國
立
交
通
大
學
生物科技系
碩
士
論
文
探討干擾素-γ及內毒素刺激誘導人類介白素 12 p40 (IL-12 p40)
基因轉錄作用分子機轉之研究
Studies of molecular mechanism of interferon-g and endotoxin induced upregulation of human interleukin-12 p40 (IL-12 p40) gene
研 究 生:吳欣怡
指導教授:袁俊傑 博士
探討干擾素-γ及內毒素刺激誘導人類介白素 12 p40 (IL-12 p40)基因轉錄作用分子 機轉之研究
Studies of molecular mechanism of interferon-γ and endotoxin induced upregulation of human interleukin-12 p40 (IL-12 p40) gene
研 究 生:吳欣怡 Student:Hsin-Yi Wu 指導教授:袁俊傑 Advisor:Chiun-Jye Yuan 國 立 交 通 大 學 生物科技系 碩 士 論 文 A Thesis
Submitted to Department of Biological Science and Technology National Chiao Tung University
in partial Fulfillment of the Requirements for the Degree of
Master in
Biological Science and Technology July 2004
Hsinchu, Taiwan, Republic of China
探討干擾素-γ及內毒素刺激誘導人類介白素 12 p40 (IL-12 p40) 基因轉錄作用分子機轉之研究 學生:吳欣怡 指導教授:袁俊傑 國立交通大學生物科技系(研究所)碩士班 摘要 介白素-12 ( Interleukin-12 )為抗原呈現細胞受到病原菌刺激後產生之細胞激素之 一。主要之功能為活化自然殺手細胞及促使輔助型T 細胞成熟。介白素 12 由 p35 及 p40 兩個次單元以雙硫鍵連接構成。多數細胞皆可表現p35 次單元,而 p40 只在活化後的抗 原呈現細胞中才大量表現。 我們選殖人類介白素 12 p40 啟動子區間-922 至+35,此區間之序列經由軟體比對發 現除了目前已知之轉錄因子辨識位置之外,仍有一些與免疫相關之轉錄因子辨識區域可 能存在。我們將啟動子區域序列自5’端選擇性截斷消去,再將這些不同長度之啟動子區 域序列置於蟲螢光酵素基因之前,來建構一系列的啟動區截斷型報導質體,以電穿孔方 法將這些質體分別送入細胞中。在本研究中我們成功建立了穩定持有不同長度之人類介 白素12 p40 啟動子區域蟲螢光酵素報導基因的小鼠巨噬細胞株 (RAW264.7),以此做為 探討LPS/IFN-γ以及中草藥初萃物分子機制之工具。我們發現 LPS/IFN-γ同時處理細胞可 以協同性地激活啟動子區間-563 至-398 及 Ets-2 辨識區間(-222 至-212)。後者為相關研 究中常被提及之部分。經由軟體比對發現-433至-413區間為 c-Rel 辨識區,以-433至-413 之DNA 序列為探針進行膠體位移實驗,結果顯示確實有蛋白質會結合至此區間。此外, 我們也利用現有之報導質體來篩選具有活化介白素12 p40 之中草藥初萃物並找出可能 作用之啟動子區間。我們發現部分中草藥之萃取物確實能有效促進巨噬細胞株之介白素 12 p40 轉錄作用之進行。往後,我們將進一步分離出具有生物活性之前導化合物,並深 入探討中草藥促進巨噬細胞介白素12 p40 作用之分子機轉。
Studies of molecular mechanism of interferon-γ and endotoxin induced upregulation of human interleukin-12 p40 (IL-12 p40) gene
Student:Hsin-Yi Wu Advisor:Dr. Chiun-Jye Yuan Department of Biological Science and Technology
National Chiao Tung University
Abstract
Interleukin-12 (IL-12) is a cytokine secreted by antigen-presenting cells (APCs) in response to pathogen infections. The major functions of IL-12 are to activate natural killer cells and promote naïve T helper cells to functional type 1 T helper cells. IL-12 is a heterodimeric cytokine composing of two distinct subunits, p35 and p40 linked by disulfide bonds. The p35 is constitutively expressing in most cells, whereas p40 is inducible and can only be detected in activated APCs.
The promoter region -922 to +35 of human IL-12 p40 gene containing several putative transcription factor binding elements has been subcloned and their sequence alignment confirmed. A set of luciferase reporter plasmids containing various length of IL-12 p40 promoter were constructed and transfected into RAW264.7 cells to generate compounding stable cell clones. The stable clones of RAW264.7 containing human IL-12 p40 promoter−based reporter vectors were used to study the molecular mechanisms underlying LPS/IFN-γ− or Chinese herbs− induced IL-12 p40 gene activation. We found that promoter regions -563 to -398 and -239 to -190 are important for the synergistic effect of LPS/IFN-γ. The region -239 to -190 contains a Ets-2 (-222 to -212) site, which is consistent with previous finding. The result from gel shift assay, two bands were seen using region -433 to -413 as probe. Upon sequence alignment, a putative c-Rel binding element was postulated in region -563 to -398. Furthermore, the reporter plasmids pIL12p40/90 containing RAW cells were
used to screen for the bioactive components of various Chinese Herbs. The result implicated that our system is useful for the future drugs screening and molecular mechanism studies of Chinese herbs.
Acknowledgment
本論文感謝指導教授 袁俊傑老師在這兩年內不倦的指導與教誨,以及 口試委員 徐祖安博士、廖光文博士和吳文勉博士於口試時給予的寶貴意 見,除了使本論文更趨完善,也讓我在未來的研究方向上有更嶄新的思維。 兩年時光匆匆流逝,大家對我的幫助成就了我的成長。感謝實驗室學長 姐 俊炫、君璇、柏文、柏翰,和我共進退的同學 珮玲、昱珊、政剛、宗 翰,以及學弟妹 宜芳、元碩,你們的陪伴使我的研究生涯更多采多姿。感 謝我的家人 爸爸、媽媽、眾姊妹們以及怡仁給我無限的關懷與支持。Content
中文提要 ……… i Abstract ……… ii Acknowledgment ……… iv Content ……… v Content of tables ……… viContent of figures ……… vii
Abbreviation Index ……… ix
I. Introduction……… 1
II. Methods and Materials……… 5
III. Result……… 16
3.1 Construction of reporter plasmids of IL-12 p40 gene…… 16
3.2 Deletion analysis of the promoter of IL-12 p40 gene 18 3.3 Generation of IL12p40/90 containing RAW264.7 stable clones ……… 22 IV. Discussion……… 27 References ……… 30 Appendix 1 ……… 71 Appendix 2 ……… 72 Appendix 3 ……… 73 Appendix 4 ……… 74
Content of Tables
Table 1 Primers in the work……..……….…37 Table 2 Plasmids and vectors use in the work……….……… .39 Table 3 Classification of Chinese Herbs in this thesis……….….41 Table 4 List of responding element in the human and murine IL-12 p40 promoter……42 Table 5 The putative transcription responding elements in the promoter region from -922 to
+33 of human IL-12 p40.………..….44 Table 6 The activation of IL-12p40 promoter induced by four fractionated layer of Chinese
Content of Figures
Figure 1 Comparison of the promoter region of murine and human IL-12 p40 gene……….47
Figure 2 Potential responding elements in 5 –flanking region of human IL-12p40 gene...48
Figure 3 Construction of pIL12p40/90 Teasy……….49
Figure 4 Construction of pIL12p40/90 Lucneo……… 50
Figure 5 pIL12p40/90 Lucneo vector circle map……….51
Figure 6 Illustration of the luciferase reporter plasnids containing various length of the promoter region of human IL-12 p40 gene.………...52
Figure 7 Time-dependent activation of IL12p40/90 promoter…………53
Figure 8 Western blotting of NPT in different RAW264.7 stable clones………….54
Figure 9 LPS and LPS plus IFN-γ induce IL-12 p40 promoter fragments activation.…...55
Figure 10 Time course of the RAW(IL12p40/56) activation by LPS, IFN-γ or both...56
Figure 11 Time course of the RAW(IL12p40/25) activation by LPS and LPS/IFN-γ...57
Figure 12 Does dependent effect of LPS or IFN-γ on the induction of IL12p40/56 dependent luciferase activity……….………58
Figure 13 Illustration of reconstituted reporter plasmids pIL12p40/56-40……...59
Figure 14 Time-dependent effect of LPS, IFN-γ or both on the luciferase activity of the RAW(IL12p40/56) cells………...………….……..60
Figure 15 Demonstration Ets and c-Rel transcription factor binding elements of human IL-12 p40 promoter……….…………..….…….…61
Figure 16 Illustration of reporter vectors containing mutations site of -69 to +33 promoter region………..……….………..………..…...62
Figure 17 Effect of LPS, IFN-γ on the promoter activity of pIL12p40/7, pIL12p40/7∆I and pIL12p40/7∆N…....……….……….……..63
Figure 19 Screen for the effective stable clones of RAW(IL12p40/90)...65
Figure 20 Chinese herb extract 5-4 …………..……….…….…66
Figure 21 Chinese herb extract 8-4 …………..………..….…67
Figure 22 Chinese herb extrac 15-2 ……….…68
Figure 23 Chinese herb extract 16-3. ……….….…69
Abbreviation Index
LPS Lipopolysaccharide IFN-γ Interferon-γ
IL-12 Interleukin-12
TF Transcription factor
NF-AT Nuclear factor of activated T cell NF-κB Nuclear factor kappa B
ICSBP Interferon regulated concensus sequence binding protein
IRF Interferon regulatory receptor C/EBP CCAAT-enhancer binding protein APC Antigen presenting cell
TLR Toll like receptor
EMSA Electrophoretic mobility shift assay
I. Introduction
Interleukin-12 (IL-12) is a key cytokine that involved in the development of innate and acquired immunities. It is a potent stimulus for the production of interferon-γ (IFN-γ) in normal lymphocyte, resting and activated T cells and in NK cells (natural killer cells) [1]. The evolvement of cell-mediated immunity is promoted by IL-12. IL-12 has been shown to play roles in activating cytotoxic T cells and NK cells, upregulating the expression of surface receptors of helper T (Th) cells and NK cells, and stimulating IFN-γ secretion from T lymphocytes (Appendix 1). The early decision toward Th1 or Th2 cells is depends on the balance between IL-12, which favors Th1 response, and IL-4, which favors Th2 response [2].
