從原核表現人類酪氨酸亞硫酸基轉移酶探討其催化活性與功能
61
0
0
全文
(2) 從原核表現人類酪氨酸亞硫酸基轉移酶 探討其催化活性與功能 Prokaryotic Expression, Purification and Characterization of Human Tyrosylprotein Sulfotransferase 2 (TPST2) 研 究 生:陳柏翰. Student: Bor-Han Chen. 指導教授:楊裕雄 教授. Advisor: Prof. Yuh-Shyong Yang. 國立交通大學 生物科技學系 碩 士 論 文 A Thesis Submitted to Department of Biological Science and Technology National Chiao Tung University in Fulfillment of the Requirements for the Degree of Master of Science in Biologic Science and Technology July 2007 Hsinchu, Taiwan, Republic of China 中華民國九十七年七月.
(3) 從原核表現人類酪氨酸亞硫酸基轉移酶 探討其催化活性與功能 學生: 陳柏翰. 指導教授: 楊裕雄 教授 國立交通大學生物科技學系碩士班. 酪氨酸亞硫酸基轉移酶負責催化許多分泌性蛋白質和膜上蛋白 質之亞硫酸基化。酪氨酸亞硫酸基轉移酶已經在許多哺乳動物和低 階的生物,如線蟲和果蠅中被發現,但在酵母菌與原核生物中卻沒 有發現到。在大部分的研究中,酪氨酸亞硫酸基轉移酶的來源通常 是由天然物與細胞培養方式來製備,為了能夠更加了解酪氨酸亞硫 酸基轉移酶的催化特性,我們成功地開發出新的表現系統,從大腸 桿菌中純化出人類酪氨酸亞硫酸基轉移酶-2,利用 Mass 指紋圖譜分 析所得到結果確實為人類酪氨酸亞硫酸基轉移酶-2。藉由同位素標 定的方式,探討重組的酪氨酸亞硫酸基轉移酶一些基本性質;包括 催化是否隨著時間呈線性增加,以及不同的溫度、pH 值與金屬離子 的濃度對酵素活性之影響,我們也針對此酵素對於 polyEAY 做酵素 動力學試驗。未來我們將使用這一套系統去表現突變的酪氨酸亞硫 酸基轉移酶,並尋找負責催化活性的胺基酸與一些尚未解決反應機 制的問題。 i.
(4) Prokaryotic Expression, Purification and Characterization of Human Tyrosylprotein Sulfotransferase 2 (TPST2) Student: Bor-Han Chen Advisor: Prof. Yuh-Shyong Yang Department of Biological Science and Tenology and Institute of Biochemical Engineering, National Chiao Tung University, Hsinchu, Taiwan, ROC. ABSTRACT Tyrosylprotein sulfotransferase (TPST) is responsible for the sulfation of a variety of secretory and membrane proteins. TPSTs have been found in mammalian and metazoans such as Caenorhabditis elegans and Drosophila melanogaster, but not in yeast and prokaryotes. TPSTs have been studied only from natural materials and eukaryotic cells following partial purification. To be able to fully characterize the biochemical properties of TPSTs in depth, we successfully overexpressed and purified human TPST2 in E. coli, and then confirmed by Mass fingerprinting identification. The properties of recombinant TPST2 were characterized using by isotope-based. analysis.. The. optimal. temperature,. pH. profile,. and. concentration-dependent of MnCl2 for human TPST2 were determined. The kinetic constants of TPST2 using polyEAY as substrate were also determined. Furthermore, the important amino acids responsible for the activity of TPST and its mechanism of actions will be studied.. ii.
(5) Acknowledgement 首先誠摯的感謝指導教授楊裕雄博士,悉心的教導使我得以一窺鍊金術領 域的深奧,不時的討論並指點我正確的方向,使我在這些年中獲益匪淺。老師 對學問的嚴謹更是我輩學習的典範。 本論文的完成另外亦得感謝實驗室大力協助,高中好友與父母的支持、張 程翔學長在 isotope 實驗上的幫忙,以及呂平江教授、汪宏達教授與林照雄教授 對我研究上的指教評點。因為有你們的幫忙與教導下,使得本論文能夠更完整 而嚴謹。 兩年裡的日子,實驗室裡共同的生活點滴,學術上的討論、言不及義的閒 扯、讓人又愛又怕的宵夜、趕作業的革命情感、因為實驗太晚而睡倒在實驗室(囧 學長亂取“燈塔”稱號),感謝大晃學長、Rich 學長、小米學姐、漢平學長、陸宜 學長與青辰學長的耐心指導下,不厭其煩的指出我研究中的缺失,且總能在我 迷惘時為我解惑。也感謝 LEPE 所有實驗室同仁,還有很愛虧我的家煒= =(讓 我箭箭重傷 XD 囧) ,還有唱歌好聽的咏馨與小 apple 當然也不能忘記,你們 的幫忙及搞笑我銘感在心。 有你們的陪伴讓兩年的研究生活變得絢麗多彩。 而好友………….(很多名)的陪伴更是令人感動,在此一併致謝。. iii.
(6) CONTENTS Chinese Abstract English Abstract Acknowledgement Contents Table Contents Figure Contents Abbreviations. …………………………………………………………… …………………………………………………………… …………………………………………………………… …………………………………………………………… …………………………………………………………… ……………………………………………………………. i ii iii iv v vi vii. 1 1.1. Literatures on tyrosine O-sulfation Some common and important post-translational modifications.. 1 1. 1.2. Sulfotransferases. 2. 1.3 1.3.1 1.3.2. Tyrosylprotein sulfotranferases Tyrosine O-sulfation of chemokine receptors Tyrosine O-sulfation of hemostasis Tyrosine O-sulfation mediate leukocyte inflammatory response. 3 4 5. 1.3.3. adhesion. and. 6. 1.4. Application of antibody to tyrosine O-sulfated proteins researches. 6. 1.5. Tyrostlprotein O-sulfation prediction by bioinformatic tools. 7. 2.1. Introduction. 8. 2.2. Materials. 13. 2.3. Experimental procedures. 14. 2.4. Results. 19. 2.5. Discussions. 23. Reference Appendix. 44 50. iv.
(7) Table contants. PAGE. Table 1.. Some common and important post-translational modifications.. 28. Table 2.. Specific effects of tyrosine O-sulfation. 29. Table 3.. Conservation of tyrosine sulfation sites in human chemokine receptors. 29. Table 4.. TPSTs purified from many source. 30. v.
(8) Figure Contents. PAGE. Figure 1.. Schematic representation of protein modifications related to the regulation of biological processes.. 31. Figure 2.. Sulfate activation and tyrosine O-sulfation.. 31. Figure 3.. Schematic representation of cell entry by HIV-1 following sulfonation of CCR5 by a tyrosylprotein sulfotransferase.. 32. Figure 4.. GPIbα from amino acid 200-294. 32. Figure 5.. Tyrosine sulfation plays an important role in the immune response.. 33. Figure 6.. Graphical presentation of the far-Western immunoblot technique.. 34. Figure 7.. The catalyzed reaction of tyrosylprotein sulfotransferase.. 34. Figure 8.. Sequence alignment and transmembrane domain analysis of tyrosyl protein sulfotransferase.. 35. Figure 9.. Prokaryotic expression and purification of human hTPST2.. 36. Figure 10.. The protein of LC-MS-MS fingerprinting analysis was identified for human TPST2.. 37. Figure 11.. Reaction of polyEAY sulfation as a function of time. 38. Figure 12.. Effect of Temperature on hTPST2 activity. 39. Figure 13.. Different pH affects the catalytic activity of hTPST2.. 40. Figure 14.. Effect of MnCl2 on hTPST2 activity. 41. Figure 15.. Kinetics of hTPST2 for polyEAY as substrate.. 42. Figure 16.. The SDS-PAGE of PSGL-1 peptide purification.. 43. vi.
(9) Abbreviations Abbreviation and Symbol. Full name. ε. Absorption (exitinction) coefficient. A280. Absorption at 280 nm. A600. Absorption at 600 nm. kcat. Turnover number. kDa. Kilodaton. Km. Michaelis constant. MALDI-TOF. Matrix-assisted laser desorption inonzation time of flight. MES. 2-[N-morpholino] ethanesulfonic acid. PAGE. Polyacrylamide gel electrophoresis. PAP. Adenosine 3',5'-diphosphate. PAPS. 3'-phosphoadenosine 5'-phosphosulfate. polyEAY. polypeptiede (Glu: Ala : Tyr = 6 :3 :1). SDS. Sodium dodecyl sulfate. SULT. Sulfotransferase. Vmax. Maximum velocity. vii.
