~Jjj,j,pJf~ Fish Pathology, 39 (1),15-23,2004.3 © 2004 The Japanese Society of Fish Pathology
Molecular Cloning and Recombinant Expression of
Tiger Shrimp Penaeus monodon Penaeidin
Shih-Hu Ho
1,Yu-Chan Chao
2,Hsiao-Wei Tsao
3,Masahiro Sakai
4,Hong-Nong Chou
5and Yen-Ling Song
1 ,3*1Institute of Zoology, National Taiwan University, Taipei, Taiwan, 106, Republic of China 21nstitute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan, 115,
Republic of China
30
epartment of Life Science, National Taiwan University, Taipei, Taiwan, 106, Republic of China4Faculty of Agriculture, Miyazaki University, Miyazaki 889-2192, Japan
51nstitute of Fisheries Science, National Taiwan University, Taipei, Taiwan, 106, Republic of China
(Received August 22, 2003)
ABSTRACT-A penaeidin cDNA sequence of tiger shrimp Penaeus monodon was obtained. In the sequence, an open reading frame that coded for a peptide composed of 74 amino acids was found. A cleavage site of secretory signal peptide was predicted between amino acids 19 and 20. The calculated molecular mass of mature penaeidin was about 6.1 kDa and the estimated pi of this peptide was 9.1. Northern blot analysis indicated that the penaeidin was mainly synthe-sized in the hemocytes. Clustering analysis of penaeidin sequences from P. monodon, P. vannamei, P.setiferus,P.japonicus andP. chinensis was performed. The recombinant penaeidin was expressed using insect-baculovirus expression system. The recombinant penaeidin showed antimicrobial activity against a bacterium Aerococcus viridans, but not Vibrio alginolyticus, Vibrio
harveyi or a yeast Debaryomyces hansenii. In addition, it delayed spore germination and growth of a filamentous fungus Neurospora crassa.
Key words: penaeidin, antimicrobial peptide, Penaeus monodon, shrimp hemocyte, insect-baculovirus expression system, antimicrobial activity.
Animal peptides with antibiotic properties are wide-spread in nature, present in mammals (Lee et al., 1989),
amphibians (Zasloff, 1987), fish (Cole et al., 1997), in-sects (Boman, 1991) and crustaceans (Destoumieux et
al., 1997). These peptides are major constituents of the innate immune system for non-specific defense of the host against microbial invaders. The innate system can be rapidly activated after injury on or invasion of the host by microbial agents. Host defense is done by combat-ing the proliferation of the pathogens immediately after infection. Antimicrobial peptides thus provide an impor-tant defense mechanism in animals as the first line of host defense.
Penaeidins are 5.5 to 6.6 kDa antimicrobial peptides isolated from the plasma and hemocytes of Pacific white
*Corresponding author E-mail: [email protected]
shrimp Penaeus vannamei. They are composed of a proline-rich N-terminus and a C-terminus containing six cysteine residues engaged in three disulfide bridges (Destoumieux et al., 1997; Destoumieux et al., 2000a). Penaeidin-2 and -3 exhibit both Gram-positive antibacte-rial and antifungal activities (Destoumieux et al., 1999). Penaeidins are constitutively synthesized and stored in the shrimp hemocytes and localized in granulo-cyte-cytoplasmic granules, and are released in response to appropriate stimuli such as infection (Destoumieux et
al., 2000a; Munoz et al., 2002).
A sequence similar to 3' part of penaeidin-3 of P.
vannamei (Destoumieux et al., 1997), which we termed
here as Ho-6 clone, was found when we sequenced cDNA library to search for defense-related genes from hemocytes of tiger shrimp (P. monodon) , an important cultured shrimp species in Taiwan and southeastern Asia. We also obtained the complete open reading
frame sequence of tiger shrimp penaeidin using PCR with specific primers.
This study presents the cDNA cloning data of tiger shrimp penaeidin and compared its sequence and prop-erties with other known penaeidins. To confirm the antimicrobial activity of the product of the open reading frame, we expressed the recombinant penaeidin using insect-baculovirus expression system. We could assume that the post-translation modification and protein folding of the recombinant penaeidin would be similar to native shrimp penaeidin because both insect and shrimp belong to Arthropoda. The antimicrobial activity of the recombinant penaeidin was likewise evaluated.
Materials and Methods
Construction of Hemocyte cONA Library and Cloning for Penaeidin
Shrimp (P. monodon) was obtained from shrimp farms in Taiwan. Before getting hemolymph, the shrimps were immersed in 0.5 mg/mL f3-glucan (from
Saccharomyces cerevisiae, Biotec Mackzymal, Norway)
for 3 h. Hemolymph from shrimp was collected using a syringe containing anticoagulant (0.1 M sodium citrate,
0.4 Msucrose, 0.01 MTris-HCI, pH 7.6, 780 mOsm/kg)
and centrifuged at 700 x
g
for 5 min. The resulting hemocyte pellet was then harvested. Total RNA was extracted and further purified by using a QuickPrep Micro mRNA Purification Kit (Pharmacia, USA). SuperScript Plasmid System (Gibco BRL, USA) was used for cDNA synthesis and plasmid cloning. The vector-ligated cDNA was introduced by transformation into E. coli cells (MAX Efficiency DH10Bac CompetentCells, Gibco BRL).
