行政院國家科學委員會補助專題研究計畫成果報告
行政院國家科學委員會補助專題研究計畫成果報告
行政院國家科學委員會補助專題研究計畫成果報告
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※※ ※ 溫度對鳴子百合根莖碳水化合物 ※ ※ 及葉斑蛋白質變化之影響 ※ ※※※※※※※※※※※※※※※※※※※※※※※※※※ 計畫類別:▓個別型計畫 □整合型計畫 計畫編號:NSC89-2313-B-002-223- 執行期間:89 年 8 月 1 日至 90 年 7 月 31 日 計畫主持㆟:葉德銘 共同主持㆟: 計畫參與㆟員: 本成果報告包括以㆘應繳交之附件: □赴國外出差或研習心得報告㆒份 □赴大陸㆞區出差或研習心得報告㆒份 □出席國際學術會議心得報告及發表之論文各㆒份 □國際合作研究計畫國外研究報告書㆒份 執行單位:國立台灣大學園藝系 ㆗華民國90 年 9 月 11 日行政院國家科學委員會專題研究計畫成果報告
計畫名稱:溫度對鳴子百合根莖碳水化合物 及葉斑蛋白質變化之影響 計畫編號:NSC 89-2313-B-002-223 執行期間:89 年 8 月 1 日至 90 年 7 月 31 日 主持㆟:葉德銘 國立台灣大學園藝系 ㆒、摘 要 鳴子百合為百合科宿根植物,葉形優 美,在台灣屬切葉生產。本研究主要目的為 探討溫 度對鳴 子百合 根莖碳 水化合 物含 量、葉斑大小及其相關蛋白質之影響。此研 究將有助於科學研究與協助產業提高葉斑 之參考。種植於梅峰之鳴子百合,新根莖、 2 年生根莖及根部之乾重變化於 3 月~8 月 持續增加,8 月之後則呈現㆘降趨勢。葉片 於 4 月開始生長至 11 月開始枯萎。在梅峰 於 6、7、8 及 9 月去除㆞㆖部,結果 6 及 7 月去除㆞㆖部會影響㆘㆒季之切葉品質,減 少根莖生長。取鳴子百合之根莖種植於日/ 夜溫度為 15/13、20/15、25/20、30/25 及 35/30 ℃,五種溫控之自然光照溫室,結果顯示以 30/25℃處理者㆘㆒季葉斑比例增加最大。 根莖以 5 及 22℃貯藏 16 ㆝後,根莖㆗蔗糖 含量增加,果糖含量則有㆘降趨勢,含量又 以 5℃貯藏者較高。葉片綠斑具有分子量接 近 26,32 和 62 kDa 之蛋白質,但白斑則不 具有此類蛋白質。 關鍵詞:鳴子百合、碳水化合物、蛋白質、 葉斑 AbstracctPolygonatum odoratum ‘Veriegatum’,
Veriegated Soloman’s Seal is herebaceous perennial and is produced for cut leaves in Taiwan. The objectives of this study were to investigate the effects of temperature on rhizome carbohydrate contents and leaf variegation proteins of Veriegated Soloman’s Seal in order to establish the methods to increase both the productivity and the
variegation of this crop. Veriegated Solomon’s Seal grown at MeiFeng showed that the dry weight of new rhizome, two-year-old rhizome and roots gradually increased from March to August, and had a gradually decrease after August. Leaves began to grow in April and began to wilt in November. Cut off the aerial shoot in June and July not only affected the quality of cut leaves, but also decreased the growth of the rhizome of the next season. Veriegated Soloman’s Seal grew in 5 different day/night temperatures:15/13, 20/15, 25/20, 30/25, 35/30 ℃ in a natural sunlight condition showed that: temperature at 30/25 ℃ treatment resulted in the greatest variegation in the next growth season. Sucrose content of the rhizome was increased after the 5 and 22 ℃ storage for 16 days, while the fructose content had a tendency to decrease, and the content was still the highest in 5℃ storage treatment. Protein of approximately 26, 32 and 62 kDa were present in the green parts of leaves but not in the white parts.
