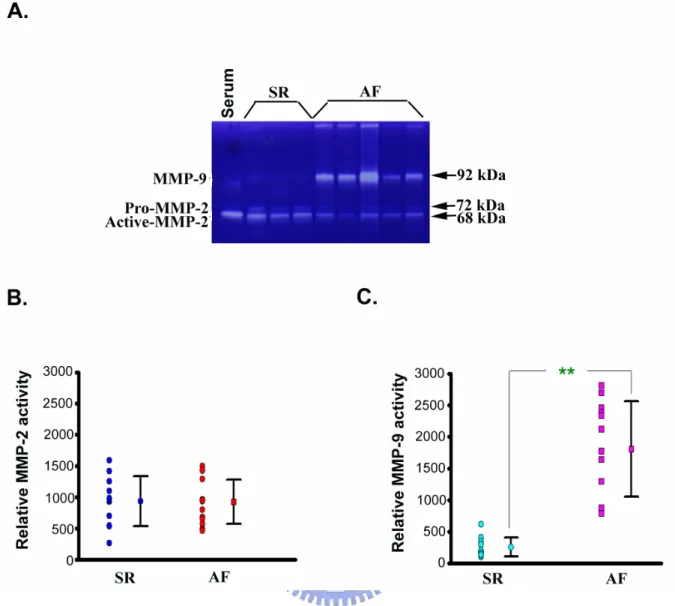
3-1-3. Enzyme activities of MMP-2 and MMP-9 in the fibrillating atria
The atria tissue isolated from hearts with AF (n = 14) and SR (n = 12) were used to detect the enzyme activity of two gelatinases, MMP-2 (gelatinase A; 72 kDa) and MMP-9 (gelatinase B; 92 kDa) by zymographic analysis(Figure 3A). As shown inFigure 3A, clear proteolytic bands on the gelatin zymographicy gels shows increased MMP-9 in the AF
subjects as compared with in the SR. The relative MMP-9 activity in the AF group was 1806 ± 753, ranging from 786 to 2809; the relative MMP-9 activity in the SR group was 254 ± 150, ranging from 100 to 627(Figure 3B). The result showed that MMP-9 activity in the AF was higher than that in the SR by 7.1 folds (p < 0.01). In the MMP-2, relative activity of this enzyme in the AF group was 923 ± 356, ranging from 496 to 1586); the relative MMP-2 activity in the SR group was 932 ± 402, ranging from 258 to 1498(Figure 3C). The enzyme activity of MMP-2 in the subjects with of with or without AF was insiginificantly different.
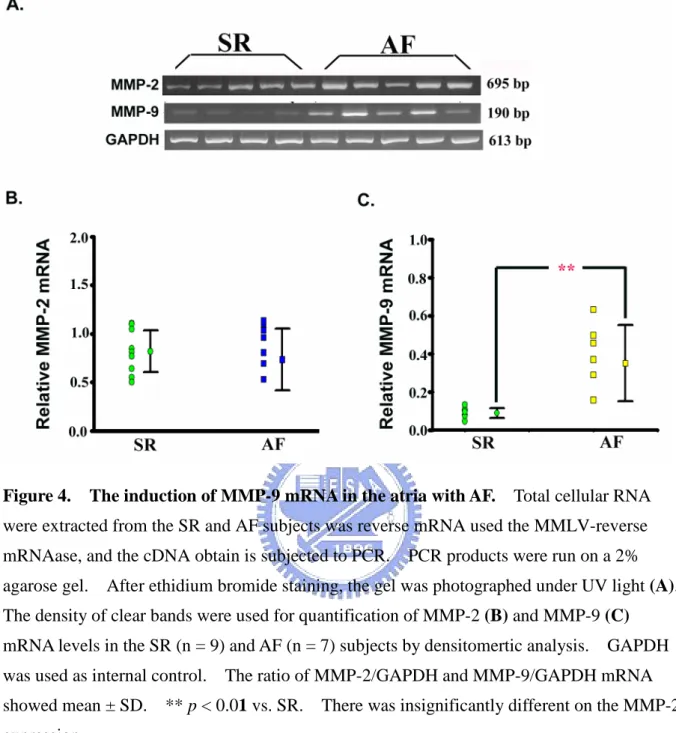
3-1-4. MMP-2 and MMP-9 mRNA levels in AF
According the results of Zymography, the markedly MMP-9 activity was observed in the fibrillating atria. Further, we were interesting to measure the MMP-2 and MMP-9 mRNA levels in the atria, because MMP family was a major protease in the ECM metabolism. The transcript (i.e., mRNA) of MMP-2 and MMP-9 in the atrial tissues with AF and SR were determined by the method of semiquantitative RT-PCR(Figure 4A). In this analysis, the expression of GAPDH mRNA was used as internal control and the