An Ho-6 clone sequence was found similar to 3' part of penaeidin-3 of P. vannamei. To get the 5' part se-quence of penaeidin, a primer 5'-CCTACATCCTTT-CCACAAGC-3' (corresponding to nucleotides 333-352 shown in Fig. 1) was designed and used as the reverse primer in the polymerase chain reaction (PCR). The vector-located T7 primer served as the forward primer, and the cDNA library plasmid mixture was the template. Twenty-five cycles were used, consisting of 1 min at 94°C, 1 min at 55°C and 1 min at 72°C. To con-firm the 3' part sequence of penaeidin, a primer 5'-(AGC)G(AGCT)CCG(AGT)T(ACT)CCCAGGCCACC-3' (corresponding to nucleotide 85-104 shown in Fig. 1) was used as forward primer in PCR. The vector-located SP6 primer served as the reverse primer. The bands of the PCR product on electrophoresis gel were eluted then were ligated to pUC-T vector (MDBio Inc., Taiwan). Every ligated vector was subsequently introduced to E. coli DH-5
a
competent cells to amplify the vector. At least 3 clones were sent to the commer-cial company (Mission Biotech, Taiwan) for sequencing. The sample strand (100 ng) was used in a cyclesequencing reaction using dye-nucleotide terminators (ABI PRISM Big Dye Terminator Cycle Sequencing Ready Reaction Kit, Applied Biosystems, USA) with a 5 pmole primer as sequencing primer. Sequencing prod-ucts were purified using ethanol precipitation procedure (Applied Biosystems protocol) and sequenced on an ABI PRISM 377-96 Sequencer (Applied Biosystems, USA).
Northern Blot Analysis
Total RNA was extracted from P. monodon tissues and hemocytes using Trizol reagent (Gibco BRL). Ethidium bromide was added into the RNA preparations and the RNA Millennium Markers (Ambion, USA). The samples were then run on 1.2% agarose gel at 50V in the presence of formaldehyde. After electrophoresis, a photography picture of the gel was taken under UV light. The RNA was then transferred to an Immobilon-Ny+ membrane (Millipore, USA) by capillary blotting overnight according to the manufacturer's instructions, followed by UV-crosslinking of the blotted membrane.
A DIG-Iabeled penaeidin-specific 360 bp-probe was amplified by PCR on a cDNA clone using 5'-AAGGCCTTGCCATGCGTCTCGTGGTCTG-3' as the forward primer and 5'-CCTACATCCTTTCCACAAGC-3' primer as the reverse primer. Thirty cycles were used, consisting of 45 s at 94°C, 1 min at 56°C and 2 min at 72°C, and using DIG DNA labeling Mix (Roche, Switzer-land) as dNTP substrate.
After hybridization at 65°C overnight, the membrane was washed twice with 2 x SSC, 0.1 % SOS at room tem-perature and two times with 0.1 x SSC, 0.1 % SOS at 50°C. Then the target bands on the membrane were immunologically detected using DIG Nucleic Acid Detec-tion Kit (Roche) and COP-Star chemiluminescent sub-strates (Applied Biosystems) exposed to BioMax MS Film (Kodak, USA).
Analysis of Nucleotide and Amino Acid Sequences
The nucleotide and the deduced amino acid sequences of the shrimp penaeidin cDNA were analyzed using Wisconsin Package Version 10.2, Genetics Com-puter Group (GCG), Madison, Wise. and NCBI BLAST program. The amino acid sequences of all penaeidin clones were retrieved from the NCBI Entrez and aligned by the DAMBE (Data Analysis in Molecular Biology and Evolution, version 4.0.75) software package (Xia, 2000; Xia and Xie, 2001). The multiple sequence alignment was created with the ClustalW. Phylogenetic and molecular evolutionary analyses were conducted using MEGA version 2.1 (Kumar et al., 2001) and the tree was constructed based on the neighbor-joining method (Nei and Kumar, 2000).
Production of Recombinant Penaeidin
Penaeidin peptide was produced by a recombinant baculovirus system. The Spodoptera frugiperda Sf21
Cloning and expression of P. monodon penaeidin
5'
GCCATGCGTCfCGTGGTCfGCCfGGTCITCCfGGCCfCCITCGCCCfGGTCfGCCAAGCC 60
M R L V veL V F L A S F A L V C
Q
At19
CAAGGGTACCAGGGTGGTIACACACGCCCGTICCCCAGACCACccrATGGGGGAGGATAT 120
Q
G Y Q G G Y T R P F P R P P Y G G G Y 39
CATCCAGTICCTGITfGCACITCATGCCACAGGCITAGCCCCITACAAGCfCGTGCITGC 180
H P V PVC T S C H R L S P L Q A RAC 59
TGCAGGCAGTIAGGACGTIGTIGTGATGCAAAGCAGACATATGGTTGATGGAGAAGACAA 240
C R
Q
L G R C C D A K
Q
T Y G
*
74
CGAAAAACfGACITCACAATGTATTAATCAGTTGTGAAGAAAGTGCAACCCTGATITIGA 300
ACTGTATITICfAGTTCCATITICITACITITGCITGTGGAAAGGATGTAGGTAITfGGA 360
TITICCATGAATGTATGATGAATGAAAGTGCATGTGGGATGTATGTGCATACAGTCGTAT 420
TIGTCCCAGCAGGTCCTCGTGTATTCACAGGAGAAAGATATCGTGTTGTTTGACITfCGT 480
TGTAGTTATTTGTAGGTATGGGTCfGTGTGTGGTIGGTGTTTGCATAITfCCCAAAGGAC 540
ATTCGGAATTGTACfACfCfCITACAMIMMTTGATATCTGTGAAAAAAAAAAAAAAA 600
AAA
Fig. 1. Nucleotide and deduced amino acid sequence of Penaeus monodon penaeidin. The nucleotide sequence is numbered from the first base at the 5' end. The first methionine (M) is numbered on the first deduced amino acid. The asterisk indicates the stop codon. The double-headed arrow indicates the predicted cleavage site of secretory signal peptide. AATAAA is a polyadenylation signal. The underlines mean the primers used in this study.
cell line was cultured in TNM-FH insect medium contain-ing 8% heat-inactivated fetal bovine serum. The cells were incubated at 26°C. 8f21 monolayers were used for the propagation of recombinant Autographa
californica multiple nuclearpolyhedrovirus (AcMNPV).