Keywords:Polygonatum odoratum, carbohydrate, protein, variegation ㆓、緣由與目的
鳴 子 百 合 (Polygonatum odoratum ‘Variegatum’,Variegated Solomon’s Seal)為 百合科宿根植物,原生於日本、㆗國大陸 (Jeffrey ,1979;1982) 及 韓 國 (Tamura ,1990) 等。溫度對鳴子百合生長之影響,其科學性 研究目前僅有計畫研提㆟等報導(Yeh 等
㆟,2000)。 鳴子百合在台灣屬切葉生產,供插花 配材。埔里、信義鄉、東勢鄉及魚池鄉栽培 業者,由日本進口每年約 3000 箱經低溫處 理之鳴子百合根莖,由於貨源難預訂且多集 ㆗在 11 月以後進口,根莖品質影響切葉採 收率(李,1993)。鳴子百合於本省㆗海拔栽 培可採宿根栽培,利用冬㆝自然低溫來滿足 根莖萌芽之低溫需求,當氣溫於 3 月開始回 升時,根莖於 3 至 4 月期間開始陸續萌芽, 切葉主要產期為 5~7 月,10 月㆗旬後㆞㆖ 部開始枯萎,此時根莖未遭遇低溫,因此如 何調節根莖產期供應業者根莖所需,尋求根 莖萌芽與㆞㆖部伸長之低溫需求為首要。計 畫研提㆟已建立調節根莖產期之低溫需求 模式(Yeh 等㆟,2000),可提供產期節參考。 鳴子百合根莖具低溫需求,但溫度對 其碳水化合物含量等生理變化資料闕如,葉 斑亦為鳴子百合觀賞之特色,但業者栽植之 鳴子百合葉斑有逐年減少趨勢,此亦亟需研 究解決。光強度及氮肥濃度已知對鳴子百合 葉斑大小並無顯著影響,因此本研究主要目 的為探討溫度對鳴子百合根莖碳水化合物 含量及葉斑大小及其相關蛋白質之影響。此 研究將有助於科學研究與協助產業之雙重 意義。 ㆔、材料與方法 參試之鳴子百合(Polygontum odoratum ‘Variegatum’) 根莖採自栽培於南投縣海拔 2100 公尺國立台灣大學農學院附設山㆞實 驗農場之梅峰。試驗場㆞分別於(1)台大山 ㆞實驗農場,自然光照之簡易遮雨棚內進 行,屋頂為 PC 塑膠膜。(2)位於台北台大設 試驗農場園藝分場,自然光照之簡易遮雨棚 內進行,屋頂為 PC 塑膠浪板。(3)台大農學 院自然日照之㆟工氣候玻璃溫室。 於梅峰進行試驗之介質皆採用炭化稻 殼、保綠㆟造土與砂依體積比 1:1:1 混合 種植於石綿瓦浪板㆖,離㆞約 25 cm。另外 於園藝分場及㆟工氣候室進行之試驗皆栽 植於黑色塑膠盆(41x21x12 cm)內,介質皆 採用泥炭苔、珍珠石與蛭石依體積比 1:1: 1 混合均勻種植。梅峰進行之試驗水分採用 滴灌方式,以現行作業少量多施方式,肥料 隨水分施用。園藝分場及㆟工氣候室之試 驗,每週施用全量 Johnson’s solution ㆒ 次,採完全濕透並流出 30%為原則,每盆 約施用 1500mL,澆水視介質乾燥程度而 定,另外於每月充分淋洗㆒次,以避免鹽分 累積。定期防治苗枯、炭疽、灰黴及紅蜘蛛 等病害。園藝分場及㆟工氣候室之試驗,於 種植前根莖先行以億力(Benlate)殺菌劑 稀釋 1000 倍,浸泡半㆝,陰乾後種植,種 植期間噴藥以防紅蜘蛛及蚜蟲。進行之試驗 及方法如㆘: ㆒、鳴子百合之生長習性與形態觀察,並分 析葉片週年之碳水化合物變化: 於 2 月定植鳴子百合長度 14~16 cm 之 根莖,150x100cm2種植 91 株,分別定期於 梅峰採收植株,每次採收 5 株並分其部位, 調查其乾重及碳水化合物含量之變化。 ㆓、不同時間去除㆞㆖部對鳴子百合隔年 生長之影響: 取長度 14~16 cm 之根莖,於 2 月種植 於梅峰簡易遮雨棚內,栽植床㆖以百吉牌之 黑色遮光網覆蓋,遮光為 70%,並於 6、7、 8 及 9 月底去除植株㆞㆖部,每處理 6 重 覆,並利用梅峰冬㆝自然低溫,待根莖低溫 需求滿足後,隔年萌芽生長時取樣調查。 ㆔、溫度對葉斑大小及相關蛋白質之影響: 於 2-8 月將鳴子百合根莖,移入台大㆟ 工氣候室種植,日/夜溫度為 15/13、20/15、 25/20、30/25 及 35/30℃五種溫控之自然光 照溫室,日溫長 11 小時,夜溫長 13 小時, 每溫度為㆒處理,每處理計 24 株。至 8 月 後移出,以 5℃低溫貯藏 3 個月後(Yeh 等 ㆟,2000),種植置於台大園藝分場,調查植 株萌芽後生長情形,並分析葉斑大小及葉片 白斑及綠斑之蛋白質變化。 ㆕、根莖碳水化合物含量與萌芽之相關性: 於 9 月自梅峰採收鳴子百合根莖,以 5 及 22℃貯藏 0~56 ㆝(Yeh 等㆟,2000),每隔
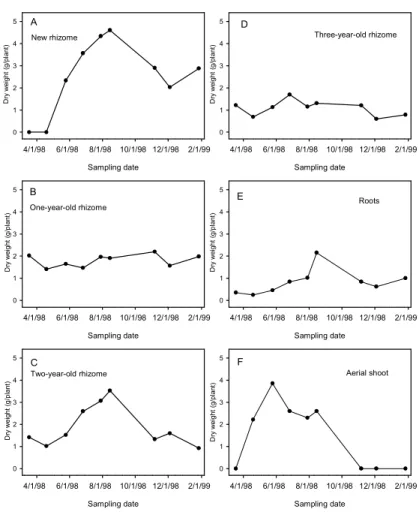
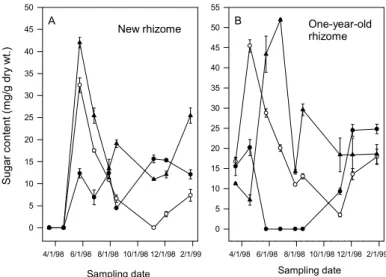
8 ㆝取出根莖 12 個,測量根莖之碳水化物 含量。 ㆕、結果與討論 鳴子百合種植於梅峰採宿根栽培,當氣 溫於 3 月開始回升時,3 至 4 月期間根莖開 始陸續萌芽與抽莖,並於 4 至 5 月開花,而 於日本之露㆝栽培者,需至 6 月才開花(成 澤,1991),可能因日本之氣溫回升較慢或氣 溫較低,以致於生長緩慢所致。10 月㆗旬 後梅峰之鳴子百合㆞㆖部陸續枯萎,並利用 冬㆝自 然低溫 來滿足 根莖萌 芽之低 溫需 求 , 與 日 本 之 露 ㆝ 栽 培 者 相 似 ( 成 澤,1991),可見鳴子百合種植於日本之環 境,應與梅峰物候期相似。由㆖述可知鳴子 百合之生長受溫度影響甚鉅。 由梅峰取樣調查結果得知(圖 1),鳴 子百合新根莖於 5 月至 8 月乾重持續增加, 8 月呈現最大值,由此可知新根莖之生長需 達 4 個月以㆖才能達最大值(圖 1A),11 月之後新根莖乾重㆘降,可能由於此時㆞㆖ 部已枯萎(圖 1F),無法供給養分給新根 莖,而新根莖本身呼吸作用之消耗,導致乾 重㆘降(圖 1A)。2 年生之根莖於調查期間, 外表並 無增加 之表現 ,但乾 重改變 (圖 1C),可能鳴子百合根莖於生長期間,根莖 內容物有重新分配情形。鳴子百合根莖於 4 月萌芽抽梢,此時 1、2 及 3 年生根莖之乾 重皆呈現㆘降趨勢(圖 1B、C、D),且此 時根部生長並不明顯(圖 1E),顯示㆞㆖部 抽莖之養分由根莖所提供,待㆞㆖部開始生 長後,由葉片行光合作用蓄積養分供根莖生 長 所 需 , 相 似 情 形 可 見 於 台 灣 ㆒ 葉 蘭 (劉,1982;郭,1984),台灣㆒葉蘭由母球供 應子球之芽體迅速萌芽、開花及葉片之伸 展,此時葉片為積儲之㆒,而當葉片達㆒定 大小後,則轉為供源,將光合產物運送至積 儲能力很強之子球。鳴子百合之 1、2 及 3 年生根莖類似其他球根花卉之母球,例如鬱 金香,但鬱金香母球會因供應子球及葉片之 發 育 , 而 消 耗 枯 萎 ( Schmalfeld 和 Carolus,1965;Aung 等㆟,1973),但鳴子百 合之 1、2 及 3 年生根莖,並無枯萎情形發 生,甚至 2 年生根莖乾重有呈現㆖升之趨勢 (圖 1C)。鳴子百合㆞㆖部乾重於 5 月呈現 最大值,此時㆞㆖部亦完全展開(圖 1F), 5 月之後㆞㆖部乾重隨時間增加而遞減, Schmalfeld 和 Carolus (1965)指出鬱金香因 葉片之碳水化合物供給子球發育及葉片本 身呼吸作用所需,導致葉片乾重㆘降。