行政院國家科學委員會專題研究計畫 成果報告
日本鰻棲地利用與族群遺傳結構之分析
研究成果報告(精簡版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2313-B-002-119- 執 行 期 間 : 95 年 10 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學漁業科學研究所 計 畫 主 持 人 : 韓玉山 計畫參與人員: 碩士班研究生-兼任助理:孫宇樑 處 理 方 式 : 本計畫可公開查詢中 華 民 國 96 年 10 月 02 日
計畫中文摘要 日本鰻在傳統上被認為是降河洄游性魚類,但近年來發現鰻魚在黃鰻時期利用淡水 棲地的現象並非絕對。本研究想要探討日本鰻的性比是否與其棲地利用相關,並且想知 道不同棲地利用的族群間是否有遺傳分化的現象存在。我們於高屏溪河口採集了 221 隻 日本鰻樣本,利用 EPMA 分析其耳石鍶/鈣比,並利用 6 組微衛星 DNA 分析不同棲地 族群間之遺傳分化情形。 根據日本鰻耳石鍶/鈣比的生活史型態,將之分成三型: 第一 型 (淡水型)、 第二型 (半淡鹹水偏淡水型) 與 第三型 (半淡鹹水偏海水型)。 雄性與雌 性日本鰻之棲地利用有顯著的不同。 雌鰻主要集中在第二型與第三型,而雄鰻則集中在 第一型與第二型。雌鰻偏好半淡鹹水棲地而雄鰻則偏好淡水棲地。此外,統計距耳石核 心 200-400 μm 區間之平均鍶/鈣比,此區間約當日本鰻性別分化時期,發現雌鰻之平均 值顯著高於雄鰻。 此結果暗示日本鰻的性別分化可能與其棲地利用有關,亦即生活在半 淡鹹水棲地之鰻魚傾向分化為雌性,而生活在淡水棲地之鰻魚則傾向分化為雄性。此外, 是否在不同鹽度的棲地中,鰻魚會有不同的遺傳結構組成,本計劃仍在進行中,約半年 內可分析完成。 關鍵詞:日本鰻、鍶鈣比、性比、族群遺傳結構、微衛星DNA、性別分化 計畫英文摘要
The Japanese eel is traditionally known as a catadromous fish but it has recently been discovered that the use of freshwater habitat at the yellow eel stage is facultative. To determine if habitat use by Japanese eels differs between the sexes and population genetic structure differs among eels of various habitats, we examined the Sr/Ca ratios in otoliths by electron probe micro analyzer and 6 microsatellite DNA loci of 221 eels to reconstruct their environmental history and identify the population genetic variety. Eels were collected from the Kaoping River estuary of southwestern Taiwan from 1998 through 2005. The habitat use of yellow-phase eels was divided into 3 types according to the life history pattern of the otolith Sr/Ca ratios: Type1 (freshwater resident), Type2 (brackish water resident with a freshwater preference) and Type3 (brackish water resident with a seawater preference). Habitat use differed significantly between male and female silver eels. Females were predominantly in Type2/3 while males were predominantly in Type1/2. Consequently, yellow stage females preferred an estuarine habitat while males preferred a freshwater habitat. In addition, the mean otolith Sr/Ca ratios from 200-400 μm from the primordium, which corresponded to the period of sex differentiation were higher in females than in males. This indicated that the sex differentiation of the eel might be related to habitat use, i.e., brackish water eels tended to differentiate as females and freshwater eels as males. In addition, the genetic structures among these 3 types of eel populations are still in progress. The final analysis will be done within half year.
Key words: Japanese eel, otolith Sr/Ca ratio, sex-ratio, population genetic structure, microsatellite DNA, sex differentiation
一、前言
The Japanese eel (Anguilla japonica) is a catadromous fish, spawning in the open ocean west of the Mariana Islands, 14°N 142-143°E (Tsukamoto, 2006) and growing in fresh water (Tesch 2003). Their larvae, leptocephali, drift with the North Equatorial and Kuroshio Currents to east Asia, metamorphose to glass eels on the continental shelf, and become elvers in the estuary, taking about 4-6 months in total (Cheng and Tzeng 1996). The elvers then grow as yellow eels in rivers for approximately 4-10 years (Tzeng 2000, Han et al. 2003a) before metamorphosing to silver eels and migrating back to the spawning ground to spawn and die.