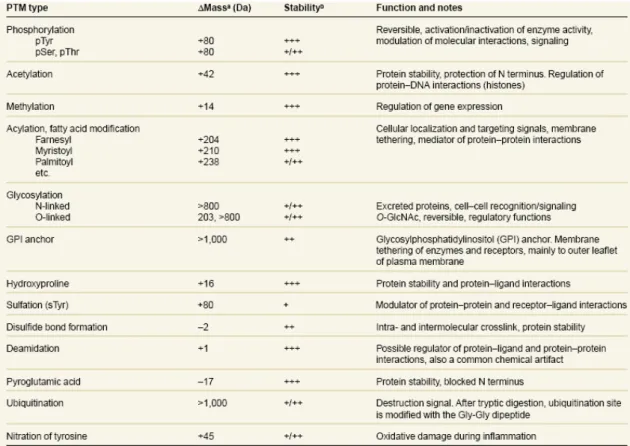
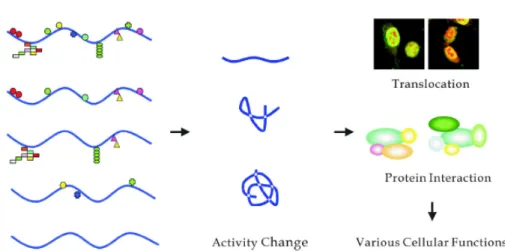
(10) 1.. Literatures on tyrosine O-sulfation. 1.1 Post-translational modification in the regulation of protein functions. The post-translational modifications of proteins can regulate the protein functions and protein activities by causing the changes of the protein structure or the affinity of dynamic interaction between proteins and compounds. (Fig. 1) (Seo et al. 2004) A series of are involved in the signaling pathway from membrane to nucleus in response to external stimuli. (Zhao Y et al. 2008; Guo et al. 2007) Some common and important post-translational modifications include sulfation,. phosphorylation,. acetylation,. methylation,. glycosylation,. and. ubiquitination. “For the discovery of ubiquitin-mediated protein degradation” was awarded 2004 Nobel Prize in chemistry. Protein sulfation has drawn a significant amount of attention recently. For instance, CXCR4 and CCR5 include modifications of the extracellular domains (N terminal, ECI, ECII, and ECIII) or the intracellular loops. Modifications of the extracellular domains include N-linked and O-linked glycosylation and tyrosine sulfation. The intracellular loops of the receptors may undergo palmitoylation, phosphorylation, and ubiquitination. These three types of modifications play major roles in receptor turnover and will be discussed in relation to receptor endocytosis and recycling. In previous literatures, post-translational modification was identified by. 1.
(11) radioisotope labeling, western analysis with antibody against specific modification, mutagenesis of modification sites and mass spectrometry. Mass spectrometry is a powerful tool to map modification sites and species. MS measures mass-to-charge ratio (m/z), yielding the molecular weight and the fragmentation pattern of peptides derived from proteins. (Mann et al. 2003). 1.2. Sulfotransferases Sulfate-containing biomolecules were identified in 1876.( Baumann et al. 1876). Sulfotransferases(STs) catalyze the transfer of sulfonated group (SO2-) from a donor, PAPS, to an acceptor which has alcohol, thiol, amine, or phenol as functional groups. There are two classes of STs: cytosolic sulfotransferases and membrane-associated sulfotransferases. Cytosolic sulfotransferases mediated small chemical compounds (Steroids, xenobiotics, dietary carcinogens, and neurotransmitters) to regulate hormone homeostasis, and metabolic detoxications/activation of xenobiotics. Membrane-associated sulfotransferases catalyze carbohydrates and proteins (Heparan, glycoproteins, oligopeptide, and proteins). Many of which have been implicated recently in crucial biological processes, for instances, modulating receptor binding, signaling processes, and intercellular communications.. 2.
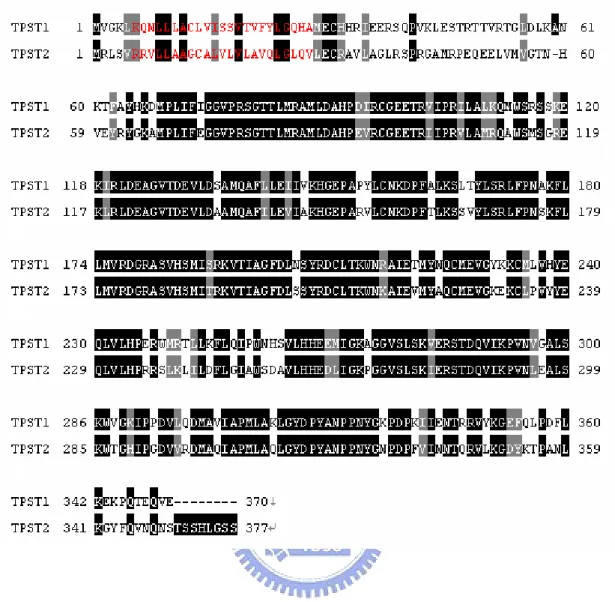
(12) 1.3. Tyrosylprotein sulfotranferases Tyrosylprotein sulfotransferases (TPSTs) are type II transmembrane proteins. with a short N-terminal cytoplasmic domain, a single about 17-residue transmembrane domain (red), and a luminal catalytic domain.(Fig. 8)(Baeuerle et al. 1987; Lee et al. 1985) TPSTs are localized in the Golgi lumen. In most species, TPSTs are classified. as TPST-1 and TPST-2, but D. melanogaster and C. elegan have only a single TPST gene, TPST-2 and TPST-A, respectively. Furthermore, no tyrosine-sulfated proteins, TPST activity, or putative TPST orthologs have been described in prokaryotes or in yeast. (Moore et al. 2003) Human TPST-1 and human TPST-2 share 68% sequence identity. (Fig. 8) A recent gene-knockout researches revealed that mice deficient in TPST-1 had reduced body mass and increased postimplantation fetal death (Ouyang et al. 2002) and that mice deficient in TPST-2 had caused male infertility. (Borghei et al. 2006) Sulfation is similar to phosphorylation, but protein sulfation in signaling pathway was less studied as compared to phosphorylation. The Golgi localization and the luminal active site orientation of TPST-1 and -2 predict that tyrosine O-sulfation occurs only on proteins that transit the trans-Golgi network.(Fig. 2) PAPS is synthesized by the action of one of two PAPS synthases in the cytosol. TPSTs used PAPS, as the donor of sulfonate group, to catalyze the protein that transit trans-Golgi network. The sulfonate protein affects lots of protein-protein interactions, in hemostasis, leukocyte 3.
(13) adhesion, inflammatory response, chemokine receptor, and bioactive peptides. The following are the details about protein-protein interactions through protein sulfation.. 1.3.1 Tyrosine sulfation of chemokine receptors Chemokines are involved in leukocyte trafficking, angiogenesis, angiostasis, battling viral infections, and the host immune response to cancer (Zlotnik et al. 1999). In last decades, the sulfation of chemokine receptors is found to play important role to affect enzyme activity, protein lifespan and the binding affinity between proteins.(Table 2) To days, sulfated tyrosines have been experimentally confirmed in only five human chemokine receptors: CXCR3, CXCR4, CCR2b, CCR5, and CX3CR1. (Farzan et al. 1999; Farzan et al. 2002; Preobrazhensky et al. 2000; Fong et al. 2002; Colvin et al. 2006) Many tyrosine sulfation was often found in the same groups of protein and. peptides. For instance, CCR5 and CXCR4 have tyrosine sulfated. CCR5 carries four tyrosine residues in the amino-terminal region. (Farzan et al. 1999) Incubation of CCR5-expressing Cf2Th cells with sodium chlorate (NaClO3)a, a global inhibitor of the sulfation (Farzan et al. 1999), decreased binding of MIP-1 d MIP-1—two natural chemokine ligands for CCR5. Sulfonate tyrosine14 of CCR5 seems to play a particularly important role in binding to the chemokines MIP-1(Chapman et al 2004). a. Sodium chlorate is a specific inhibitor of the ATP sulfurylase activity of PAPS 4.
(14) synthetase, and a global inhibitor of the sulfation. (Farzan et al. 1999) In addition to binding native chemokines, CCR5 also serves as a co-receptor for HIV-1 gp120, composing together with CD4 to mediate attachment of the virus and its subsequent infections.(Fig. 3) (Kuhmann et al. 2004; , Chapman et al. 2004) Mutation of the four sulfotyrosine residues in CCR5 to phenylalanine inhibits HIV infection by 50–75% in cultured cells, depending on the HIV isolate tested (Farzan et al. 1999). This information suggests that inhibitors of tyrosine sulfation might serve as anti-HIV herapeutics, as long as disruption of the modification is not globally toxic. Alternatively, peptides that mimic the tyrosine-sulfated sequence of CCR5 might compete for binding to HIV-1 gp120 and block viral adhesion.. 1.3.2 Tyrosine sulfation of hemostasis Tyrosine sulfation is also involved in hemostasis. This modification is crucial in the interaction between many plasma proteins such as fibronectin and fibrin (Suiko et al. 1988), hirudin and thrombin (Stone et al. 1986), coagulation factor VIII, and von Willebrand factor (VWF)(Leyte et al. 1991) and glycoprotein (GP) Ibα with both VWF and thrombin(Marchese et al. 1995; Ward et al. 1996; Fredrickson et al. 1998; Dong. et al. 2001; Murata et al. 1991). The interaction between the platelet. membrane protein (GP) Ibα and VWF is essential for initiation of hemostasis. For. 5.
