國
立
交
通
大
學
理學院網路學習學程
碩
士
論
文
構築及表現白色念珠菌
ORF19.2730 的重組 DNA
及其功能性研究
The construction and functional study of ORF19.2730
recombinant DNA of Candida albicans
研 究 生:戴勝賢
構築白色念珠菌ORF19.2730 的重組 DNA 及其功能性研究 The construction and functional study of ORF19.2730
recombinant DNA of Candida albicans
研 究 生:戴勝賢 Student:Sheng-Shyan Tai 指導教授:楊昀良 Advisor:Yun-Liang Yang 國 立 交 通 大 學 理學院網路學習學程 碩 士 論 文 A Thesis
Submitted to Degree Program of E-Learning College of Science
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Degree Program of E-Learning July 2008
Hsinchu, Taiwan, Republic of China
構築及表現白色念珠菌
ORF19.2730 的重組 DNA 及其功能性研究
研究生:戴勝賢 指導教授:楊昀良 博士摘要
白色念珠菌(Candida albicans)是人體重要的伺機性致病真菌之一,具有多 形態的特性。當人體免疫力下降或受藥物副作用影響而改變體內菌落生態時,便 會伺機感染黏膜組織;局部性臨床症狀如鵝口瘡、陰道炎等,嚴重者會造成全身 性系統感染而死亡。近年來因為免疫不全患者增加及使用抗生素不當而產生抗藥 性等因素,導致院內感染的病例不斷攀升。對白色念珠菌的基因研究將有助於感 染的預防、治療、及解決抗藥性等問題。 Braun等人(2005)綜合Standford、CandidaDB、CGD等主要的白色念珠菌 基因資料庫,比對總數11615個推定的ORF片段,排除重複及錯誤的部份,確認 出6354個基因片段,其中包括功能不明或未確定的部分基因。實驗室先前經抑制 相減雜交之比對發現orf19.2730的序列可能涉及形態變化。orf19.2730是位於第四 對染色體上的ORF片段,屬於Contig19-10158,和orf19.10244為對偶基因,計有1821 個鹼基對。本研究以目前電腦資料庫推定而未經實驗確認的orf19.2730為標 的,因其編碼蛋白質與啤酒酵母菌的RPH1有同源性,與DNA修補有關。以啤酒 酵母(Saccharomyces cerevisiae)為表現平台,藉著偵測其蛋白質表現來推測此ORF的功能性。首先以PCR的方式自白色念珠菌的Genomic DNA 擷取此ORF 片段及其上游的promoter共2192個鹼基對插入YEP363質體,構築成YEP363-TA1 質體。在E.coli增殖後轉殖到啤酒酵母
,
以Coomassie blue staining檢測,未觀察到 明顯的表現。另由實驗室已構築的pET∆5T系列質體擷取HAHis Tag序列做為 標籤,連接在去除3’端終止碼的orf19.2730下游,再以西方轉漬分析偵測, 發現其確實是一個能產生蛋白質的ORF。又將orf19.2730上游371個鹼基的序 列分成三個片段進行β-galactosidase filter assay觀察,發現在中段-257個鹼基 到-315個鹼基的區域可能有未知的負向調控之調控子存在。The construction and functional study of ORF19.2730 recombinant DNA of Candida albicans
Student: Sheng-Shyan Tai Adviser: Dr. Yun-Liang Yang Degree Program of E-Learning
College of Science
National Chiao Tung University ABSTRACT
Candida albicans is one of the important opportunistic fungal pathogens in
human body. It has various morphologies. When body immunity deteriorates or microbial ecology in the body changes due to medical reasons, the fungus would have opportunity to infect mucous tissue; typical clinical symptoms include thrush and vaginitis. In the worst case, it would lead to general system infection and death. Recently, because of the increase of immune deficiency patients and drug resistance due to improper use of antibiotics, the cases of nosocomial infection keeps rising. Genetic and functional studies on Candida albicans may help in issues of infection prevention, treatment, and drug resistance.
Previously, the laboratory has conducted suppression subtractive hybridization and found the sequence of orf19.2730 might have involved in the morphogenesis of Candida albicans. Orf19.2730 is a hypothetical gene consisting 1821 nucleotides and is in Contig19-10158 on the fourth chromosome of C. albicans. The homologous allele is orf19.10244. The sequence of orf19.2730 and the upstream promoter, 2192 nucleotides in total, was amplified by PCR. The ORF was then cloned into 2µ vector for expression in
Saccharomyces cerevisiae. The promoter sequence was fused to lacZ gene and
then was transformed into Saccharomyces cerevisiae for the purpose of identifying cis-acting sequences regulating the promoter activity. Western blot analysis showed that the ORF indeed could manifest, able to produce protein.
β-galactosidase filter assay also showed that there may have inhibitory sequence
誌謝
終於可以昭告諸親朋好友,我的論文完成了。這一篇拖遲不出橫亙數 年的論文,今日能夠勉力完成,有賴太多貴人的加持與協助! 感謝志鴻指引交大理學院專班這一條明路並告誡我既入寶山就不要空 手而歸!感謝林文彬校長獨排眾議並給予祝福讓我得以一圓而立之前未能 完成的研究所之夢!感謝專班提供進修的機會及選擇研究主題的包容! 最要感謝指導教授楊昀良老師不惜跌股的接納、容忍、引領與提攜, 給我最大的空間與時間,允許我能在專班廣泛探索了兩年知識的寶庫後, 再進實驗室享受三年生命科學的焠鍊、思維邏輯的訓練及自我實現私慾的 滿足!還要感謝羅秀容老師有關白色念珠菌的啟蒙與指導!論文的完成有 幸承蒙彭慧玲老師、藍忠昱老師、謝家慶老師及徐琅老師悉心的審查及愷 切的指正與建議,使本論文得以更為詳實與縝密。另外從各位老師身上所 領略到博學鴻儒待人治學與處事的態度,更使我獲益匪淺!在此謹致最大 的謝忱! 實驗室裡忙碌而喜樂的一千多個日子裡,感恩建斈、婷尹、志豪、怡 瑾、宗翰、佳叡、育穎毫無保留的技術指導與協助,使我節省許多摸索的 時間提早進入狀況。也要感謝欣彬、淑萍、旻秀、惠敏、敏書、淑貞、柏 伶、瀞云及萍芳這群充滿陽光活力的年輕夥伴們一路同行!有幸與你們共 渡這一段學習成長的歲月! 實驗室多少漫漫長夜,感謝生科系葉大哥的夜半關懷送暖!延宕多時 的論文得以產出,感謝專班張小姐臨門一腳的催促! 一路走來,感恩菩薩加持維、汝平安長大沒有太多的顢頇。感激內子 玉珍支持體諒包容與付出,讓我能無後顧之憂。感謝岳母的鼓勵與關懷。 謝謝父母的支持與打氣。 感激您們及所有一路相伴的貴人們! 戴勝賢 謹誌 于 2008 的生日目錄
中文摘要... i 英文摘要... ii 誌謝...iii 目錄... iv 圖目錄... vii 1.一、緒論... 1 1.1.前言 ... 1 1.2.白色念珠菌... 21.3. DNA 修補(DNA Repair)之探討... 2
1.3.1. 光恢復修補(photoreactivation)... 3
1.3.2. 切除修補(Excision repair)... 3
1.3.3. 錯配修復(Mismatch Repair,MMR) ... 4
1.3.4. 雙股螺旋斷裂修補 (double-strand break DNA repair)... 5
1.3.5. 錯誤傾向修復作用(Error-prone repair) ... 6 1.3.6. 有關白色念珠菌的 DNA 修補之研究... 7 1.4.本論文之研究目標... 8 2.二、材料... 9 2.1.菌株... 9 2.2.質體... 10 2.3.引子(primer)... 11 2.4.藥品試劑... 11 2.5.緩衝溶液及溶液 ... 14 2.6.培養基配製... 16 2.7.儀器設備... 17 3.三、實驗方法... 19