In preclinical studies, IL-12 augments the cytotoxic activity of lymphocytes and the antitumor activity of NK cells from patients with tumors can be augmented the treatment of IL-12 [3-6]. IL-12 is used clinically to cure some inflammatory diseases, such as airway hyper-responsiveness, asthma and allergy [8-10]. However, IL-12 may involve in the pathological process of some autoimmune diseases. The peripheral blood mononuclear cells (PBMCs) from patients with progressive multiple sclerosis were shown to produce more IL-12 [11]. NOD (nonobese diabetic) mice, like type I diabetic patients, develop insulitis, an early infiltration of leukocytes into the pancreas that leads to inflammatory lesions within the islets [12 and 13]. The macrophage depleted NOD mice appear normal without the characteristics of type 1 diabetes, such as death of β cell and the type 1 response of the auto-reactive T-helper cells. After injecting IL-12, the symptom of diabetes mellitus appeared in the macrophage-depleted NOD mice, suggesting that pathogenic effect of T-cell in type I diabetic patients might be mediated by IL-12 [14].
Active IL-12 is a heterdimeric complex consisting of two subunits, p40 and p35. Recent studies demonstrate that p40 can associate with either p35 to form
the active IL-12 p70 heterodimer, with p19 to form the active IL-23 p59 heterodimer or with itself to form an inhibitory p80 homodimer. The IL-12 p70 can be released by antigen- presenting cells (APCs), such as dendritic cell, macrophage, monocyte, and activated B cell by the stimulation of cytokines, pathogens, nutrients and stresses [15]. The p35 subunit of IL-12 is thought to be expressed constitutively, and regulated post-transcriptionally in most cell types [16 and 17]. Whereas, p40 subunit of IL-12 is inducible upon the stimulation of various stimuli.
Although genes can be regulated through several levels, transcription is the primary control for the gene expression [18]. Upon stimulation, the transcription of IL-12 p40 gene can be upregulated by transcription factors, such as Ets-2, ICSBP (IFN consensus sequence binding protein, also called IRF-8), NF-κB and IRF-1 (interferon regulatory factor-1) (Table 4).
Lipopolysaccharide (LPS), derived from the cell wall of Gram negative bacteria, is a potential activator of macrophages. It has been shown that LPS can induce the expression of IL-12 p40 gene in murine peritoneal macrophages by binding to receptor complexes, consisting of lipopolysaccharide-binding protein (LBP), CD14, Toll like receptor-4 (TLR4) and the accessory protein MD-2 [19 and 20]. Upon the binding its receptor stimulates the activity of mitogenic-activated protein kinase (MAPK) cascades (Appendix 2) [21]. These MAPKs activate different transcription factors, including Elk-1, AP-1, and CREB by phosphorylation. LPS may also activate IKKβ by serially activating MyD88, IRAK,-TRAF-6, and TAK1/MEKK1. Subsequently, IκBs is phosphorylated by IKKβ and targeted for degradation [22 and 23]. The degradation of IκBs permits NF-κB/Rel complex, e.g. p65/RelA, to translocate to nuclear [20]. NF-κB is a primary transcription factor that binds to the DNA sequence between -118 and -107 of the human IL-12 p40 gene [24] or between -132 and -122 of the murine IL-12 p40 gene [25]. Further studies found that
two other sites, Ets and C/EBP residing upstream (-211 to -205) and downstream (-75 to -64) of Rel/NF-κB site, respectively, are important for the trans-activation of Rel/NF-kB [15, 25 and 26]. Interestingly, Ets and C/EBP are conserved in both human and mouse genes.
Interferon-γ (IFN-γ), a cytokine mainly produced by T and NK cells, regulates the inflammatory and the antimicrobial/tumoricidal activity of macrophages by binding to its specific receptor [27]. The IFN-γ receptor, in turn, enhances the production of mediators, such as TNF-α, IL-1, NO, and IL-12, from macrophage [28]. The IFN-γ receptor consists of α and β chains, forming a heterodimer upon the binding of IFN-γ. Dimerization triggers receptor trans- phosphorylation and activates the associated JAK1 (Janus kinase) and JAK2 tyrosine kinases [29 and 30]. Subsequently, the activated JAK1 and JAK2 phosphorylate and activate downstream γ-activated factor (GAF), consisting of STAT1 (signal transduction ans activator of transcription) α dimmer and possibly another DNA binding protein. The activated GAF then upregulates transcription of IFN-γ-responsive genes by binding to their GAS (γ-activated site) [31-34]. IRF-1 and ICSBP, two GAF-activated proteins, have been shown to modulate important cellular events in response to IFN-γ [31 and 35]. ICSBP and IRF-1 can form a complex and binds to the Ets site in the promoter region from -224 to -214 of human IL-12 p40 gene [35]. In murine IL-12 p40 gene, ICSBP can associate with NF-AT (nuclear factor of activated T cell) and binds to the NF-AT site (-68 to -63) [17]. On the other hand, IRF-1 was found to activate the transcription by binding at site (-63 to -56) downstream the Rel/NF-κB. The binding of IRF-1 at this site could either up or down regulate the transcription of IL-12 p40 in response to different stimuli [31]. Mice with deficient of the IRF-1, IRF-2, or ICSBP exhibit a defect in IL-12 production [17, 36 and 37], suggesting the essential role of these transcription factors in the expression of IL-12. A synergistic
effect was observed on promoting the human IL-12 p40 gene in RAW264.7 cells with pretreatment of IFN-γ 8 h prior to LPS stimulation [2]. The promoter region responsible for the synergistic response of LPS and IFN-γ was located within -222 and -212 relative to the transcription start site of human IL-12 p40 gene [2]. These results suggest that multiple transcription factors probably work cooperatively to enhance the transcription of IL-12 p40 gene. However, the cooperation between LPS and IFN-γ signal pathway is still unknown. Earlier works have shown that most known functional transcription binding site were located in the region between -250 and -50 of human IL-12 p40 gene (Fig. 1 and Table 4). Since the total length of reported promoter region of p40 gene is about 2500 basepairs, the promoter region upstream of -250 remain to be explored.
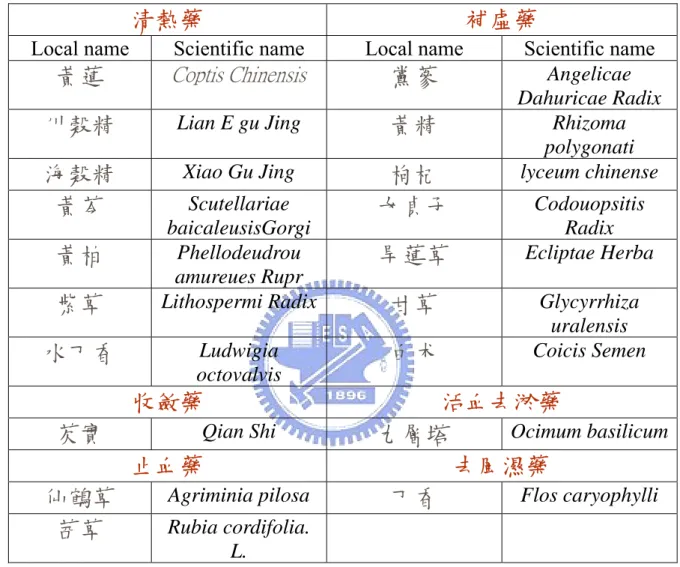
Owning to the importance of IL-12 on regulating cellular immune response and acting as bridge between innate and adaptive immunities, we have constructed a cell-based assay system for the screening of functional lead compounds from Chinese herbs that can modulate the transcription of IL-12 p40 gene. Various traditional Chinese herbs (Table 3) were chosen to screen for the potential lead components.
II. Materials and Methods
I. Materials
1. Cell Line
Murine macrophage cell line RAW264.7 and human embryonic kidney cell line HEK 293 cell lines were purchased from Food Industrial Research and Development Institute, Hsinchu.
2. Reagents
The crude extracts of various Chinese herb including coptis chinensis,
Lian E Gu Jing, and Xiao Gu Jing, were provides by Ho, Jiau-Ching of Ta-Hwa
Institute of Technology. Lipopolysaccharide (LPS) of Salmonella typhimurium and Escherichia coli G418, inhibitors or antagonists were from Calbiochem. Restriction enzymes were from New England BioLabs. Prestained protein standard marker (broad range) and Bradford’s reagent were obtained from BioRad. BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit was purchased from ABI. Biomax film was from Kodak. Hyperfilm, PVDF and ECL were from Amersham. Nylon membrance was from Schleicher &Schaell. The pfuTurbo DNA polymerase was from STRATAGENE. Taq DNA polymerase was from Viogene. Agarose, gel DNA extraction kit and BSA were from Boehringer Mannheim. T4-DNA ligase was from MBI. Fetal bovine serum (FBS) and Dulbecco’s modified egale medium (DMEM) were purchased from Hyclone. Penicillin, streptomycin, fetal calf serum (FCS), agarose, 0.25% trypsin-EDTA and ethidium bromide (EtBr) were purchased from GIBCO BRL. 3’end Biotin labeling kit, electropherotic mobility shift assay (EMSA) kit and Nonidat P-40 (NP-40) was purchased from PIERCE. All other reagents and chemicals used in the experiments were reagent grade.