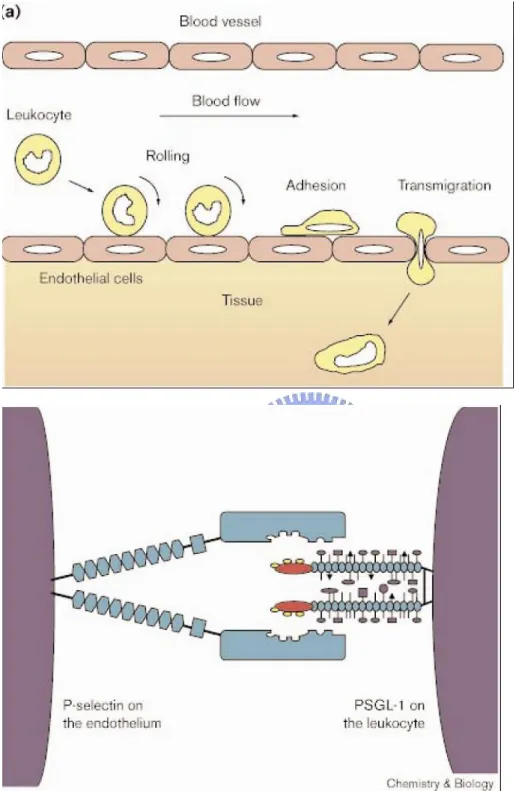
(15) instance, in previous studies, they found many sulfated tyrosine residues, located within this region at positions 276, 278, and 279, in the platelet membrane protein (GP) Ibα . (Fig. 4) (Marchese et al. 1995; Ward et al. 1996; Fredrickson et al. 1998) The region has been demonstrated that how sulfated tyrosine residues affect both the conformation of the amino-terminal region of GPIbα and the electrostatic interaction with VWF in the presence of both ristocetin and botrocetin.(Lee et al. 1983). 1.3.3 Tyrosine sulfation mediate leukocyte adhesions and inflammatory responses The inflammatory response results from producing and releasing of chemical agents by cells in the infected, injured or diseased tissue. The chemical agents induced the leukocyte into the site of the inflammatory. In the processes, leukocytes roll upon, adhere to and transmigrate between endothelial cells at sites of inflammation. Tyrosine sulfation near the amino terminus of PSGL-1 plays an essential role in P-selectin binding.(Fig. 5) (Pouyani et al. 1995; Sako et al. 1995; Wilkins 1995). 1.4. Application of antibody to researches of tyrosylproteins. Antibody is a useful tool to discovery lots of things, including staining tissue,. immunoprecipitation, immunoblotting, immunoaffinity purification, tagging proteins and the epitope mapping. (Kasinathan et al. 2005; William et al. 1997; Ramaprasad et al. 1998; Hoffhines et al. 2006; United States Patent 5716836). 6.
(16) Antibodies in researches of tyrosine O-sulfation were widely used. I am interested in the epitope mapping of tyrosine O-sulfation. In 1992, united states patent, patent number:5716836, Masahito suiko was successfully produced the anti-sulfated tyrosine antibody specific for sulfated tyrosine but not to unsulfated tyrosine.(United States Patent 5716836), the antibody of anti-sulfated tyrosine cound be not purchased but polyclonal anti-TPST1 anibody, abnova (H00008460-A01) and polyclonal anti-TPST2 antibody, abcam (ab59958), could. If we could buy the anti-sulfated tyrosine antibody, we would use proteomics tools to discovery lots of the sulfations of proteins, and how to affect biological functions in cells. In previous studies, most of these used antibody to purification enzymes, location in cells, and detection the quantity of protein expressions.(Ramaprasad et al. 1998; Kasinathan et al. 2005). More interacted proteins could be found out and then to deduce the functions and pathways by using the antibody to help to facilitating the technique of protein-protein interaction such as far western blotting (Wu et al. 2007) (Fig. 6), but previous researches don’t study in these aspects.. 1.5. Predication the sulfation of protein by bioinformation tools The post-translational modifications (PTMs) modulate all aspects of cellular. life. Many PTMs have been discovered including protein kinases, sulfation, acetylation, methylation, amidation….etc.(Table1) PTMs affect lots of functions 7.
(17) thought protein modification in physiology. How to predict PTMs sites from a protein. sequence. is. very. important.. Previous. researches. used. Position-Specific-Scoring-Matrix (PSSM) to compare the distributions of amino acids around the query tyrosine sites with those of known sulfated and non-sulfated tyrosine sites in Swiss-Prot.(Liu et al. 2008) Each query tyrosine site was assigned a score measuring the similarity of the amino acids around the tyrosine to those around either the known sulfated tyrosine sites or those around non-sulfated tyrosine sites. The site of tyrosine o-sulfation took high score. (Nicholas et al. 1999; Lin et al. 2003) The softwares, Sulfonator(http://ca.expasy.org/tools/sulfinator/) and SulfoSite (http://sulfosite.mbc.nctu.edu.tw/) , are often used to predict sulfation sites. The figure 1 as the example, Justin Liu et al. screened all human chemokine receptors and found that many tyrosines predicted to be sulfated were in a region localized to the N-terminus of their receptor.(Table 3.)(Farzan et al. 1999) The following tyrosines are known to be sulfated: Y12 and Y21 of CXCR4; Y26 of CCR2; Y3, Y10, Y14, and Y15 of CCR5; and Y14 of CX3CR1. CCR8 is known to be tyrosine sulfated in the mouse, probably in both Y14 and Y15. These tyrosine sulfation sites were frequently completely conserved in predictions. Previous researches also discoveried some conditions to increase the chances of the tyrosine sulfation. There is the presence of negatively charged acidic amino acid. 8.
(18) residues around the site of tyrosine o-sulfation.(Baeuerle et al.1986;Liu et al. 2008) The secondary and tertiary protein structure promotes to expose adjacent tyrosines to TPSTs. (Huttner et al. 1984) The bioinformatics tool for prediction of specific post-translational modifications will be useful in discovering these important protein modifications.. 9.
(19) Prokaryotic Expression, Purification and Characterization of Human Tyrosylprotein Sulfotransferase 2 (TPST2). 2.1. Introduction Tyrosine O-sulfation is a common post-translational modification of protein in. lots of mammalian cells. Lee and Huttner directly demonstrated tyrosine O-sulfation is catalyzed by tyrosylprotein sulfotransferases (TPSTs, EC 2.8.2.20) (Wilkins et al. 1995). So far only two distinct human TPSTs were indentified, TPST1 and TPST2.. They are in similar size (370–377 residues) and share 67% identity in primary sequence. TPSTs are type II transmembrane topology with a short N-terminal cytoplasmic domain and a luminal catalytic domain. It transfers the sulfonate group (SO3-1) from 3’-phosphoadenosine-5’-phosphosulfate (PAPS), the only known sulfonate donor (Robbins P, 1956), onto a tyrosyl residue within target proteins as shown in Fig. 7. The sulfation plays an important role in many biological mechanisms described as followed: 1) Modulation of receptor binding (Preobrazhensky et al., 2000; Farzan et al., 2002; Wilkins et al., 1995); 2) Intercellular communication (Rossi et al., 2000); 3) Signaling processes (Goettsch et al., 2002). To this day, tyrosine O-sulfation affects lots of critical pathology. In the previous research, the experiment of TPST double knockouts mice had discoveried. 10.
(20) that TPST-1 and TPST-2 both affected the functions of thyroid.(Westmuckett et al. 2008) The absence of TPST1 and TPST2 caused euthyroid and primary hypothyroidism, respectively. Furthermore, the mouse lungs of TPST double knockouts failed to expand at birth so that the mouse died in asphyxia during the early postnatal period (Westmuckett et al. 2008). Tyrosine O-sulfation also come into notice about HIV infections and inflammations in recent years. Sulfonated CCR5 renders gp120 of HIV-1 interacts with the sulfonated CCR5/CD4 complex, and then leads to the entry of the virus into the host cell. (Choe et al. 2003) The tyrosine O-sulfation near the amino terminus of PSGL-1 also plays an essential role in P-selectin binding.(Kehoe et al, 2000) Sulfation enhances the affinity binding interaction so that sulfonated PSGL-1 helps rolling leukocyte adheres to endothelial cells at sites of inflammation. According to the analyses of known peptides, some acidic and basic amino acid around the tyrosine from +5 residues to -5 residues affect the catalytic activity of tyrosine O-sulfation in its substrates. (Mishiro et al. 2006; Bundgaard et al. 1997; Lin et al. 1992) Substrate specificity and mechanism have not been well-known in the past years, so that developing the powerful and quick method of the purification to obtain adequate TPST is necessary. Furthermore, the crystal structure and antibody preparation are also urged to facilitate the study in advanced.. 11.
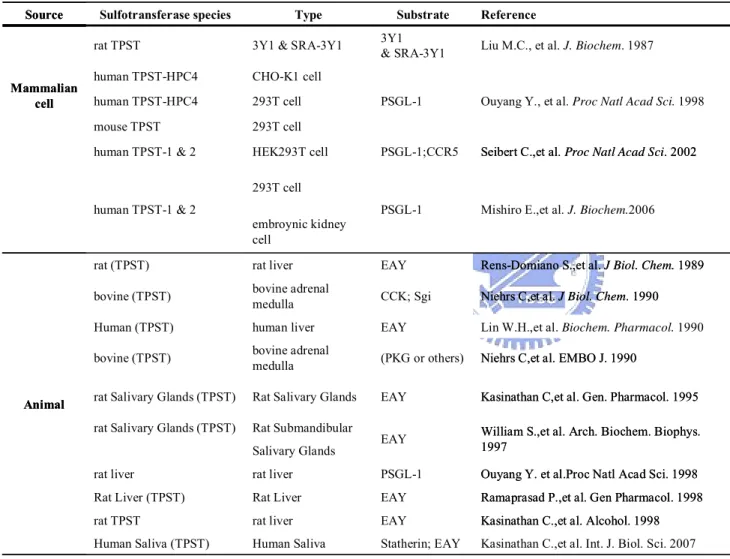
(21) Previous studies investigated the characterization of TPSTs, whose enzyme source were purified from crude and partially purified preparations from a variety of mammalian cells neither in yeast nor prokaryote (Table 4). Expresses desired enzyme from prokaryotic system will be convenient, stable, time-and-effort-saving, and acquire lots of the enzyme quantities. Firstly we set up the prokaryotic expression system to express human TPST2, and further study the characterizations of recombinant TPST2. In the future, we will facilitate this platform to discover the important amino acid regulating PAPS binding, substrate specifity, catalysis, and enzymatic mechanism.. 12.