3.2.以 Excel Pure Plasmid mini kit 抽取 E. coli 質體 ... 19 3.3.聚合酶連鎖反應(PCR) ... 19 3.4. PCR 產物的 T-vector 檢驗 ... 20 3.5.限制酶反應(Sambrook et al, 1989) ... 21 3.6.萃取洋菜膠內之 DNA 片段 ... 21 3.7. E. coli 的連接反應(Ligation)和轉殖作用(Transformation)... 22 3.8.真菌的轉殖作用(Yeast transformation) ... 22 3.9.抽取啤酒酵母質體... 23 3.10.萃取真菌細胞萃取液... 23
3.11.SDS PAGE 進行 Coomassie blue stain 測試... 23
3.12.西方轉漬分析(Western Blotting) ... 24
3.13.Yeast β-galactosidase filter assay ... 25
3.14.侵犯力分析(invasion) ... 25
4.四、結果... 27
4.1.白色念珠菌 Genomic DNA 的資料庫分析 ... 27
4.2.將 Candida albicans Genomic DNA 的 orf19.2730 及其上游序列 轉殖到 YEP363 和 pRs426 質體... 28
4.3.將 Ta2Y 質體轉殖到啤酒酵母菌建構 10560-2B-Ta2y... 29
4.4. Coomassie blue staining... 29
4.5.建構 Ta2HAS 質體... 29
4.6.將 Ta2HAS 質體轉殖到啤酒酵母建構 10560-2B-Ta2HAS... 31
4.7. Western bloting ... 32
4.8.建構帶有 URA3 序列的 YEP363URA3 和 YEP363TA1URA3 質體 ... 32
4.9.建構酵母菌 SLO69Tu 和 SLO69Yu ... 33
4.10.侵犯力分析... 34
4.11.以 Yeast β-galactosidase filter assay 對 orf19.2730 的 promoter 檢驗... 35
5.五、討論... 36
5.1. orf19.2730 資料庫分析之探討 ... 36
5.2.以啤酒酵母菌觀察 orf19.2730 表現 ... 37
5.3.西方墨點法檢驗 orf19.2730 之表現 ... 38
5.5. orf19.2730 上游 promoter 的探討 ... 40 5.6.侵犯力分析之探討... 41 5.7.結語與未來展望... 42 六、參考文獻... 43 附錄 1:SB021 序列 ... 74 附錄 2:SB021 和 orf19.2730 比對結果 ... 75 附錄 3:SB021 和 orf19.2743 比對結果 ... 76 附錄 4:orf19.2730 序列 ... 77 附錄 5:orf19.2743 序列 ... 79
附錄 6:CGD 資料庫中和 DNA repair 相關的 ORF ... 81
圖目錄
圖 1. SB021 序列和 orf19.2730 對應之相關位置 ... 49
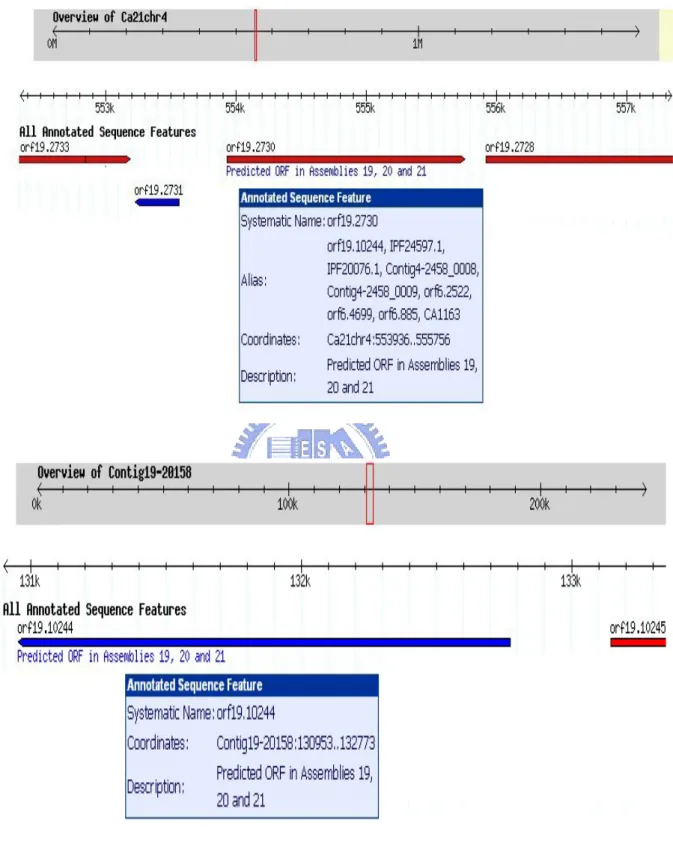
圖 2. orf19.2730 及其對偶基因 orf19.10244 在 Candida albicans 第四對 染色體上的相對位置... 50
圖 3. orf19.2730 及其上游序列之限制酶切割部位分析... 51
圖 4. PCR 切取 orf19.2730 及其上游序列,長度為 2192 個鹼基 ... 52
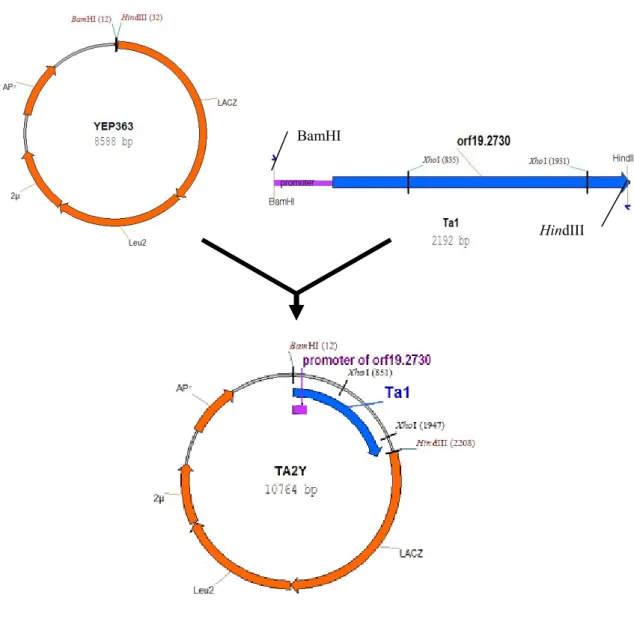
圖 5. Ta2Y 質體的構築... 53
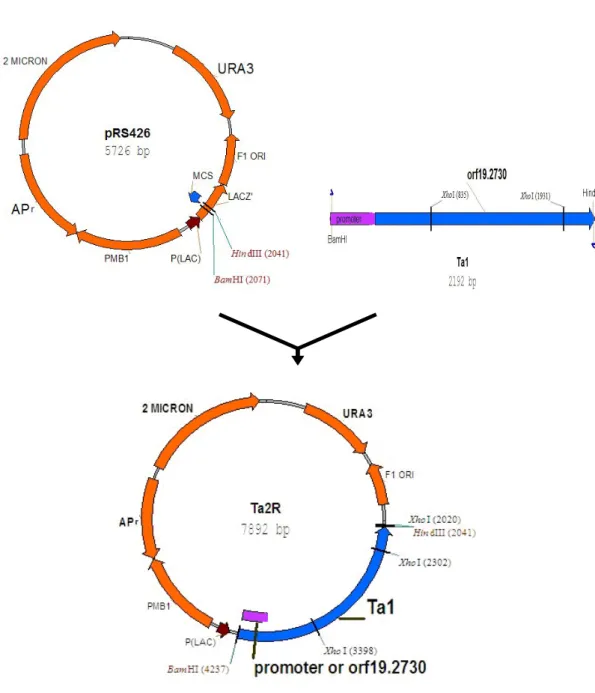
圖 6. Ta2R 質體的構築... 54
圖 7. Ta2Y 及 Ta2R 以限制酶 XhoI 切割後膠體電泳分析... 55
圖 8. Ta2Y 質體轉型到啤酒酵母菌 10560-2B,建構 10560-2B-Ta2Y... 56
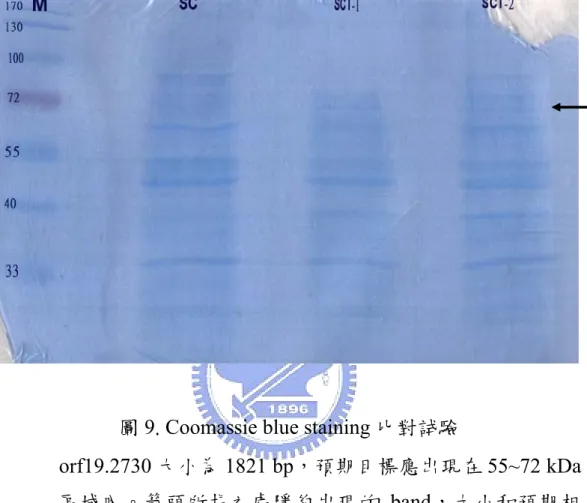
圖 9. Coomassie blue staining 比對試驗... 57
圖 10.以 PCR 方式自 Ta2Y 切取 orf19.2730∆stop ... 58
圖 11.以 BamHI 和 HindIII 切割 YEP363 質體及 orf19.2730∆stop ... 59
圖 12. Ta2Y∆stop 質體 ... 60 圖 13.以限制酶 XhoI 切割確認 Ta2Y∆stop 質體 ... 61 圖 14.以 PCR 方式在 pET∆5T-D24B-HAHis 質體上擷取 HA3His6 及其 下游共 996 個鹼基的 pHas 片段 ... 62 圖 15. Ta2HAS 質體... 63 圖 16.經限制酶 XhoI 處理確認建構 Ta2HAS 質體... 64 圖 17.檢驗 10560-2B-Ta2HAS 轉型株 ... 65
圖 18.啤酒酵母菌及 10560-2B-Ta2HAS 進行 Coomassie blue staining 及 Western bloting ... 66 圖 19. proURA3 ... 67 圖 20.建構 YEP363URA3 質體及 YEP363TA1URA3 質體 ... 68 圖 21.建構質體 YEP363URA3 和 YEP363TA1URA3 的確認 ... 69 圖 22.對酵母菌 SLO69Yu 和 SLO69Tu 抽取質體後進行 PCR 檢驗 ... 70 圖 23.侵犯力分析(Invasion assay)結果... 71 圖 24.建構 YpL7、YpL5,及 YpL2 等三種質體的建構... 72
圖 25.對 SCYpL7、SCYpL5 及 SCYpL2,進行 Yeast ß-galactosidase filter assay ... 73
1.
一、緒論
1.1.
前言 白色念珠菌(Candida albicans)是人體重要的伺機性致病(opportunistic pathogen)真菌之一,具有多形態的特性。通常可以少量存在於一般人的 口腔、喉部、皮膚、指甲溝、大腸、陰道等黏膜組織,與人體及其他體內 菌種保持平衡。正常情形下健康而營養良好的成年人有著功能正常的免疫 系統可以維持這種平衡。當這種平衡受到化學療法、使用抗生素、口服避 孕藥、懷孕、衰弱的免疫系統、營養不良、過多的壓力和生病等因素衝擊 而失調,或受藥物副作用影響而改變體內菌落生態時,便會伺機感染黏膜 組織。局部性臨床症狀如鵝口瘡、陰道炎等,嚴重者會造成全身性系統感 染而死亡(陳豪勇等, 2000),其中白色念珠菌血症的死亡率甚至可高達 40%(Barelle et al, 2003; Wenzel and Gennings, 2005)。抗生素是對抗細菌感染的重要藥物,但它沒法區別體內菌落。因此抗 生素在殺掉有害菌種的同時也殺掉有益菌種,而給白色念珠菌有機會增殖 的空間。近年來因為免疫不全患者增加及使用抗生素不當等因素,導致院 內感染的病例不斷攀升。在台灣醫院內真菌培養分離出白色念珠菌所佔的 比例可高達60.4﹪(Hsueh et al, 2002)。一般真菌的代謝方式與哺乳動物 的細胞相似,所以在開發抗真菌感染的藥物相當困難。而真菌侵入細胞內 部後,其會轉變以脂質來取代醣類產生能量,這使得它們有更強的致病力, 也可能成為藥物攻擊的目標(Lornenz & Fink, 2001)。目前抗真菌藥物主 要有Polyenes、Azole-base 與 5-Flucytosin 及 Caspofungin 等四類(Dupont, 2001;Yang and Lo, 2001;Ripeau et al, 2002)。但是這些藥物的使用有其 細胞毒性及副作用等不良反應與限制。隨著後抗生素時代全球性抗藥性病 菌的崛起肆虐,抗真菌藥物的使用量日增(Chen et al, 1997;陳宜君, 2002),目前甚至已有白色念珠菌抗藥性菌株產生(White et al., 1998; Schuetzer-Muehlbauer et al., 2003;Akins, 2005),使得念珠菌感染成為不 容忽視的醫學課題。以分子生物學方法對白色念珠菌的基因探討及其調控 機制等研究將可協助尋找新的藥物標的,進而有助於感染的預防、治療、 及解決抗藥性等問題。
1.2.
白色念珠菌念珠菌屬約有 81 種,多數為人體正常菌群,少數對人有致病性。致病 性念珠菌主要有白色念珠菌(C. albicans)、C. tropicalis、C. parapsilokis、 C. krusei 等,其中最為常見的是白色念珠菌。白色念珠菌的菌體成圓形或
卵圓形(2×4µm)。出芽繁殖時若芽管延長不與母體脫離,則形成假菌絲。 在 YPD 培養基無血清時生長良好。生長時需氧。室溫或 37℃培養 3 天長 出類酵母型菌落,表面光滑,呈灰白色或奶油色。隨培養時間稍延長,菌 落增大,顏色變深並皺褶。用玉米培養基培養時則易形成厚膜孢子(郭曉 奎, 2007)。白色念珠菌可以酵母菌型(yeast form)、假菌絲型(pseudohyphal form)、菌絲型(true hyphal form)及厚膜孢子型(chlamydospores)等四 種型態存在,型態之間的轉換主要受環境條件影響如溫度、酸鹼值、氮源 或碳源是否充足、細胞密度等(Ernst, 2000)。環境條件的訊息通常經由 信號傳導途徑傳達給細胞,使得特定基因被誘導或被抑制,而啟動形態轉 變的機制(Brown, 2002; Soll, 2002)。其中酵母菌型和菌絲型之間的轉 變決定著白念珠菌的致病能力(Odds, 1994; Lo et al., 1997),酵母菌型 有助於菌體的散佈到宿主其他位置,而菌絲型則可能有助於侵犯組織逃避 巨噬細胞的攻擊,所以生成菌絲的能力為白色念珠菌產生致病性不可或缺 的條件之一。
1.3.