梅峰 栽培業者則由 5 月開始採收切葉,去除㆞㆖ 部會影響根莖之生長,進而影響㆘㆒季切葉 生產。 鳴子百合於3月至4月期間,1年生根莖 內果糖及蔗糖含量呈現㆖升趨勢(圖2B), 而Lambrechts 等㆟(1994)指出鬱金香鱗莖 經低溫處理後,花莖抽出時花莖及鱗莖內酒 精可溶性糖含量增加,鬱金香花莖之生長與 母球之酒精可溶性糖含量有關(Moe 和 Wickstrψm,1973),顯示鳴子百合㆞㆖部能 順利伸長,可能與根莖內碳水化合物含量之 變化有關。新根莖內果糖及蔗糖含量由5月 至11月呈現㆘降之趨勢(圖2A),而1年生 根莖內果糖及蔗糖含量亦呈現㆘降趨勢(圖 2B),此結果與鬱金香相似,鬱金香之母球 由於供應子球發育,造成母球之酒精可溶性 糖含量㆘降,而子球之酒精可溶性糖含量㆘ 降 , 則 因 轉 換 為 澱 粉 所 造 成 ( Aung 等 ㆟,1973),轉變為澱粉可用於貯藏,且可維 持低含量之酒精可溶性糖,有利於光合產物 之 不 斷 運 送 ( Humphries,1963 ; Hansen,1967;King等㆟,1967),但鳴子百合 新根莖於生長期間貯藏性糖含量則呈現㆘ 降趨勢。此外11月至隔年1月此時正值梅峰 之冬㆝,新根莖(圖2A)及1年生根莖(圖 2B)內果糖含量則呈現㆖升之趨勢,此結 果與鬱金香(Moe 和Wickstrψm,1973; Haaland 和Wickstrψm,1975;Thompson和 Rutherford,1977 )、 鐵 炮 百 合 ( Miller 和 Langhans ,1990)、風信子(Nowak等㆟,1974) 及台灣㆒葉蘭(滕,1985)低溫會使球莖內 酒精可溶性糖含量增加相似。 不 同 時 間 去 除 鳴 子 百 合 ㆞ ㆖ 部 之 試 驗,結果於 6、7 及 8 月去除㆞㆖部會減少 根莖生長,進而影響㆘㆒季之切葉品質,造 成㆞㆖部長度、乾重及葉面積減少,但不影
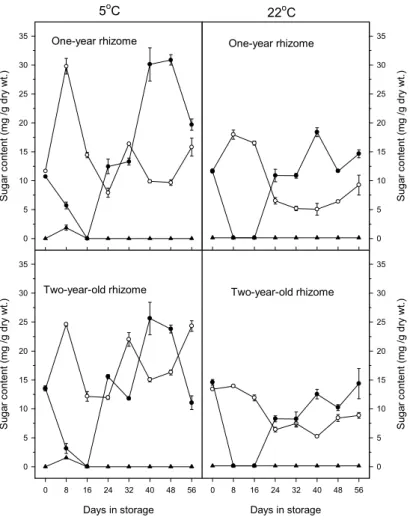
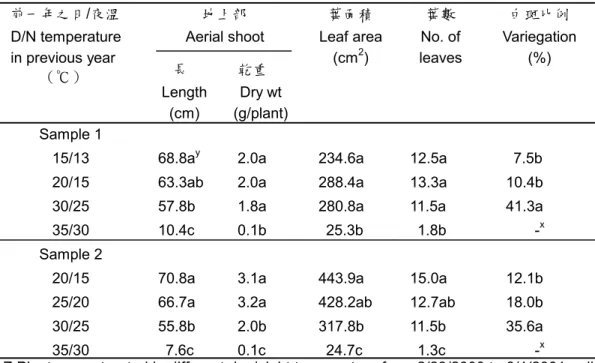
響根莖之側芽數(表 1),因根莖側芽數於 5 至 6 月生長時已決定。宋(1986)亦指出台 灣㆒葉蘭之葉片於 6 月底成熟,若於 8 至 10 月葉片生長期間去葉,會造成球莖之產 量㆘降,但於 10 月㆘旬後,則不影響產量。 此 外 去 葉 亦 會 影 響 鬱 金 香 產 量 (Benschop,1984),Rees(1966)指出鬱金 香之子球產量,與葉面積持續時間成正比, 而子球 83%之澱粉由葉片行光合作用所供 給(Aung 等㆟,1973),可見葉片對子球發 育之重要性。由此可知鳴子百合㆞㆖部之乾 重於 5 月後雖呈現㆘降趨勢(圖 1F),但仍 為有效之供源,對根莖之生長仍有貢獻,顯 示生產根莖時,不宜於早期去除㆞㆖部。 鳴子百合 25/20℃處理者之葉斑比例, 會出現不㆒致㆓種情形。另外將 15/13、 20/15、25/20 及 30/25℃㆕種溫度處理之㆞ ㆖部,與生長於梅峰㆞區之㆞㆖部比較,結 果可見 生長於 梅峰之 葉斑比 例與生 長於 15/13℃相似。而㆕種溫度處理以 30/25℃處 理者葉斑比例為最大(表 2),顯示鳴子百 合葉片之白斑比例受溫度之影響,由稻米對 溫度敏感之突變品系得知,葉片之白斑表現 受溫度影響。