(22) 2.2. Materials The human TPST2 clone (accession No. is AAH17509) was obtained from. GenDiscovery Biotechnology, Inc. PfuTurbo DNA polymerase was purchased from Stratagene, and T4 DNA ligase, BanHI, XhoI restriction endonucleases were purchased from New England Biolabs. Oligonucleotide primers were synthesized by Mission Biotech Co., Ltd. (Taiwan). MES, NaOH, Trizma base, NaCl, Triton X-100 and imidazole were products of Sigma (St. Louis, MO, USA). HisTrap fastflow sepharose, HiTrap Q fast flow, and glutathione S-transferase sepharose fast flow were obtained from GE Healthcare. Cellulose thin-layer chromatography plates were obtained from MERCK, Ltd. All other reagents were of the highest grades commercially available.. 13.
(23) 2.3. Experimental procedures. Protein sequence alignment and prediction of transmembrane domain of TPST1 and TPST2—The sequence pairwise alignment was performed by ClustalW (http://www.ebi.ac.uk/Tools/clustalw2/index.html). and. sorted. shading. by. BOXSHADE server (http://www.ch.embnet.org/software/BOX_form.html). The black background indicated identity to each other and the gray one meant conserved substitutions. The residue colored in red is the predicted transmembrane domain calculated by PSIPRED (http://bioinf.cs.ucl.ac.uk/psipred/psiform.html) ranged from residue 6 to 28 both for human TPST1 and TPST2.. Cloning of cytosolic domain of TPST2—The human TPST2 incorporated into the vector pOTB7 originally was subcloned into pET-43a (Appendix 2). The potential cytosolic domains of TPST was amplified by PCR through specific primers designed to contain BamHI restriction site in the forward direction was 5’-tgaaggatccctagagtgccgggcggtgctggc-3’, and the reverse one contained XhoI restriction site was 5’-gccactcgagtcacgagcttcctaagtgggagg-3’. cDNA fragments were inserted into the BamHI/XhoI doubly-restriction sites and then confirmed using an ABI Prism 377 DNA sequencer (Applied Biosystems, Foster City, CA) following the standard protocol.. 14.
(24) Cloning of substrate peptides—The specific primers were designed to produce the self-annealed. cDNA. (ATEYEYLDYDFL),. encoding CCR5. the. N-terminal. region. (DYQVSSPIYDINFYTSE),. of and. PSGL-1 Ror2. (MCRNKQKASASTPQRRQLMASPSQDMEMPLINQHKQAKLKEISLSAVRFM E), respectively. The self-annealed oligonucleotides were subcloned into BamHI–XhoI restriction site of pGEX-4T1 and transformed into BL21 (DE3) to express further.. Protein expression and purification of human TPST2 and its substrates—A single colony of BL21 (DE3) consisted of TPST2 or substrates (PSGL-1, and CCR5) plasmid was used to inoculate in the LB broth with ampicillin as the antibiotic at 37 °C. Growth was continued to an ODA600 of 0.4–0.6 and then induced with 0.4 mM isopropyl-thio-β-D-galactoside (IPTG) for 16 hours incubation at 20 °C. The cultures were centrifuged at 14000¯g for 20 minutes, and the pellet was sonicated in IMAC5 buffer (20 mM Tris-HCl, pH 8.0, 500 mM NaCl, 5 mM imidazole, and 10% glycerol) for TPST2 and GST lysis buffer (20 mM Tris–HCl, pH 8.0, 150 mM NaCl, and 10% glycerol) for substrates. Further the HisTrap sepharose charged with NiSO4 was facilitated to the TPST2 purification. GSTrap for Substrates were purified by GSTrap sepharose following the digestion against the bovine thrombin at 4 °C for 16 hours. 15.
(25) and then applied to HiTrap Q chromatography. A homogeneous protein (Figs. 9 and 16) was obtained as determined by SDS-polyacrylamide gel electrophoresis (38).. Protein identification by LC-MS-MS spectrometry—For MS analysis, protein spots were excised from the gel and digested with trypsin according to published procedures (Shevchenko et al. 1996). The protein spots were sent to Academia Sinica, Core Facilities for Proteomics and Structural Biology Research, for Mass Spectrometric analysis. Proteins were identified by searching the protein databases NCBInr using MASCOT (http://www.matrixscience.com ). To denote a protein as unambiguously identified, the Mowse scoring algorithms were sued. (Fig. 10). TPST enzymatic activity assay—For the determination of TPST activity from bacterial expression, we detected radiation of. 35. S using [35S]PAPS as donor and. transferred sulfate group to substrate, such as PSGL-1 peptides, polyEAY, or CCR5 peptides. The complete assay mixture consisted of sulfate acceptor (TPST substrate), 40 μM polyEAY peptides, 50 mM MES at pH 6.0, 25 mM MnCl2, 50mM NaF,0.5 μM [35S]PAPS (15Ci/mmole), and 4 μg human TPST2 in a final volume of 15 μl. Catalytic reactions were initiated by the addition of the enzyme and incubated for 30 minutes at 30 °C. The reactions were terminated by heating at 95 °C for 2 minutes.. 16.
(26) The supernatant was collected and analyzed by spotting 1 μl aliquot of the reaction mixture on a cellulose thin-layer chromatography (TLC) plate and developed with n-butanol/pyridine/88%formic acid/water (4:2:2:4; by volume) as the solvent system. The dried plate was exposed with Kodak BioMax MR films which provided the optimal resolution for 35S autoradiography.. Time-dependent TPST enzymatic activity assay—This activity of TPST2 was measured as described above. Each assay consisted of 50 mM MES at pH6.0, 25 mM MnCl2 , 50 mM NaF, 0.5 μM [35S]PAPS (15Ci/mmole), 40 μM polyEAY, 0.5% Triton-X100 and 4 μg human TPST2 in a final volume of 10 μl. Data shown here is the result of a typical experiment performed in triplicate.. pH profile of TPST enzymatic activity—Each assay contained 50 mM MES at desired pH (5.5, 6.0, 6.5, 7), 25 mM MnCl2 , 50 mM NaF, 0.5 μM [35S]PAPS (15Ci/mmole), 40 μM polyEAY, and 4 μg human TPST2 in a final volume of 10 μl. Data shown here are the result of a typical experiment performed in triplicate. This experiment was repeated three times with similar results.. Temperaturen effect on TPST activity—This experiment was performed described. 17.
(27) above under various temperatures (25, 30, 35, 37, 40, 45, 50 °C). Data shown here is the result of a typical experiment performed in triplicate. and repeated three times with similar results.. MnCl2 dependence of TPST enzyme assay—Various concentrations of MnCl2 were examined in the enzyme assay. Data shown here is the result of a typical experiment performed in triplicate.. Kinetic constants determinations of TPST—Measurements of kinetic constants were performed by varying the polyEAY concentration while keeping the PAP at a fixed and near saturating concentrations. The apparent Km and Vmax were determined using nonlinear regression by SigmaPlot 2001. V7.0 and Enzyme module, V1.1.. 18.
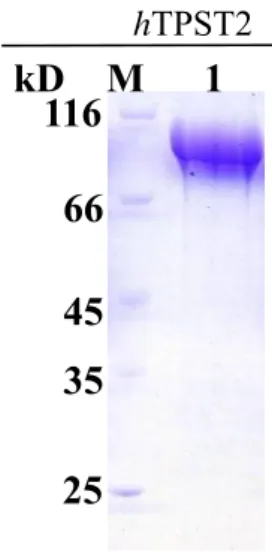
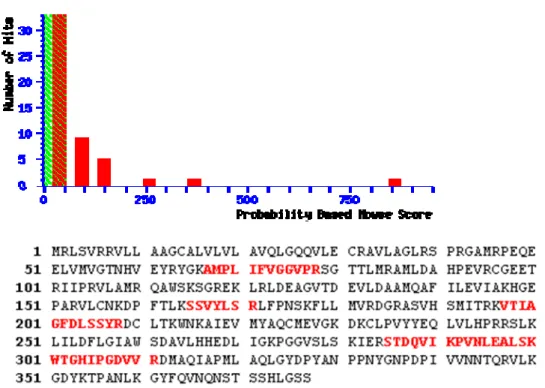
(28) 2.4. Result. Expression of recombinant human TPST2 in prokaryote expression system. The human TPST2 is localized in the membrane of Golgi apparatus network and the transmembrane domain is shown as Fig 8. In our experiment, the catalytic domain of TPST2 from residue 29 to 377 was incorporated into various expression vectors. Various expression vectors incorporated human TPST2 cDNA in the open reading frame were examined (data not shown). Most of the expression vector could not prevent TPST2 from inclusion body. pET-43a (Appendix 2), a vector of expressing proteins was purchased from Novagen, was competent to express recombinant human TPST2 in E. coli with the reducing amount of inclusion body. The prokaryotic expression of human TPST2 was optimized to reach the maximal soluble amount and purified to nearly homogeneity (Fig. 9). A band on the SDS-PAGE of 100 kDa protein composed of NUS-Tag fusion protein (60 kDa) and TPST protein (40 kDa) upon treatment in coomassie blue R350. The spots excised from SDS-PAGE were analyzed by LC-MS/MS. (Fig. 10) There were many peptides (colored in red) come after trypsin digestion and they indicated the peptide sequences of human TPST2. The alignment of these peptide sequences showing homology to human TPST2 with high scores of confidences.. 19.