For transfection, Lipofectin (Life Technologes, U8A) was used.
FlankingStuI and8g/11 restriction sites (in italic) for cloning of penaeidin cDNA into the transfer vector were introduced by PCR mutagenesis with a sense primer
(5'-AAGGCCTTGCCATGCGTCTCGTGGTCTG-3') and an
antisense primer (5'
-AGATCTTCAGTGGTGGTGGTGG-TGGTGACCATATGTCTGCTTTGGAT-3'). Penaeidin cDNA fragments that contained the translation start codon ATG, entire coding region of penaeidin (including the signal sequence), 6xHis tag codon, stop codon, and
StuI and 8g/11 restriction sites at 5' and 3' ends of the insert were cloned into the pABhRpX transfer vector and introduced into Escherichia coliDH-5
a.
The correct orientation and sequence of the penaeidin insert were verified by sequencing. After cotransfection of thepABhRpX-penaeidin transfer plasmid and BaculoGold Linearized Baculovirus DNA (BD) into insect 8f21 cells, we selected viable recombinant baculovirus clones that performed fluorescence of DsRed.
Recombinant penaeidin was generated from 8f21 insect cells infected with recombinant baculovirus. For large scale expression, 8f21 cells were grown in 75 cm2 T-flasks. 8f21 monolayers were infected at a multiplic-ity of infection between 3 and 10. The cells were kept in an incubator at 26°C and harvested 3 days post-infec-tion.
Harvested infected 8f21 cells (1.5 x 108 cells) were
lysed in 40 mL of lysis buffer (50 mM NaH2P04 ,300 mM
NaGI, 10 mM imidazole, pH 8.0, supplemented with 1% Igepal CA-630) and incubated for 10 min on ice. The lysate was centrifuged at 10,000xg for 10 min at 4°C and the supernatant was collected. Then about 2 mL of 50% Ni-NTA Agarose (Qiagen, Germany) slurry was added into the 40 mL supernatant and was gently mixed by shaking at 4°G for 1.5 h. The mixture was centri-fuged at 500xg for 5 min at 4°C. The agarose pellet
Table 1. Effect of the recombinant penaeidin on the growth of some pathogens of aquatic organisms using radial diffusion assay. Area of inhibition zone (mm2
)
Species Strain Source Recombinant Cecropin
penaeidina P1 b Lysozyme
C
Gram-positive bacteria
Aerococcus viridans ATCC 11563 Air sample 11.64 5.50 178.05
A4icrococcusluteus ATCC 9341 0 89.26 86.24
Gram-negative bacteria
Vibrio alginolyticus Val Diseased shrimp (Penaeus monodon) 0 36.14 0
V. harveyi 13b Diseased shrimp (P. monodon) 0 19.33 0
Yeast
Debaryomyces hansenii QF87-25 Giant freshwater prawn (Macrobrachium rosenbergil) 0 19.76 52 Filamentous Fungus
Neurospora crassa ATCC 14692 15.62d
0 22.42
Area of inhibition zone
=
Jrr -
area of the well. r is the measured radius.aRecombinant penaeidin, 30 f.1M; bporcine synthetic cecropin, 30 f.1M; cpurified from chicken egg white, 30 f.1M. Four f.1L sample was
loaded in each well.
dApplying 77 f.1M recombinant penaeidin, the inhibited zone had fewer fungal growth than the surrounding area.
was added with 8 mL wash-buffer (50 mM NaH2P04 ,300
mM NaCI, 50 mM imidazole, pH 8.0) and loaded on a column. Then the non-specifically bound proteins were removed by washing twice with 8 mL wash buffer. Elution of 6xHis-tagged penaeidin from the Ni-NTA Aga-rose was achieved using elution buffer (50 mM NaH2P04 ,
300 mM NaCI, 250 mM imidazole, pH 8.0). The eluate containing penaeidin was desalted using Microcon YM-3 (Millipore) and stored at -20°C before antimicrobial test.
Antimicrobial Activity Test of Recombinant Penaeidin
Four bacterial strains, one yeast strain and one fila-mentous fungus were collected for the test (Table 1). The bacterial strains were grown in tryptone soya broth (TSB, Difco), except Vibrio strains which were cul-tured in the same medium but supplemented with 2.5% NaCI. Debaryomyces hansenii was grown in yeast malt
broth (YMB, Difco, USA). Neurospora crassa was
grown in potato dextrose agar (PDA, Difco). All bacte-rial and yeast strains were incubated with shaking at 28°C. Cells were prepared for testing when they grew to mid-log phase. Spores of Neurospora crassa were collected for following assays.