DNA 修補(DNA Repair)之探討DNA 修補是細胞內維持 DNA 結構完整及修復 DNA 損害最重要的一
個機制。當生物體 DNA 受到環境突變原(environmental mutagens)例如
x-ray,紫外線,以及有毒化學物質等刺激而導致DNA 分子結構的破壞時,
會改變細胞閱讀信息和基因編碼的方式,所以 DNA 修複必須經常運行以 迅速改正 DNA 結構上的任何錯誤之處,維持整個基因庫在細胞複製時的 正確性。遭受DNA 損傷的細胞需經 DNA 修補後才能再繼續 DNA 複製及 細胞分裂等過程(Bao et al., 2001)。目前已知細胞內的 DNA 修復機制中, 由 DNA 損害斷裂的程度可以分為兩種類型,一種是單股損害的修復;當 DNA 兩股的其中一股發生損害時,就可以另一股為模版(template)作為修
正的依據。另一種則是DNA 雙股斷裂;DNA 雙股斷裂是在缺乏另一股序 列當模版的情況下,透過同源的染色體(homologous chromosome)序列或姊 妹染色分體(sister chromatid)來尋求支援。DNA 雙股斷裂的修復有時候可能 無須任何序列當模版,而逕行將斷裂部分直接接合,這種 DNA 修復方式 則隱含錯誤的機率(error-prone)(Wikipedia, 2008)。 1.3.1. 光恢復修補(photoreactivation)
光恢復修補是指紫外線使 DNA 產生嘧啶二聚體(purimidine dimer)造 成 的 損 傷 , 其 中 共 價 鍵 的 連 接 可 直 接 被 一 種 光 依 賴 型 的 光 分 解 酶 (photolysae)所逆轉。光分解酶是細菌內的修復蛋白,當光分解酶結合到 DNA 的變形區域時,即具近紫外光及藍色的光譜區域的吸收帶。吸收光子 造成激動態切割二聚體回到其原始鹼基,故可將紫外線所造成的雙嘧啶鍵 結在可見光照射後恢復成完整無損的狀態。啤酒酵母菌 DNA 中的 PHR1 基因即可編碼光分解酶,是在轉錄層級上和光恢復修補有關的基因。PHR1 的主要調節者是一段含有39 個鹼基,稱為 URS(PHR1) 的序列,這個序列 是組成damage-responsive repressor PRP 蛋白的 binding site。RPH1 是 PHR1 的抑制者之一。Rph1p 有兩個 zinc fingers 的構造,可辨識 URS(PHR1)上的 連結區域 AG(4) (C(4)T)序列而加以連結,進行對 PHR1 的抑制調控。當 RPH1 基因被刪除後,PHR1 基因不再被抑制,其所表現的光恢復修補速率 也因此加快 (Jang, Wang, and Sancar, 1999) 。由於這一類的蛋白可以直接 將遭受破壞的 DNA 或核苷酸還原,因此不需要另一股當作修復的模版, 是單一蛋白所參與的直接修復機制 (Direct repair) 的一種方式。
1.3.2. 切除修補(Excision repair)
切除修補是細胞內 DNA 損傷的主要修補機制。其作用是由一種識別 酶(Recognition enzyme)起始,此酶能發現損傷鹼基或 DNA 空間結構的變 化。識別後把含有損傷鹼基的序列切除掉,然後合成一段新的 DNA 來代 替切除掉的部分。這種修復系統很常見,有些能識別 DNA 的一般損傷, 其他的則僅作用於鹼基損傷的特殊類型,如糖基化酶(DNA glycosylase) 去除特異的鹼基改變;AP 內切核酸酶(AP endonuclease)則去除無嘌呤位 置的殘基。在單細胞類型中通常存在多種切除修復系統,能夠處理發生的
大部分損傷。切除修復系統主要可分為鹼基切除修復(Base-excision repair) 和核苷酸切除修復(Nucleotide-excision repair)。
完整的 DNA 的骨架中一個鹼基遺失所形成的缺口稱為 AP 位置 (apurinic-apyrimidinic site , AP site)。鹼基切除修復是指先用糖基化酶把單 一個錯誤的含氮鹽基移除,再用 AP 內切核酸酶切在鹽基被移除後的空位 以移除磷酸鍵形成nick。接著 DNA polymerase I 從 nick 處的 free 3’OH 開 始修復,拿掉部分壞損的DNA,同時加上未損壞 DNA。DNA polymerase I 脫落後,DNA ligase 會把 nick 封好,完成修復(David-Cordonnier et al., 2002)。 核苷酸切除修復則先用ABC excinuclease 在發生錯誤的部分(DNA lesion) 左右各切下數個核苷,切下的數目因物種和損壞的鹼基數目而定。再經過 DNA helicase 作用後,將含有損壞鹼基的 DNA 去除,然後由 DNA polymerase I 把缺少的核苷補上,再經由 DNA ligase 把 nick 封補(Van Houten
et al, 1987)。 核苷酸切除修復和鹼基切除修復最大的不同是拿走一小段核苷酸而不 是單獨一個鹼基。不同切除修復系統的專一性是不同的,但是它們每個系 統都是去除DNA 錯配或損傷的鹼基,然後用一段新合成的 DNA 序列代替。 1.3.3. 錯配修復(Mismatch Repair,MMR) 錯配修復主要是負責 DNA 在複製過程發生嘌呤—嘧啶錯誤配對的校 對工作。舊有的DNA strand 是被甲基化的,而剛合成出來的 DNA 則否。 藉由辨識DNA 是否被甲基化可以辨識出 DNA 的新舊。比對新舊之間的差 異,當發現 DNA 不一樣時,就可進行除錯。因此,大部分的修復都要在 複製完成後的很短時間內進行,才可根據是否有甲基化而分別。一但新股 被Dam methylase 甲基化之後,就無法校對兩股的對錯了。不正確配對的 錯配鹼基可能是一個C 與 A 錯誤配對,或者是 polymerase 跳過或重複一些 鹼基,形成1 至 5 個不成對的鹼基。
大腸桿菌(E. coli)參與錯配修復的酵素主要有 MutH、MutL、MutS 和SSB 等。其中 MutH 會辨識 GATC 序列;MutL 用來連接 MutH 和 MutS 的蛋白,形成一個複合物(MutH- MutS complex);MutS 則辨識突變的核 苷並與其相接。SSB 會保護 ss DNA(single strand DNA)。藉由 MutH- MutS complex 的形成,DNA 會被拉近形成了 DNA loop。MutH 帶有一個
site-specific endonuclease,要在遇到 hemimethylated GATC 序列時才會被活 化。GATC endonuclease 會對未甲基化的新股上的 GATC 中 G 的 5’端作用 (nick),標記出這個新股是要被修復的。再以 DNA helicase II 由 3’到 5’把 DNA 打開(unwind),然後 exonuclease I 將出錯的這一股 DNA 從 GATC 的 地方開始去除核苷,一直到突變的部位為止。如果突變的部分位於兩個 GATC 中間,上述 nick 和去除核苷的動作也可以從有 GATC 的兩端進行。 由於DNA 發生損傷,所以新的 DNA 會在相對原來發生損傷的部位出現缺 口。從另一條未受損的姐妹股(sister strand)上可以切下相應的核苷酸部 分填到這個缺口中,最後再用DNA polymerase III 和 DNA ligase 把空缺 部份補起來。(Li, 2008)
啤酒酵母的 Msh 修復系統與大腸桿菌的 Mut 系統同源。以 MSH2 為 識別錯配裝置的主體,MSH3 和 MSH6 提供特異性的參數。MSH2-MSH3 複合物可結合2 至 4 個核苷酸的錯配環,而 MSH2-MSH6 複合物則與單一 鹼基錯配或插入或缺失部位結合。(Marsischky et al, 1999; Harrington & Kolodner, 2007)
1.3.4. 雙股螺旋斷裂修補 (double-strand break DNA repair)
當 DNA 的傷害型式是雙股螺旋斷裂時,就無法利用互補的特性進行 修復,所以雙股 DNA 的斷裂所造成的傷害對細胞來說是最嚴重也是最致 命的的 DNA 傷害。對於 DNA 的雙股螺旋斷裂,生物細胞發展出兩種主 要機制進行修復:一種為同源重組修復系統(homologous recombination, HR),另一種為非同源的黏合系統(non-homologous end-joining,NHEJ)。 HR 是利用細胞內的染色體兩兩對應的特性,若其中一條染色體上的 DNA 發生雙股斷裂,則另一條染色體上對應的 DNA 序列即可當作修復的 模版來回復斷裂前的序列。HR 修復路徑與細胞週期有很高的依賴性。在 G1 期,染色體套數為 2 倍體,同源染色體是 HR 唯一可使用的模版。等到 在S 期及 G2 期時,有了姊妹染色分體(sister chromatid)的加入,染色體套 數倍增為4 倍體,這時候 HR 機制就有更多的修復模版可供選擇,因此一 般認為HR 修復的運作在 S 期/G2 期比較活躍(Ciudad et al., 2004)。此種 修復所憑藉的是正確的模版,所以其修復的準確度極高,但是細胞內的染 色體散佈在細胞核內,要將相對應的同源染色體與同源序列正確的並排在
一起來進行雙股螺旋斷裂的修復是十分耗時費事的工作。對於高等生物而 言,因為基因體太複雜,要進行同源重組較困難,所以除非在細胞進行複 製後又尚未將新複製序列與原本序列分離,同源序列容易取得的時期,一 般是比較不會利用此一系統。 另一個進行 DNA 雙股螺旋斷裂修復的模式是 NHEJ。NHEJ 與 HR 最 大的差異就在於不需要任何模版的幫助。NHEJ 的修復蛋白可以直接將雙 股裂斷的末端彼此拉近,再藉由 DNA 黏合酶(ligase)的幫助,將斷裂的兩 股重新接合。它的主要作用僅是將斷裂的 DNA 雙股螺旋重新加以黏合, 並未對於斷裂的部份進行實質的比對與修復。NHEJ 的機制比 HR 簡單又 不依靠模版,在基因體複雜、包含越多無用 DNA(junk DNA)的生物體, NHEJ 會比 HR 系統有效率。但是在基因體簡單的單細胞型態生物,NHEJ 則很可能破壞原本序列完整性。然而對細胞而言,DNA 雙股螺旋斷裂是 一種致命的傷害形式,在這種緊迫的情況之下,NHEJ 修復的效率比 HR 修復的正確性更重要。(沈志陽,2006;黃憲達, 2005;Lewin, 2000)。 在啤酒酵母菌中的基因體十分複雜,雖然使用同源重組修復系統雖然 修復結果不容易出錯,但在效率的考量之下,對於非同源的NHEJ 依賴程 度亦非常深。NHEJ 僅發生在缺乏 Rad52p 或大量的 DNA 損壞發生之後 (Ciudad et al., 2004)。
1.3.5. 錯誤傾向修復作用(Error-prone repair)
當細胞暴露在強的輻射線照射下,DNA 被破壞的程度遠超過之前的幾 個修補系統所能修補的程度,DNA 就會整個四分五裂。為此細胞只好選擇 一條極度不精確而且會造成高突變率的 SOS 反應(SOS response),啟動錯 誤傾向修復作用(Error-prone repair)。
SOS 反應是藉著 DNA 損傷釋放出誘發信號以啟動 RecA 蛋白酶的活 性。RecA 啟動後可活化 LexA。LexA 在未經處理的細胞中相對穩定,可 結合到與啟動子重疊的 SOS 框(SOS box)序列而抑制許多修復功能基因的 操縱子表現。LexA 的蛋白酶活性被 RecA 啟動後,會因為發生自我切割而 失效,使得所有與其關聯的操從子開始作用而表現基因修復的功能。RecA 和 LexA 互為拮抗,RecA 可切割 LexA,而 LexA 又會抑制 RecA,所以 SOS 反應可使 RecA 蛋白和 LexA 的抑制物均被擴增。當 DNA 修補完成,