3. Primers
All the primers used in the thesis are listed in Table. 1.
4. Plasmids and Vector
II. Methods
1. Cell Culture
Human embryonic kidney cell line HEK293 cell was grown in DMEM medium containing 10% FBS, 100 µg/mL penicillin, and 100 U/mL streptomycin in a 37℃, humidified incubation chamber in the presence of 95% air and 5% CO2.
Cells were subcultured in an interval of 2-3 days by detaching cells with 0.05% trypsin-EDTA and replating in the ratio of 1:2 to 1:4. Murine macrophage cell line RAW264.7 was grown in DMEM medium containing 10% FBS, 100 µg/mL penicillin and 100 U/mL streptomycin in a 37℃, humidified incubation chamber containing 95% air and 5% CO2. Cells were subcultured every 2-3 days.
Cells were from cultural flask detached using gently patting, and subcultivated in the ratio of 1:3 to 1:6.
2. Preparation of Genomic DNA
HEK 293 cells (2×107 cells) were harvested and resuspended in cold PBS (pH 7.4). Cells were washed once with PBS and centrifuged at 3,000×g for 2 min to pellet cells. Following the removal of the supernatant, 400 µL lysis buffer (200 mM NaCl, 20 mM EDTA, 40 mM Tris-HCl (pH 8.0), 0.2% triton X-100, 5 µM DTT, 100mg/mL lysozyme and 20 µg/mL of protease K) were added and resulting mixture was incubated at 50℃ for overnight. Place the digesting mixture on ice after incubation, followed by adding 250 µL saturated NaCl and incubating at 0 ℃ for 10 min. Centrifuge the mixture at 6,000×g for 10 min, and transfer the supernatant to a new 1.5mL eppendorf tube for DNA precipitation. The precipitation of genomic DNA was performed by adding two volumes of 95% ethanol (about 0.4 mL) into the tube, mixed well, and incubated at -20℃ for 2 hr. Decant the upper layer and wash the genomic DNA pellet twice with 1mL cold 75% ethanol. Remove the residue solution and air
dry the DNA pellet by placing tube at room temperature with cup open for 3-5 min. Dissolve genomic DNA in 100µL TE buffer, pH 8.0.
3. Plasmids construction
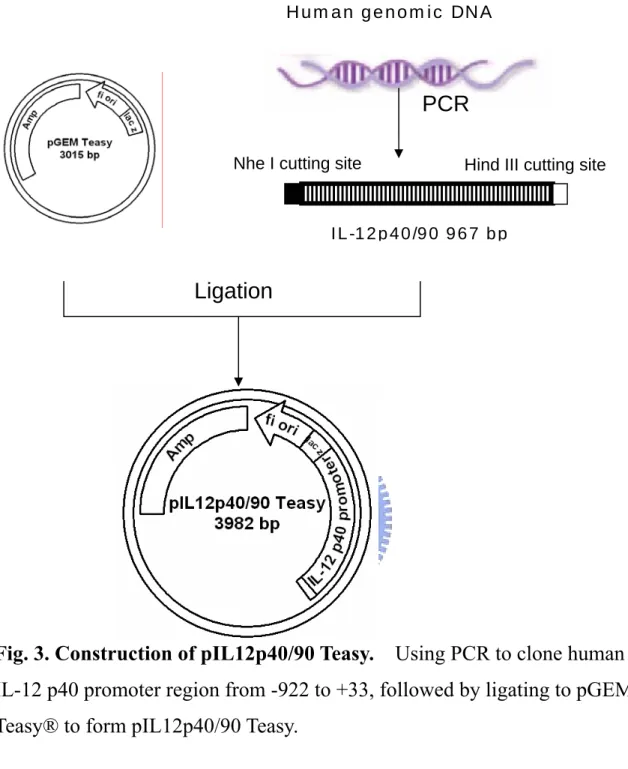
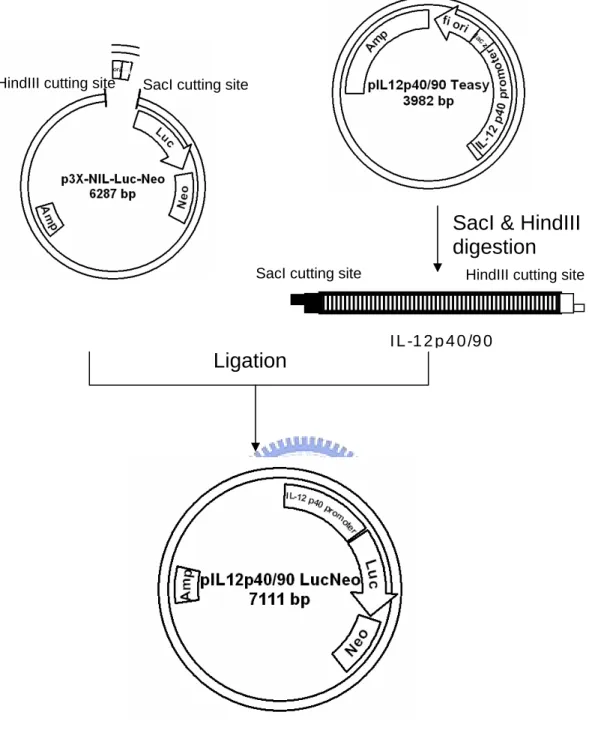
To generate the human IL-12 p40 promoter/luciferase reporter constructs, we subcloned promoter region (-922 to +35) of IL-12 p40 ( +1 is starting site of transcription ) by PCR by using genomic DNA isolated from HEK293 as template. After purification, the corresponding DNA fragments were ligated into pEasyTTM vector to form pIL12p40/90 Teasy (Appendix 3 and Fig. 3). The DNA sequence of the subcloned promoter of human IL-12 p40 was confirmed by ABI377 DNA sequencerTM. The serially deleted gene promoter fragments (Fig. 6) of IL-12 p40 were generated by PCR and ligated into pEasy TTM vector. The DNA sequence of the subcloned promoter fragments was confirmed by ABI377 DNA sequencerTM. The promoter fragments were excised from pEasy TTM vectors using SacI/HindIII enzyme and inserted into the same sites of of p3X-NIL-Lucneo [52] after removing 3X-NF-IL6 responding
element fragments (Fig. 4 and 5). The resulting reporter vectors for IL-12 p40 promoter activity are illustrated in Fig 3. pIL12p40/5 Leuneo means that the
reporter plasmid was inserting from -48 to +33 and containing TATA box only. pIL12p40/0 Leuneo means that the reporter plasmid was inserting no promoter
region. In order to removal the promoter inserting, we excised and religated in NotI/HindIII region from pCDNA3.0 to p3X-NIL-Lucneo.
4. Preparation of reporter plasmids stably transfected cell lines
To generate stably transfected cell lines, the reporter plasmids were electroporated into RAW264.7 cells by using Multiporator (Eppendorf, AG) following manufacture’s instruction. RAW264.7 cells were harvested and pellet by centrifugation at 3000rpm for 5 min under the room temperature.
Wash cells once with DMEM containing 0.5% FCS and calculate the cell density. Resuspend cells by Hypoosmolar Electroporation Buffer (Eppendorf, AG) in a concentration of 5 × 106 cells/mL. Add plasmid DNA to the cells in a final concentration of 25 µg/mL. Transfer cell and plasmid mixture (800 µL) into electroporation cuvettes (4 mm gap width) and carefully avoid the formation of air bubbles. Electroporation was performed under the condition indicated as following : The mode was set at Eukaryotes “µ”;Voltag was at 570
V; “the time constant (µ)” and “No. of pulses” were adjusted to 50 µs and 2,
respectively. After the electric shock, the cells were allowed to stand in the cuvette for 5-10 minutes at room temperature. Carefully transfer the cells suspension from the cuvette to T-25 containing 5mL DMEM containing 10% FCS. The cells were then recovered in a humidified 37 ℃ incubator for about 24 h.
Replacing the conventional cultural medium with selection medium (DMEM containing 10% FBS, 100 µg/mL penicillin and 100 U/mL streptomycin, and 500 µg/mL G418) and cultivated for another 3 days. Some untransfected cells should be eliminated under this constrained selective culturing condition. Harvest the remaining cells and determine the concentration of cells, and replate the cells in the T25 flask. The cells were further grown in the medium containing 750 µg/mL G418 for another 3 days. The selective stress was further constrained by increasing G418 from 750 µg/mL to 1000 ng/mL during the final stage of selection. Replacing selection medium every 2-3 days until the resistant clones cover the surface of cultural well. Transfer the neomycin resistant clones to a new 96-well cultural plate in a cell concentration of 1 cell/well for further amplifying the cell number. The cell number B21, A1 and B30 of neomycin-resistant cell clones were gradually amplified from 12-well plate, a 6-well plate, a T-25 and to a T-75 flask. The stably transfected cell clones were then harvested and transfered to liquid N2
tank for long term storage.