(29) Time-dependence of human TPST2-catalyzed tyrosine O-sulfation polyEAY is a substrate of TPST2 with higher specificity activity than other endogenous substrates as described in the previous reports. Firstly the time dependence of the activity of the human TPST2 with polyEAY as substrate was examined. The concentrations of PAPS and polyEAY, were both saturated in the reactions. The tyrosine O-sulfation of polyEAY increased linearly with the incubation time as shown in the Fig. 11. The slope in this reaction was not appreciably reduced up to 120 mins.. Temperature effect on TPST activity So far the catalytic activity of human TPST2 in different temperatures was not studied clear in the past. The experiment demonstrated that human TPST2 exhibited the activity at 25 °C was three times than that at 40 °C (Fig. 12). This catalytic reation was almost quenched under the treatment of 45 °C for 30 mins. The human TPST2 might denature and thus lost the fuction of tyrosine O-sulfation catalysis. The activity at 37 °C (body temperature) was also investigated and it was apparently lower than the activity at 25 °C. It might be attributed to the reason as decribed above.. 20.
(30) pH profile of TPST pH affects the electricity of amino acid and further contributes to the substrate binding affinity, enzymatic catalysis, and protein conformational structure. The pH profiles of the recombinant human TPST2 were determined by measuring the activity at various pH values. The pH optimum was ranged from 5.5 to 6.0 within the error tolerance as showed in Fig. 13. The catalytic activity of human TPST2 appreciably decreased from pH 6.0 to 6.5 and was nearly nondetectable at pH 7. pH values higher than 6.0 might result in the instability of tyrosine O-sulfated peptide, or the dysfunction of human TPST2 in catalysis.. Metal ion effect on TPST2 It is known to require exogenous metal ions for activity with Mn2+ and Mg2+ to activate the highest activity of TPSTs (Mishiro et al. 2006). The data indicated that whether there is metal ion in enzyme catalysis or not, it is not necessary for human TPST2 (Fig. 14). The concentration of Mn2+ at 25 mM performed the maximal activity and 2.5 folds higher than the absence of Mn2+.. 21.
(31) Kinetics of TPST2 utilized polyEAY as substrate Previous studies indicated that tyrosine sulfation was studied in the subcellular fractions containing the enzyme activity.(William et al. 1997; Sane et al. 1993; Lin et al. 1990) The most widely used sulfonate acceptor was EAY as a positive control. The kinetic constants toward polyEAY, the synthetic polypeptides0 composed of Glu, Ala, and Tyr in the ratio 6:3:1, demonstrated that Km was 10.6 μM and Vmax was 4.8 pmole/min/mg (Fig. 15). It revealed that the heterologous expression of human TPST2 was active in the catalysis and performed the similar kinetic constants compared to the previous studies. (Sane et al. 1993). 22.
(32) 2.5 Discussion Tyrosine O-sulfation is firstly discovered in 1954. (Bettelheim, F.R.1954) Tyrosylprotein sulfotransferase (TPST) is demonstrated to catalyze tyrosine O-sulfation by Lee and Huttner in 1983. In opposition to the researches of kinases, the ones of TPST are extremely few. It may be attributed to some characteristics of TPST, such as the difficulty to purify the homogenous and ample amount of TPST. TPST is reported to be labile and is hard to purify during the process of purification (Ouyang et al. 1998). In the previous studies, the enzyme source came from the nature materials or mammalian cell lines, and further purified through affinity column whose beads conjugated with its substrate or antibody.(Ramaprasad et al. 1998; Kasinathan et al. 2005) In this research, the heterologous expression system utilizing E. coli as host to purify the human TPST2 was optimized with high recovery. In the process of the purifications, the inclusion body and contamination of chaperonin 60 (GroEL) resulted in the difficulties to overcome. The formation of inclusion body includes solubility limitation, protein size, type of promoter, and improper disulfide formation. (Hartley et al. 1988; Marston et al. 1986) The choice of vector and expression host can significantly increase the activity and amount of target protein present in the soluble fraction. In the previous study, the truncated form of human TPST2 comprising the catalytic domain was. 23.
(33) secreted from stably transfected Chinese hamster ovary (CHO) cells (Ouyang et al.1998). According to the topological analysis of primary sequence of TPST2, the N-terminal transmembrane domain in TPST2 was truncated to prevent hydrophobic domain from interfering in this study. Moreover, the fusion protein, Nus•Tag, on the expression vector was utilized to enhance the solubility of target proteins (Davis et al. 1999; Harrison et al. 2000). Furthermore, the contamination of chaperonin 60 (GroEL) was found to co-elute with TPST2 in this study. Common features of chaperone action are transient interaction with non-native species in the prevention of aggregation and promotion of correct folding and assembly (Young et al. 2004; Bukau et al. 2006; Anken et al. 2005). The existence of the GroEL represented that TPST was not easy to fold or not fold into the correct stage. This interaction between TPST2 and GroEL was interfered through Triton X-100 to be the competence of the hydrophobic force and separate each other. This purification procedure in the study is simple, straightforward, and can produce great quantities and homogeneous sources of TPST2 (Fig. 9). The homogeneous human TPST2 was measured further to understand the characterizations and mechanism of action. The catalytic activity of zebrafish TPST with N-terminal PSGL-1 peptide as substrate indicated the optimal activity ranged from 28 to 37 °C (Mishiro et al. 2004). The temperature profile of human TPST2 with. 24.
(34) polyEAY as substrate in this study was different from the catalytic activity of zebrafish TPST (Fig. 12). The activity of TPST2 decreased with the increasing temperature and approached to inactive while the temperature was higher than 45 °C. Furthermore, previous research revealed that the TPST under the treatment of detergent possessed the half life of 48 hours at 4 °C (Niehrs et al. 1990). The catalytic specificity of human TPST2 is unknow in this studies. Previous studies had revealed that TPST1 and TPST2 are localized in the Golgi apparatus and the catalytic domain is situated in the lumen, which is an acidic environment (Baeuerle et al 1987; Lee et al.1985). The recombinant human TPST2 indicated that the activity is adaptable under acidic environment and become labile while the pH value was higher than 6.0 (Fig. 13). It might be resulted from the influence of the affinity of PAPS or substrate binding sites. The information from previous studies also indicated that the optimal pH of human TPST2 expressed from E. coli was the same as that from 293T cells, but was different from human saliva.(Mishiro et al. 2006; Kasinathan et al. 2005) It might infer that human TPST2 possessed the isoforms so that resulting in the different optimal pH towards the catalytic activity. Membrane lysates of Golgi apparatus have revealed the stimulatory effects of Mn2+ on the activity of TPST.(Mishiro et al. 2006; Kasinathan et al. 2005) The. 25.
(35) catalytic activity of human TPST from salivary and PC12 cells is stimulated by the divalent cations, such as Mn2+, Ca2+ and Mg2+, and is inhibited by EDTA.(Kasinathan et al. 2005) On the contrary, tyrosine O-sulfation of the endogenous membrane. proteins in A431 cell is not inhibited by EDTA.(Liu et al. 1986) The stimulatory effect and mechanism of metal inons, however, was still not clear so far. When MnCl2 was up to 40 mM in the reaction mixture, the enzyme activity was apparently decreased (Fig. 14). High concentration of Mn2+ might affect the structure and render the protein denaturation. The appropriate concentration of Mn2+ might also stabilize the sulfonate groups while catalysis and further to reduce the activation energy. It could be comparable to the role of Mg2+ in the kinase catalysis. According to the previous studies, the Km of platelet TPST for polyEAY as substrate was 3.7 μM and the Vmax was 0.09 pmol/min (Sane et al. 1993). In our study, the kinetic constants indicated that the Km and Vmax were 10.5 μM and 4.8 pmole/min/mg, respectively (Fig. 15). This difference might result from that the polyEAY is synthetized with distinct ratio of components (Glu, Ala, and Tyr), and the various composition of the sequences also led to the different catalytic efficiency. According to these characterizations of recombinant human TPST2, the NusA protein fused TPST2 expressed from E. coli was similar to that either from natual materials or eukaryotic expression. The NusA protein obviously did not affect the. 26.
(36) catalytic activity of human TPST2 and render the high solubility to facilitate TPST folding. In summary, we first purified TPST from prokaryote systems (E. coli) with catalytic activity. By means of this purification procedure, the time-and-effort-saving, inexpensive, high quality and quantity platform was established to express and purify homogenous human TPST2 with only one chromatography step for further biochemical characterization. The catalytic mechanism of substrate specificity, for example PSGL-1(Fig. 16), metal ion effect, and the regulatory residues will be examined. Furthermore, the crystal structure and antibody will be pursued to study in advanced for either the physiological or pathological functions and regulations.. 27.
(37) Table 1a. Some common and important post-translational modifications. a. Adapted from Mann et al. 2003.. 28.
(38) Table 2. Specific effects of tyrosine O-sulfationa. a. Adapted from Liu et al. 2008. Table 3a. Conservation of tyrosine sulfation sites in human chemokine receptors. Sulfation sites with scores of 2.5 or higher are in black. Sites with intermediate scores between 1.5 and 2.5 are in gray. a Adapted from Liu ,et al. 2008 29.