DNA 損傷的信號被去除後,RecA 蛋白質即失去活性。此時,LexA 蛋白 質則不再自我切割,而以未切割的形式重新和相關的啟動子結合而終止 SOS 反應(Maor-Shoshani et al., 2000)。
1.3.6. 有關白色念珠菌的 DNA 修補之研究
真菌類(Yeast)的 DNA 修補機制可區成分基因相關(Associated with gene)和非基因相關(not associated with gene)兩類途徑。其中和基因相 關的機制包括鹼基切除修復(base excision repair)、核苷酸切除修復 (nucleotide excision repair)、錯配修復(mismatch repair pathway)及雙股 斷裂修復(double-strand break repair pathway)等。
根據 CGD 資料庫的註解,目前經過電腦推定和白色念珠菌相關的基 因或 ORF 有 53 個,但只有四種基因被證實(附錄六),其中 DPB2 (orf19.7564)是從序列或結構相似性推斷而來(Inferred from Sequence or structural Similarity, ISS),目前僅初步證實和 DNA 修補有關,電腦推測 可能和錯配修復或是和核苷切除修復有關的部份則尚未確認(Backen et al., 2000)。另外三種基因是從突變體表現型實驗推斷而來(Inferred from Mutant Phenotype, IMP),包括MRE11(orf19.6915)、RAD50(orf19.1648) 及RAD52(orf19.4208),三者都涉及 DNA 修補,同時也和白色念珠菌對 藥物的敏感性或耐受性的轉變及對氧化壓力(oxidative stress)的反應有關 (Legrand et al., 2007; Chibana et al., 2005)。RAD50(orf19.1648)及 RAD52 (orf19.4208)又都和雙股 DNA 斷裂的修補有關(Legrand et al., 2007; Uppuluri et al., 2007; Ciudad et al., 2004)。
以啤酒酵母做為研究 DNA 修復的模式生物,根據單突變和雙突變的 敏感性及上位基因(epistatic gene)相互作用的資料可將酵母細胞對射線敏 感的基因(radiation sensitive, RAD)分為三個上位顯性組(epistasis group),分 別是RAD3 組、RAD6 組和 RAD52 組(Zawel et al., 1995)。RAD3 組與 核 苷 酸 的 切 除 修 復 有 關 ,RAD3 蛋 白 是 切 割 階 段 需 要 的 一 種 解 旋 酶 (unwindase),與 RNA 聚合酶有關的轉錄機制有關。RAD6 組與複製後 的修復有關。RAD6 是染色體複製後的修復所必須,同時也涉及菌絲的形 成。Rad6 蛋白除了對紫外線的傷害有保護功能之外,也可以透過未知的反 應途徑抑制菌絲的成長;Rad6 蛋白的缺陷則會引起假菌絲的表現(Leng et
al., 2000)。RAD52 則與重組過程中 DNA 的修復有關。在啤酒酵母中,
RAD52 更是唯一在所有 HR 修復過程都需要的基因(Ciudad et al., 2004)。 白色念珠菌其他的 49 種和 DNA 修復相關的基因或 ORF 則都是從電 子註釋推斷而來(Inferred from Electronic Annotation, IEA),包括從資料 庫紀錄轉移的註釋或是經由 DNA 序列比對的結果來推測,目前都尚未有 經實驗證實的文獻記載。整體而言,目前對白色念珠菌的 DNA 修補機制 相關基因的研究仍相當欠缺。
1.4.
本論文之研究目標 DNA修補是一種生物對環境耐受度的表現(Sheedy et al, 2005),如果 能對白色念珠菌有關DNA修補的基因加以確認,能了解DNA修補和白色念 珠菌的型態改變的關聯性,或是能了解DNA修補和致病性或產生抗藥性有 關的作用機制而與以中止,應該可以對白色念珠菌疾病的治療提供更多的 選擇。實驗室先前運用相減雜交技術(suppression subtractive hybridization, SSH),共得到991個選殖體,用限制酶圖析(restriction mapping)的方式,將 有相同片段數量及大小的選殖體分組定序,接著將定序結果在Stanford''s Current Assembly of Candida albicans Sequence及NCBI中做比對,把所得到 的基因依照功能分類,共得到52個已知基因及48個未知基因(郭大榮, 2002)。本研究將實驗室先前所得之cDNA序列SB021的643個核酸序列在 CandidaDB 比對結果,發現其和有1821個鹼基對,推定可能和DNA修補有 關的orf19.2730相關度最高。故選擇以orf19.2730為標的,以啤酒酵母菌為表 現平台,以PCR的方式自白色念珠菌的Genomic DNA擷取此ORF片段及其 上游的promoter共2192個鹼基對,插入YEP363質體,轉殖到啤酒酵母。另 將自實驗室已構築的pET∆5T系列質體擷取HA3His6序列做為標籤,以西方 轉漬分析偵測是否有His及HA標誌的蛋白表現,目標在於能藉此確認 orf19.2730的表現,並進行功能性探討。
2.
二、材料
2.1. 菌株
2.1.1. Candida albicans SC5314:(Gillum et al., 1984)
為野生株白色念珠菌,染色體用於PCR 模板以選殖特定基因。
2.1.2. Escherichia coli :DH5α
作為質體增殖及保存之宿主。
2.1.3. Saccharomyces cerevisiae :
10560-2B:(Chen et al., 2004) Mating type 為 MATa
基因型為ura3-52;his3::hisG;leu2::hisG。 將質體YEP363 轉形至 10560-2B 中,命名為 SCY 將質體Ta2Y 轉形至 10560-2B 中,命名為 10560-2B-Ta2Y 將質體Ta2HAS 轉形至 10560-2B 中,命名為 10560-2B-Ta2HAS 將質體YpL7 轉形至 10560-2B 中,命名為 SCYL7 將質體YpL5 轉形至 10560-2B 中,命名為 SCYL5 將質體YpL2 轉形至 10560-2B 中,命名為 SCYL2 SLO69:(Chen et al., 2004)
Mating type 為 MATa
基因型為his3 : : hisG ura3–52;leu2 : : hisG;ade3 : : CDR1p-lacZ-LEU2
將質體Ta2YURA3 轉形至 SLO69 中,命名為 SLO69Tu 將質體YEP363URA3 轉形至 SLO69 中,命名為 SLO69Yu
2.2. 質體
質體 特性 Reference YEP363 在E. coli 的篩選標記為抗 Ampicillin, 在Saccharomyces cerevisiae 的篩選標 記為LEU2,在yeast 中為高複製倍數 的質體,MCS 後接 lacZ 為 Reporter gene。 Valenzuela et al., 1998 pRS426 篩選標記為抗 Ampicillin,MCS 位於 lacZ 中,可用於藍白篩選,其上有 T7 promoter。 Nakanishi et al., 2004pET∆5T-D24B-HAHis 篩 選 標 記 為 抗promoter 並帶有 HAHis-tag。 Kanamycin , 含 T7 楊昀良 實驗室 Ta2Y 將YEP363 中,篩選標記為抗 Ampicillin。 orf19.2730 及其上游序列置入質體 本實驗 Ta2R 將pRS426 中,篩選標記為抗 Ampicillin。 orf19.2730 及其上游序列置入質體 本實驗
Ta2Y∆stop 將 orf19.2730 去除 3’端中止碼後,置 入質體 YEP363 中,篩選標記為抗 Ampicillin。 本實驗 Ta2HAS 將Ta2Y∆stop 上 orf19.2730 的 3’端加 入HAHis-tag,破壞 lacZ,篩選標記為 抗Ampicillin。 本實驗
Ta2YURA3 將標記為抗pRS426 的 URA3 置入 Ta2Y,篩選Ampicillin。 本實驗 YEP363URA3 將選標記為抗pRS426 的 URA3 置入 YEP363,篩Ampicillin。 本實驗 YpL7 將端加入orf19.2730 上游的 371 個鹼基在 3’AUG 啟動碼後置入到 YEP363
的lacZ 前,篩選標記為抗 Ampicillin。 本實驗 YpL5 將orf19.2730 上游的 315 個鹼基在 3’ 端加入AUG 啟動碼後置入到 YEP363 的lacZ 前,篩選標記為抗 Ampicillin。 本實驗 YpL2 將orf19.2730 上游的 257 個鹼基在 3’ 端加入AUG 啟動碼後置入到 YEP363 的lacZ 前,篩選標記為抗 Ampicillin。 本實驗
2.3. 引子(primer)
註:方框為限制酶切位,灰階為外加之核甘酸,非位置欄所標記。
名稱 序列5’~3’ 位置
R158Hd30 TTGAAGCTTATACAACATTAATTATGCTTGTCG +1821~ +1796orf19.2730: F158Bm10 ATAGGATCCGTCAAATTTTCCAGTCTCA orf19.2730:-371 ~ -353 FYB20 ATAGGATCCATGCAATCAGTGTATCC orf19.2730:
+1 ~ +17 RYH20 CGGAAGCTTTACAACATTAATTATGCT orf19.2730: +1819 ~ +1801 FPH30 ATTAAGCTTTACCCATACGATGTTCCT pET∆5T-D24B-HAHis: +5864 ~ +5881 RPB32 TTAGCGCGCATATTTGAATGTATTTA pET∆5T-D24B-HAHis: +538 ~ +521 FPUR3 TGCGGATCCCTAAGAAACCATTATT pRS426: +5662 ~ +5677 RPUR3 ATTGGATCCTTAGTTTTGCTGGCC pRS426: +1220 ~ +1206 RH200 CGAAAGCTTTCATTGTGAATGTGTTGTTT orf19.2730: -16 ~ +3 FB371 TAAGGATCCGTCAAATTTTCCAGTCTCA orf19.2730: -371 ~ -353 FB315 ATAGGATCCGATGATGGCTCAACAAAC orf19.2730: -315 ~ -298 FB257 GCCGGATCCTCATTTACTAACAGAGTTGT orf19.2730: -257 ~ -238
2.4. 藥品試劑
z Amresco:
Acryl/Bis 37.5:1 solution (Cat.No.0254) Glycerol (Cat.No.0854-1L-PTM)