5. Luciferase Assay
Plating various stably transfected cell clones in the 24-well plates in a concentration of 2.5 × 105 cells/well. Following incubation at 37 ℃ for 24 h, cells were treated with cultural medium containing LPS, IFN-γ, drugs, herb extracts, inhibitors or medicines at the indicated concentration. Treatment cells were harvested at the indicated times. Briefly, following removal cultural medium, cells were washed once with 1X PBS, pH7.4, followed by adding 50 µL cell lysis buffer (25 mM Tris-HCl, pH 7.8, 2 mM DTT, 2 mM EDTA, 10% glycerol, 1% Triton X-100). The resulting cell lysates were stored in the -80 ℃ freezer.
The luciferase activity of cell lysate was performed by mixing 50 µL luciferase assay reagent (0.47 mM D-luciferin, 2.67 mM MgSO4, 20 mM Tricine,
0.55 mM ATP, 1.07 mM (MgCO3)4-Mg(OH)2.5H2O, 0.27 mM CoA, 33.3 mM
DTT, 0.1 mM EDTA) with 10 µL cell lysate in a 100×12 mm plastics tube. The activity of luciferase was determined on the luminometer (BioOrbit) with a detection protocol: Delay 90 seconds, detect 20 seconds and integrate the relative light units (RLU) detected within 20 seconds. The luciferase activity of each set of samples is normalized using the level constitutively expressed neomycin phospho-transferase (NPT) in each sample.
The protein concentration of cell lysate was determined using Bradford reagent (Bio-Rad) under constant reaction volume of 200 µL containing 4 volumes of didiluted water solution to one volume of Bradford’s reagent. Following vigously vortexing, the mixture incubated in room temperature for 10 ~20 min, determine the absorbance (OD595) at wavelength in 595 nm by
The pIL12p40/90, 56, 40, 25, 20, 10, 7, 5, 0 Lucneo stably transfected
RAW264.7 stable clones (RAW(IL12p40/90), RAW(IL12p40/56), RAW(IL12p40/40), RAW(IL12p40/25), RAW(IL12p40/20), RAW(IL12p40/10), RAW(IL12p40/7), RAW(IL12p40/5), RAW(IL12p40/0)) were generated and maintained in the culture medium containing 250 µg/mL G-418 sulfate. The reporter plasmid stably transfected RAW264.7 clones were further tested for their responses to LPS.
(4)Data analysis
After the process of luciferase assay and protein assay, we could obtain the value of the RLU and concentration of cell lysate. Determine the value of RLU divided by the amount of protein in 10 µL lysate. This value (RLU/µg. of prot) indicates the luciferase expressional level and trans-activation activity of the transcription factors in IL-12 p40 promoter region.
Neomycin phosphor-transferase is expression constitutively. In order to normalize copy numbers of different length of promoter reporter plasmids, we check the expression amounts of neomycin phospho-transferase of different stable clones by western blotting (see western blotting). Each sample loads 60 µg of protein amount. The NPT protein and β-actin expression amounts of RAW(IL12p40/90) are used as 100% and others are compared with RAW (IL12p40/90). Thereafter, we use NPT relative percentage to divide β-actin relative percentage to get the NPT/β-actin ratio, which is used as Normalization Index and each RLU/µg. of protein is further divided by Normalization Index.
6. Western Blotting
RAW264.7 cells were grown in a 100 mm dish to about 60-70% confluence before being treated with medium, drugs, stimuli or Chinese herb extracts. After incubation for the indicated time internals, the cells were harvested and
incubated with cell lysis buffer (20 mM PIPES, pH 7.2, 100 mM NaCl, 1 mM EDTA, 0.1% CHAPS, 10% sucrose) at 4 ℃ for 15 min. The total protein of each cell sample was loaded onto a 10% SDS polyacrylamide electrophoresis gel (SDS-PAGE). After electrophoresis at 150 V for 2 hr, the separated protein bands were transferred to a polyvinylidine difluroide (PVDF) membrane through a Mini-Trans Blot Electrophoretic Transfer Cell soak in the presence of Transfer buffer (12.5 nM Tris, 96 mM glycine, 20% methanol, pH8.9). After transblotting, PVDF membrane was blocked with 5% skim milk in PBST (1X PBS with 0.5% Tween 20) at room temperature for 1 h or 4 ℃ for 8~16 h. The membrane is then incubated with first antibody in PBST with 5% skim milk at room temperature for 1 h or 4 ℃ for 8~16 h. After incubation, the membrane was washed three times with PBST, followed by incubating with horseradish peroxidase conjugated secondary antibody at room temperature for 1 hr. After washing three times with PBST, 1 mL ECL (Amersham) was lay on the membrane for chemiluminescent reaction. The specific protein was visualized by exposing to an x-ray film (Hyperfilm). Protein concentration for SDS-PAGE was determined by Bradford’s assay (Bio-Rad). Stripping buffer we used is containing 65 mM Tris-HCl (pH 6.8), 2% SDS, and 100 mM β-mercaptol.
7. Subcellular Fractionation
After treatment, cells were washed twice with cold 1X PBS, pH 7.4, and then suspended in 100 µL buffer I (10 mM Tris-HCl, pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM DTT, 0.5 mM PMSF, and protease inhibitor cocktail) and kept on ice for 15 minutes. After incubation, 25 µL 10% Nonidet P-40 was added followed by a vigorously vortexing for 10 seconds. This procedure will be used to help weakening the cell plasma membrane. The resulting cell lysate was then centrifuged at 14000 rpm for 15 minute at 4 ℃.
The supernatant containing cytoplasmic proteins was removed and stored at -80 ℃ freezer. The pellet containing intact nuclear was further subjected to nucleaus preparation. The intact nuclei were resuspended in 50 µL buffer II (20 mM Tris-HCl, pH 7.9, 0.4 M NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM DTT and 1 mM PMSF, 0.1 µg/mL protease inhibitor cocktail) and vigorously vortexing for 15 minutes. Centrifugate the nuclei/buffer I suspension at 14000 rpm for 10 minutes at 4 ℃ to collect the soluble extracted fraction. The nuclear fraction can be stored at least six months at -80 ℃ freezer.
8. Site-directed Mutagenesis
The site-directed mutagenesis toward transcription factor binding sites on
IL-12 p40 promoter were manipulated according to the instruction manual of QuickChangeTM site-directed mutagenesis kit (STRATAGENE). In the beginning, prepare the sample reaction(s) as indicated bellow: 10X reaction buffer, primer pairs for target site (200 ng), PfuTurbo DNA Polymerase (2.5 U), template DNA (75 ng/mL) and 10 mM of dNTP.
PCR thermocycle profile was processed at 2 min at 95 ℃, 2 min at 64 ℃ and 10 min at 72 ℃ during 18 cycles. After PCR thermocycles finishing, utilize the restriction enzyme Dpn I to digest the original template, overnight. Thereafter, transfer the Dpn I-treated DNA for transformation.
9. DNA-sequencing
By Sequencing of all constructed and mutant plasmids was preformed by using Applied Biosystems BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit reagent. The sequencing mixture was prepared as described by manufacture’s instruction. The reaction was performed on a thermal cycles with a cycling condition of 30-sec at 95 ℃ for denaturing, 20-sec at 50 ℃ for annealing, and 4-min at 60 ℃ for elongation. Total 30 cycles were carried out.
The resulting fluorescence-labeled sequence was then product on an ABI 377 automatic sequencer.
10. EMSA (Electrophoretic mobility shift assay)
The probes were prepared by using biotin 3’end DNA labeling kit (PIERCE). Biotinylated probes were added to the binding reaction that also contained 2 µL binding buffer (100 mM Tris, 10 mM DTT, pH7.5), 2% glycerol, 0.5% NP-40, 5 mM MgCl2, 10 mM EDTA, and 2 µL labeled probe ( labeling efficiency is about 70% to 80% ). The binding mixtures were incubated at room temperature for 1.5 h, followed by adding 5 µL loading buffer. For supershift assays, 2 mg of specific Abs were used. For competition analysis, an excess of unlabeled oligocucleotides containing concensus binding sites for transcription factors was added to the binding reaction. Complex formation was proceeding for 1.5 h at 4 ℃. Finally, the samples were fractionated on a 6% nondenaturing polyacryamide gels in 0.5 X TBE buffer at 100 V for about 2.5 hour. The separated oligonucleotide probe and DNA/protein complex were transblotted to a nylon membrane (S&S) in the same buffer (0.5 X TBE buffer) at 380 mA for 45 min and cross-link the transferred membrane at 120 mJ/cm2 by using UV-light cross linker instrument, SPECTRO LINKERTM (Spectronics Inc.). After blocking at room temperature for 15 min, conjugating with strepavidin-HPR at room temperature for 15~20 min, washing using washing buffer for 6 min 3 times, and substrate equilibrium buffer for 5 min. Then, adding ECL reagent on the membrane for chemiluminescent reaction. The biotin labeled probes were visualized by exposing to an x-ray film (Hyperfilm).
11. MTT assay
Following removal of culture supernatants, cell viability was assessed by replacing medium and adding 500 µg/mL of 3-(4,5-dimethylthiazol-2-yl)
-2,5-diphenyl-tetra-zolium bromide (MTT). After incubation for 3 h at 37 ℃, medium was removed totally and the precipitate formed in metabolically active cells by the reduction of MTT to its insoluble MTT-formazan was dissolved in acidic isopropanol (1N), and the absorbance at 570 nm was determined.