In the recent years, otolith growth patterns and chemical compositions have provided detailed chronologies of habitat use for individual eels (Campana 1999). Otolith strontium/calcium (Sr/Ca) ratio levels are correlated with the ambient salinity and thus were used to trace the salinity life history of the eels (Tzeng et al. 1997, Campana 1999). The eel’s life history strategy is not only complicated but also plastic. Some of the American, European and Japanese eels may skip the freshwater phase and can grow up in the brackish water and seawater until silver eel stage (Tzeng et al. 1997, 1999, 2000, Tsukamoto et al. 1998, Jessop et al. 2004). The eel also has different migratory contingents and should be regarded as a facultatively catadromous fish (Tsukamoto and Arai 2001, Tzeng et al. 2002a, 2003, Jessop et al. 2004,). Although the migratory behavior of the eel is well documented, differences in habitat preference between the sexes, however, are still poorly understood.
The sex composition of wild eels in each habitat varies widely, ranging from almost all males to predominantly females (Matsui 1972, Tzeng et al. 2002a, Oliveira 1999, Oliveira et al. 2001, Tesch 2003, Han and Tzeng 2006). The uneven distributions of sexes in habitats must result from either different habitat choices by each sex or significant effects on their sex differentiation by the habitat in which they grow. The sexual differentiation of the eel is labile and thought to be environmentally determined and to occur at the yellow eel stage around 15-35 cm (Matsui 1972, Colombo and Grandi 1996, Krueger and Oliveira 1999). Individual growth rate (Helfman et al. 1987, Holmgren and Mosegard 1996), population density (Parsons et al. 1977, Krueger and Oliveira 1999, Tesch 2003, Han and Tzeng 2006), temperature (Holmgren 1996), latitude (Helfman et al. 1987) and river types (Sinha and Jones 1967, Oliveira et al. 2001) are possible extrinsic cues. The higher percentage of males in estuarine habitat than in freshwater in some studies had led to the suggestion that higher salinity water might favor male development (Sinha and Jones 1967, Helfman et al. 1987, Tesch 2003). However, many studies observed the opposite distribution, with males predominant in freshwater and females predominant in brackish water (Sinha and Jones 1967, Holmgren 1996, Kotake et al. 2003, 2005). In the estuary habitats, the changes in salinity are highly variable and eels can choose the preferred salinity microhabitat (Han et al. 2003b). The migratory behavior of some eels between freshwater and seawater habitats further disturbs the possible effect of salinity on sex-determination. Thus, the use of chronologies of salinity history recorded in the otoliths of individual eels as revealed by Sr/Ca ratios (Tzeng et al. 1997, Campana 1999) may
enable assessment of whether habitats with different salinity will affect eel sex-determination.
二、研究目的
This study aims to identify different strategies of habitat selection by Japanese eels of different sex by analyzing otolith Sr/Ca ratios and to assess the potential effect of different habitat use by juvenile eels on their future sex determination. In addition, another purpose of this study is to test the diversity of population genetic structure among the three contingents
using 6 microsatellite loci. Results from sex-dependent habitat use and the genetic diversity among the three contingents can be applied to eel stock releasing strategy.
三、研究方法
Sample collection. Wild Japanese eels were collected monthly with eel traps in the estuary of the
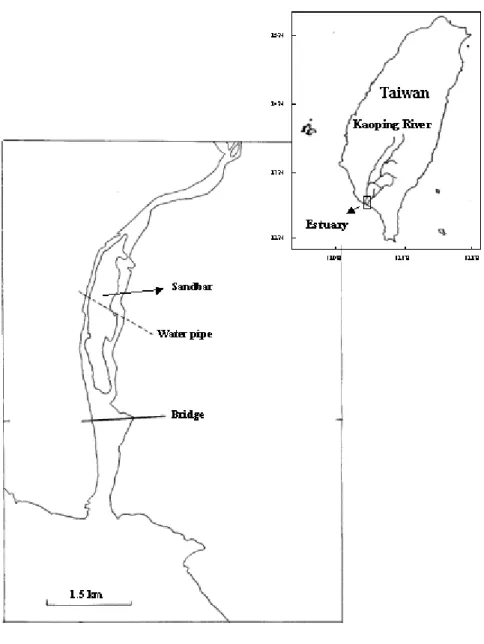
Kaoping River in southwestern Taiwan (120°50’E, 22°40’N) during the period from 1998 to 2005 (Fig. 1). The eel pots were set at the bottom of the riverbank in the estuary, where the salinity variated between 0-30 ‰ influenced by both the freshwater and tide. Total length (TL, ± 1 mm) and body weight (BW, ± 0.1 g) of the eels were measured. The sex and developmental stage of each eel were determined by visual inspection or by histology when possible (Han et al. 2003a).