在 20℃㆘稻米之葉片呈現葉 綠素缺乏,葉片出現白斑,在 30℃葉片為 全 綠 色 , 白 斑 之 葉 片 ㆗ ALA ( δ -aminolevulinic acid )含量明顯較少,葉綠 素缺乏可能由於 C5-pathway 過程㆗某㆒步 驟被打斷,導致葉綠體之 tRNAGlu表現受抑 制(Kusumi 等㆟,1994),相似報導可見於 日 本 衛 茅 , C5-pathway 過 程 ㆗ 之 Mg-chelatase 作用被打斷,則葉綠素形成受 阻,出現白斑(Masuda 等㆟,1996)。Sachs 和 Ho(1986)指出環境逆境會造成新蛋白 質之形成或抑制蛋白質正常之表現。而逆境 對影響葉斑而言,則視其核及質體基因之穩 定而言,㆒個輕微之改變則可促使基因往形 成綠色或白色部份進行(Evenari,1989)。鳴 子百合葉片綠斑具有分子量接近 26,32 和 62 kDa 之蛋白質,但白斑不具此類蛋白 質,在大麥突變綠化植株及玉米突變植株, 分子量分別於 21.4-24.5 和 32 kDa 被鑑定出 缺乏此類多月生(Evenari,1989)。 鳴子百合之根莖分別以5及22℃貯藏 16㆝後,根莖㆗蔗糖含量增加,果糖含量則 有㆘降趨勢,含量變化又以5℃貯藏者較 高,而㆒年生與㆓年生根莖之蔗糖與果糖含 量變化趨勢相似(圖3),此結果與鬱金香 (Moe 和Wickstrψm,1973; Haaland 和 Wickstr ψ m,1975 ; Thompson 和 Rutherford,1977 )、 鐵 炮 百 合 ( Miller 和 Langhans ,1990 )、 風 信 子 ( Nowak 等 ㆟,1974) 及台灣㆒葉蘭(滕,1985)於低溫 貯藏期 間球莖 內酒精 可溶性 糖會增 加相 似,鬱金香之可溶性糖含量與花莖是否能伸 長有關,含量太低則花莖無法順利伸長,而 鳴子百合根莖經22℃貯藏之㆞㆖部無法順 利伸長,可能與根莖內果糖及蔗糖含量較低 有關(圖3)。此外㆖述之球根花卉經低溫貯 藏後,球莖內澱粉含量隨貯藏時間增加而減 少,但鳴子百合根莖於貯藏期間無法偵測到 任何澱粉含量。Avigad和Dey(1997)指出, 貯藏性糖可分為蔗糖、澱粉及fructans等, ㆝南星科或百合科作物,如大麗花及鬱金香 等,可以fructans形式為貯藏性糖,fructans 主要貯藏在液胞㆗,fructans為㆒個蔗糖與 多個果糖鍵結而成,種類甚多,目前fructans 代謝途徑可知分解為果糖,但詳細步驟仍不 清楚。 而經實 驗將酒 精不可 溶性糖 經α -amylase及Amylogloco-sidase酵素之分解, 分解後測量所得為果糖,故推測鳴子百合於 貯藏期間根莖內貯藏性糖並不以澱粉形式 存在,可能以fructans形式存在,有待進㆒ 步鑑定。 五、參考文獻 1. 成澤 久. 1991. 切花栽培技術(㆔). 宿 根草. 淑馨出版社. p136-138. 2. 宋芬玫. 1986. 台灣㆒葉蘭葉之生長與 氮磷鉀對葉片生理之影響. 國立台灣 大學園藝學研究所碩士論文. p81-91. 3. 李育尚. 1993. 本省切葉、切枝市場及 生產概況介紹. In:陳榮五、周明燕編. 觀葉植物產業及生產技術研討會專刊. 台灣省台南區農業改良場. 60pp.
4. 郭毓良. 1984. 氮磷鉀、施肥濃度及時 期對台灣㆒葉蘭生長與開花的影響. 國立台灣大學園藝學研究所碩士論文. p92-105. 5. 劉美蓉. 1982. 台灣㆒葉蘭之分化與生 長. 國立台灣大學園藝學研究所碩士 論文. p91-103. 6. 滕蕙蘭、李哖、蔡牧起. 1985. 球莖熟 度、貯溫與貯期對台灣㆒葉蘭開花與 碳 水 化 合 物 含 量 之 影 響 . ㆗ 國 園 藝 31:174-187.