III. Result
3.1 Construction of reporter plasmids of IL-12 p40 gene
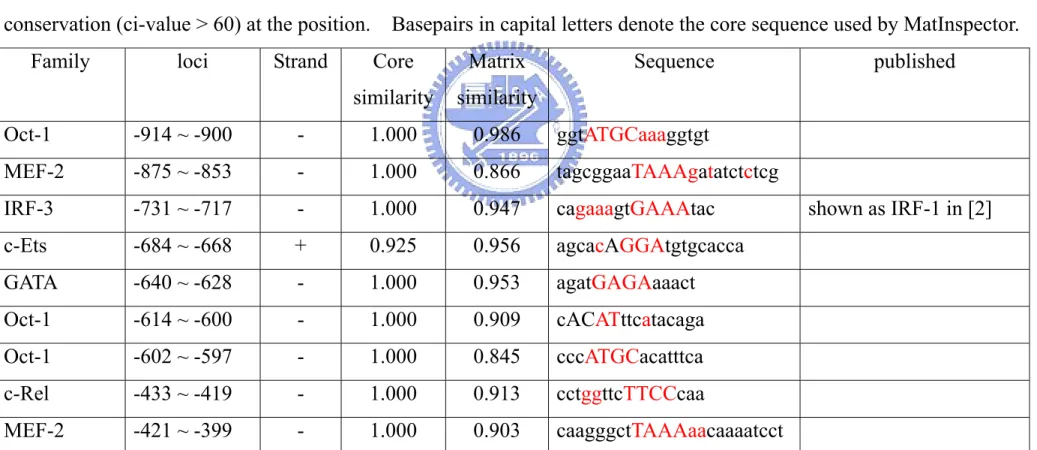
In primary peritoneal or bone marrow-derived macrophages, the protein and mRNA level of IL-12 p40 can be induced by the cotreatment of LPS and IFN-γ [2, 5, 42 and 43]. Similar result was also found in murine macrophage cell line, RAW264.7 [15]. Earlier work have shown that the proximal promoter region of human (-248 to +10) and murine (-286 to +10) share high sequence identify (~84%) upon DNA sequence alignment using “BLAST 2 sequence” sequence alignment program in NCBI web site. The region was found to contain several transcriptional elements, including NF-AT, IRF-1, AP-1. C/EBP, NF-κB, GA-12 and Ets (Fig. 1). To facilitate the reaching of putative responding elements in the rest promoter region (upstream of -280), we analyzed the promoter region (-922 to +33) of human IL-12 p40 gene using MatInspector® (published in [38] and cited by [39-41]) to find out potential responding elements in their region. In additional to known transcription factor binding sites, there are several putative responding elements including c-Rel, GATA, c-Ets, MEF-2 or Oct-1 in the IL-12 p40 promoter, were shown to have the highest similarities with consensus transcription factor binding sites (Table 5 and Fig. 2).
To further elucidates the molecular mechanism of LPS/IFN-γ-induced upregulation of IL-12 p40 gene transcription, we subcloned promoter region (-922 to +33) of human IL-12 p40 gene using genomic DNA from HEK293 cells as the template. The subcloned promoter fragment (~1kb) was then used to construct the pIL12p40/90 Lucneo reporter plasmid, which consisted of IL-12
p40 promoter region (-922 to +33) of IL-12 p40 gene, a luciferase reporter gene and a neomycin resistant gene/(Neor) (Fig. 6). In order to identify the promoter region in IL-12 p40 gene that may responsible for the activation of various
stimuli. A set of luciferase reporter plasmids containing various length of IL-12 p40 promoter, pIL12p40/90, pIL12p40/56, pIL12p40/40, pIL12p40/25, pIL12p40/20, pIL12p40/10, pIL12p40/7, pIL12p40/5, pIL12p40/0, were constructed (Fig. 6). The time-dependent effect of LPS and LPS/IFN-γ to the RAW(IL12p40/90) stable clone was first demonstrated by incubating with 1 µg/mL LPS at 37℃ for 3 to 15 h (Fig. 7). The result shows that the maximum stimutory effect of LPS was observed after incubation for 12 h, while, the activity reached maximum at about 6 h in response to LPS (1 µg/mL) and IFN-γ (100 U/mL).
Determine the NPT expression level in each RAW264.7 stable clone
The transfection efficiency of each stable clone was determined based on the protein level of constitutively expressed neomycin resistant gene, neomycin phospho-transferase (NPT) by Western blotting (Fig. 8A), β-actin was used as an internal control. The expression level of NPT protein in each stable clone was quantified using densitometry and normalized by comparing with the amount of β-actin. The RAW264.7 cells without transfecting were used as a negative control. Normalization index (NPT/β-actin ratio or transfection efficiency) of NPT in each stable clone is calculated as the relative expression level of NPT in each clone to the level of NPT in RAW(IL12p40/90) (Fig. 8B). Thus, the normalization index of RAW(IL12p40/90), RAW(IL12p40/56), RAW(IL12p40/40), RAW(IL12p40/25), RAW(IL12p40/20), RAW(IL12p40/10), RAW(IL12p40/7), RAW(IL12p40/5), RAW(IL12p40/0) were determined as 1.00, 0.63, 0.70, 0.97, 0.85, 0.55, 0.64, 0.61 and 0.44, respectively.
3.2 Deletion analysis of the promoter of IL-12 p40 gene
In previous studies, Rel/NF-κB (-116 to -106) and Ets-2 (-211 to -205) have been shown to be the potential transcription factors triggering the transcription of murine IL-12 p40 gene in response to LPS [2, 24, 25 and 26]. In constract to this observation, LPS induced significant luciferase activity in RAW(IL12p40/56) in addition to RAW(IL12p40/40) and RAW(IL12p40/25). The luciferase activity decreased markedly when the region upstream -190 was deleted (Fig. 9). Apparently, the region between -563 and -190 may contain essential responding elements that are important for the LPS-induced actiovation.
When stable clones cotreated with LPS and IFN-γ, a significant synergistic effect (p<0.001) was observed in the stable clone containing pIL-12p40/56, although RAW(IL12p40/25) (p<0.05), RAW(IL12p40/20) (p<0.05) and RAW(IL12p40/7) (p<0.05) exhibited moderate synergistic effect in the presence of both LPS and IFN-γ (Fig. 9). The LPS/IFN-induced luciferase activity reduced sharply when the region -563 to -398 was cleaved, suggesting the existence of the essential transcription responding element(s). Interestingly, a responding element c-Rel (-433 to -413) was potential to be responsible for the stimulation of LPS and LPS/IFN-γ. Consistent with previous studies, Ets-2 (-211 to -205), Rel/NF-κB (-116 to -106), C/EBP cooperated with AP-1 (-96 to -88), IRF-1 (-63to -53) and NF-AT (-54 to -42) may be involved in the upregulation of IL-12 p40 gene transcription in response to LPS and IFN-γ [2, 17, 26, 42, and 48-51]. The cell viability was not affected by the treatment of both LPS (1 µg/mL) and IFN-γ (100 U/mL) (data not shown).
The deletion analysis also reveals that LPS and IFN-γ can promote the luciferase activity of RAW(IL12p40/7) cells possibly via NF-AT and IRF-1 responding elements (Fig. 9). This result is consistent with the observation on murine IL-12 p40 gene [17 and 31].
The result from MatInspector® reveals that the promoter region from -68 to -48 may contain responding elements for IRF-1/NF-AT (Fig. 2). When treating RAW(IL12p40/7) with 1 µg/mL of LPS, the luciferase activity elevated five folds compared with the cells treating with medium only. A synergistic effect was also observed (~10 folds induction) by treating with both 1 µg/mL LPS and 100 U/mL IFN-γ . NF-AT is an essential transcription factor in mediating T cell activation and maturation. It will be interesting to known more about its role in macrophage.
Characterization of IL-12p40/56
The time-dependent effects of LPS, IFN-γ and LPS/IFN-γ on the stimulation of pIL-12p40/56 reporter plasmid were examined. As shown in Fig. 10, the stimulatory effects of LPS and IFN-γ could be clearly seen after 9 h of incubation, while co-treatment with LPS and IFN-γ the luciferase activity started increasing at 6 h. With treatment of LPS and IFN-γ, the activities of luciferase reached maximum at 12 h and 24 h, respectively. Compared with RAW(IL12p40/25), another region shown previously to be activated by LPS/IFN-γ co-treatment, the maximum activation reached after 8 h of incubation (Fig. 11).
The luciferase activity of RAW(IL12p40/56) in response to LPS/IFN-γ co-treatment could be significantly induced after 6 h incubation (Fig. 10). The stimulatory effect of LPS/IFN-γ co-treatment reached maximum at 12 h and declined slowly afterward. In the preliminary study, the luciferase activity of RAW(IL12p40/90) could be induced by LPS started from 3 h. The activity reaches maximum at about 12 h and rapidly decline to close to basal level after incubation for 24 h (data not shown). This result suggests that some unknown transcription factors that counter act at the stimulatory effect of LPS may bind to the promoter region -922 to -563 to suppress the transcription of luciferase.
Previous work has shown that LPS and IFN-γ exhibits strong synergistic effect to the luciferase activity of RAW(IL12p40/56) from 10 fold increase (with 1 µg/mL LPS) to 60 folds increased (with both 1 µg/mL LPS and 100 U/mL IFN-γ). Compared with stimulatory effect of LPS/IFN-γ to regions -245 to -190 (containing Rel/NF-κB and Ets sites) and -68 to -48 (containing NF-AT and IRF-1 sites), the region between -563 to -398 is more essential than above two regions.