Otolith preparation for Sr/Ca ratio analysis. A total of 221 Japanese eels, which were selected
from batches collecting in the late autumn and winter when most silver eels occurred, were used for otolith analysis. This included 39 silver males, 2 yellow males, 64 yellow females, 87 silver females, and 29 unknown-sex juveniles. Sagittal otoliths, the largest of the three pairs of otoliths in the inner ear, were used for Sr/Ca ratio analysis and age determination. The procedure for preparation of the otolith for Sr/Ca ratio analysis followed Tzeng et al. (1997). For electron probe microanalysis, the polished otoliths were coated with carbon under a high vacuum evaporator. Strontium (Sr) and calcium (Ca) concentrations (wt %) in the otolith were measured from the primordium to the edge of otolith at an interval of 10 μm with an electron beam of 5 μm in diameter, using an electron probe microanalyzer (EPMA, JEOL JXA-800M). After microchemistry analysis, the otolith was polished to remove the carbon layer, etched 1 to 2 minutes with 5% EDTA (PH = 7.4) to reveal the annular marks for age determination (Tzeng et al. 1997). Previous studies had identified divergent migratory contingents of Japanese eel for samples coming from upstream of river and lake, estuary and marine habitats (Tzeng et al. 2003). The Sr/Ca ratio of 4 ‰ was used as the boundary to discriminate freshwater and brackish water habitats. Based on this, the eel was classified into different migratory contingents.
DNA extraction. Genomic DNA extraction was carried out from a small piece of muscle tissues of
the glass eels. Extraction was done using a DNA purification and extraction kit (Bioman Scientific Ltd.). Extracted DNA was stored under -20 ℃ temperature before polymerase chain reaction
(PCR).
PCR and genotyping. Six microsatellite loci were selected including 5 (GT)n (AJM-1, AJMS-3,
AJMS-5, AJMS-6 and AjTR-37) loci and 1 (GA)n (AjTR-12) loci. The Accession numbers were AM062762 (AJM-1), AJ297601 (AJMS-3), AJ297602 (AJMS-5), AJ297603 (AJMS-6), AB051084 (AjTR-12) and AB051094 (AjTR-37). Microsatellites were amplified via the PCR. Reverse primers contain FAM, TAMRA or HEX fluorescence labels for genotyping. 1 μl of PCR product was diluted with 12 μl of Milli-Q H2O for genotyping performed using a capillary Megabase 1000 DNA
analysis system (Amersham Biosciences). Scoring of data was done by Genetic ProfilerTM Version 2.0 (Amersham Biosciences) and the sizes of each allele were manually checked with the naked eye.