relative MMP mRNA was represented as MMP/GAPDH mRNA detected. The relative mRNA level of MMP-9 in the AF group was 0.43 ± 0.18 (ranging from 0.16 to 0.49) and in the SR group was 0.0.10 ± 0.03 (ranging from 0.05 to 0.13)(Figure 4B). The results showed that the relative mRNA level of MMP-2 was 0.90 ± 0.22 (ranging from 0.53 to 1.13) in the AF group and 0.82 ± 0.23 (ranging from 0.50 to 1.11) in the SR group(Figure 4C). The statistical result shows that MMP-9 mRNA level in the AF was higher than that in the SR by 4.3 folds ( p < 0.01); but mRNA level of MMP-2 in the SR was not significantly different.
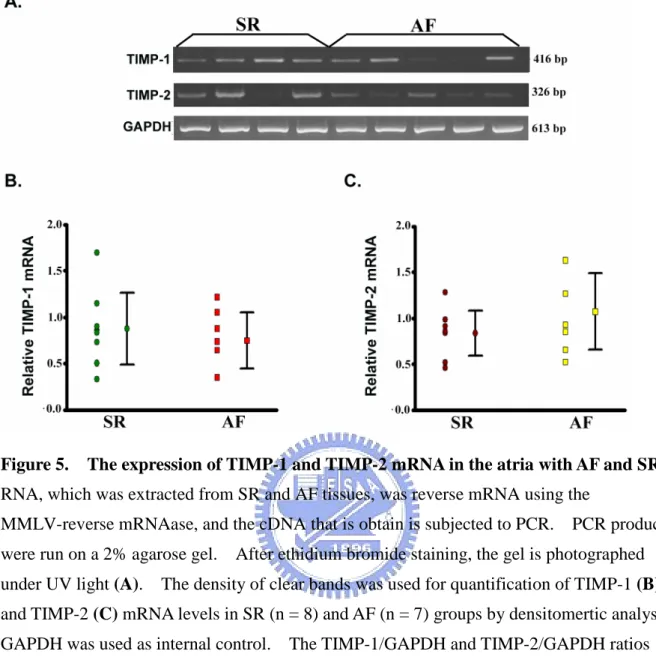
3-1-5. TIMP-1 and TIMP-2 mRNA levels in AF
Several studies have demonstrated the pathological effects of both MMPs and TIMPs in cardiovascular diseases involved vascular remodeling and cardiac remodeling in congestive heart failure or myocardial infarction[Hijova, 2005]. Therefore, the relative abundances of TIMP-1 and TIMP-2 in the atrial tissues were also determined by semiquantitative RT-PCR analysis(Figure 5A). In this analysis, the expression of GAPDH mRNA was used as internal control and the relative TIMP mRNA was represented as TIMP/GAPDH mRNA detected. The relative mRNA level of TIMP-1 in the AF group was 0.75 ± 0.33 (ranging from 0.35 to 1.22) and in the SR group was 0.88 ± 0.41 (ranging from 0.35 to 1.70)(Figure
5B). The results showed that the relative mRNA level of TIMP-2 was 1.07 ± 0.45 (ranging from 0.52 to 1.63) in the AF group and 0.84 ± 0.26 (ranging from 0.52 to 1.28) in the SR group(Figure 5C). There were insignificantly different between the two groups on TIMP-1 and TIMP-2.