The diffusion method was done mainly according to Hultmark et al. (1982, 1983). Assay plates (9 cm diam-eter petri dish) were made on a leveling table from 6.8 mL of the melting culture medium agarose (1 %) and the final concentration of cultured cells was about 105
CFU/ mL. Cecropin was included as a positive control because it has been reported to inhibit the growth of bac-teria, both Gram-positive and Gram-negative (Qu et al., 1982; Lee et al., 1989, van't Hof et al., 2001). Lysozyme cleaving f3-(1A)-glycosidic bonds in the pepti-doglycan layer of Gram-positive bacterial cell wall was used as antimicrobial peptide control. Wells 3 mm in diameter were punched on the harden assay
plates. Four,uL recombinant penaeidin (30 ,uM), cecropin P1 (30 ,uM, synth!3tic porcine peptide, Sigma) or lysozyme (30 ,uM, purified from chicken egg white, Sigma) was loaded into wells. Plates were read after overnight incubation at 28°C using a dissecting microscope. Two diameter readings at 90° angles were taken. Positive inhibitory response was defined as the area of the clear zone being bigger than the area of the well.
Determination of minimum inhibitory concentration (MIC) values was performed by liquid growth inhibition assays as described by Destoumieux et al. (1999). Briefly, 10 ,uL aliquots from each dilution were incubated in 96-well microtiter plates, each well with 80 ,uL of fungal spores (final concentration 104 spores/mL) and 10 ,uL of water. Antifungal assays were performed in potato dextrose broth (Difco). Cultures were grown for 48 h at 25°C in the dark and in a moist chamber. Growth inhi-bition of filamentous fungi was observed microscopically after 24 h and measured at 590 nm optical density after 48 h.
Results
The Ho-6 clone contained a fragment from position 200 of the nucleotide to the poly A tail while the other clone obtained from PCR consisted of the nucleotides from position 1 to 5'-CCTACATCCTTTCCACAAGC-3' primer (position 352-332) (Fig. 1). There were 133 overlapping nucleotides that were completely identified in these two clones. A 603-bp penaeidin sequence was obtained (GenBank accession number: Banklt450249 AF475082). An open reading frame (ORF) of 225 bp and a 375 bp of 3' untranslated region containing the poly A tail were found in the sequence that coded for a peptide of 74 amino acids with a putative initiation
methionine codon (ATG) starting at position 4 and end-ing at position 226 of stop codon. A cleavage site of secretory signal peptide was predicted between amino
acid 19-20 using SPScan program of GCG. If other
post-translational modifications were excluded, the
cal-culated molecular mass of mature penaeidin was about 6.1 kDa and the estimated pi of this peptide was 9.1.
A probe was designed to cover the complete ORF and used in Northern blot analysis. A hybridization was done with RNA preparations extracted from shrimp
1
2
3
4
5
6
7
8
Penaeidin
(750 b)
18S RNA
(1900 b)
~"ti~ ~,. '"'~l ~ ~~'t;' .- • • • ~,:,~tJ':.
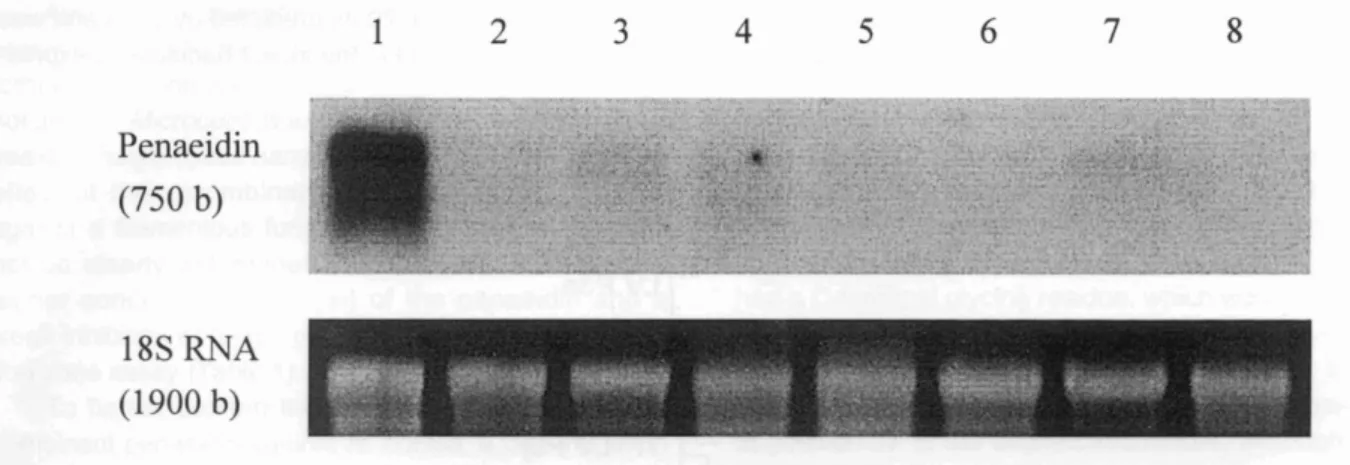
l ,-I"~ -1l.... ,,:;,~ ..._~)"....~'~~: ·~~~-':':~~~;\f: ~~1....it f .4};" ~.". t·,:;}:"'/l ~'i':' . ,1 .'~ .. ,-.Fig. 2. Northern blot analysis of total RNAs from various shrimp tissues. About 5 /1g of total RNAs each from various tissues was separated on 1.2% agarose-formaldehyde gel through electrophoresis, blotted to a Nylon membrane and hybridized with mG-labeled penaeidin probe. 18S RNAs were stained by EtBr on the gel. Lanes 1- hemocytes; 2- heart; 3- intestine; 4-gills; 5- subcuticular epithelium; 6- lymphoid organ; 7- hepatopancreas; and 8- muscle.