四、結果
Otolith Sr/Ca ratio patterns
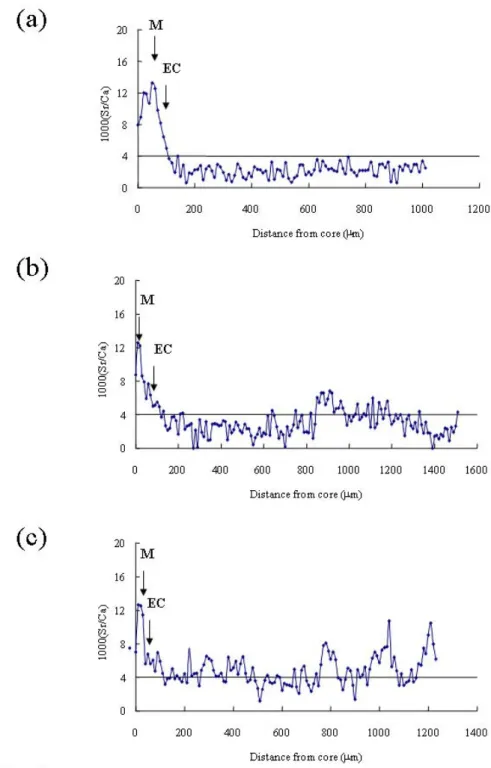
The Sr/Ca ratios in eel otoliths before the elver stage, approximately 0-150 μm from the primordium, were similar among individuals, indicating that the migratory environmental history was similar among individuals during the marine leptocephalus stage (Fig. 2). However, otolith Sr/Ca ratios beyond the elver stage varied markedly and were classified into 3 types of migratory contingents (Fig. 2) described as follows: Type 1 (freshwater resident), the >90 % of Sr/Ca ratios in the otolith beyond the elver check fluctuated below 4 ‰, indicating that the eels after elver stage
had been migrating to freshwater habitat to grow until metamorphosis to silver eel (Fig.2a). Type 2 (brackish water resident with freshwater preference), the 50-90 % of Sr/Ca ratios in the otolith beyond the elver check were lower than 4 ‰,indicating that the eels after elver stage had been migrating between freshwater and brackish water but showed a freshwater preference (Fig. 2b). Type 3 (brackish water resident with seawater preference), the > 50 % of Sr/Ca ratios in the otolith beyond the elver check were higher than 4 ‰,indicating that the eels after elver stage had been migrating between freshwater and brackish water but showed a brackish preference (Fig. 2c).
Sex-dependent habitat use in the Kaoping River estuary
From 1998 through 2005, a total of 221 Japanese eels collected in the Kaoping River estuary were analyzed (Table 1). More silver stage eels were chosen in order to trace their whole environmental life history. The mean age, TL, and BW of the silver females were all largest in silver females and lowest in the silver males (age: F = 23.2, df = 3, p < 0.001; TL: F = 110.2, df = 3, p < 0.001; BW: F = 76.7, df = 3, p < 0.001) (Table 1). The distribution of the 3 type life histories of the eels varied between sexes (Table 2). Silver stage males and females differed significantly in habitat use (χ2 = 22.52, df = 2, p < 0.005) (Table 2). There was no significant difference between females of yellow and silver stages (χ2 = 7.91, df = 2, 0.01< p <0.025) (Table 2). Male silver eels predominated
in the Type 1 (54%) and Type 2 (38%) contingents and silver females were dominant in the Type 2 (47%) and 3 (37%) contingents (Table 2).
Habitat-related sex determination between sexes
The sex differentiation of the eels occurs during the juvenile stage around 15-35 cm for the temperate eel species of Japanese, American and European eels (Matsui, 1972; Helfman et al., 1987; Kruger and Oliveira, 1999). This approximately corresponded to the region of eel otoliths 200-400 μm from the primordium, as evaluated by back-calculated growth histories from annual otolith increments (Tzeng et al. 2000). The mean Sr/Ca ratio of female eels was significantly higher (p = 0.001) than for males during the supposed eel sex-differentiated period (Table 3). Mean otolith Sr/Ca ratios beyond the elver check also differed between the sexes, with higher mean values for females than for males (p < 0.001). The mean otolith Sr/Ca ratios between 200-400 μm and beyond the 150 μm from the primordium were not significant in both the male (p = 0.308) and female (p = 0.437).
五、討論
Life-history scans of otolith Sr/Ca ratios has validated that yellow-stage Japanese eels occupy habitats that included freshwater and brackish water with some eels migrating between the two habitats (Fig. 2). The diverse life-history strategies of eels indicated that freshwater residence of eels during the growth phase is opportunistic, not obligatory (Tsukamoto and Arai 2001, Kotake et al. 2003, 2005). The rivers in Taiwan, however, are mostly small without stable freshwater input, and thus most eels may be compelled to grow in the estuary (Tzeng et al. 2002a). The catch experience of local fishermen indicates that Japanese eels are seldom caught in the middle and upper reaches of the Kaoping River, and there are many weirs along the River that may block the upstream migration of the eels. Thus, most of the Japanese eels are forced to concentrate in the estuary of Kaoping River (Han and Tzeng 2006). Consequently, most individual eels analyzed in this study should come from the estuarine stock. The fluctuant salinity in the Kaoping River estuary (0-30 ‰) provides diverse microhabitats, from almost freshwater up the sandbar of estuary to near seawater down the bridge of estuary (Fig. 1), for the eel to select. Since silver stage eels have almost finished their entire continental life, their otolith Sr/Ca ratios represent their whole environmental life history. Most (84%) silver females showed brackish water preference (Type 2 and 3) (Table 2). In contrast, most (54%) silver males preferred the freshwater habitat (Type 1) (Table 2). There was also significant difference on the mean Sr/Ca ratios between sexes. Beyond the elver stage, the mean otolith Sr/Ca ratio of female eels was higher than for males (Table 3). Consequently, male and female Japanese eels showed different habitat preferences, with males preferring freshwater habitat and females favoring brackish water one.