3-2. MMP-2 expression in cardiac fibroblasts treated with Ang II
and Ang-(1-7)
Alterations in the normally circulating Ang II and/or Ang-(1-7) concentrations and the ratio of Ang II and Ang-(1-7) might reflected cardiac remodeling in response to physiologic stresses[Schupp et al., 2005]. We were interesting to understand the effects on the MMPs and TIMPs expression in cardiovascular system under abnormal conditions of Ang II and Ang-(1-7). In the following experiments, a cardiac fibroblast cell line H9c2 was used to treat with serious concentrations (10-9, 10-7and 10-5M) of Ang II and Ang-(1-7), respectively. The mRNA levels of collagen type I, MMP-2, MMP-9, TIMP-1 and TIMP-2, and enzyme activity of MMP-2 as well as MMP-9 in the angiotensin peptides treated cells were detected and compared.
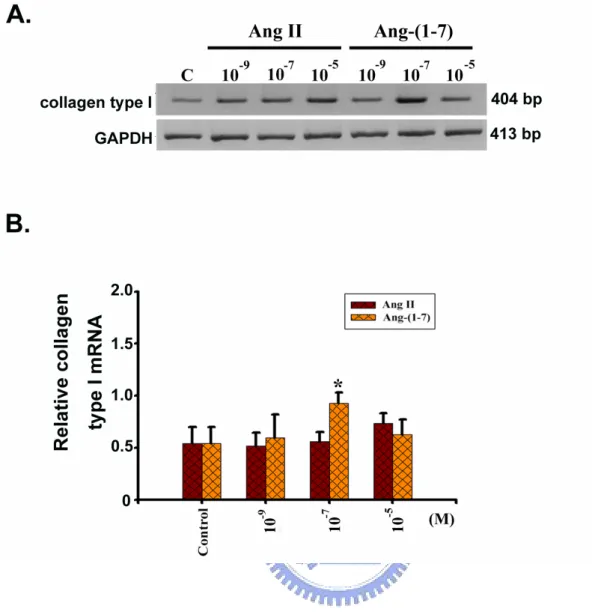
3-2-1. Collagen typeⅠmRNA in the H9c2 treated with Ang II and Ang-(1-7)
The relative collagen type I expression in control, 10-9, 10-7and 10-5M of Ang II treated H9c2 cells was 0.54 ± 0.16, 0.52 ± 0.12, 0.56 ± 0.08, and 0.73 ± 0.10, respectively. In the same condition, the cells were treated with control, 10-9, 10-7and 10-5M of Ang(1-7) which was 0.54 ± 0.16, 0.59 ± 0.22, 0.93 ± 0.10 and 0.63 ± 0.14, respectively(Figure 7A). In this analysis, the expression of GAPDH mRNA was used as internal control and the relative collagen type I mRNA was represented as collagen type I/GAPDH mRNA detected. The
significant data showed 0.92 ± 0.10 in 10-7M of Ang-(1-7); but, the other concentrations were insignificantly different, whether Ang II or Ang-(1-7) treatments(Figure 7B).
3-2-2. MMP-2 activity in H9c2 cells treated with Ang
Ⅱ
and Ang
-(1-7)
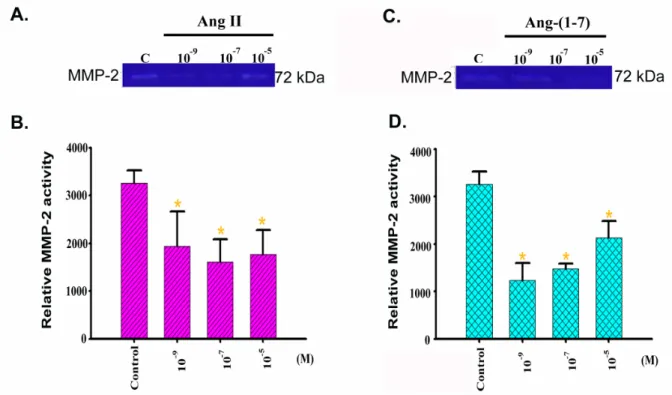
The condition medium isolated from Ang II and Ang-(1-7)-treated in H9c2 cells were used to detect the enzyme activity of gelatinase, MMP-2 (gelatinase A; 72 kDa) by
zymographic analysis(Figure 8A, C). The relative MMP-2 activity in control, 10-9, 10-7and 10-5M of Ang II were 3257 ± 184, 1943 ± 512, 1603 ± 334, and 1737 ± 357, respectively. In the same condition, the cells were treated with control, 10-9, 10-7and 10-5M of Ang-(1-7) which was 3257 ± 184, 1219 ± 261, 1477 ± 73 and 2140 ± 247, respectively. The enzyme activities were significantly decreased in all concentrations of Ang II(Figure 8B)and Ang-(1-7)(Figure 8D); in Ang-(1-7)-treated, the enzyme activities had dose-dependent, but not in Ang II-treated.