The dosage of LPS and IFN-γ to activation of RAW(IL12p40/56) was further studied. The result of Fig. 12A showed that in the presence of fixed concentration of 100 U/mL IFN-γ, the stimulatory affect of LPS reached maximum at about 5 µg/mL. However, with fixed concentration of LPS (1 µg/mL) the stimulatory effect of IFN-γ increase slightly in the concentration range from 50 U/mL to 1000 U/mL (Fig. 12B). The result shows LPS and IFN-γ can induce the expression of the IL-12 p40 gene in a does-dependent manner. Moreover, LPS exhibits a stronger effect than IFN-γ does.
To further elucidate the role of the region -563 to -398, we transfer the promoter region -563 to -398 to the upstream of TATA box in the pIL12p40/5 (Fig 13). Τhe resulting plasmids pIL12p40/56-40 was then transfected into RAW264.7 to generate a stable clone RAW(IL12p40/56-40) as described in
Materials and Methods. The transfection efficiency of the reporter plasmid in
both RAW(IL12p40/56) and RAW(IL12p40/56-40) were determined by Western Blotting (Fig. 13). As showed in Fig. 14, RAW(IL12p40/56-40) can be activated by 1 µg/mL LPS alone, or LPS/IFN-γ in RAW(IL12p40/56-40) in a time-dependent manner. The luciferase activity of RAW(IL12p40/56-40) could also be induced by LPS and LPS/IFN-γ reached maximum at about 12 h and declined afterward. Notably, the synergistic effect of LPS/IFN-γ could not be seen in RAW(IL12p40/56-40) stable clone. It is postulated that an IFN-γ responsive element may located at the border around -398. Alternatively, the
transcription factor that binds to promoter region -563 and -398 depends on other TF downstream its binding site for its trans-activation activity. Thus, the removal of -398 to -48 may lead to lost of synergistic effect of LPS and IFN-γ.
Elucidation of novel c-Rel binding site
Based on the sequence alignment using MatInspector® (Genomatrix), we found that -431GGGAAGAACC-424 is similar to c-Rel responding element, a member of NF-κB family. c-Rel, a member of NF-κB family, shares a highly conserved 300-amino acid Rel homology domain at N-terminal (which encompasses sequences required for DNA binding, protein dimerization, and nuclear translocation) with other Rel/NF-κB proteins [44]. IL-12 p40 production would be blocked by c-Rel-deficient cell [45]. All published studies demonstrated that c-Rel could bind to the promoter region from -132 to -122 and cooperated with C/EBP and p50, another NF-κB member, to promote the transcription of murine and human IL-12 p40 gene. Interestingly, a putative c-Rel site was also found in region -433 to -412 upstream of the known Ets site, according to the result of MatInspector® (Table 5 and Fig. 2), we like to study whether this site is responsible for the LPS/IFN-γ-induced IL-12 p40 upregulation. To demonstrate that c-Rel responsing element is located in the promoter region -563 and -398, we synthesized double stranded oligonucleotide probe (25 bp) that directly copied the proposed c-Rel corresponding element in the region. This probe was used in eletrophoretic mobility shift assay (EMSA) to determine whether c-Rel is activated in response to LPS or LPS/IFN-γ treatment. Nuclear protein from RAW264.7 cell treated with medium upon formed two complexes, we named it complex I and II. And then cell treatement with LPS for 2, 4 and 6 h, complex I declined in a time-independent manner and complex II is increased by treating with LPS. When co-treating of LPS and IFN-γ, complex II is enhanced along with incubation time (see Fig. 15).
Characterizing the NF-AT binding site in the human IL-12 p40 gene
The NF-AT binding site in -69 to -48 of the IL-12 p40 promoter is also an interesting target that is worth to understand more about it. The promoter region in reporter plasmid pIL12p40/7 Lucneo has been shown to contain both
NF-AT and IRF-1 responding elements along with a TATA sequence. In previous studies, we have demonstrated that LPS/IFN-γ could synergistically induced the luciferase activity of RAW(IL12p40/7) (Fig. 9). As indicated by Zhu and colleagues that LPS and IFN-γ could activate murine IL-12 p40 gene promoter by facilitating a functional interaction between NF-AT and ICSBP [17]. The DNA sequence of NF-AT binding site in human IL-12 p40 gene
-54AAGTTTCCATCAG-42 exhibits (purine nucleotides are underlined) exhibits a
high similarity to the one in murine IL-12 p40 gene, -62GGGTTTCCATCAG-52. Upon sequence alignment, the human IL-12 p40 gene also contains an IRF-1 site--72TTCTACTTTGGGTTT-58, which is highly homologous to IRF-1 site--63TTCTATGTTTAAGTTT-49 in murine IL-12 p40 gene. In murine IRF-1 site the short stretch -61 to -63 is critical for the binding of transcription factor, which is not conserved in human proposed IRF-1 site. We then performed site-directed mutagenesis to selectively remove IRF-1 or NF-AT responding element to verify the role of these two transcription factors in the LPS and IFN-γ-induced human IL-12 p40 gene transcription. Two mutants of reporter plasmids pIL-12p40/7 Lucneo, termed pIL-12p40/7∆I Lucneo and pIL12p40/7∆N
Lucneo, were constructed (Fig. 16A) and transfected into RAW264.7 cell lines to
generate corresponding stable clones as described previously. The copy number of RAW(IL12p40/7∆I) and RAW(IL12p40/7∆N) were determined as the NPT expression through Western Blotting (Fig. 16B). Interestingly, we found that mutation on NF-AT site of pIL12p40/7 totally lost ability to respond to the stimulation of LPS or LPS/IFN-γ (Fig. 17). The mutation on IRF-1 binding site, however exhibit normally in respond to the induction of LPS or LPS/IFN-γ. Hence, it is apparent that LPS or LPS/IFN-γ induces luciferase
activity of RAW(IL12p40/7) cells through the binding site of NF-AT(Fig. 17). CsA (Cyclopsprine A) is an immunosuppressent for organ transplanted patients. In T cells, CsA could inhibit a phosphatase called calcineurin and blocked the dephosphorylation and nuclear translocation of NF-AT [46]. In macrophage, the role and signaling pathway of NF-AT is still unknown. Although CsA was shown to block the function of NF-AT in T cells, the RAW(IL12p40/7) cells pretreated with CsA for 1h, followed by adding LPS enhanced the luciferase activity of the cell up to 3-fold. CsA had alittle effect on the IFN-γ and LPS/IFN-γ treated cells. Interestingly, the mutation on IRF-1 binding site of IL12p40/7 suppressed the CsA potentiated LPS-induced activity (Fig. 18).
3.3 Generation of IL-12p40/90 containing RAW264.7 stable clones
We have generated the pIL-12p40/90 Lucneo stably transfected RAW264.7
clones (RAWIL12p40/90/A1~A20 and RAWIL12p40/90/B1~B30) as a cell-based drug screening system. The difference between these single stably clones and previously stable clones was that all single stable cells were divided from a single transfected cell. The positive clones were screened under the selection stress of neomycin. LPS, is a potent activator to induce IL-12 production in antigen-presenting cells (APCs), the effectiveness of the neomycin resistant clones were further verified for their responses to the stimulation of LPS. Further selection was performed by treating cells with 1 µg/mL of LPS at 37 ℃ for 12 h. We obtained fifty single stably transfected cells and promoter activity between 0 and 10-fold included thirty seven clones, between 10 and 20-fold included two clones, between 40 and 50-fold also included two clones (Fig. 19A). Eight clones which could not only resist the selection of 1000 µg/mL of G418-sulfate, but also could sensitively respond to the stimulation of 1 µg/mL of LPS. Based on the luciferase activity of each stable clone in response to the stimulation of LPS, we found that clones A1, B21 and B30 exhibited strong responses (over 25-fold relative to control cells), toward the stimulation of LPS (Fig. 19B). In order to respond the activation of herbs extracts-induced promoter activity, we picked B21 clones as a cell-based drug screening model for the screening of bioactive components in the Chinese herbs.
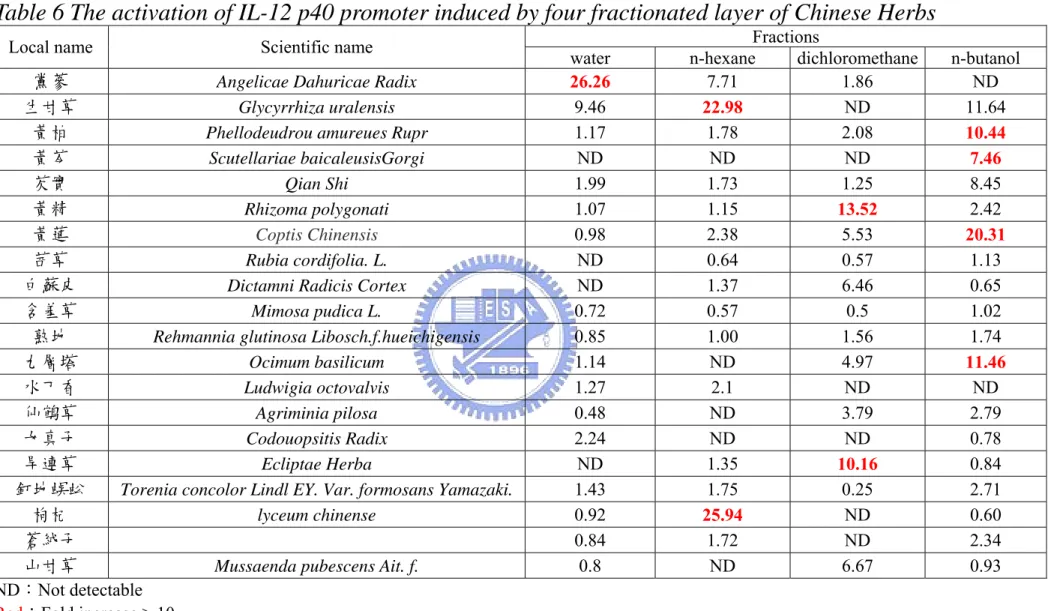
Table 3 demonstrated about 25 kinds of Chinese herbs as our source for drug screening and they were classified into six types of herbs. The Chinese herbs were then fractionated into four parts based their polarity with the help of professor, Ho Jian-Ching of the Ta-Hwa Institude of Technology. A code was assigned to each fraction from Chinese herbs. The B21 cells were treated with variously Chinese herb extracts (100 µg/mL) at 37 ℃ for 12 h. Table 6 showed the luciferase activity induced by the Chinese herb extracts and red words meant
the activity higher than 10-fold. Herb extracts in dichloromethane or n-butanol layers exhibited the potential stimuli toward activating IL-12 p40 promoter activity.