PM P ESTPMI2 ESTPMCU19 ESTPMI20 LV Pl LV P2b LV P2 LS P2d FC PS LV P3e LV P3i LV P3a LV P3b LV P3j LV P3f LV P3h LV P3d LV P3c LV P3q LS P3k LS P3n LS P3m LS P31 LS P4d LV P4a LV P4c MJ P2
*
20 _ . . • QGYQI'·
. . •
~GYQI"
. . •
QGYQ1'-
. . •
QGYQ*
40*
,·---PYGGGYH VPVCTS--'F ··---PYGGGYHP.VPVCTS- ··FP'·---PYGGGYHPVPVCTS- ··Fp··---TYGGGYHPVPVCTS-• P ··Fp··---TYGGGYHPVPVCTS-•··Fp··---TYGGGYHPVPVCTS-• ---;PIGRP--PLRLVVC p··---'PIGRP--PLRPVCN p··---'PIGRP--PFRPVCN p··---'PHGRP--PLGPICN s··---PYGGGYG--NVCTS-··PP 'PGGPIGPYNGCPVS ··PP 'PGGPIGPYNGRPVS P··PP 'PGGPIGPYNGCPVS PI' • PP 'PGGPIGPYNGCPVS P"P- 'PGGPIGPYNGCPVS "PP 'PGGPIGPYNGCPIS PI· • PP 'PGGPIGPYNGCPIS ··PP • PGGPIGPYNGCPIS ··P- • PGGPIGPYNGCPVS P"PP 'PGGPISPYNGCPVS S---YNACTLS S---YNACTLS S---YNA TLS S---YNVCTLS 'IFIRP----IGCDV- ·IFIRP----IGCDV-'ISRPP---SRAACYS*
80 AKQTYG- LVK---- LVKG---PRRGYSG GKGYSG GKGYSG GKGYSG VGKGYSG GKGYSG GKGYSG GKGYSG GKGYSG GKGYSG GKGYSG AKGYSG AKGYSG AKGYSG AKGYSG LG--- RG--- LVKG---74 74 74 74 50 72 72 72 71 82 82 82 82 81 82 82 82 81 82 75 75 75 75 67 67 67 63Fig. 3. Sequence comparison of 27 penaeidins amino acid or deduced amino acid sequences of their coding region. The multiple sequence alignment was created with the ClustalW. Conservative residues are in boxes. PM P: Penaeus monodon penaeidin (this study); ESTPMI2: Shrimp Lambda Zip Express LibraryP. monodon cDNA clone (GenBank Accession num-ber: 81784441); ESTPMI20: Shrimp Lambda Zip Express LibraryP. monodon cDNA clone (81784459); ESTPMCU19: Shrimp Lambda Zap Express LibraryP. monodon cDNA clone (81018089); LV P1: Penaeus vannamei penaeidin-1 (P81056); LV P2: P. vannamei penaeidin-2 (P81057); LV P2b: P. vannamei penaeidin-2b (AAK77539); LS P2d: P. setiferus penaeidin-2d (AAK83453); LV P3a: P. vannamei penaeidin-3a (P81058); LV P3b: P. vannamei penaeidin-3b (P81059); LV P3c: P. vannamei penaeidin-3c (P81060); LV P3d: P. vannamei penaeidin-3d (AAK77533); LV P3e: P. vannamei penaeidin-3e (AAK77534); LV P3f: P. vannamei penaeidin-3f (AAK77535); LV P3g: P. vannamei penaeidin-3g (AAK77536); LV P3h: P. vannamei penaeidin-3h (AAK77537); LV P3i: P. vannamei penaeidin-3i (AAK77538); LV P3j: P. vannamei penaeidin-3j (AAK73083); LS P3k: P. setiferus penaeidin-3k (AAK83450); LS P31: P. setiferus penaeidin-31 (AAK83454); LS P3m: P. setiferus penaeidin-3m (AAK83451); LS P3n: P. setiferus penaeidin-3n (AAK83452); LV P4a: P. vannamei penaeidin-4a (AAK77540); LV P4c: P. vannamei penaeidin-4c (AAK77542); LS P4d: P. setiferus penaeidin-4d (AAK83455); FC P5: P. chinensis penaeidin-5 (AAP33450); MJ P2: P. japonicus penaeidin-2 (EST data, Rojtinnakorn et al., 2002).
hemocytes, heart, intestine, gills, subcuticular epithe-lium, lymphoid organ, hepatopancreas, and muscle. A strong 750 b transcript signal was detected in the shrimp hemocytes (Fig. 2). This clearly shows that penaeidin is mainly synthesized in the hemocytes.
We collected 27 sequences of penaeidins from P.
monodon, P. vannamei, P. setiferus,P.japonicusandP.
chinensis. The mUltiple sequence alignment was
cre-ated with the ClustalW (Fig. 3). Based on the neighbor-joining analysis, a cluster tree was construct (Fig. 4).