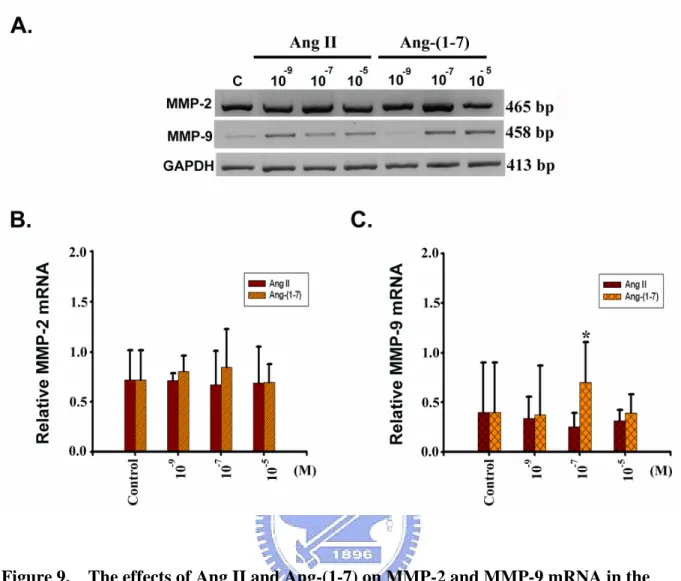
3-2-3. MMPs and TIMPs mRNA levels in the H9c2 cells treated with Ang Ⅱ
and Ang-(1-7)
The relative MMP-2 expression in control, 10-9, 10-7and 10-5M of Ang II treated H9c2 cells was 0.72 ± 0.29, 0.71 ± 0.07, 0.67 ± 0.34, and 0.69 ± 0.36, respectively; in MMP-9 expression, the values were 0.40 ± 0.20, 0.34 ± 0.22, 0.25 ± 0.14, and 0.32 ± 0.11,
respectively; in TIMP-1 expression, the values were 0.81 ± 0.23, 0.88 ± 0.15, 0.96 ± 0.17, and 0.94 ± 0.20, respectively; in TIMP2 expression, the values were 0.80 ± 0.35, 0.80 ± 0.20, 0.92 ± 0.15, and 0.94 ± 0.12, respectively. In the same condition, the MMP-2 activity in cells treated with control, 10-9, 10-7and 10-5M of Ang(1-7) which was 0.72 ± 0.29, 0.80 ± 0.16, 0.85 ± 0.38, and 0.69 ± 0.18, respectively; in MMP-9 expression, the values were 0.40 ± 0.20,
0.38 ± 0.49, 0.70 ± 0.40 and 0.39 ± 0.19, respectively; in TIMP-1 expression, the values were 0.81 ± 0.23, 0.93 ± 0.14, 0.80 ± 0.10 and 0.77 ± 0.03, respectively; in the TIMP-2 expression, the values were, 0.80 ± 0.35, 0.60 ± 0.22, 0.93 ± 0.10 and 0.63 ± 0.14, respectively. In this analysis, the expression of GAPDH mRNA was used as internal control and the relative MMP-2, MMP-9, TIMP-1 and TIMP-2 mRNA were represented as MMP-2/GAPDH,
MMP-9/GAPDH(Figure 9), TIMP-1/GAPDH and TIMP-2/GAPDH(Figure 10)mRNA
detected. The relative MMP-9 expression was significant by 10-7M of Ang-(1-7)-treated (Figure 9C). There were no significantly different between the three groups on MMP-2 (Figure 9B), TIMP-1(Figure 10B)and TIMP-2(Figure 10C).