sex-determination of young juvenile stage eels may also have been affected by salinity. The mean Sr/Ca ratios differed significantly between sexes not only beyond elver stage but also during the time window in which eel sex-determination occurred (Table 3). This indicated that the habitat use not only differed between the sexes but the habitat itself might also affect eel sex determination, e.g., high salinity prefers female sex differentiation and freshwater prefers male sex differentiation. The environmental sex determination (ESD) has been proved in many species (Docker and Beamish 1994), and ESD evolves when an environmental factor is more advantageous to one sex than to the other, as the salinity factor in the case of eel. However, many other environmental factors that might affect eel sex are also being reported. Individuals experiencing rapid growth in the juvenile stage prior to sex differentiation tend to develop as males, whereas eels that grow slowly initially are more likely to develop into females (Helfman et al. 1987, Holmgren and Mosegard 1996). Males tend to predominate at high environmental densities while females predominate at low density (Krueger and Oliveira 1999, Tesch 2003, Han and Tzeng 2006). High temperature conditions have also been proposed to favor development as males (Holmgren 1996). Some studies showed that the proportion of females of migratory silver eels tends to increase with latitude (Helfman et al. 1987). However, the interrelated environmental factors that might be responsible for eel sex determination make it difficult to evaluate their relative contribution and dominance. Kotake et al. (2003, 2005) study the migratory history of the Japanese eel in the Amakusa Islands and Mikawa Bay of Japan by otolith Sr/Ca ratios. They find high percentage of females in seawater and brackish waters, and most river type of the eel is the male. The better feeding conditions and lower population density in the coast areas of Japan than in the streams are used to explain the habitat- dependent sex determination of the eel. In the Kaoping River of Taiwan, seawater eels are rare in contrast to the study in Amakusa Islands and Mikawa Bay of Japan. This may be due to the difference on habitat, with the marine habitats in the sampling site of Japan but estuary habitat in that of Taiwan. The sex ratio of the Japanese eel in the Kaoping River estuary was strongly skewed towards females, which was believed due to the low-density effect (Han and Tzeng 2006).
Kotake et al. (2005) report that after sex determination, more females may tend to move to marine or brackish water habitats, while males may remain in the freshwater until spawning migration. In the present study, the percentage of freshwater (type 1) females is slightly higher in yellow than in silver stages, while percentage of brackish water (type 2 and 3) females is slightly lower in yellow than in silver stage. However, no significant difference is found. In addition, mean otolith Sr/Ca ratios between 200-400 μm and beyond the elver check are similar for each sex, indicating that the sex differentiated individuals may tend to stay in the similar habitats without sex-dependent difference on habitat shift.
In summary, sex-undetermined eels in the Kaoping River estuary tended to develop as females in a brackish water habitat and as males in a freshwater habitat. Sexually differentiated individuals continued to show a sex-specific habitat preference. Phenotypic-dependent eel sex determination may have evolved as an adaptation to a spatially heterogeneous habitat. The sex determination of
the eel may be facultative and determined by multiple environmental factors, which vary among habitats. More studies are needed to clarify the mechanism of how extrinsic factors affect eel sex determination and subsequent habitat selection.