(39) Table 4. TPSTs purified from many source. Source. Mammalian cell. Sulfotransferase species. Type. Substrate. rat TPST. 3Y1 & SRA-3Y1. human TPST-HPC4. CHO-K1 cell. human TPST-HPC4. 293T cell. mouse TPST. 293T cell. human TPST-1 & 2. HEK293T cell. Reference. 3Y1 & SRA-3Y1. Liu M.C., et al. J. Biochem. 1987. PSGL-1. Ouyang Y., et al. Proc Natl Acad Sci. 1998. PSGL-1;CCR5. Seibert C.,et al. Proc Natl Acad Sci. 2002. PSGL-1. Mishiro E.,et al. J. Biochem.2006. 293T cell human TPST-1 & 2 embroynic kidney cell. Animal. rat (TPST). rat liver. EAY. Rens-Domiano S.,et al. J Biol. Chem. 1989. bovine (TPST). bovine adrenal medulla. CCK; Sgi. Niehrs C,et al. J Biol. Chem. 1990. Human (TPST). human liver. EAY. Lin W.H.,et al. Biochem. Pharmacol. 1990. bovine (TPST). bovine adrenal medulla. (PKG or others). Niehrs C,et al. EMBO J. 1990. rat Salivary Glands (TPST). Rat Salivary Glands. EAY. Kasinathan C,et al. Gen. Pharmacol. 1995. rat Salivary Glands (TPST). Rat Submandibular. EAY. William S.,et al. Arch. Biochem. Biophys. 1997. Salivary Glands rat liver. rat liver. PSGL-1. Ouyang Y. et al.Proc Natl Acad Sci. 1998. Rat Liver (TPST). Rat Liver. EAY. Ramaprasad P.,et al. Gen Pharmacol. 1998. rat TPST. rat liver. EAY. Kasinathan C.,et al. Alcohol. 1998. Human Saliva (TPST). Human Saliva. Statherin; EAY. Kasinathan C.,et al. Int. J. Biol. Sci. 2007. 30.
(40) Figure 1a Schematic representation of protein modifications related to the regulation of biological processes. a. Adapted from Seo et al. 2004. Figure 2 a. Sulfate activation and tyrosine O-sulfation. a Adapted from Moore et al. 2003. 31.
(41) Figure 3a. Schematic representation of cell entry by HIV-1 following sulfation of CCR5 by a tyrosylprotein sulfotransferase. a. Adapted from Chapman et al. 2004.. Y276, Y278, Y279. Figure 4a GPIbα from amino acid 200-294 a Adapted from Murata et al. 1991. 32.
(42) Figure 5 a. Tyrosine sulfation plays an important role in the immune response. a Adapted from Kehoe et al. 2000. 33.
(43) Figure 6. Graphical presentation of the far-Western immunoblot technique. ☆ = TPSTs, Y = Anti-TPST antibody, Y = HRP-linked secondary antibodie. O OH. -. O. S O O. R1. NH NH. R2. Tyrosylprotein sulfotransferases. O. NH. R1 NH O. NH2 N. O O O S O P O O. O. O. -. -2 O 3 PO. N. OH. N N. R2. NH2 O O P O. N O. N. -. O. -2. O3 PO. N N. OH. PAP. PAPS. Figure 7. The catalyzed reaction of tyrosylprotein sulfotransferase.. 34.
(44) Figure 8. Sequence alignment and transmembrane domain analysis of tyrosyl protein sulfotransferase. The sequence pairwise alignment is performed by ClustalW (http://www.ebi.ac.uk/Tools/clustalw2/index.html) and sorted shading by BOXSHADE server (http://www.ch.embnet.org/software/BOX_form.html). The black background indicated identity to each other and the gray one meant conserved substitutions. The residue colored in red is the predicted transmembrane domain calculated by PSIPRED (http://bioinf.cs.ucl.ac.uk/psipred/psiform.html) ranged from residue 6 to 28 both for human TPST1 and TPST2.. 35.
(45) hTPST2. kD M 116. 1. 66 45 35 25. Figure 9. Prokaryotic expression and purification of human hTPST2. The prokaryotic expression of human TPST2 was optimized to reach the maximal soluble amount and purified to near homogeneity. Lane 1 was purified from HisTrap chromatography followed the procedure as described in the “Experimental procedures.” Lane M was protein marker consisted of β-galactossidase (116.0 kDa), bovine serum albumin (66.2 kDa), ovalbumin (44 kDa), lactate dehydrogenase (35 kDa), and restriction endonuclease Bsp98I (25 kDa).. 36.
(46) Figure 10. The protein of LC-MS-MS fingerprinting analysis was identified for human TPST2. The excised spot from SDS-PAGE was identified for human TPST2 by LC-MS-MS. The sequence (red) by mass fingerprinting was mapped to the protein sequence with high confidence. (The result was particularly descrided in appendix). 37.
(47) 7. Sulfation number (pmole). 6 5 4 3 2 1 0. 0. 20. 40. 60. 80. 100. 120. 140. Time (mins). Figure 11. Reaction of polyEAY sulfation as a function of time. This experiment was performed as described under the “Experimental procedures.” Each assay consisted of 50 mM MES at pH 6.0, 25 mM MnCl2 , 50 mM NaF, 0.5 μM [35S]PAPS (15Ci/mmole), 40 μM TSH-R signaling polyEAY, 0.5 % Triton-X100 and 4 μg human TPST2 in a final volume of 10 μl. Data shown here is the result of a typical experiment performed in triplicate.. 38.
(48) Specific activity (pmole/min/mg). 6 5 4 3 2 1 0. 20. 25. 30. 35. 40. 45. 50. 55. Temperature (oC). Figure 12. Effect of Temperature on hTPST2 activity. This experiment was performed as described under the “Experimental procedures.” Using different temperature in the same mixture resulted in changes of the catalytic activity of human TPST2. Data shown here is the result of a typical experiment performed in triplicate.. 39.
(49) 6. Sepcific activity (pmole/min/mg). 5 4 3 2 1 0 -1 5.5. 6.0. pH. 6.5. 7.0. Figure 13. Different pH affects the catalytic activity of human TPST2. The experiment was performed as described under the “Experimental procedures.” The result indicated pH affects of tyrosine O-sulfation in human TPST2 enzyme assay. The data was performed in triplicate.. 40.
(50) Specific activity (pmole/min/mg). 7 6 5 4 3 2 1 0. A. B. 0. 5. 15 20 25 MnCl2 concentration 10. 30. 35. 40. Figure 14. Effect of MnCl2 on hTPST2 activity. This experiment was performed as described under the “Experimental procedures.”.Data shown here was the result of a typical experiment performed in triplicate. This experiment was repeated three times with similar results. Point A indicated the total reaction without polyEAY and MnCl2, and point B meant the total reaction without human TPST2 and MnC2.. 41.
(51) Specific Activity (pmol/min/mg). 5. 4. 3. 2. 1. 0 0. 10. 20. 30. 40. 50. EAY (microM). Figure 15. Kinetics of human TPST2 for polyEAY as substrate. Each assay consisted of 50 mM MES at pH 6.0, 25 mM MnCl2 , 50 mM NaF, 0.5 μM [35S]PAPS (15Ci/mmole), various concentrations of polyEAY, and 4 μg human TPST2 in a final volume of 10 μl. Data shown here was the result of a typical experiment performed in triplicate. The data indicated the Km was 10.5 ± 2.1 μM and Vmax was 4.8 ± 0.3 pmole/min/mg. This experiment was repeated three times with similar results.. 42.
(52) M. PSGL-1 1. 2. kDa 6. PSGL-1. Figure 16. The SDS-PAGE of PSGL-1 peptide purification. Lane M was protein marker consisted of β-galactossidase (116.0 kDa), bovine serum albumin (66.2 kDa), ovalbumin (44 kDa), lactate dehydrogenase (35 kDa), and restriction endonuclease Bsp98I (25 kDa). Lane 1 was the elution of GST-fused PSGL-1. Lane 2 was PSGL-1 peptide purified from GSTTrap chromatography after the thrombin digestion.. 43.
(53) Reference 1. Anastasi, A., Erspamer, V., and Endean, R. (1968) Isolation and amino acid sequence of caerulein, the active decapeptide of the skin of hyla caerulea. Arch. 2.. Biochem. Biophys. 125:57-68 van Anken E., and Braakman I. (2005) Versatility of the endoplasmic reticulum. 3.. protein folding factory, Crit. Rev. Biochem. Mol. Biol. 40:191–228 Baeuerle P.A., and Huttner W.B. (1986) Chlorate-a potent inhibitor of protein. 4.. sulfation in intact cells, Biochem. Biophys. Res. Commun. 141:870–877 Baeuerle P. A., and Huttner, W. B. (1987) Tyrosine sulfation is a. 5. 6.. trans-Golgi-specific protein modification. J. Cell Biol. 105: 2655–2664 Baumann E. (1876) Ber. Dtsch. Chem. Ges. 9:54-58 Bettelheim, F.R. (1954) Tyrosine-O-sulfate in a peptide from fibrinogen. J. Am.. 7.. 8. 9.. Chem. Soc. 76:2838–2839 Borghei A., Ouyang Y.B., and Westmuckett A.D. (2006) Marcello M.R., Landel C.P., Evans J.P., Moore K.L. Targeted disruption of tyrosylprotein sulfotransferase-2, an enzyme that catalyzes post-translational protein tyrosine O-sulfation, causes male infertility. J Biol Chem. 281:9423-9431 Bukau B., Weissman J., and Horwich A. (2006) Molecular chaperones and protein quality control, Cell. 125: 443–451 Bundgaard J.R., Vuust J., and Rehfeld J.F. (1997) New consensus features for tyrosine O-sulfation determined by mutational analysis. J Biol Chem.. 272:21700-21705 10. Chapman E., Best M.D., Hanson S.R., and Wong C.H. (2004) Sulfotransferases: structure, mechanism, biological activity, inhibition, and synthetic utility. Angew. Chem. Int. Ed. Engl. 43:3526-3548 11. Colvin R.A., Campanella G.S., Manice L.A., and Luster A.D. (2006) CXCR3 requires tyrosine sulfation for ligand binding and a second extracellular loop arginine residue for ligand-induced chemotaxis. Mol. Cell. Biol. 26:5838–5849 12. Davis, G.D., Elisee, C., Newham, D.M., and Harrison, R.G. (1999) New fusion protein systems designed to give soluble expression in Escherichia coli. Biotechnol. Bioeng. 65:382–388 13. Dong J.F., Li C.Q., and Lopez J.A. (1994) Tyrosine sulfation of the glycoprotein IbIX complex: identification of sulfated residues and effect on ligand binding. Biochemistry. 33:13946-13953 14. Dong J., Ye P., Schade A.J., Gao S., Romo G.M., Turner N.T., McIntire L.V., and López J.A. (2001) Tyrosine sulfation of glycoprotein Ibα: role of electrostatic interactions in von Willebrand factor binding. J. Biol. Chem. 276:16690-16694 44.