The molecular weight (MW) of recombinant 6xHis-tagged penaeidin in this study was 6860.0 Da based on mass spectrometry data (data not shown). It indicated that the cleavage of secretory signal peptide was be-tween amino acids 19-20 as predicted by GCG and was performed as expected in the insect-baculovirus
expres-53
LV P3i LV P3b 71 LV P3j LV P3h LV P3f 63 LV P3d 100 LV P3e 19 LV P3g 23 LV P3a 33 47 LV P3c 67 100 LS P31 LS P3k LS P3n LS P3m . - - - FC P535
60 48 100 63 59 ESTPMI20 ESTPMCU19 ESTPMI2 PM P 1 . - - - MJ P2 98 99 , - - - LS P2d LVP1
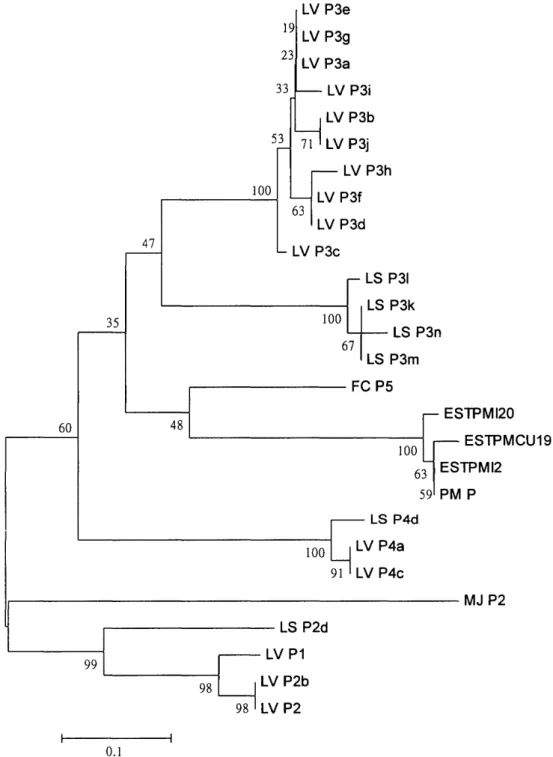
LV P2b LV P2 0.1Fig. 4. A comparison of 27 penaeidins based on a neighbor-joining analysis of their amino acid or deduced amino acid sequences. The distance is the proportion of amino acid sites at which the two sequences compared are different. Bootstrap values are given in percentage (Felsenstein, 1985).
si on system. In addition, the mass obtained (6860.0 Da) was 22.75 Da less than the estimated MW (6882.75 Da) of penaeidin tagged with 6xHis without modification. The difference could be explained by the formation of 3 intramolecular disulfide bridges and blocked N-terminal by a pyroglutamic acid.
Among the microbial species screened in the present study, the recombinant penaeidin (30 f.lM) could form an inhibition zone againstAerococcus viridans, but
not againstMicrococcus luteus, two Vibrio species and a
yeast Debaryomyces hansenii (Table 1). The inhibitory effect of the recombinant penaeidin of tiger shrimp against a filamentous fungus Neurospora crassa could
not be clearly determined. We, therefore, applied a higher concentration (77 f.lM) of the penaeidin and a weak inhibition of fungal growth was noted in the inhibi-tion zone assay (Table 1).
To further confirm the inhibitory activity of the re-combinant penaeidin against N. crassa, a determination
of minimum inhibitory concentration (MIC) was carried out. The recombinant penaeidin at different concentra-tions of 10, 20 and 40 f.lM delayed spore germination and growth of N. crassa but a complete inhibition of spore
germination was not observed.
Discussion
Although the length of cDNA sequence in the present study was less than the result obtained from Northern blot analysis, it could be explained by the size of 5' untranslated region, the length of poly A and/or the possible splicing. We already obtained the complete coding region of penaeidin. This was confirmed by a comparison of the sequences and an activity test of its product.
Sequence analysis with the BLAST algorithm showed that the deduced amino acid sequence of theP.
monodon penaeidin had the highest similarity to white
shrimp Penaeus setiferus penaeidin-3k (E value = 7x10-6
), similarity
=
61.6% using Gap program of GCG.Significant similarities with other penaeidins such as penaeidin-2, -3, -4 and -5 from white shrimp P.
vannamei, P. setiferus and Chinese shrimpP. chinensis
were also found. In the nucleotide database of NCBI, there were some penaeidin-similar EST (expressed se-quence tag) sequences such as ESTPMCU19 (Supungul et al., 2002), ESTPMI2 and ESTPMI20 of P.
monodon submitted from Thailand (accession numbers
81018089, 81784441 and 81784459 respectively). Comparing the sequence cloned in this study with those sequences using the Gap program of GCG, it shows that the sequences are highly identical (98.8-99.5%) and the deduced amino acid sequences are almost identical (98.6-100%).
The sequences of penaedins from different sources (Fig. 3) showed a conserved NH2-terminal proline-rich
domain and a COOH-terminal domain containing 6 cys-teines that could engage in 3 disulfide bridges residues as mentioned by Bachere et al. (2000). These two do-mains are usually observed in distinct groups of antimi-crobial peptides. Another feature of these penaeidin sequences is a frequent presence of basic amino acid lysine and arginine, which contributed positive charge to the antimicrobial peptides. Except LV P1 sequence that was identified solely on the basis of amino acid se-quence (Destoumieux et al., 1997), examination of all
penaeidin-coding sequences revealed that the leader sequence (1-19 residues) was almost completely con-served across most sequences, regardless of genus or species excluding P. japonicus. However, all of them had a C-terminal glycine residue, which was amidated in their mature form of Pen-2 and -3a for a possible in-crease in stability (Destoumieux et al., 2000a). In addi-tion, we observed that threonine was strongly conserved at position 27 of the aligned sequences, although Yang
et al. (2003) proved that the change of the amino acid
threonine to alanine could not affect an antimicrobial activity. It may suggest that penaeidins have another function other than the antimicrobial activity.
Penaeidins of white shrimp could be classified into penaeidin-2, -3, -4 groups. Penaeidin-1 was grouped together with penaeidin-2 group (Fig. 4). The cluster analysis in the present study showed similar pattern as noted by Cuthbertson et al. (2002). However P. monodon and P. chinensis penaeidins could not be
reli-ably clustered in penaeidin-3 group. It indicates that there is variance between penaeidins from different groups of penaeid shrimp. The same phenomenon was also observed on P. japonicus penaeidin, it could
not be reliably clustered in penaeidin-2 group.