Phenol saturated solution (Cat.No.0945) Tris
z AppliChem:
Ampicillin (Cat.No.A2839) z Bio-Rad:
APS (Cat.No.161-0700)
Ethylenediaminetetraacetic acid (EDTA) (Cat.No.161-0729) z Difco:
Bacto agar (Cat.No.143175)
Yeast nitrogen base w/o amino acid (Cat.No.145368) YPD broth (Cat.No.135141XB)
Nutrient Broth (Cat.No.149018) z Fermentas:
T4 DNA ligase (Cat.No.1812) Restriction Enzyme:HindIII, PauI, z Fluka:
Acetic acid (Cat.No.33209) PMSF (Cat.No.78830) Urea (Cat.No.SK2644U) z GiBco BRL:
Goat serum (Cat.No.16210-072) z J.T.Baker:
Coomassie Brilliant Blue R-250 (Cat.No.F792-01) Dextrost (Cat.No.1916-01)
Triton® X-100 (Cat.No.X198-07) z Mallinckrodt: Methanol (Cat.No.3016-08) z MDBio,Inc: X-gal (Cat.No.613049) z Merck:
Dodecyl Sulfate Sat(SDS)(Cat.No.1.12012.0500) Ethanol (Cat.No.1.009833.2500)
Tris-HCl (Cat.No.1.01547.1000)
Sodium dihydrogen phosphate monohydrate (Cat.No.1.06346.0500) Di-sodium hydrogen phosphate dehydrate (Cat.No.1.06580.0500) Maleic acid (Cat.No.8.17058.1000)
β-mercaptoethanol (Cat.No.1.1543.0100)
z Midwest Scientific: X-film (Cat.No.LA7111) z NEB:
Restriction Enzyme:BamHI, HindIII, XhoI, BssHII, z Riedel-de Haën:
Chloroform (Cat.No.32211)
Sodium hydroxide (Cat.No.30620) Sodium chloride (Cat.No.31434) z Scharlau:
LB agar (Cat.No.01-385) LB broth (Cat.No.02-385) z Schleicher&Schuell
Nitrocellulose Transfer Membrane (Cat.No.10401396) z SibEnzyme:
1kb DNA ladder (Cat.No.SEM11C001) z Sigma:
Crystal Violet (Cat.No.C-3886) EtBr (Cat.No.E-7637)
Glass Beads (Cat.No.G-9268) Kanamycin (Cat.No.K4000) Lithium acetate (Cat.No.L-6883) Uridine (Cat.No.U-0750)
L-Histidine (Cat.No.H-8125)
Polyethylene Glyco3350 (Cat.No.P-4338)
polyoxyethene-sorbitan monolaurate(Tween20) (Cat.No.p-1379) Phenol (Cat.No.P-4682)
TEMED (Cat.No.T-9281) z TBB:
Protein marker (Cat.No.0901) z VEGONIA:
Agarose (Cat.No.9201-05)
2.5. 緩衝溶液及溶液
z 50X TAE buffer48.4 g Tris base,0.5 M EDTA(pH 8.0)20 ml,11.42 ml acetic acid added ddH2O to 200 ml
z 5M EDTA stock solution
186.1 g EDTA added ddH2O to 800 ml(pH 8.0)
z TE buffer
10 mM Tris-HCl(pH 8.0),1 mM EDTA(pH 8.0) z LATE buffer
0.1 M Lithium, 10 mM Tris-HCl (pH 7.5), 1 mM EDTA z PLATE buffer
40% polyethylene glycol3350 in LATE buffer
z 1M 10X LiOAC
40.8 g Lithium Acetate added ddH2O to 400 ml (pH 7.5)
z 50% PEG
75 g polyethylene glycol3350 added ddH2O to 150 ml
z 40% dextrose
40 g Dextrose added ddH2O to 100 ml
z Shearing buffer
100 mM NaCl, 10 mM Tris, 1 mM EDTA, 0.1%SDS, pH=8.0 z Breaking buffer
0.1 M Tris-HCl, 20% glycerol (v/v), 1 mM Dithiothreitol z Z buffer
Na2HPO4 7H2O 16.1 g, NaH2PO4 H2O 5.5 g, KCl 0.75 g, MgSO4 7H20 0.246 g,
β-Mercaptoethonol 2.7 ml added ddH2O to 1000 ml pH 7.0
z Lysis buffer
2% Triton X-100,1% SDS,100 mM NaCl,10 mM Tris-HCl, pH 8.0,1 mM EDTA, pH 8.0
z 10X SDS-PAGE running buffer
0.25 M Tris base,1.92 M Glycine,1% SDS z 2X SDS-PAGE loading buffer
0.5% bromphenol blue,0.5 M Tris-HCl(pH 6.8),10% SDS,100% glycerol z 0.25% Coomassive blue stain solution
2.5 g Coomassive brilliant blue,50% methanol,10% acetic acid added ddH2O
z Destain solution
30% methanol,10% acetic acid z 10X transfer buffer
39 mM Glycine,48 mM Tris base,10% SDS,20% methanol z Blocking buffer
2.5 g nonfat powdered milk dissolved in 50 ml 1X TBS buffer z TBS buffer
10 mM Tris(pH 8.0),150 mM NaCl z TBST buffer
10 mM Tris(pH 8.0),150 mM NaCl,0.05% Tween 20
2.6. 培養基配製
z LB(Luria-Bertni)培養液
1% tryptone,0.5% yeast extract,1% NaCl z LB(Luria-Bertni)/Ampicillin 培養基
1% tryptone,0.5% yeast extract,1% NaCl,1.5% agar,50 µg/ml Ampicillin z LB(Luria-Bertni)/Kanamycin 培養基
1% tryptone,0.5% yeast extract,1% NaCl,1.5% agar,50 µg/ml Kanamycin z YPD 培養液
2% Bacto-peptone,1% yeast extract,2% dextrose z YPD 培養基
2% Bacto-peptone,1% yeast extract,2% dextrose,2% agar z SD 培養液
0.67% Baco-yeast nitrogen base w/o amino acid,2% dextrose z SD 培養基
0.67% Baco-yeast nitrogen base w/o amino acid,2% dextrose,2% agar z SD/Uridine/ Histidine 培養基
0.67% Baco-yeast nitrogen base w/o amino acid,2% dextrose,2% agar, 80 mg/l uridine,20mg/l histidine
z SD/ Histidine 培養基
0.67% Baco-yeast nitrogen base w/o amino acid,2% dextrose,2% agar, 20 mg/l histidine
z Solid Spider 培養基
10 g of nutrient broth,10 g of mannitol, 2 g of K2HPO4 and 13.5 g of agar in
1 liter H2O
2.7. 儀器設備
-20℃直立冷凍櫃(WHITE-WESTINGHOUSE) 4℃三門冰藏櫃 KS-101-MS(MINI KINGDOM) -80℃超低溫冷凍櫃 925/926(FORMA SCIENCE) 水平式電泳槽MJ-105(MEDCLUB)震盪器VORTEX-GENIE G560(SCIENTIFIC INDUSTRICS) 水平式震盪器S-101(FIRSTEK)
試管震盪器VXR S1(IKA-VIERAX-VXR) 加熱攪拌器PC-420(CORNING)
直立式電泳槽(BIO-RAD)
半乾式電泳轉印槽TRANS-BLOT® SD CELL 221BR(BIO-RAD) 往復式恆溫水槽B206-T1(FIRSTEK SCIENTIFIC)
恆溫式震盪培養箱S300R(FIRSTEK SCIENTIFIC)
核酸計算儀GeneQuant pro(AMERSHAM PHARMACIA BIOTECH) 桌上型低溫高速離心機Centrifuge 5804 R(eppendorf)
桌上型高速離心機5100(KUBOTA CORPORATION) 微量離心機MICRO 240A(DENVILLE SCIENTIFIC INC.) 桌上型微量離心機Biofuge(KENDRO)
乾燥加熱板VH-01(VIOLET BIO SCIENCE)
程式溫度控制儀PTC-100TM(MJ RESEARCH INC.) 電泳影像處理系統GEL DOC 2000(BIO-RED) 酸鹼值檢測器Φ360(BECKMAN)
電子天平PB153-S(METTLER TOLEDO) 電子防潮箱DX106(台灣防潮科技)
倒立顯微鏡CK40(OLYMPUS) 數位相機C-5050Z(OLYMPUS)
3.