六、參考文獻
Cheng PW, Tzeng WN (1996) Timing of metamorphosis and estuarine arrival across the dispersal range of the Japanese eel Anguilla japonica. Mar Ecol Prog Ser 131:87-96
Colombo G, Grandi G. (1996) Histological study of the development and sex differentiation of the gonad in the European eel. J Fish Biol 48:493-512
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms, and applications. Mar Ecol Prog Ser 188:263-297
Docker MF, Beamish FWH (1994) Age, growth, and sex ratio among populations of least brook lamprey, Lampetra aepyptera, larvae: an argument for environmental sex determination. Env Biol Fish 41:191-205
Han YS, Tzeng WN (2006) Sex ratio as a means of resource assessment for the Japanese eel
Anguilla japonica: A case study in the Kaoping River of Taiwan. Zool Stud 45: 255-263
Han YS, Liao IC, Huang YS, He JT, Chang CW, Tzeng WN (2003a) Synchronous changes of morphology and gonadal development of silvering Japanese eel Anguilla japonica. Aquaculture 219:783-796
Han YS, Yu JYL, Liao IC, TzengWN (2003b) Salinity preference of the silvering Japanese eel
Anguilla japonica: evidences from the pituitary prolactin mRNA levels and otolith
strontium/calcium ratios. Mar Ecol Prog Ser 259:253-261
Helfman GS, Facey DE, Stanton HL, Bozeman EL (1987) Reproductive ecology of the American eel. Am Fish Soc Symp 1:42-56
Holmgren K (1996) Effect of water temperature and growth variation on the sex ratio of experimentally reared eels. Ecol Freshw Fish 5:203-212
Holmgren K, Mosegaard H (1996) Implications of individual growth status on the future sex of the European eel. J Fish Biol 49:910-925
Jessop BM, Shiao JC, Iizuka Y, Tzeng WN (2004) Variation in the annual growth, by sex and migratory history, of silver American eels Anguilla rostrata. Mar Ecol Prog Ser 272:231-244 Kotake A, Arai T, Ozawa T, Nojima S, Miller MJ, Tsukamoto K (2003) Variation in migratory
history of Japanese eels, Anguilla japonica, collected in coastal waters of the Amakusa Islands ,Japan, inferred from otolith Sr/Ca ratios. Mar Ecol 142:849-854
Kotake A, Okamura A, Yamada Y, Utoh T, Arai T, Miller MJ, Oka HP, Tsukamoto K (2005) Seasonal variation in migratory history of Japanese eel Anguilla japonica in Mikawa Bay,Japan. Mar Ecol Prog Ser 293:213-221
Krueger W, Oliveira K (1999) Evidence for environmental sex determination in the American eel,
Anguilla rostrata. Environ Biol Fish 55:381-389
Matsui I (1972) Unagigaku: eel biology. Kosei-sha Kosei-Kaku, Tokyo
Can J Fish Aquat Sci 56:795-802
Oliveira K, McCleave JD, Wippelhauser GS (2001) Regional variation and the effect of lake: river area on sex distribution of American eels. J Fish Biol 58:943-952
Parsons J, Vickers KU, Warden Y (1977) Relationship between elver recruitment and changes in the sex ratio of silver eels Anguilla anguilla L. migrating from Lough Neagh, Northern Ireland. J Fish Biol 10:211-229
Sinha VRP, Jones JW (1967) On the age and growth of the freshwater eel (Anguilla anguilla). J Zool 153:119–137
Tesch FW (2003) The eel. Blackwell Science, Oxford, UK
Tsukamoto K, Nak I, Tesch WV (1998) Do all fresh water eels migrate? Nature 396:635-636
Tsukamoto K (2006) Spawning of eels near a seamount: Tiny transparent larvae of the Japanese eel collected in the open ocean reveal a strategic spawning site. Nature 493:929
Tsukamoto K, Arai T (2001) Facultative catadromy of the eel Anguilla japonica between freshwater and seawater habitats. Mar Ecol Prog Ser 220:265-276
Tzeng WN, Severin KP, Wickström H (1997) Use of otolith microchemistry to investigate the environmental history of European eel Anguilla anguilla. Mar Ecol Prog Ser 149:73-81