Analyzing the bioactivities of Chinese herb extracts
After high-throughout herbs screening, we chose some Chinese herb extracts to further study the molecular mechanism.
Fig. 20, 21, 22 and 23 showed four kinds of Chinese herb extracts induced human IL-12 p40 gene transcription in a concentration dependent manner. We used deletion analysis to understand which region of promoter was critical for the activation by Chinese herb extracts. The deletion analysis was also normalized as described previously. Different Chinese herb extracts exhibited different activation pattern toward the same set of reporter plasmids. It means that different Chinese herb extracts may mediate IL-12 p40 gene via distinct pathways. For example, 8-4 enhanced significant promoter activity in RAW(IL12p40/90), RAW(L-12p40/56), RAW(IL12p40/40) and RAW(IL12p40/25). The luciferase activity decreased markedly when the region between nucleotide sequences -245 to -190 was deleted. This region from -211 to -205 of IL-12 p40 promoter contains a cis-responding element for Ets-2, c-Rel and ICSBP.
Furthermore, we detected cell viability by MTT to confirm the Chinese herbs induced IL-12 p40 promoter activation was not resulted from cell proliferation, see Fig. 24. Three herbs extracts, including 5-4, 8-4 and 15-2 treated cells with no significant change of cell viability (data of 5-4 is not shown). Surprising, 100 µg/mL 16-3 reduced the cell viability, but the same of dosage could also enhance the IL-12 p40 promoter. Maybe, there are some bioactive compounds and some cytotoxic compounds residing the Chinese herb extracts, 16-3. When the dosage of 16-3 reduces from 100 µg/mL to 50 µg/mL, the cell viability has no significant division and promoter activity only reduces 10%.
IV. Discussion
IL-12 plays a regulatory role in the innate and acquired immunities. The physiological functions have been identified, but the molecular mechanism study is still in the initial stage.
It has been demonstrated that the Ets-2 (-211 to -205), NF-κB (-116 to -106) and C/EBP (-75 to -64) sites in IL-12p40 promoter are essential for the stimulatory effect of LPS+IFN-γ in APCs [2, 24 and 26]. In our work, LPS or LPS/IFN-γ stimulate pIL12p40/25 (containing Ets-2, NF-κB and C/EBP binding sites) at 37 ℃ for 12 h to 8 and 25 folds (fold increase of medium only and the fold indicated fold increase without normalization). When truncated the Ets-2 binding element, the promoter activity reduced 50%. Hence, our result confirmed the previously studies of LPS- or LPS/IFN-γ-induced IL-12 p40 promoter activity. Interestingly, promoter fragment -563 to -398 of IL-12 p40 promoter could also be stimulated after LPS and LPS/IFN-γ to about 8 and ~50 fold. In the time course of RAWIL12p40/56 and RAWIL12p40/25 (Fig. 10 and 14), activation of IL12p40/56 was long-life and stronger than IL12p40/25. In other words, inducing the putative transcription factor(s) to bind to the promoter region from -563 to -398 can hold the activation of IL-12p40 gene. In the reporter of Ma et al, they used human IL-12 p40 promoter reporter plasmids in murine monocyte-like cell line. They constructed whole IL-12 p40 promoter (from -3300) and depleted to -292, -265, -243, -222, -204 and TATA. When truncated to -292, the promoter activity reduced to 10 fold and only 40% of whole promoter reporter plasmids contained cells. Ma et al suggested that the reduction of promoter activity was not result from TFs binding to the NF-IL6 (-512 to -505) and IRF-1 (-703 to -719) [2]. Therefore, they had even tried to find some TF binding element(s) which was upstream of Ets-2. Utilizing the bioinformatics method, we found the putative transcription factor(s) residing the
promoter region of –563 to –398 might contain c-Rel, a member of NF-κB family. In order to narrow down the critical region of LPS/IFN-γ-induced activity, we transfer the promoter region -563 to -398 to the upstream of TATA box in the IL-12p40/5 (Fig 13). Alternatively, the transcription factor that binds to promoter region -563 and -398 depends on other TF downstream its binding site for its trans-activity. Thus, the removal of -398 to -48 may lead to lost of synergistic effect of LPS and IFN-γ. Thereafter, we performed the EMSA by using Biotin-labeled oligo-probe containing the putative c-Rel binding site (-433 to -413) (Fig. 15). Manson and colleagues identified the c-Rel-dependent and -independent pathways of IL-12 production during infection and inflammatory stimuli [45]. They also demonstrated that c-Rel was required for IL-12 p40 production by macrophages in response to Coryne-bacterium parvum, CpG oligodeoxynucleotides, anti-CD40 and low molecular weight hyaluronic acid. To data, c-Rel has not been characterized as a major enhancer transcription factor of IL-12 p40 activation. In the future, we want to further confirm the TF complex components and the role of putative c-Rel in regulation human IL-12 p40 gene transcription.
IL-10 could abrogate LPS- or IFN-γ- induced murine IL-12p40 promoter by interaction of NF-AT and ICSBP [17]. Another similar reporter revealed that ICSBP could activate the p40 promoter in cooperation with IRF-1 through an ISRE (interferon stimulated responsive element)-like element [47]. To regard with the role of NF-AT and IRF-1 in the regulation of IL-12 p40 gene transcription, we identified the relative locations of NF-AT and IRF-1 binding sites in human IL-12 p40 promoter by alignment (using BLAST 2 sequence and MatInspector®) and constructed the RAW(IL12p40/7), RAW(IL12p40/7∆I) and RAW(IL12p40/7∆N). Zhu et al demonstrated that LPS, IFN-γ or both can activate transcription proceeding of murine IL-12 p40 gene by NF-AT to recruit ICSBP to bind to the NF-AT binding element. To mention with regulation of human IL-12 p40 gene transcription level, NF-AT can upregulate the
transcription activity of IL-12 p40, too. IRF-1,aAnother TF onto the promoter region of -63 to -53, may be an inhibitory factor, because the promoter pIL12p40/7 with truncation of IRF-1 binding site leads to enhance the promoter activity. Μaruyama et al reported that IRF-1 played an inhibitory role in down-regulating the transcription of IL-12 p40 and the production of IL-12 p70, when mice were infected by Plasmodium berghri [53].
We also utilized the pIL12p40/90 Lucneo stably transfected cell clones, the
promoter-based screening system, to select the bioactive function extracts from about 30 Chinese herbs. To find out which component(s) is capable of activation the IL-12 p40 promoter, we proceed with the further isolation of these extracts and sieving out the active component(s) in these extracts. Table 6 shows that the herb extracts induced IL-12 p40 promoter transcription activity significantly. In order to understand the regulation of herbs extracts-induced promoter activity, deletion analysis of human IL-12 p40 promoter is to be used. Fig. 20, 21, 22 and 23 demonstrated four kinds of herb extracts induced human IL-12 p40 gene transcription in a concentration dependent manner. Based on the understanding of regulation of IL-12 p40 transcription level, 8-4 and 15-2 maybe enhance the NF-κB/Rel (-210 to –205, relevant to transcription starting site) or the interaction of NF-AT and ICSBP (-62 to -57). 16-3 may induce IL-12 p40 promoter transcription through the putative c-Rel TF binding element. However, 50 µg/mL 16-3 is a proper dosage to treating cells without triggering cells to apoptosis. Hence, 16-3 treated RAW264.7 might activate IL-12 p40 through other pathway.
References
[1] Roberson, M. J. and Ritz, J. Interleukin 12: Basic biology and potential applications in cancer treatment. The Oncologist. 1, 88-97. 1996.
[2] Ma, X., Chow, J. M., Gri, Giorgia., Carra, G., Gerosa, F. and Trinchieri, G. The interleukin 12 p40 gene promoter is primed by interferon-γ in
monocytic cells. J. Exp. Med. 183, 147-157, 1996.
[3] Soiffer, R. J., Roberson M. J., Murray, C. and Ritz, J. Interleukin-12
augments cytolytic activity of peripheral blood lymphocytes from patients with hematologic and solid malignancies. Blood 82, 2790-2796, 1993. [4] Zeh, H. J. III., Hurd, S., Storkus, W. J. and Lotze, M. T. Interleukin 12
promotes the proliferation and cytolytic activity of immune effectors: implications for the immunotherapy of cancer. J. Immunother. 14, 155-161, 1993.