3-3. MMP-2 expression in the different tissues
The MMPs and TIMPs had until been distributed the quantity in any tissues of rat. The balance of MMPs and TIMPs was demonstrated relative of cardiac diseases. We were interesting the study, we collected different source tissues in rat and analyzed the content of MMP-2, MMP-9, TIMP-1 and TIMP-2 mRNA, enzyme activity of MMP-2 and MMP-9.
The activity of MMP-2 and MMP-9 in the different tissues, including heart, liver, spleen, lung, kidney, brain, muscle, intestine and testis, were detected by gelatin zymography(Figure 13A). The actual activity of MMP-2 was estimated via MMP-2 standard curve(Figure 12). The activity MMP-2 in the heart, lung, muscle and testis was 10475 ± 7454, 14444 ± 8918, 13782 ± 10288 and 6993 ± 4483, respectively(Figure 13B). We noted the conspicuous gelatinolytic activity in the heart, lung, muscle and testis as compared with others. However, the activity of MMP-9 in the several tissues of rat was undetectable.
The mRNA of MMP-2 was measured in the different tissues of rat with heart, liver, spleen, lung, kidney, brain, muscle, intestine and testis, were detected by the method of
semiquantitative RT-PCR. In this analysis, the expression of 18s RNA mRNA was used as internal control and the relative MMP-2 mRNA was respresented as MMP-2/18s RNA mRNA detected(Figure 14A). The values were that a markedly high expression in heart (5.15 ± 1.06), lung (5.05 ± 0.95) and muscle (8.02 ± 0.79) was detected as compared with those in the others(Figure 14B).
3-4. Ratios of MMP2/TIMP-2 and MMP-9/TIMP-2
MMPs and TIMPs played a crucial role in physiological and pathological matrix turnover[Moche et al., 2005]. Inhibition of MMPs or targeted deletion of MMP gene attenuated cardiac remodeling. Correlation between MMP-9 and TIMP-1 (MMP-9/TIMP-1) and MMP-2 and TIMP-2 (MMP-2/TIMP-2) can be estimated and applied as a physiological marker for diagnosis of disease processing.
The ratio of MMP-9/TIMP-1 showed positive correlation with mRNA levels in the AF group (0.32 ± 0.23, ranging from 0.10 to 0.75) compared with the SR group (0.11 ± 0.05, ranging from 0.05 to 0.19). The result showed that ratio of MMP-9/TIMP-1 in the AF was higher than that in the SR by 2.9 folds (p < 0.05). In contrast, the ratio of MMP-2/TIMP-2 was insignificantly different(Figure 6).
Similarly, the ratio of MMP-9/TIMP-1 in 10-7M Ang-(1-7)-treated showed positive correlation with mRNA levels control (0.70 ± 0.40). The results showed that ratio of MMP-9/TIMP-1 treated 10-7M of Ang-(1-7) in H9c2 cells was higher than that in the control by 1.8 folds (p < 0.05). In contrast, the ratio of other concentration with Ang II or Ang-(1-7)-treated was insignificantly different(Figure 11).
IV. Discussion
The major finding of this study was that the process of fibrillation follow AF might regulate MMP-9 mRNA through cardiac fibroblast activation. In constant, cardiac
fibroblasts were associated with, high levels of active MMPs. This raises the possibility that an in vivo interaction between the fibrillar collagen network and cardiac fibroblasts works as a “motor” to generate active MMPs, which may, in turn, be involved in myocardical ECM turnover. A point of view about cardiac fibroblasts play an important role to regulate the ECM remodeling has been a focus of increasing recent investigation. Accurate
understanding the molecular mechanism of ECM remodeling would help make possible the development of successful therapeutics strategies during the AF.
In this article, we had successful established the sustained AF model by pacing-induced, the myocardial fibrosis markedly accumulated in interstitial cardiomyocytes in the AF by Masson’s trichrome staining(Figure 1). These results were corresponsed to Masson’s trichrome staining. First at all, Western blot demonstrated the increased protein levels of collagen typeⅠinthe AF(Figure 2). The values indicated that MMP-9 activity showed different between SR and AF and the quantitative values demonstrated that the increase of MMP-9 activity was more pronounced in the AF than SR. In fact, the zymographic activity of MMP-9 increased in the AF was reported here for the first time. These AF had benefits to imitate actual human pathological AF symptom compared with other animal models.