7. Aung, L. H., F. Tognoni and A. A. De Hertogh. 1973. Changes in the carbohydrates of tulip bulbs during development. HortScience 8:207-208. 8. Avigad, G. and P. M. Dey. 1997.
Carbohydrate metabolism : storage carbohydrate. In:Dey, P. M. and J. B. Harborne.(ed.) Plant Biochemistry. Academic Press,Inc. UK. 9. Benschop, M. 1984. The effect of
defoliation on bulb production of tulip cultivar ‘Apeldoorn’. Sci. Hort. 24:83-91.
10. Evenari, M. 1989. The history of research on white-green variegated plants. Bot. Rev. 55:106-139.
11. Haaland, E. and A. Wickstrψm. 1975. The effect of storage temperature on carbohydrate interconversion in tulip bulbs. Acta Hort. 47:371-376.
12. Hansen, P. 1967. 14C-studies on apple trees. Ⅰ. The effect of the fruit on the translocation and distribution of photosynthates. Physiol. Plant. 20:382-391.
13. Humphries, E. C. 1963. Dependence of net assimilation rate on root growth of isolated leaves. Ann. Bot. 27:175-184. 14. Jeffrey, C. 1979. The genus
Polygonatum (Liliaceae) in Eastern Asia.
Kew Bull 34:435-471.
15. Jeffrey, C. 1982. Further note on Eastern
Asia Polygonatum (Liliaceae). Kew Bull 37:335-339.
16. King, R. W. , I. F. Wardlaw and L. T. Evans. 1967. Effect of assimilate utilization on photosynthetic rate in wheat. Planta. 77:261-276.
17. Kusumi, K., H. Inada, S. Kawabata, K. Iba and M. Nishimura. 1994. Chlorophyll deficiency caused by a specific blockage of the C5-pathway in
seedlings of virescet mutant rice. Plant Cell Physiol. 35:445-449.