The recombinant penaeidins derived from the white shrimpP. vannamei were synthesized in a yeast
expres-sion system (Destoumieux et al., 1999). Their antimi-crobial activities were almost indistinguishable from those of the native molecules. However, an additional glycine residue at the COOH-terminus instead of the naturally occurring a-amide was noted. In addition, two distinct Pen-3a isoforms were obtained either with an unprocessed glutamine residue or with the natural pyroglutamate at position 1. Moreover, about 50% of the recombinant molecules were shown to be O-substi-tuted by a dimannosyl moiety (Destoumieux et al.,
2000a). According to the molecular weight measure-ment of our recombinant penaeidin, it indicated an es-tablishment of a secretory signal peptide on cleavage, a formation of 3 intramolecular disulfide bridges, and an N-terminal blocked by a pyroglutamic acid. These post-translational modifications were consistent with the native modifications of the white shrimp penaeidin observed by Destoumieux et al. (1997). There was no sign of any glycosylation in our recombinant penaeidin. However, since we added 6xHis before the stop codon,
we could not determine the COOH-terminal amidation in the baculovirus-insect expression system.
We tried to isolate the recombinant penaeidin from insect cell lysate and culture medium. We were only able to collect significant amount of penaeidin in the cell lysate, which was consequently purified, but not in the culture medium. It suggested that the recombinant penaeidin, which was led by shrimp signal sequence, was preserved in Sf21 cells, in the same way the shrimp penaeidin is stored in the shrimp hemocytes and local-ized in granulocyte-cytoplasmic granules. (Destoumieux
et al., 2000b).
The results of the inhibition zone assays were simi-lar to the observations of Destoumieux et al. (1999) on white shrimp penaeidins whose antibacterial activities were noted against some Gram-positive bacteria but not against Gram-negative bacteria and yeasts. We could not observe, however, an inhibition zone formed by the recombinant penaeidin of tiger shrimp against M.luteus
that showed a certain degree of sensitivity to white shrimp penaeidins. It is possible that a higher concen-tration of the recombinant penaeidin is needed to form inhibition zones against M. luteus. Destoumieux et al.
(1999) showed that the MICs against M. luteus were higher by 2-4 times than A. viridans. In addition, the inhibition zone assay used in the present study may have low sensitivity in distinguishing the zones against
M.luteus.
Regarding an anti-fungal activity, data on white shrimp penaeidin activity demonstrated a broad-spec-trum antifungal property associated with fungicidal activity. Lower concentrations (below the MIC value) caused delayed growth of hyphae and abnormal r'!10r-phology of Fusarium oxysporum (Destoumieux et al.,
1999). In this study, a higher concentration (>40 J1M) seemed to be needed to observe the fungicidal activity. It may also mean that the recombinant penaeidin has no fungicidal activity but can inhibit fungal growth only. Comparing the MICs (1.25-5 J1M) of the white shrimp penaeidins (Destoumieuxet al., 1999) and discounting the possibility that the natural tiger shrimp penaeidin has lower antifungal activity than the white shrimp penaeidin, we speculate that the weakening of the anti-fungal activity might be due to the addition of 6xHis tag that followed the recombinant penaeidin COOH-terminal according to the inference that binding of penaeidins to chitin, and consequently their antifungal activity, is mediated by their COOH-terminal domain (Destoumieux et al.,2000a).
A striking feature of shrimp hemocyte libraries is the abundance of messages for antimicrobial peptides. Antimicrobial messages comprise 17.7% and 21% of all sequences isolated from the hemocyte cDNA libraries of
P. vannamei and P. setiferus, respectively, and penaeidin ESTs predominate among antimicrobial pep-tides (Gross et al., 2001). Direct screening of the P.
vannamei?L,Zap hemocytes library with penaeidin probes confirmed the robust nature of penaeidin expression in hemocytes with approximately 6.12% of all clones being penaeidin (Cuthbertson et al., 2002). In an EST library from hemocytes ofP. monodon,8.9% were identified as putative immune-related genes. Abundant penaeidin ESTs (frequency 7 among 47) were found in defense and homeostasis groups (Supungul et al., 2002).
Penaeidin mRNAs seem to appear frequently in hemocytes of shrimp. Recently, many antimicrobial peptides were reported to be involved in the regulation of immunity that included phagocytosis, opsonization, de-granulation, chemoattraction and complement reaction (Perregaux et al., 2002; Yang et al., 2002; Zasloff, 2002). We also aimed to determine if penaeidins have other biological function aside from antibiotic-like activity. In this study, we have demonstrated the molecular cloning, tissue localization of mRNA and antimicrobial activity of P. monodon penaeidin. We compared the sequence and antimicrobial property of the penaeidin with other shrimp penaeidins. In addition, the recombi-nant penaeidin has been expressed successfully using insect-baculovirus expression system, especially on the post-translational modifications.
Acknowledgements
The authors are grateful to Drs. J. P. Hsu and K. K. Lee, for donating the yeast and bacterial strains. Thanks are also extended to Dr. Shau-Chi Chi for kindly providing inverted fluorescence microscope facilities. Computer analysis was provided by GCG Sequence Analysis Service of National Health Research Institutes, Taiwan, Republic of China. The ESI-MS work was per-formed by Core Facilities for Proteomic Research, Insti-tute of Biological Chemistry, Academia Sinica, Taiwan, Republic of China. Council of Agriculture, Republic of China under the grant No. 92AS-4.2.3-FD-Z3 supported this research financially.
References
Bachere, E., D. Destoumieux and P. Bulet (2000): Penaeidins, antimicrobial peptides of shrimp: a comparison with other effectors of innate immunity. Aquaculture, 191, 71-88. Boman, H. G. (1991): Antibacterial peptides: key components
needed in immunity. Cell, 65, 205-207.