Several lines of evidence point to the importance of MMP-2 and MMP-9 in cardiac
remodeling after myocardial infarction[Lu et al., 2000; Spinale, 2002; Spinale et al., 2000], as well as in vascular remodeling after injury and during atherogenesis[Johnson andGalis, 2004; Whatling et al., 2004]. Therefore, we presented a useful detection method that would be applied to follow the process of fibrosis in the AF atria by MMP-9 activity.
AF is a characteristic feature of heart failure and there is evidence that an ECM imbalance between levels of MMP-9 and TIMP-1 was associated with cardiac remodeling
[Hornebeck et al., 2005; Moshal et al., 2005]. Increased cardiac expression of TIMP-1 and TIMP-2 were related to cardiac fibrosis and dysfunction in the chronic pressure-overloaded human heart [Heymans et al., 2005]. In accordance with the documents, we further analyzed the MMPs and TIMPs mRNA levels in the AF. As shown inFigure 3 and Figure 4,the MMP-9 mRNA levels were corresponded with MMP-9 activity. Although the mRNA levels of TIMP-1 and TIMP-2 were down-regulated in the AF, however, the trends of reduction did not reach statistical significance (p > 0.05)(Figure 5). These results seem to correspond to the induction of MMP-9 mRNA levels and zymographic activity. Besider, TIMP-1 would down regulated MMP-9 expression in different tissues. To summarize these data, we supposed that the imbalance between MMP-9 and TIMP-1 mRNA levels in the AF were reached statistical significance in the AF as compare with SR (p < 0.01). This imbalance ratio of MMP-1/TIMP-1 might reveal the tendency toward fibrillation.
In the application of genome research, we would get the whole human genome sequence easily and application to molecular biology. But, we had problems with got the whole pig genome sequence. Therefore, we capitalize on human and rat genome sequence to design MMP-9 and MMP-2 primers for pig. Because, there were almost 70% conserved sequences on MMP-9 and MMP-2 between human and rat. After RT-PCR analysis, the specific product of MMP-9 and MMP-2 were cloned using the TA cloning kit and sequenced. After automated sequencing, comparative analyses were performed with the human MMP-9 and MMP-2 mRNA sequence and demonstrate that the conserve sequence of MMP-9 and MMP-2 from pig compare with human would reach to 70%. Cardiac fibroblasts were the most abundant cell type in the heart and play a major role in synthesizing components of the
cardiac ECM[Eghbali, 1992]. Cardiac fibroblasts had also been shown to play an important
role in the regulation of collagen degradation by MMPs[Cleutjens et al., 1995]. We used the culture systems to mechanically stimulate cardiac fibroblasts allows for the systematic
examination of mechanical regulation of cardiac fibroblast function in the absence of systemic effects. In the present study, the rat cardiac fibroblast H9c2 cells were selected as an in vitro cell line model to investigate what was an important effects in the process of fibrosis in AF. In fact, the primary cultured of cardiac fibroblasts from AF atria should be selected as an in vitro cell line model; however, the whole pig genome was difficult to get. For the reasons, rat cardiac fibroblast H9c2 cells were selected and challenged with Ang II and Ang-(1-7) for 12 and 24 h. As shown inFigure 7, the critical step of fibrosis was based on the induction of collagen type I mRNA levels. The values indicate that Ang II and Ang-(1-7) induced the collagen type I mRNA levels significant in a dose-dependent manner, especially in the dosage of 10-7M of Ang-(1-7) which had significantly different than control (p < 0.05). It was an interesting finding that Ang-(1-7) would induce collagen type I mRNA levels. However, the regulate mechanism of Ang-(1-7) on collagen type I induction was still unknown. After the H9c2 cells challenged with Ang II and Ang-(1-7), we anticipated that the MMP-2 and
MMP-9 mRNA levels would corresponded with collagen type I induction. However, it was not anticipated that the signal ratios of MMP-2/GAPDH, TIMP-1/GAPDH and
TIMP-2/GAPDH were not increased in dose-dependent manner after Ang II and Ang-(1-7) treatment, respectively(Figure 9 and 10). In contrast, the MMP-9 mRNA levels were induced in the dosage of 10-7M of Ang-(1-7) which had significantly different than control (p < 0.05). But, the mRNA levels of MMP-9 in H9c2 cells were too weak to be detected by RT-PCR. Then we even tried to amplify the MMP-9 gene expression for 38 cycles by RT-PCR. Here, the difference from MMP-9 mRNA levels and zymographic activity between cardiac fibroblast H9c2 cells and AF model, we though about the species different between rat and pig.