三、實驗方法
3.1. Bust n’ Grab Protocol 快速抽取 DNA(Harju et al., 2004)
取Candida albicans 在 5ml YPD 培養液中以 37℃,200 轉培養 18小時,取1.5 ml 菌液加入 eppendorf 中,以 13000 rpm 離心 5 分鐘,移 除上清液。加200 µl lysis buffer 沖起沉澱,震盪 1 分鐘,移入-80℃急 凍 2 分鐘,再移入 95℃乾熱,重複操作後震盪 30 秒。加 200 µl Chloroform,震盪 2 分鐘,再 13000 rpm 離心 3 分鐘後,取出上清液。 加400 µl ice-cold 100% ethanol,輕微震盪,移入-20℃靜置 5 分鐘。以 13000 rpm 在 4℃離心 5 分鐘後移除上清液。再加入 500 µl 70% ethanol, 以13000 rpm 在 4℃離心 5 分鐘,移除上清液。以 60℃乾熱 5 分鐘去除 殘留ethanol,以 25 µl 1X TE 回溶,在 4℃保存。
3.2. 以 Excel Pure Plasmid mini kit 抽取 E. coli 質體
YEP363 以加入 5 µl Ampicillin 的 LB 培養液,pET∆5 以加入 25 µ l Kanamycin 的 LB 培養液,在 37℃,200 rpm 培養 17 小時。取 1.5 ml 菌液離心1 分鐘,去除上清液。加 200 µl Solution I 沖起沉澱,再依序 加200 µl Solution II,輕微震蕩;加 200 µl Solution III,輕微震蕩;以 以13000 rpm 在 4℃離心 5 分鐘取上清液。將 Spin Colum 和 collection tube 組合,倒入上清液,以 13000 rpm 離心 1 分鐘後倒掉濾液。加 700 µl Washing Solution,以 13000 rpm 離心 1 分鐘,倒掉濾液,重複一次之 後再以13000 rpm 離心 3 分鐘,將 Spin colum 移入 Eppendorf,於 60℃ 加熱板乾熱5 分鐘去除殘留 ethanol,加 50 µl Elution Solution,以 13000 rpm 離心 1 分鐘,保留濾液。
3.3. 聚合酶連鎖反應(PCR)
進行一般PCR 反應在 0.5 ml 的微量離心管內加入 1 unit(U)的 Taq polymerase(5U/µl)、 5 µl 10X PCR buffer,各 1 µl primerd(50 µM),4 µl dNTPs mixture(2.5mM), 0.1 µg Template DNA,再加入二次無菌水將體積調整到 50 µl,置於 PCR 溫度控制儀內進行聚合連鎖反應。待反應完成後,以洋菜膠電泳確認 PCR 產物的大小。 設定PCR 的溫度控制 1. 94℃,3~5 分鐘 2. 94℃,Denaturation,1 分鐘 3. 50~55℃,Annealing,1~1.5 分鐘 4. 72℃,Extension,1 分鐘 5. 重複 2~4 步驟,循環 25~30 回 6. 72℃,最後一回 Extension,5 分鐘 7. 4℃,停止反應。
PCR Clean Up (by Excel Band PCR)
將 PCR 產物移入 Eppendorf,加 500 µl Binding Solution,震盪混合, 裝進組合好的spin colum,以 13000 轉離心 1 分鐘,移除濾液。再加 700 µl Washing Solution,以 13000 轉離心 1 分鐘,移除濾液,重複一次後, 再以 13000 轉離心 3 分鐘。將 Spin colum 裝入新的 Eppendorf, 60℃ 乾熱5 分鐘。加 30 µl Elution Solution,以 13000 轉離心 1 分鐘後,保 存於-20℃。
3.4. PCR 產物的 T-vector 檢驗
Ligation將 0.5 unit 的 T4 DNA ligase、1~2 unit 的 10X buffer、PCR 產生的 DNA 片段和T-Vector(pGEM®-T)以莫爾濃度 3:1 混合於 1.5 ml 微量離心管內, 總反應體積為10~20 µl,於 14 ℃ 水浴 18 小時。
將 Ligation 產物移入 65℃加熱板 10 分鐘以停止反應。將勝任細胞 (competence cell)移入冰桶解凍 3 分鐘。取 3 管 50 µl 的勝任細胞分別加 入plus control 2.5 µl、Background Control 2.5 µl 及 PCR 產物 2.5 µl。冰浴 15 分鐘後移入 42℃水浴槽 heat shock 45 秒。再冰浴 15 分鐘。分別滴在含 Ampicilin 之 LB 培養盤,以沾酒精加熱滅菌之玻棒冷卻後均勻塗到乾燥。 在37℃培養 18 小時後,挑取菌落加入含 Ampicillin 之 LB 培養液,在 37℃, 200 rpm 培養 18 小時,挑選質體大於 T-Vector(3KB)者進行 Restriction Enzyme digestion,確認切割後的質體片段大小。
3.5. 限制酶反應(Sambrook et al, 1989)
取適量 DNA(約 0.5~10 µg;適反應需要)到適量反應體積(30 µl) 以限制酵素切割。酵素的用量及反應溫度、時間都依照廠商所提供的條件 資料進行。反應完後,利用洋菜膠體電泳分析。所需的 DNA 片段在經限 制酵素切割後,以Gel Extraction System 或 PCR Clean-up Kit 來純化 DNA 並去除限制酵素及鹽類。3.6. 萃取洋菜膠內之 DNA 片段
配製 0.8% 洋菜膠,稍微冷卻後,加 30 µl crystal violet 搖動錐型瓶充 分混合,等待凝膠。樣品加入專用 6X dye,跑膠到 band 明顯分離。切取 含有目標band 的部份。以 Wizard® S. V. Gel & PCR Cleanup System 進行 Cleanup。先計算出洋菜膠重。以 10 µl solution : 10 mg gel 的比例加入 Membrane Binding Solution,在 60℃加熱板加熱並震盪到膠完全熔化。組 裝S. V. minicolum 到 collection tube。將熔膠混合液置入 minicolum,室溫 下靜置1 分鐘。以 13000 rpm 離心 1 分鐘,移除濾液。加 700 µl Membrane Wash Solution,以 13000 rpm 離心 1 分鐘去除濾液。再加 500 µl Membrane Wash Solution,以 13000 rpm 離心 1 分鐘,倒掉濾液。以 13000 rpm 離心 3 分鐘,將 Minicolum 移到新的 Eppendorf,在 60℃加熱板靜置 5 分鐘。加 50 µl Nuclease-Free Water,室溫靜置 1 分鐘,以 13000 rpm 離心 1 分鐘, 移去Minicolum,於-20℃儲存。
3.7. E. coli 的連接反應(Ligation)和轉殖作用(Transformation)
Ligation:DNA 片段(Insert DNA)和載體(Vector)在加入 0.5 unit 的 T4 DNA ligase 和適量緩衝液的作用下進行接合反應,反應體積 20 µl。其中 DNA 片段和載體的莫爾濃度比為3:1,於 14℃水浴培養 18 小時。 Transformation: 將連接反應的產物移入 65℃乾熱 10 分鐘以停止反應。將勝任細胞移 入冰桶解凍3 分鐘。取 50µl 勝任細胞加 2.5 µl 連接反應後的 DNA。冰浴 15 分鐘後移入 42℃水浴槽 heat shock 45 秒後再冰浴 15 分鐘。菌液滴在含 Ampicilline 之 LB 培養盤,以沾酒精加熱滅菌之玻璃棒均勻塗布到乾燥, 置於 37℃恆溫箱中培養 18 小時。挑取菌落加入含 Ampicilline 之 LB 培養 液,在37℃,200 rpm 培養 18 小時後抽質體進行限制酶反應確認。
3.8. 真菌的轉殖作用(Yeast transformation)
取啤酒酵母 在 5 ml YPD 培養液,30℃,150 rpm 培養 18 小時後轉養 到15 ml YPD 培養液,30℃,150 rpm 至 OD 600>0.9,移置 15 ml 離心管, 室溫下以2500 rpm,離心 5 min,倒去上清液後加 4 ml LATE buffer,室溫 下2500 rpm,離心 5 min,去除上清液後加 1ml LATE buffer 懸浮菌液,移 入eppendorf tube,室溫下靜置 10 分鐘即為勝任細胞。取 4 µl 10mg/ml 的 Salmon Sperm DNA 在 95℃乾熱 1 分鐘立即冰浴 1 分鐘,連同欲轉形的質 體 DNA 2µl (約 0.5-1 µg)一併加入 100 µl 的勝任細胞。混合均勻後加入 700 µl PLATE buffer,在 30℃,150 rpm 振盪培養 30 分鐘後,置於 42℃水浴 槽Heat shock 7 分鐘,再冰浴 2 分鐘。以 2500 rpm 離心 5 min 去除上清液, 加入200 µl 1X TE buffer 懸浮菌體,將菌液塗抹到適當的 SD 培養基,在 30℃恆溫培養 3 天,挑選適當之菌落。3.9. 抽取啤酒酵母質體
取啤酒酵母在加入適當營養基的 10 ml SD 培養液以 30℃,150 rpm 培 養2 天後,以 3000 rpm 離心 5 分鐘,除去上清液。加入 5 ml d.dH2O 重新
懸浮菌液,以3000 rpm 離心 5 分鐘後除去上清液,倒置吸乾。加入 200 µl Shearing buffer,再倒入經 acid-washed 過的玻璃珠至液面下。在 4℃高速震 盪1 分鐘,接著靜置 1 分鐘,重複 4 次;加入 200 µl TE 震盪 10 秒鐘後以 3000 rpm 於 4℃低溫離心 5 分鐘,取上清液至預冷之 Eppendorf,加入同體 積Phenol: Chloroform: isoamylalcohol=25: 24: 1 混合,在室溫下震盪 60 秒, 再以13000 rpm 離心 1 分鐘,取上清液。
3.10. 萃取真菌細胞萃取液
取啤酒酵母在加入適當營養基的 10 ml SD 培養液以 30℃,150 rpm 培 養到OD600= 1.5~1.6,取 1.5ml 菌液轉養至新鮮 15~30 ml 加入適當營養基 的SD 培養液,以 30℃,150 rpm 培養到 OD600=1.5~1.6,取全部菌液在 4 , ℃ 5000 rpm 離心 5 分鐘,去除上清液。加入 10 ml Z buffer 清洗細胞,轉入 15 ml 離心管,以 5000 rpm 離心 5 分鐘後去除上清液。加入 500 µl Breaking buffer 懸浮菌體,再加入 25 µl PMSF(40 mM) ,倒入玻璃珠至液面下,於 4℃高速震盪 15 秒,靜置 15 秒,重複 5 次,使細胞完全破裂。加入 250 µl Breaking buffer,於 4℃高速震盪 10 秒後高速離心 15 分鐘,取上清液即為 細胞萃取液,於-20℃保存。3.11. SDS PAGE 進行 Coomassie blue stain 測試
製膠: 組裝製膠組件,玻璃板先用 70%酒精噴洗擦乾淨,確認不留纖維屑後, 組合在製膠架上,注意玻片要垂直壓於泡棉墊上以免發生漏液。先配製下 層14%膠,用塑膠滴管導入玻璃模版到固定夾上緣高度,上方以 Isopropanol 壓平,待30 分鐘凝膠後,注水除去 Isopropanol,用注射筒以針頭抽出水, 重複洗滌。再配製上層4%膠,塑膠滴管導入注入到滿,插入 Comb,待凝膠後用保鮮膜加水包裹備用。 膠體電泳: 先以 1:9 的比例混合 10X ß-M. E. ( ß-mercaptoenthanol )和 2X SDS loading dye,再加入等體積的蛋白質萃取液,室溫下靜置 30 分鐘等待變性 (denature)後注入膠體準備電泳。先以 120 V 電泳 20 分鐘待 band 壓成一 線,再以100 V 電泳 2~5 小時,到目標區的 band 明顯分離。
Coomassie blue stain:
取下玻片,截去上層膠,在下層膠片上滴足量的水保持濕潤,剝下膠 片移入Coomassie blue stain。以 40rpm 搖動 20 分鐘染色。回收 Coomassie blue stain 後,膠片移入塑膠盒,加入 Destain 淹沒膠片到可搖動高度,退 染到band 清晰且膠片無色,以清水清洗數回。 收膠: 取玻璃紙充分浸濕,平鋪在厚玻片上,去除氣泡。將退染後的膠片平 鋪在玻璃紙上,壓去氣泡,加水保持濕潤。取另一浸濕的玻璃紙平鋪在膠 片上,壓去氣泡。以4 個夾子在四周夾住固定,垂直放置待完全乾燥即可 收存觀測。
3.12. 西方轉漬分析(Western Blotting)
Transfer:將濾紙及 Nitrocellulose membrane 在 1x transfer buffer 浸泡 10 分鐘。 裝配Bio-Rad Trans-Blot® SD CELL 221BR semidry。由下而上依序鋪二層 濾紙、Nitrocellulose membrane、SDS-PAGE 電泳膠體、上層再鋪一層濾紙, 充分淋澆 buffer;用試管滾壓驅除氣泡後再滴加 buffer。架上電極板,以 0.09 A 電泳 40 分鐘,使膠體上的 protein 轉漬到 Nitrocellulose membrane 上。
夾去上層濾紙及 SDS-PAGE 電泳膠體,將 Nitrocellulose membrane 移入 20 ml Blocking buffer,於室溫下,40 rpm 平面震盪 1 小時。
Detection:
將 Nitrocellulose membrane 置入塑膠袋,加入含稀釋 2000 倍 Anti-His 抗體(一抗)的2ml Blocking buffer,壓出氣泡後封口。用玻片包夾固定, 平放在 Waver,35 rpm 平面震盪 1 小時。夾起 membrane 浸入 20 ml 的 TBST,以 55 rpm 平面震盪 5 分鐘,重複清洗 3 次。再夾起 membrane 置 入塑膠袋,改加入含稀釋 8000 倍 Anti-His 抗體(二抗)的 2ml Blocking buffer,壓出氣泡後封口。用玻片包夾固定,平放在 Waver,35 rpm 平面震 盪1 小時。夾起 membrane 浸入 20 ml 的 TBST,以 55 rpm 平面震盪 5 分 鐘,重複清洗3 次。最後將 800 µl 的 ECL substrate(Pierce)均勻加到膜 上,在暗房內以 X 光底片進行壓片 1 至 5 分鐘,取出底片浸入顯影液 (Develop)搖晃約 30 秒,將底片換到清水中漂洗 30 秒,接著浸入定影液 (Fixer)搖晃約 1 分鐘到底片變黑,以大量清水沖洗底片,將底片晾乾保 存。
3.13. Yeast β-galactosidase filter assay
將欲測的菌體畫在同一培養基上培養 3 天,取滅菌過的 colony filter 直接覆蓋於培養盤的菌體上3 分鐘,將菌落轉印至濾紙,置入新的培養基 中,擠出氣泡,於30℃培養一天。將帶有 colonies 的濾紙菌體面向上,放 入液態氮1~2 分鐘破壞菌體後取出,置於室溫下 1 分鐘使菌體完全破裂。 另取assay filter 置於乾淨培養皿,加入適量(約 1 ml) X-gal / Z buffer (Z buffer + 1 mg / ml X-gal)充分浸潤,濾紙不可過於潮濕,以免菌體漂浮。 將處理好之含菌濾紙菌體面朝上覆蓋在其上方,濾紙間避免氣泡出現,置 於30℃呈色。
3.14. 侵犯力分析(invasion)
培養液中添加uridine,於 37℃恆溫培養 7 天,觀察菌落型態後,以水流沖 洗菌落,觀察菌落是否因形成菌絲侵入培養基而殘留。
4.