(54) 15. Farzan M., Mirzabekov T., Kolchinsky P., Wyatt R., Cayabyab M., Gerard N.P., Gerard C., Sodroski J., and Choe H. (1999) Tyrosine sulfation of the amino terminus of CCR5 facilitates HIV-1 entry. Cell. 96:667–676 16. Farzan M., Babcock G.J., Vasilieva N., Wright P.L., Kiprilov E., Mirzabekov T., and Choe H. (2002) The role of post-translational modifications of the CXCR4 amino terminus in stromal-derived factor 1 alpha association and hiv-1 entry. J. Biol. Chem. 227: 29484-29489 17. Fong A.M., Alam S.M., Imai T., Haribabu B., and Patel D.D. (2002) CX3CR1 tyrosine sulfation enhances fractalkine-induced cell adhesion. J. Biol. Chem. 277:19418–19423 18. Fredrickson B.J., Dong J.F., McIntire L.V., and Lo´pez J.A. (1998) Shear-dependent rolling on von Willebrand factor of mammalian cells expressing the platelet glycoprotein Ib-IX-V complex. Blood. 92:3684-3693. 19. Guo B., Yang S.H., Witty J., and Sharrocks A.D. (2007) Signalling pathways and the regulation of SUMO modification. Biochemical Society transactions. 35:1414-1418. 20. Hanai H., Nakayama D., Yang H., Matsubayashi Y., Hirota Y., and Sakagami Y. (2000) Existence of a plant tyrosylprotein sulfotransferase: novel plant enzyme catalyzing tyrosine O-sulfation of preprophytosulfokine variants in vitro. FEBS Lett. 470:97-101 21. Harrison, R.G. (2000) Expression of soluble heterologous proteins via fusion with NusA protein. inNovations 11:4–7 22. Hartley D.L., and Kane J.F. (1988) Properties of inclusion bodies from recombinant Escherichia coli. Biochem Soc Trans. 16:101-2 23. Hoffhines A.J., Damoc E., Bridges K.G., Leary J.A., and Moore K.L. (2006) Detection and purification of tyrosine-sulfated proteins using a novel anti-sulfotyrosine monoclonal antibody. J. Biol. Chem. 281:37877-37887 24. Hortin, G., Folz, R., Gordon, J.I., and Strauss, A.W. (1986) Characterization of sites of tyrosine sulfation in proteins and criteria for predicting their occurrence. Biochem. Biophys. Res. Commun. 141:326–333 25. Huttner, W.B. (1984) Determination and occurrence of tyrosine O-sulfate in proteins. Methods Enzymol. 107:200–223 26. Kasinathan C., Gandhi N., Ramaprasad P., Sundaram P., and Ramasubbu N. (2007) Tyrosine sulfation of statherin. Int. J. Biol Sci. 3:237-241. 27. Kasinathan C., Ramaprasad P., William S., and Espina N. (1998) Stimulation of tyrosylprotein sulfotransferase activity by ethanol: role of increased enzyme level. Alcohol. 15:271-276. 28. Kasinathan C., Ramaprasad P., and Sundaram P. (2005) Identification and 45.
(55) characterization of tyrosylprotein sulfotransferase from human saliva. Int. J. Biol. Sci. 1:141-145 29. Kasinathan C, Sundaram R, and William S. (1995) Effect of prostaglandins on tyrosylprotein sulfotransferase activity in rat submandibular salivary glands. Gen. Pharmacol. 26:577-580. 30. Kuhmann S.E., Platt E.J., Kozak S.L., and Kabat D. (2000) Cooperation of multiple CCR5 coreceptors is required for infections by human immunodeficiency virus type 1. J. Virol. 74:7005-7015 31. Lázaro-Diéguez F., Jiménez N., Barth H., Koster A.J., Renau-Piqueras J., Llopis J.L., Burger K.N., and Egea G. (2006) Actin filaments are involved in the maintenance of Golgi cisternae morphology and intra-Golgi pH. Cell. Motil. Cytoskeleton. 63:778-791 32. Lee, R. W. H., and Huttner, W. B. (1983) Tyrosine-O-sulfated proteins of PC12 pheochromocytoma cells and their sulfation by a tyrosylprotein sulfotransferase J. Biol. Chem. 258:11326–11334 33. Lee, R.W.H., and Huttner, W.B. (1985) (Glu62, Ala30, Tyr8)n serves as high-affinity substrate for tyrosylprotein sulfotransferase: a Golgi enzyme. Proc. Natl. Acad. Sci. U. S. A. 82:6143–6147 34. Leyte A., van Schijndel H.B., Niehrs C, Huttner W.B., Verbeet M.P., Mertens K., and van Mourik J.A. (1991) Sulfation of Tyr1680 of human blood coagulation factor VIII is essential for the interaction of factor VIII with von Willebrand factor. J Biol Chem. 266:740-746. 35. Lin H.C., Tsai K., Chang B.L., Liu J., Young M., Hsu W., Louie S., Nicholas H.B., and Jr, Rosenquist G.L. (2003) Prediction of tyrosine sulfation sites in animal viruses. Biochemical and Biophysical Research Communications 312:1154–1158 36. Lin WH, Larsen K, Hortin GL, and Roth JA. (1992) Recognition of substrates by tyrosylprotein sulfotransferase. Determination of affinity by acidic amino acids near the target sites. J Biol Chem. 267:2876-2879 37. Lin W.H., and Roth J.A. (1990) Characterization of a tyrosylprotein sulfotransferase in human liver. 40:629-635. 38. Liu J., Louie S., Hsu W., Yu K.M., Nicholas H.B., and Jr, Rosenquist G.L. (2008) Tyrosine Sulfation is prevalent in human chemokine receptors important in lung disease. Am. J. Respir. Cell. Mol. Biol. 38:738–743. 39. Liu M. C., Suiko M., and Lipmann F. (1987) Studies of the decrease of tyrosine-O-sulphated proteins in Rous sarcoma-virus-transformed rat embryo fibroblasts, line 3Y1. Examination of the sulphate activation and tyrosyl-protein sulphotransferase systems. J. Biochem . 247: 201–206. 46.
(56) 40. Mann M, and Jensen O.N. (2003) Proteomic analysis of post-translational modifications. Nat. Biotechnol. 21:255-261. 41. Marchese P., Murata M., Mazzucato M., Pradella P., De Marco L., Ware J., and Ruggeri Z.M. (1995) Identification of three tyrosine residues of glycoprotein Ibα with distinct roles in von Willebrand factor and α-thrombin binding. J. Biol. Chem. 270:9571- 9578. 42. Marston F.A. (1986) The purification of eukaryotic polypeptides synthesized in Escherichia coli Biochem J. 240:1-12. 43. Mishiro E., Liu M.Y., Sakakibara Y., Suiko M., and Liu M.C. (2004) Zebrafish tyrosylprotein sulfotransferase: molecular cloning, expression, and functional characterization. Biochem Cell Biol. 82:295-303. 44. Mishiro E., Sakakibara Y., Liu M.C., and Suiko M. (2006) Differential enzymatic characteristics and tissue-specific expression of human TPST-1 and TPST-2. J Biochem. 140:731-737. 45. Moore K.L. (2003) The biology and enzymology of protein tyrosine O-sulfation. J. Biol. Chem. 278:24243-24246. 46. Murata M., Ware J., and Ruggeri Z.M., (1991) Site-directed mutagenesis of a soluble recombinant fragment of platelet glycoprotein Ibα demonstrating negatively charged residues involved in von Willebrand factor binding. J. Biol. Chem. 266:15474-15480. 47. Nicholas H.B., and Jr, Chan S.S. (1999) Rosenquist G.L. Reevaluation of the determinants of tyrosine sulfation, Endocrine. 11:285-292 48. Niehrs C., and Huttner W.B. (1990) Purification and characterization of tyrosylprotein sulfotransferase. EMBO J. 9:35-42 49. Niehrs C, Kraft M, Lee RW, Huttner WB. (1990) Analysis of the substrate specificity of tyrosylprotein sulfotransferase using synthetic peptides. J. Biol. Chem. 265:8525-8532. 50. Ouyang, Y., Lane, W. S., and Moore, K. L. (1998) Tyrosylprotein sulfotransferase: purification and molecular cloning of an enzyme that catalyzes tyrosine O-sulfation, a common posttranslational modification of eukaryotic proteins. Proc. Natl. Acad.Sci. U.S.A. 95: 2896 51. Ouyang, Y.B., Crawley, J.T., Aston, C.E., and Moore, K.L. (2002) Reduced body weight and increased postimplantation fetal death in tyrosylprotein sulfotransferase-1-deficient mice. J. Biol. Chem. 277:23781–23787 52. Preobrazhensky A.A., Dragan S., Kawano T., Gavrilin M.A., G0lina I.V., Chakravarty L., and Kolattukudy P.E. (2000) Monocyte chemotactic protein-1 receptor CCR2B is a glycoprotein that has tyrosine sulfation in a conserved extracellular n-terminal region. J Immunol 47. 165:5295–5303.