The effects of MMPs were determined in different tissues of rat. We further studied the MMP-2 zymographic activity in different tissues of rat by gelatin zymography. In this study, MMP-2 zymographic activity was significantly greater in heart, lung and muscle as compared with other tissues. The induction of MMP-2 zymographic activity in heart, lung and muscle than other tissues was a novel observation. A recent study showed that ECM protein
remodeling enhances MMP-2 enzyme stability as well as its activity[Itoh et al., 1998]. Taken together, we thought that certain tissues had high level of MMPs zymographic activity would trend to develop fibrosis after tissue injury or damaged. Several previous rat studies had reported that the high level of MMP-2 was detected in common, but might be predict to increase during cardiac remodeling. Weber et al.[Weber et al., 1992]reported that the severity of cardiac fibrosis might become significantly apparent with the development of remodeling by increasing MMP activity. That was, once the heart was damaged, ECM that connected cardiomyocytes would be degenerated by increased MMP for the adaptation of cardiomyocytes enlargement and fibroblasts invasion. The similar results were observed in lung and muscle, too[Cheng et al., 2005; Hsu et al., 2005].
The MMP-2 mRNA levels were only induced in heart, lung and muscle based on the different mRNA levels between MMP-2 in multiple tissues of rat(Figure 13). The result suggested the development of heart failure characterized by the unusual expression of MMP-2 in the fibrosis. We believe that pathologic cardiac dysfunction can be predicted to occur if the experiment period was prolonged. In fact, cardiac expression of TIMP-1 and TIMP-2 was significantly increased in chronic pressure-overloaded human hearts compared with controls and is related to the degree of interstitial fibrosis[Heymans et al., 2005]. We supposed that the MMPs zymographic activity was as manifest cardiac dysfunction marker.
End-stage of human dilated cardiomyopathy (DCM) was characterized by myocytes loss and fibrosis, and associated with ventricular dilatation and reduced cardiac function. MMPs
and TIMPs have been involved in the myocardial remodeling[Rouet-Benzineb et al., 1999]. In the AF model, the imblance of MMP-9/TIMP-1 was compared with the SR (p < 0.05). The increased ratio of MMP-9/TIMP-1 associated with the process of fibrosis during fibrillating atria(Figure 6). Recently studies showed that the imbalance between
macrophage-derived MMP-9 and TIMP-1 in bleomycin-induced pulmonary fibrosis in mice
[Li et al., 2004]. In fact, the imbalance between MMP-9 and TIMP-1 in AF might be reported here for the first time. Furthermore, we would through the selected inhibitors of MMP-9 RNAi and chemical agent treatment for the correlation between MMP-9 and TIMP-1 in the AF was more clear. In the previous studies, Ang II mediated fibroblast function and collagen production over a 72-h period, while increasing MMP-2 expression and activity
[Mookerjee et al., 2005]. But, there were not significant induction of MMPs mRNA and activity in H9c2 cells. Although the ratio of MMP-9/TIMP-1 had significantly different (p < 0.05) on the concentration of 10-7M of Ang-(1-7) treated-cells(Figure 11). But, it was stranged in the induction of MMP-9 mRNA levels by single dosage of Ang-(1-7) treatment. We speculated that the H9c2 cells were not sensitive to Ang II and Ang-(1-7) treatment.
Our findings in the presentation studies include the induction of MMP-9 mRNA levels, zymographic activity and imbalance of MMP-9/TIMP-1 ratio were predicted what an important role of MMP-9 during the ECM remodeling in AF. They might be, at least partially, the possible molecular mechanisms behind how regulated the ECM remodeling during the increased risk of AF reported in pig. Particularly, because cardiac fibrosis was a more typical end-stage condition, it is more beneficial to alleviate these problems before the end-stage is reached. Therefore, it would seem appropriate to follow AF when considering possible agents that might be helpful to control the development of fibrosis-related cardiac diseases.