四、結果
4.1. 白色念珠菌 Genomic DNA 的資料庫分析
將實驗室先前所得之 Candida albicans 的 cDNA 序列 SB021 的 643 個 核酸序列(附錄一)在 CGD 資料庫(http://www.candidagenome.org),選擇 Assembly 20 資料,以 Washington University BLAST (WU BLAST) version 2.0 程式作比對,可推定得兩個可能,其一為 orf19.2730,積分為 1731(Score = 265.8 bits),E-value= 1.3e-74(附錄二)。另一為 orf19.2743,積分為 1722( Score = 264.4 bits),E-value= 3.3e-74(附錄三)兩者都是由 1821 個鹼基所構成,且都位於第4 對染色體上。其中 orf19.2730 共有 1821 個鹼 基,可決定607 個胺基酸,而 orf19.2743 同樣有 1821 個鹼基,也決定 607 個胺基酸。將兩者的鹼基序列作比較,共有34 個分散的鹼基不同,而再將 它們所編碼的胺基酸做比較,則可發現有10 個胺基酸不同(附錄二、三)。 對兩者的功能做查詢,發現都是推測對應到啤酒酵母菌的 RPH1 基因 (YER169W),和 DNA 修補有關,且兩者也都是未經實驗證實的電腦資 料庫推定結果。將相同的序列在 BRI、CandidaDB 等資料庫比對,結果相 似,兩者應是同源基因。由於兩者差異不大,故依資料庫比對積分最高而 E 值最低的原則評估,選擇以 orf19.2730 做為研究對象。 SB021 的核酸序列在 orf19.2730 有 2 個對應區域。其中一個區域可在 orf19.2730 的第 99 至 503 個鹼基的範圍對應到的 415 個鹼基,有 92%的 identity,3%的 gaps。另一個區域可在 orf19.2730 的第 1 個鹼基起始的範圍 對應到148 個鹼基,有 80.8%的 identity,其中有 9.8%的 gaps(圖 1)。將 此序列貼到CandidaDB 資料庫(http://genolist.pasteur.fr/CandidaDB),分別選擇 以NCBI Blast V2 和 WU BLAST version 2.0 程式進行比對,結果積分最高 的都是指向 IPF15357,即 orf19.2730。兩個資料庫比對的結果一致,故推 定此序列屬於 orf19.2730 的一部份。 依 CGD 資 料 庫 比 對 的 結 果 , orf19.2730 在 Assembly 19 , 位 於 Contig19-10158 上第 139713 個鹼基到第 137893 個鹼基之間,其對偶基因 orf19.10244 則位於 Contig19-20158 上,起始於第 132773 個鹼基,終止於 第130953 個鹼基,共有 1821 個鹼基,可決定 607 個胺基酸。依 Assembly21 定義,orf19.2730 位於 Candida albicans 的第 4 對染色體上,起始於第 553936
個鹼基而終止於第 555756 個鹼基(圖 2)。對 orf19.2730 及其上游序列的限 制酶切割部位分析,發現其中沒有HindIII 及 BamHI 的切位(圖 3),而這兩
個限制酶在質體 YEP363 和質體 pRS426 上都恰有一個切位,故在 5’端引 子加上 BamHI 序列(GGATCC),而在 3’端引子加上 HindIII 序列
(AAGCTT)作為切位。引子分別位於 orf19.2730 上 5’端的 -371 至 -353 個鹼基及3’端+1821 至 +1796 個鹼基的位置。
4.2.
將 Candida albicans Genomic DNA 的 orf19.2730 及其上游序
列轉殖到
YEP363 和 pRs426 質體
以 R158Hd30 和 F158Bm10 為引子在 Candida albicans 的 Genomic DNA 上以 PCR 方式切取長度為 2192 個鹼基的 orf19.2730 及其上游序列,經膠 體電泳顯示,切取的片段大小介於2000 個鹼基到 2500 個鹼基之間,大小 符合預期(圖 4),暫以 Ta1 表示。將 Ta1 連結在大小為 3000 個鹼基的 T-vector 上增殖後,以限制酶 HindIII 和 BamHI 切下。另外以 HindIII 和
BamHI 切取有 8588 個鹼基的質體 YEP363 和有 5726 個鹼基的質體
pRS426。將切割後的 YEP363 和 Ta1 連結建構大小為 10764 個鹼基的質體, 暫以Ta2Y 表示。因為 YEP363 質體本身沒有 XhoI 的切位,而 Ta1 上則有 2 個切位,若以 XhoI 對 Ta2Y 進行切割,應可得到大小為 1096 個鹼基和 9668 個鹼基的兩個片段(圖 5)。將切割後的 pRS426 和 Ta1 連結後建構 大小為7892 個鹼基的質體,暫以 Ta2R 表示。其中 pRS426 質體本身有一 個 XhoI 的切位,而 Ta1 片段上有 2 個切位。若以 XhoI 對 Ta2R 切割,應
可得到大小為282、1096 和 6514 個鹼基的三個片段(圖 6)。將質體分別 轉殖至勝任細胞中培養,抽取質體後以XhoI 限制酶切割,經膠體電泳驗證 的結果如圖 6。依電泳結果顯示,Ta2Y 經限制酶 XhoI 切割後產生兩個片 段,其中y1 片段介於 8000 到 10000 個鹼基之間偏向 10000 鹼基,y2 片段 介於1000 到 1500 個鹼基之間偏向 1000 鹼基,和預期的 9668 個鹼基及 1096 個鹼基兩片段應屬符合。Ta2R 經限制酶 XhoI 切割後產生三個片段,r1 片 段介於6000 到 8000 個鹼基之間,r2 片段介於 1000 到 1500 個鹼基之間略 偏向1000 鹼基,r3 則介於 200 到 500 個鹼基之間,和預期中的 6514、1097 及281 個鹼基的三片段亦屬相當,應可確定 Ta1 已分別接合到 YEP363 質
體及pRS426 質體上,建構成 Ta2Y 和 Ta2R 質體(如圖 7)。
4.3.
將
Ta2Y 質體轉殖到啤酒酵母菌建構 10560-2B-Ta2y
培養啤酒酵母菌 10560-2B,製作勝任細胞,並將先前抽取之 Ta2Y 質 體轉殖到勝任細胞中。因Ta2Y 質體上含有 LEU2 基因,可表現 β-isopropyl malate dehydrogenase,此酵素與 pyruvate 轉化為 leucine 的過程相關。啤酒 酵母菌若經轉殖成功,則帶有Ta2Y 質體,可以生長在不含 leucine 的培養 基中,故以不含 leucine 的 SD/Uridine/Histidine 培養基篩選,在 30℃恆溫 培養3 天後,挑出能夠生長的菌落,轉養到 SD/Uridine/ Histidine 培養液中, 以30℃,150 轉震盪培養 17 小時後抽取真菌質體,經酒精沉澱後,以未經 限制酶切割的圓形 Ta2Y 質體當參考指標,與從轉殖後的啤酒酵母菌質體 中抽出的抽取液作比較,不經限制酶處理進行膠體電泳。結果如圖 8,可 看到選殖株的抽取液有明顯的band 出現在和 Ta2Y 質體相當的區域,而未 經轉殖的啤酒酵母菌抽出液則沒有,應可初步確認 Ta2Y 已成功轉殖到啤 酒酵母菌中,建構10560-2B-Ta2Y。
4.4. Coomassie blue staining
將啤酒酵母菌及 10560-2B-Ta2Y 細胞打破,分別萃取上清液,經 SDS-PAGE 電泳後,進行 Coomassie blue staining 比對試驗。orf19.2730 大 小為1821 個鹼基,如果能夠表現,其所合成的蛋白質應有 607 個胺基酸, 大小約為67 kDa,預期應在 55~72 kDa 的區域內出現目標。經比對結果如 圖 9,在 55~72 kDa 的目標區域內隱約觀察到 10560-2B-Ta2Y 上清液有 band 出現,而啤酒酵母菌的上清液則沒有,大致符合預期,但表現量較低, 需進一步以Western bloting 佐證。
4.5.