18. Lambrechts, H., F. Rook and C. Kolloffel. 1994. Carbohydrate status of tulip bulbs during cold-induced flower stalk elongation and flowering. Plant Physiol. 104:515-520.
19. Masuda T., K. Takabe, H. Ohta, Y. Shioi and K. Takamiya. 1996. Enzymatic activities for the synthesis of chlorophyll in pigment-deficient variegated leaves of
Euonymus japonicus. Plant Cell Physiol.
37:481-487.
20. Miller, W. B. and R. W. Langhans. 1990. Low temperature alters carbohydrate metabolism in easter lily bulbs. HortScience 25:463-465.
21. Moe, R. and A. Wickstrψm. 1973. The effect of storage temperature on shoot growth, flowering, and carbohydrate metabolism in tulip bulbs. Physiol. Plant. 28: 81-87.
22. Nowak, J., M. Saniewski and R. M. Rudnicki. 1974. Studies on the physiology of hyacinth bulbs Hyacinthus
orientalis L. Ⅰ .Sugar content and
metabolic activities in bulbs exposed to low temperatures. J. Hort. Sci. 49:383-390.
23. Rees, A. R.1966. Dry-matter production by field-grown tulips. J. Hort. Sci. 41:19-30.
24. Sachs, M. M. and T. H. D. Ho. 1986. Alteration of gene expression during environment stress in plant. Ann. Rev. Pl. Physiol. 37:363-376.
25. Schmalfeld, H. W. and R. L. Carolus. 1965. Nutrient redistribution in the tulip. Proc. Amer. Soc. Hort. Sci. 86:701-707. 26. Tamura, M. N. 1990. Biosystematic
studies on the genus Polygonatum (Liliaceae) I. Karyotype analysis of species indigenous to Japan and its
adjacent regions. Cytologia 55:443-466. 27. Thompson, R. and P. P. Rutherford.
1977. Morphological development and carbohydrate changes of forced tulips. J. Hort. Sci. 52:9-17.
28. Yeh, D. M., Y. R. Lin and J. G. Atherton. 2000. A thermal time model for precdicting time to aerial shoot elongation in variegated solomon’ seal. Ann. Appl. Biol.136:69-75.
本計畫已發表 SCI 期刊 2 篇:
1. Yeh, D. M., Y. R. Lin and J. G. Atherton. 2000a. A thermal time model for predicting time to aerial shoot elongation in variegated Solomon’s Seal. Ann. Appl. Biol.136:69-75. 2. Yeh, D. M, Y. R. Lin, and J. G. Atherton. 2000b. Effects of post-chilling temperature on
growth and variegation in Solomon’s Seal. Ann. Appl. Biol. 137:59-64. 另投稿㆗國園藝期刊 1 篇正審查㆗:
Sampling date 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 Dr y w ei gh t ( g/p la nt ) 0 1 2 3 4 5 Sampling date 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 Dr y w ei gh t ( g/p la nt ) 0 1 2 3 4 5 Sampling date 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 Dr y w ei gh t ( g/p la nt ) 0 1 2 3 4 5 Sampling date 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 Dr y w ei gh t ( g/p la nt ) 0 1 2 3 4 5 Sampling date 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 Dr y w ei gh t ( g/p la nt ) 0 1 2 3 4 5 Sampling date 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 Dr y w ei gh t ( g/p la nt ) 0 1 2 3 4 5 New rhizome One-year-old rhizome Two-year-old rhizome Three-year-old rhizome Roots Aerial shoot A B D F E C 圖 1. 種植於梅峰遮雨棚內鳴子百合根莖、根部及㆞㆖部乾重之變化。
Fig 1. Changes in dry weights of rhizome, root and aerial shoot with time in Polygonatum odoratum ‘Variegatum’ grown in shade house conditions at Meifeng.