(57) 53. Pouyani, T., and Seed, B. (1995) PSGL-1 recognition of P-selectin is controlled by a tyrosine sulfation consensus at the PSGL-1 amino terminus. Cell. 83:333-343 54. Ramaprasad P., and Kasinathan C. (1998) Isolation of tyrosylprotein sulfotransferase from rat liver. Gen. Pharmacol. 30:555-559 55. Ramaprasad P., and Kasinathan C. (1998) In vivo induction of tyrosylprotein sulfotransferase by ethanol: role of increased enzyme synthesis. Alcohol. Clin. Exp. Res. 22:1120-1124 56. Rens-Domiano S., and Roth J.A. (1989) Characterization of tyrosylprotein sulfotransferase from rat liver and other tissues. J. Biol. Chem. 264:899-905. 57. Sako D., Comess K.M., Barone K.M., Camphausen R.T., Cumming D.A., and Shaw G.D. (1995) A sulfated peptide segment at the amino terminus of PSGL-1 is critical for P-selectin binding. Cell. 83, 323-331 58. Sane D.C, and Baker M.S., (1993) Human platelets possess tyrosylprotein sulfotransferase (TPST) activity. Thromb Haemost. 69:272-275 59. Seibert C., Cadene M., Sanfiz A., Chait B.T, and Sakmar T.P. (2002) Tyrosine sulfation of CCR5 N-terminal peptide by tyrosylprotein sulfotransferases 1 and 2 follows a discrete pattern and temporal sequence. Proc Natl Acad Sci. 99:11031-11036. 60. Seo J., and Lee K.J. (2004) Post-translational modifications and their biological functions: proteomic analysis and systematic approaches. J. Biochem. Mol. Biol. 37:35-44 61. Shevchenko A., Wilm M., Vorm O. and Mann M. (1996) Mass spectrometric sequencing of proteins from silver stained polyacrylamide gels. Anal. Chem. 68:850-858 62. Stone S.R., and Hofsteenge J. (1986) Kinetics of the inhibition of thrombin by hirudin. Biochemistry. 25:4622-4628 63. Suiko M., and Liu M.C. (1988) Change in binding affinities of 3Y1 secreted fibronectin upon desulfation of tyrosine-O-sulfate. Biochem. Biophys. Res. Commun. 154:1094-1098 64. United States Patent, patent number : 5716836. 65. Ward C.M., Andrews R.K., Smith A.I., and Berndt M.C. (1996) Mocarhagin, a novel cobra venom metalloproteinase, cleaves the platelet von Willebrand factor receptor glycoprotein Ibα Identification of the sulphated tyrosine/anionic sequence Tyr-276–Glu-282 of glycoprotein Ibαas a binding site for von Willebrand factor and α-thrombin. Biochemistry. 35:4929-4938 66. Westmuckett A.D., Hoffhines A.J., Borghei A., and Moore K.L. (2008) Early postnatal pulmonary failure and primary hypothyroidism in mice with 48.
(58) combined TPST-1 and TPST-2 deficiency. Gen Comp Endocrinol. 156:145-153 67. William S., Ramaprasad P., and Kasinathan C. (1997) Purification of tyrosylprotein sulfotransferase from rat submandibular salivary glands. Arch. Biochem. Biophys. 338:90-96. 68. Wilkins, P.P., Moore, K.L., McEver, R.P. and Cummings, R.D. (1995) Tyrosine sulfation of P-selectin glycoprotein ligand-1 is required for high affinity binding to P-selectin. J. Biol. Chem. 270:22677-22680 69. William S., Ramaprasad P., and Kasinathan C. (1997) Purification of tyrosylprotein sulfotransferase from rat submandibular salivary glands. Arch. Biochem. Biophys. 338:90-96 70. Wu Y., Li Q., and Chen X.Z. (2007) Detecting protein-protein interactions by Far western blotting. Nat. Protoc. 2:3278-3284 71. Young J.C., Agashe V.R., Siegers K., and Hartl F.U. (2004) Pathways of chaperone-mediated protein folding in the cytosol, Nat. Rev. Mol. Cell. Biol. 5:781–791 72. Zhao Y., Sato Y., Isaji T., Fukuda T., Matsumoto A., Miyoshi E., Gu J., and Taniguchi N. (2008) Branched N-glycans regulate the biological functions of integrins and cadherins. Federation of European Biochemical Societies. 275:1939-1948. 73. Zlotnik A., Morales J. and Hedrick J.A. (1999) Recent advances in chemokines and chemokine receptors. Crit. Rev. Immunol. 19:1-47. 49.
(59) Appendix 1. 1. FJEC Mass: 54837 Score: 858 Queries matched: 25 transcription termination-antitermination factor nusA - Escherichia coli (strain K-12) 2. AAG41947 Mass: 66027 Score: 352 Queries matched: 8 AF304164 NID: - Homo sapiens 3. Q4R863_MACFA Mass: 41823 Score: 254 Queries matched: 6 Testis cDNA clone: QtsA-13311, similar to human tyrosylprotein sulfotransferase 2 ),.- Macaca fascicularis (Crab eating macaque) (Cynomolgus monkey) 50.
(60) 4. Q4VAQ2_HUMAN Mass: 65393 Score: 176 Queries matched: 3 Keratin 2A (Epidermal ichthyosis bullosa of Siemens).- Homo sapiens (Human). 5. BAC29428 Mass: 42056 Score: 165 Queries matched: 4 AK036436 NID: - Mus musculus 6. KRHU0 Mass: 59492 Score: 147 Queries matched: 3 keratin 10, type I, cytoskeletal - human 7. K2C1_MOUSE Mass: 65052 Score: 144 Queries matched: 3 Keratin, type II cytoskeletal 1 (Cytokeratin-1) (CK-1) (Keratin-1) (K1) 8. Q2C7T3_9GAMM Mass: 54979 Score: 122 Queries matched: 5 Transcription elongation factor NusA.- Photobacterium sp. SKA34. 9. Q6IG03_RAT Score: 119 Queries matched: 2 Type II keratin Kb36.- Rattus norvegicus (Rat). 10. Q7VLI1_HAEDU Mass: 55131 Score: 103 Queries matched: 3 Transcription termination-antitermination factor nusA, N utilization substance protein A 11. Q8D2X5_WIGBR Mass: 55614 Score: 101 Queries matched: 4 NusA protein.- Wigglesworthia glossinidia brevipalpis. 12. Q3TTY5_MOUSE Mass: 70880 Score: 89 Queries matched: 2 10 days neonate skin cDNA, RIKEN full-length enriched library, clone:4732404G19 product: 13. BAA19418 Mass: 25913 Score: 86 Queries matched: 2 AB001594 NID: - Homo sapiens 14. TRPGTR Mass: 24394 Score: 85 Queries matched: 2 trypsin (EC 3.4.21.4) precursor - pig (tentative sequence) 15. Q1V532_VIBAL Mass: 54909 Score: 69 Queries matched: 2 Transcription elongation factor NusA.- Vibrio alginolyticus 12G01. 16. AAH03630 Score: 68 Queries matched: 2 BC003630 NID: - Homo sapiens 17. Q1ZCR5_9GAMM Mass: 55527 Score: 68 Queries matched: 2 Transcription elongation factor NusA.- Psychromonas sp. CNPT3. 18. Q32MB2_HUMAN Score: 65 Queries matched: 2 K6IRS3 protein.- Homo sapiens (Human). 19. Q163L6_ROSDE Mass: 25428 Score: 56 Queries matched: 1 Oxidoreductase, short chain dehydrogenase/reductase family (EC 1.1.1.47).Roseobacter. 51.
(61) Appendix 2. 52.
(62)
數據

+7


相關文件
Valor acrescentado bruto : Receitas do jogo e dos serviços relacionados menos compras de bens e serviços para venda, menos comissões pagas menos despesas de ofertas a clientes
•Last month I watched a dance class in 崇文 Elementary School and learned the new..
2.1.1 The pre-primary educator must have specialised knowledge about the characteristics of child development before they can be responsive to the needs of children, set
To stimulate creativity, smart learning, critical thinking and logical reasoning in students, drama and arts play a pivotal role in the..
Reading Task 6: Genre Structure and Language Features. • Now let’s look at how language features (e.g. sentence patterns) are connected to the structure
Promote project learning, mathematical modeling, and problem-based learning to strengthen the ability to integrate and apply knowledge and skills, and make. calculated
The teacher explains to learners their duties: to present their ideas and findings on the questions on their role sheet, and lead the other group members to discuss the
This is especially important if the play incorporates the use of (a) flashbacks to an earlier time in the history of the characters (not the main focus of the play, but perhaps the