Tzeng WN, Shiao JC, Iizuka Y (2002a) Use of otolith Sr: Ca ratios to study the riverine migratory behaviors of Japanese eel Anguila japonica. Mar Ecol Prog Ser 245:213-221
Tzeng WN, Han YS, He JT (2002b) The sex ratios and growth strategies of wild and captive Japanese eels Anguilla japonica. In: Small B, MacKinlay D (eds) Developments in understanding fish growth. International Congress on the Biology of Fish. Vancouver, Canada, Univ. of British Columbia, p 25-42
Tzeng WN, Severin KP, Wickström H, Wang CH (1999) Strontium bands in relation to age marks in otoliths of Eurpean eel Anguilla anguilla. Zool Stud 28:452-457
Tzeng WN, Lin HR, Wang CH, Xu SN (2000) Differences in size and growth rates of male and female migrating Japanese eels in Pearl River, China. J Fish Biol 57:1245-1253
Tzeng WN, Iizuka Y, Shiao JC, Yamada Y, Oka H (2003) Identification and growth rates comparison of divergent migratory contingents of Japanese eel (Anguilla japonica). Aquaculture 216:77–86
Vøllestad LA, Jonsson B, (1986) Life-history characteristics of the European eel Anguilla anguilla in the Imsa River, Norway. Trans Am Fish Soc 115:864-871
Table 1. Mean (± SD) age (yr), TL (mm) and BW (g) of the Japanese eels used for otolith Sr/Ca ratio analysis and collected in the Kaoping River estuary during 1998-2005
Male Female Unknown
Yellow Silver (A) Yellow (B) Silver (C) Yellow (D) Tukey HSD
n 2 39 64 87 29
Age 3.0 ± 0.0 5.8 ± 1.3 4.8 ± 1.3 6.4 ± 1.1 3.8 ± 0.7 D < B < A < C
TL 35.9 ± 0.32 54.5 ± 7.2 45.7 ± 8.8 62.2 ± 7.4 34.7 ± 7.7 D < B < A < C
BW 52.8 ± 2.7 246.8 ± 101.1 146.9 ± 104.7 432.2 ± 191.6 57.1 ± 44.6 D < B < A < C
Abbreviations: (A) Silver male; (B) Yellow female; (C) Silver female; (D) Unknown-sex eel. The yellow males were excluded from analysis due to the low sample size.
Table 2. Composition of life history types (%) of the Japanese eels between sexes Number of eels (%)
Sex n Type 1 Type 2 Type 3
Sex unknown 29 12 (41) 9 (31) 8 (28)
Female yellow 64 23 (36) 24 (37) 17 (27)
silver 87 14 (16) 41 (47) 32 (37)
Male yellow 2 2 (100) 0 (0) 0 (0)
silver 39 21 (54) 15 (38) 3 (8)
Type 1: freshwater resident; Type 2: brackish water resident with freshwater preference; Type3: brackish water resident with seawater preference; number in the parenthesis indicating percentage of each contingent for each eel stage
Table 3. Comparison of mean Sr/Ca ratios (‰) (range) between sexes in otolith regions of 200-400 μm from the primordium and that from elver check to the otolith edge (> 150 μm) in silver Japanese eels
Sex 200-400 μm*
>150 μm
Male 0.26 ± 0.11 (0.10 – 0.55) 0.27 ± 0.09 (0.16 – 0.54) Female 0.34 ± 0.14 (0.16 – 0.59) 0.35 ± 0.08 (0.18 – 0.50) * The region corresponding to the period of sex-differentiation of the eel
Fig. 1. Sampling location for wild Japanese eels in the estuary of the Kaoping River southwestern of Taiwan. The inset indicates the sampling area.
Fig. 2. Temporal changes in the Sr/Ca ratios of Japanese eel otoliths from different migratory contingents collected from the estuary of the Kaoping River. (a) Type 1: freshwater resident; (b) Type 2: brackish water resident with freshwater preference; (c) Type3: brackish water resident with seawater preference. M: metamorphosis from leptocephalus to glass eel; EC: elver check from glass eel to elver. Silver female: (a) 56.0 cm; (b) 72.6 cm; (c) 69.6 cm.
七、計畫成果自評 本研究業已收集足夠之雌雄日本鰻,分析各性別在三型棲地利用之比例,透過統計 驗證鰻魚的性別是否會受環境鹽度所影響。結果顯示日本鰻的性別分化與其棲地利用有 關,亦即生活在半淡鹹水棲地之鰻魚傾向分化為雌性,而生活在淡水棲地之鰻魚則傾向 分化為雄性。此外,是否在不同鹽度的棲地中,鰻魚會有不同的遺傳結構組成,本計劃 仍在進行中,約半年內可分析完成。 本研究結果可應用在放流復育工作上,由於在天然河川中鰻魚性比偏向雌性,因此宜選 擇偏好雄性分化之淡水放流幼鰻,用以補充天然雄鰻之不足。
本研究結果業已彙整撰寫為英文paper,投稿於 Marine Ecology- Progress Series,已獲接