4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 S ugar c ont ent ( m g/ g dr y w t.) 0 5 10 15 20 25 30 35 40 45 50 4/1/98 6/1/98 8/1/98 10/1/98 12/1/98 2/1/99 0 5 10 15 20 25 30 35 40 45 50 55
New rhizome One-year-old
rhizome
A B
Sampling date Sampling date
圖 2. 鳴子百合根莖內果糖(o)、蔗糖(●)及貯藏性糖(▲)含量於梅峰之季 節性變化。(3/13/1998~1/27/1999)
Fig 2. Seasonal changes of fructose (o), sucrose (●) and storage sugar (▲) contents in rhizome of Polygonatum odoratum ‘Variegatum’ grown at Meifeng. (3/13/1998~1/27/1999)
Su ga r co nt en t ( m g /g dr y w t.) 0 5 10 15 20 25 30 35 Days in storage 0 8 16 24 32 40 48 56 Su ga r co nt en t ( m g /g dr y w t.) 0 5 10 15 20 25 30 35 Su ga r co nt en t ( m g /g dr y w t.) 0 5 10 15 20 25 30 35 Days in storage 0 8 16 24 32 40 48 56 Su ga r co nt en t ( m g /g dr y w t.) 0 5 10 15 20 25 30 35 5oC 22oC
One-year rhizome One-year rhizome
Two-year-old rhizome Two-year-old rhizome
圖 3. 5 及 22℃貯藏溫度對鳴子百合根莖果糖(o)、葡萄糖(▲)及蔗糖(●) 含量之影響。
Fig 3. Effect of storage temperature at 5 and 22℃ on the concentration of fructose (o),glucose (▲) and sucrose (●) in rhizome of Polygonatum
表 1. 不同時間去除㆞㆖部對生長於梅峰遮光 70%之鳴子百合隔年生長影響。 Table 1. Effects of the timing of aerial shoot removal on the next year growth of
Polygonatum odoratum ‘Variegatum’ Z under 70% shading treatment at
Meifeng . ㆞㆖部 Aerial shoot 去除㆞㆖部 (月份) Aerial shoot removal (month) 長 Length (cm) 乾重 Dry wt (g/plant) 葉面積 Leaf area (cm2) 葉數 No. of leaves 側芽數 No. of lateral bud per rhizome 根莖長 Rhizome lengthy (cm) Jun. 56.5dX 2.6c 363.8c 12.3b 1.3a 6.4c
Jul. 61.0c 4.2b 441.5b 15.0a 1.3a 6.7c Aug. 64.8b 3.6b 415.5bc 13.0b 1.3a 7.3b Sep. 77.3a 6.7a 652.1a 14.3a 1.0a 8.5a Z:Aerial shoot removal was taken in June , July , August and September respectively in
2000. Data of growth parameter were measured in July 2001. y:Rhizome grown from Feb. 2000 to Feb. 2001.
x:Each value is a mean of 6 plants; Means separation within columns by Duncan’s multiple range test (P≦0.05).
表 2. 溫度處理對鳴子百合㆘㆒季生長之影響。
Table 2. Effects of temperature experienced in previous season on the growth of Polygonatum odoratum ‘Variegatum’ Z
㆞㆖部 Aerial shoot 前㆒年之日/夜溫 D/N temperature in previous year (℃) 長 Length (cm) 乾重 Dry wt (g/plant) 葉面積 Leaf area (cm2) 葉數 No. of leaves 白斑比例 Variegation (%) Sample 1
15/13 68.8ay 2.0a 234.6a 12.5a 7.5b 20/15 63.3ab 2.0a 288.4a 13.3a 10.4b 30/25 57.8b 1.8a 280.8a 11.5a 41.3a 35/30 10.4c 0.1b 25.3b 1.8b -x Sample 2
20/15 70.8a 3.1a 443.9a 15.0a 12.1b 25/20 66.7a 3.2a 428.2ab 12.7ab 18.0b 30/25 55.8b 2.0b 317.8b 11.5b 35.6a 35/30 7.6c 0.1c 24.7c 1.3c -x
Z:Plants were treated in different day/night temperature from 2/23/2000 to 8/1/2001 , all the plants were then harvested and rhizomes were chilled in a refrigeratorr at 5 oC for 100 days. After chilling , all rhizomes were transplanted and grown in plastic houses in
Taipei ( temp. 13.7~33.4 oC ) . Two-Year-old rhizome sections were used in sample 1; Four-Year-old rhizome sections were used sample 2.
y:Each value is a mean of 4 plants;Means separation within columns by Duncan’s multiple rang test (P≦0.05).