建構
Ta2HAS 質體
端終止碼的 orf19.2730 片段,暫名為 orf19.2730∆stop。設計引子時已在其 5’端加入 BamHI 切位,而 3’端則在去除終止碼之後的位置加入 HindIII 切 位,在兩個限制酶的切位之間可夾出大小為1818 個鹼基的片段。經膠體電 泳顯示,PCR 放大的產物在 1500 個鹼基到 2000 個鹼基的區間內出現 band,位置略偏上方,和預期 1818 個鹼基大小的區域符合(圖 10)。將 orf19.2730∆stop 連結在 T-vector 上增殖後,將質體抽出,再以 BamHI 和
HindIII 切割質體,可以得到大小為 1818 個鹼基和 3000 個鹼基的兩條片
段。同時也對YEP363 質體用 BamHI 和 HindIII 切割,可得 8568 個鹼基和 20 個鹼基的二個片段。以膠體電泳檢視,結果如圖 11,其中 O3、O4 為對 orf19.2730∆stop 切割的結果,可見到在 3000 個鹼基及 2000 個鹼基的附近 有明顯的band。而 Y1、Y2 為 YEP363 質體切割後的結果,在 8000 到 10000 個鹼基的區域內出現band,兩者都符合預期。
接著以 gel extraction 取出 1818 個鹼基的 orf19.2730∆stop 片段和 YEP363 切割後所得的 8568 個鹼基片段,兩端分別含有 BamHI 和 HindIII 的接合位置,將兩者進行接合,經過轉殖選殖後,在 YEP363 質體的基礎 上加入去除終止碼的 orf19.2730,建構大小為 10392 個鹼基的質體,暫以 Ta2Y∆stop 命名。Ta2Y∆stop 保留原本 YEP363 質體所具有的 lacZ
、
LEU2、
2µ 及 Apr 基因,也具有 orf19.2730 上限制酶 XhoI 的兩個切位,分別在第
480 個鹼基和第 1576 個鹼基的位置(圖 12)。YEP363 質體本身沒有 XhoI 的切位,而轉殖後的質體則有 2 個 XhoI 的切位,若以 XhoI 對 Ta2Y∆stop 切割,應可得1097 個鹼基和 9295 個鹼基的兩個片段,經膠體電泳結果如 圖 13。圖中 Ye 為 YEP363 質體,Yx 為 YEP363 經限制酶 XhoI 處理後的 結果,兩者沒有顯著差異,可確認限制酶XhoI 不會對 YEP363 質體發生作
用,所以 YEP363 質體沒有被切割。而 Xo1 和 Xo2 為限制酶 XhoI 對 Ta2Y∆stop 質體作用後的結果,可觀察到有兩條 band 出現,其中一條約在 1000 個鹼基附近,另外一條則在 10000 個鹼基附近,兩條 band 分佈的位 置恰好和預期的1097 個鹼基和 9295 個鹼基的預期位置相當,應可確認成 功建構Ta2Y∆stop 質體。 自實驗室取得含有 pET∆5T-D24B-HAHis 質體(大小為 6310 個鹼基)之 菌種,以加入25µl Kanamycin 的 5ml LB broth 震盪培養 18 小時後抽取質 體,再以RPB32 和 FPH30 為引子,用 PCR 的方式自 pET∆5T-D24B-HAHis 質體擷取HA3His6 及其下游 872 個鹼基的片段。起自第 5864 個鹼基,終
止於第538 個鹼基,中間跨越第 6310 個鹼基,大小共 985 個鹼基。加上先 前設計引子時在此片段的 5’端加入限制酶 HindIII 的切位,而在 3’端加上 限制酶 BssHII 的切位,建構成大小為 996 個鹼基的片段,暫名為 pHas。
將此PCR 的產物以膠體電泳檢視,結果如圖 14。在 1000 個鹼基附近的區 域可看到明顯的 band 出現,驗證 PCR 的產物大小符合預期。再以相同的 引子對pHas 進行 PCR 增殖,以限制酶 HindIII 和 BssHII 進行切割,經 gel extraction 純化。另外再以限制酶 HindIII 和 BssHII 切割 Ta2Y∆stop,同時 也會把 HindIII 之後 lacZ 基因的部份片段切除。經膠體電泳後以 gel
extraction 擷取大小約為 8890 個鹼基的片段,將這個片段和處理過的 pHas 片段進行接合作用,經過轉殖選殖後,建構帶有 HA3His6,大小為 9880 個鹼基的質體,暫名為 Ta2HAS(圖 15)。其中 HA3His6 正好接續在去 除終止碼的orf19.2730 序列之後,兩者間恰有 6 個鹼基的 HindIII 切位,如 果orf19.2730 能夠表現,則 HA3His6 也將能夠表現而轉譯出胺基酸。而原 來存在於YEP363 質體上的 lacZ 基因則遭到破壞而被去除。
Ta2HAS 質體上有三個 XhoI 的切位,比 Ta2Y∆stop 多一個。經限制酶
XhoI 處理後,應可切出大小為 551、1097 及 8232 個鹼基的三個片段。由 圖 16 可見 Ta2HAS 質體經限制酶 XhoI 切割後再經膠體電泳,果然在 500~750、1000~1500 及 8000~10000 個鹼基之間確有三條 band 出現,其片 段數及大小都和預期的結果相符,應可確認 pHas 已成功和 Ta2Y∆stop 接 合,建構帶有HA3His6 的 Ta2HAS 質體。
4.6.
將
Ta2HAS 質體轉殖到啤酒酵母建構 10560-2B-Ta2HAS
將啤酒酵母菌種菌到 5ml YPD 培養液中,以 30℃培養 17 小時後再轉 養5 小時,製作勝任細胞。將先前抽取之 Ta2HAS 質體轉殖到勝任細胞中, 建構啤酒酵母菌10560-2B-Ta2HAS。其中所含的 Ta2HAS 質體已將原來存 在於 YEP363 質體上的 lacZ 基因破壞,但仍含有 2µ 及 LEU2 基因,故仍 可以生長在缺少Leucine 的 SD/Uridine/Histidine 培養基中。以 SD/Uridine/ Histidine 培養基於 30℃培養 3 天後進行選殖,挑出能夠生長的菌落,種菌 到SD/Uridine/ Histidine 培養液中,再經 30℃,150 轉震盪培養 17 小時, 抽取真菌質體並經酒精沉澱。觀察圖17 的結果,圖中 TH 是未經切割的圓形Ta2HAS 質體。#6 和#7 則是從 10560-2B-Ta2HAS 中抽出的真菌質體。 經過膠體電泳後,可以在 5000 到 6000 個鹼基的範圍內觀察到都有 band 出現。但因為Ta2HAS 是環狀質體,電泳時移動的速率會比相同大小的線 形鹼基片段還快,所以未經切割則無法明確認定其大小的落點位置。再以 抽取的#6 和#7 真菌質體當做板模,對包含 HA3His6 及其下游序列共 996 個鹼基的pHas 片段進行 PCR 增殖。純化後得到#6P 和#7P。經膠體電泳可 觀察到在接近1000 個鹼基的位置有明顯的 band 出現,大小和 996 個鹼基 的 pHas 片段相仿,初步推測 pHas 片段已被成功的增殖。而 pHas 片段原 來並不存在於啤酒酵母菌中,至此應可確認帶有 pHas 的 Ta2HAS 質體已 被轉殖到啤酒酵母菌10560-2B 中而成功建構了 10560-2B-Ta2HAS。
4.7.
Western bloting
培養啤酒酵母菌及 10560-2B-Ta2HAS 細胞,將細胞打破後萃取上清液 進 行 Coomassie blue staining 比 對 試 驗 及 Western bloting 。 因 為 orf19.2730∆stop 本身大小為 1818 個鹼基,加上限制酶 HindIII 的 6 個鹼基 及HA3His6 的 114 個鹼基,共計 1938 個鹼基,可轉譯成有 646 個胺基酸。 預估合成蛋白質大小約為71 kDa(646 × 0.11 Kda),經 SDS-PAGE 電泳 後,預期應出現在55~72 kDa 區域內出現 band。經進行 Coomassie blue stain 比對試驗後,仍未觀察到明顯的band 出現。繼續將上清液以 anti His 抗體 進行Western blot,結果如圖 18。將 Western bloting 的結果和 Coomassie blue staining 的結果做對照,可見到 10560-2B-Ta2HAS 上清液約在 72 kDa 附近 有明顯的 band 出現,但是啤酒酵母菌萃取上清液並未在相對的區域出現 band,表示這條 band 呈現之處有胺基酸 His 存在,所以可以和抗體結合而 被顯影,而啤酒酵母菌上並沒有這個表現His 的序列。此外,Band 出現的 位置和原先推估約71 kDa 的位置大致相符,應可確認這個區域出現的 His 序列是帶有HA3His6 的 Ta2HAS 質體所表現。至此應可確認 orf19.2730 可 以啟動胺基酸的合成,是一個能產生蛋白質的ORF。
以 FPUR3 及 RPUR3 為引子,用 PCR 方式擷取 pRS426 質體上的 URA3 及其上游自第5662 到第 1220 個鹼基(487 個鹼基)的片段共 1285 個鹼基 的序列。其中URA3 上游 487 個鹼基的片段是為了保留其中的限制酶切位 備用,並未帶有任何 ORF。另於設計引子時在 5’端都加入 BamHI 的鑑識 序列,形成兩端都有 BamHI 切位,大小為 1297 個鹼基的片段,暫名為
proURA3(圖 19)。proURA3 片段經 PCR 放大並 clean up 後,先以 BamHI 同 時切割5’和 3’端。另外也對 YEP363 和 Ta2Y 質體以 BamHI 進行切割。在 切割後隨即去除其5’端磷酸根(Shrimp Alkaline Phosphatase, SAP) 避免切 割後的YEP363 或 Ta2Y 在接合時發生自體連接。再與先前經 BamHI 切割 後的proURA3 片段進行接合,建構 YEP363URA3 和 YEP363TA1URA3 質 體。依圖 20 的分析,YEP363URA3 質體應有 9879 個鹼基,經 BamHI 切 割後應可得到8588 及 1291 個鹼基的兩條片段。而 YEP363TA1URA3 質體 應有12055 個鹼基,經 BamHI 切割後應可得到大小為 10764 及 1291 個鹼 基的兩條片段。 將建構的質體轉殖到 E. coli 勝任細胞上,用加入 Ampicillin 的 LB 培 養盤於37℃培養 18 小時。挑出數個菌落在 LB broth 培養 18 小時後抽取質 體。經膠體電泳後,分別挑出比YEP363(8588 個鹼基)及 Ta2Y(10764 個鹼基)大的質體,再以限制酶 BamHI 來確認大小。電泳結果如圖 21,
圖中Ta1 是經 BamHI 切割後形成線狀結構,大小為 10764 個鹼基的 Ta2Y 質體,可作為參考座標。Tu 為經 BamHI 切割的 YEP363TA1URA3 質體, 分別在 10764 附近及 1000 到 1500 個鹼基之間出現兩條片段。Yu 是 YEP363URA3 質體經 BamHI 切割,也分別在 8000 到 10000 及及 1000 到 1500 個鹼基之間出現兩條片段。這些片段都出現在符合預期的範圍,應可 確認質體中都已成功的加入了 proURA3,完成質體 YEP363URA3 和 YEP363TA1URA3 的建構。