Cole, A. M., P. Weis and G. Diamond (1997): Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. BioI. Chem., 272, 12008-12013.
Cuthbertson, B. J., E. F. Shepard, R. W. Chapman and P. S. Gross (2002): Diversity of the penaeidin antimicrobial pep-tides in two shrimp species. Immunogenetics, 54, 442-445.
Destoumieux, D., P. Bulet, D. Loew, A. Van Dorsselaer, J. Rodriguez and E. Bachere (1997): Penaeidins, a new fam-ily of antimicrobial peptides isolated from the shrimp Penaeus vannamei (Decapoda). J. BioI. Chem., 272,
28398-28406.
Destoumieux, D., P. Bulet, J. M. Strub, A. Van Dorsselaer and E. Bachere (1999): Recombinant expression and range of activity of penaeidins, antimicrobial peptides from penaeid shrimp. Eur. J. Biochem., 266, 335-346.
Destoumieux, D., M. Munoz, P. Bulet and E. Bachere (2000a): Penaeidins, a family of antimicrobial peptides from penaeid shrimp (Crustacea, Decapoda). Cell Mol. Life. Sci., 57, 1260-1271.
Destoumieux, D., M. Munoz, C. Cosseau, J. Rodriguez, P. Bulet, M. Comps and E. Bachere (2000b): Penaeidins, anti-microbial peptides with chitin-binding activity, are produced and stored in shrimp granulocytes and released after mi-crobial challenge. J.Cell Sci., 113,461-469.
Felsenstein, J. (1985): Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39, 783-791. Gross, P. S., T. C. Bartlett, C.l. Browdy, R. W. Chapman and
G. W. Warr (2001): Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific white shrimp,Litopenaeus vannamei, and the Atlantic white shrimp, L. setiferus. Dev. Comp. Immunol., 25, 565-577.
Hultmark, D., A. Engstrom, H. Bennich, K. Rashmi and H. G. Boman (1982): Insect immunity: Isolation and structure of cecropin D and four minor antibacterial components from Cecropia pupae. Eur. J. Biochem., 127, 207-217. Hultmark, D., A. Engstrom, K. Anderson, H. Steiner, H. Bennich
and H. G. Boman (1983): Insect immunity. Attacins, a family of antibacterial proteins from Hyalophora cecropia. EMBO J., 2, 571-576.
Kumar, S., K. Tamura, I. B. Jakobsen and M. Nei (2001): MEGA2: Molecular Evolutionary Genetics Analysis software. Bioinformatics, 17, 1244-1245.
Lee, J. Y., A. Boman, S. Chuanxin, M. Anderson, H. Jounvall, V. Mutt and H. G. Boman (1989): Antibacterial peptides from pig intestine: isolation of a mammalian cecropin. Proc. Nat!. Acad. Sci. USA, 86, 9159-9162.
Munoz, M., F. Vandenbulcke, D. Saulnier and E. Bachere (2002): Expression and distribution of penaeidin antimicro-bial peptides are regulated by haemocyte reactions in microbial challenged shrimp. Eur. J. Biochem., 269,
2678-2689.
Nei, M. and S. Kumar (2000): Molecular Evolution and Phylogenetics. Oxford University Press, New York. p. 87. Perregaux, D. G., K. Bhavsar, L. Contillo, J. Shi and C. A. Gabel (2002): Antimicrobial peptides initiate IL-1 beta posttransla-tional processing: a novel role beyond innate immunity. J. Immunol., 168,3024-3032.
Qu, X., H. Steiner, A. Engstrom, G. Bennich and H. G. Boman (1982): Insect immunity: Isolation and structure of cecropins Band D from pupae of the Chinese oak silk moth,Antheraea pernyi. Eur. J. Biochem., 127,219-224. Rojtinnakorn, J, I. Hirono, T. Itami, Y. Takahashi and T. Aoki (2002): Gene expression in haemocytes of kuruma prawn, Penaeus japonicus, in response to infection with WSSV by EST approach. Fish Shellfish Immunol., 13, 69-83. Supungul, P., S. Klinbunga, R. Pichyangkura, S. Jitrapakdee, I.
Hirono, T. Aoki and A. Tassanakajon (2002): Identification of immune-related genes in hemocytes of black tiger shrimp(Penaeus monodon) Mar. Biotechnol. 4, 487-494. van't Hof, W., E. C. Veerman, E. J. Helmerhorst and A. V.
Amerongen (2001): Antimicrobial peptides: properties and applicability. BioI. Chem., 382, 597-619.
Xia, X. (2000): Data Analysis in Molecular Biology and Evolution. Kluwer Academic Publishers. 277 pp. Xia, X. and Z. Xie, (2001): DAMBE: software package for data
analysis in molecular biology and evolution. J.Hered., 92, 371-373.
Yang, D., A. Biragyn, L. W. Kwak and J. J. Oppenheim (2002): Mammalian defensins in immunity: more than just microbicidal. Trends Immunol., 23, 291-296.
Yang, Y., J. Poncet, J. Garnier, C. Zatylny, E. Bachere and A.
Aumelas (2003): Solution structure of the recombinant penaeidin-3, a shrimp antimicrobial peptide. J. BioI. Chem., 278, 36859-36867.
Zasloff, M. (1987): Magainins, a class of antimicrobial peptides fromXenopus skin: isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA, 84, 5449-5453.
Zasloff, M. (2002): Antimicrobial peptides of multicellular organisms. Nature, 415, 389-395.