[5] Bigda, J., Mysliwska, J., Dziadzuszko, R., Bigda, J., Mysliswki, A. and Hellmann, A. Interleukin 12 augments natural killer cell mediated cytolytic in hairt cell leukemia. Leuk. Lymphoma 10, 121-125, 1993.
[6] Hiraki, A., Kiura, K. Yamane, H., Nogami, N., Tabata, M., Takigawa, N., Ueoka, H., Tanimoto, M. And Harada, M. Interleukin-12 augments
cytolytic activity of peripheral blood mononuclear cells against autologous lung cancer cells in conbination with IL-2. Lung Cancer 35, 329-333, 2002.
[7] Parihar, R., Dierksheide, Y., Hu, Y. and Carson, W. E. IL-12 enhances the natural killer cell cytokine response to Ab coated tumor cells. J. Clin.
Invest. 110, 983-992, 2002.
[8] Schwarze, J., Hamelmann, E., Cieslewicz, G., Tomkinson, A., Joetham, A., Bradley, K. and Gelfand, E. W. Local treatment with IL-12 is an effective inhibitor of airway hyperresponsiveness and lung eosinophilia after airway challenge in sensitized mice. J. Allergy Clin. Immunol. 102, 86-93, 1998.
[9] Stirling, R. G. and Chung, K. F. New immunological approaches and
cytokine targets in asthma and allergy. Eur. Respir. J. 16, 1158-1174, 2000. [10] Sur, S., Bouchard, P., Holbert, D. and Van Scott, M. R. Mucosal IL-12
inhibits airway reactivity to methacholine and respiratory failure in murine asthma. Exp. Lung Res. 26, 477-489, 2000.
[11] Karp, C. L., Biron, C. A. and Irani, D. Interferon-γ in multiple sclerosis:is IL-12 supression the key?Immunology today 21, 24-28, 2000.
[12] Rabinovitch, A., Suarez-Pinzon, W. L. and Sorensen,O. Interleukin 12 mRNA expression in islets correlates with beta-cell destruction in NOD mice. J Autoimmun, 9, 645-51, 1996.
[13] Liu, J. and Beller, D. I. Distinct pathways for NF-kappa B regulation are associated with aberrant macrophage IL-12 production in lupus- and diabetes-prone mouse strains. J Immunol, 170, 4489-96, 2003.
[14] Jun, H. S., Yoon, C. S., Zbytnuik, L., Van Rooijen, N. and Yoon, J. W. The role of macrophage in the T cell-mediated autoimmune diabetes in nonbese diabetic mice. J. Exp. Med. 189, 347-358, 1999.
[15] Xiong, H., Zhu, C., Li, F., Hegazi, R., He, K., Babyatsky, M., Bauer, A. J., and Plevy, S. E. Inhibition of interleukin-12 p40 transcription and NF-κB activation by nitric oxide in murine macrophages and dendritic cells. J.
Biol. Chem. 279, 10776-10783, 2004.
[16] Kang, B. Y. Chung, S. W., Cho, D. and Kim, T. S. Involvement of p38 mitogen-activated protein kinase in the induction of interleukin-12 p40 production in mouse macrophages by berberine, a benzodioxoloquinolizine alkaloid. Biochem Pharmaco. l63, 1901-1910, 2002.
[17] Zhu, C., Rao, K., Xiong, H., Gsgnidze, K., Li, F., and Horvath, C. activation of the murine interleukin-12 p40 promoter by functional
interactions between NFAT and ICSBP. J. Biol. Chem. 278, 39372-39382, 2003.
[18] Cor, F. C. and Geert, A. B. Multiple steps in the regulation of
transcription-factor level and activity. Biochem. J. 317,329-342, 1996. [19] Goodridge, H.S., Harnett, W., Liew, F.Y. and Harnett, M. Differential
regulation of interleukin-12 p40 and p35 induction via Erk
mitogen-activated protein kinase-dependent and -independent mechanisms and the implications for bioactive IL-12 and IL-23 responses. Immunology
109, 415-425, 2003
[20] Mausumee, G. and Nackman. LPS induction of gene expression in human monocytes. Cellular Singnalling 13, 85-94, 2001.
[21] Trinchieri Giorgio. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. 3, 133-146, 2003.
[22] Sun, Z. and Roland, A. NF-κB activation and inhibition: a review. Shock 18, 99-106, 2002.
[23] Medzbitov, R. and Janeway, C. The toll receptor family and microbial recognition. Trends in Microbiology. 8, 452-256, 2000.
[24] Becker, C., Wirtz, S., Ma, X., Blessing, M., Galle, P. R., and Neurath, M. F. Regulation of IL-12 p40 promoter activity in primary human monocytes: roles of NF-κB, CCAAT/enhancer-binding protein , and PU.1 and
identification of a novel repressor element (GA-12) that responds to IL-4 and prostaglandin E2. J. Immunol. 167, 2608-2618, 2001.
[25] Murphy, T. L., Clevelend, M. G., Kuleaza, P., Magnam, J. and Murphy, K. M. Regulation of interleukin 12 p40 expression through an NF-κB half-site. Mol.Cell. Biol. 15, 5258-5267, 1995.
[26] Ma, X., Neurath, M., Gri, G., and Trinchieri, G. Identificaiton and
characterization of a novel Ets-2-related nuclear complex impacted in the activation of the human interleukin-12 p40 gene promoter. J. Biol. Chem.
272, 10389-10395, 1997.
interferon-γ. Annu. Rev. Immunol., 15, 749-795, 1997.
[28] Brewington, R., Chatterji, M., Zoubine, M., Miranda, R.N., Norimatsu, M., and Shnyra. IFN-γ-independent autocrine cytokine regulatory mechanism in reprogramming of macrophage responses to bacterial
lipopolysaccharide. J. Immunol. 167, 392-398, 2001.
[29] Kotento, S. V., Izotova, L. S. Pollack, B. P., Mariano, T. M., Donnelly L., Muthukumaran, G., Cook, J. R., Garotta, G., Silvennoinen, O., Ihle, J. N. and Pestka, S. Interaction between components of the interferon-γ receptor complex. J. Biol. Chem. 270, 20915-20921, 1995.
[30] Kotento, S. V., Izotova, L. S.,Pollack, B. P. Muthukumaran, G., Paukku, K., Sivennoinen, O., Ihle, J.N. and Pestka, K. Other kinases can substitute for Jak2 in signal transduction by interferon-γ. J. Biol. Chem. 271,
17174-17182, 1996.
[31] Maruyama, S., Sumita, K., Shen, H., Kanoh, M., Xu. X., Sato, M., Matsumoto, M., Shinomiya H., and Asano, Y. Identification of IFN regulatory factor-1 binding site in IL-12 p40 gene promoter. J. Immonol.
170, 997-1001, 2003.
[32] Gri, G., Chiodoni, C., Gallo, C. E. Stoppacciaro, A., Liow F. Y. and
Colombo, M. P. Antitumor effect of interleukin (IL)-12 in the absence of endogenous IFN-γ: a role fro intrinsic tumor immunogenicity and IL-15.
Cancer Res. 62, 4390-4397, 2003.
[33] Taniguchi, T., Lamphier, M., and Tanaka, N. IRF-1: the transcription factor linking the interferon response and oncogenesis. Biochim. Biophys. Acta.
1333, M9-17, 1997.
[34] Taniguchi, T. Cytokine signal through nonreceptor protein tyrosine kinases.
Science 268, 251-255, 1995.
[35] Wang, I. M., Contursi, C. Masumi, A., Ma, X., Trincheri, G., and Ozato, K. An IFN-γ-inducible transcription factor, IFN consensus sequence binding
protein (ICSBP), stimulates IL-12 p40 expression in macrophages. J.
Immonol. 165, 271-279, 2000.
[36] Scharton-Kerten, T., contursi, C., Masumi, A., Sher, A., and Ozato, K. Interferon Consensus Sequence Binding Protein-deficient Mice Display Impaired Resistance to Intracellular Infection Due to a Primary Defect in Interleukin 12 p40 Induction. J. Exp. Med. 186, 1523-1534. 1997.
[37] McElligott, D. L., Philips, J. A., Stillman, C. A., Koch, R. J., Mosier, D. E. and Hobbs, M. V. CD4+ T cells from IRF-1-deficient mice exhibit altered patterns of cytokine expression and cell subset homeostasis. J. Immunol.
159, 4180-4186, 1997.
[38] Quandt, K., Frech, K., Karas, H., Wingender, E., Werner, T. MatInd and MatInspector: new fast and versatile tools for detection of consensus matches in nucleotide sequence data. Nucleic Acids Res. 23, 4878-4884, 1995.
[39] Alpy F., Boulay A., Moog-Lutz C., Andarawewa, K. L., Degot S., Stoll I., Rio M. C. and Tomasetto C. Metastatic lymph node 64 (MLN64), a gene overexpressed in breast cancers, is regulated by Sp/KLF transcription factors. Oncogene 22, 3770-3780, 2003.
[40] Liedtke C., Groger N., Manns M. P., Trautwein C. The human caspase-8 promoter sustains basal activity through SP1 and ETS-like transcription factors and can be up-regulated by a p53-dependent mechanism. J Biol
Chem. 278, 27593-27604, 2003.
[41] Ufer C., Borchert A., Kuhn H. Functional characterization of cis- and trans-regulatory elements involved in expression of phospholipid
hydroperoxide glutathione peroxidase. Nucleic Acids Res. 31, 4293-4303, 2003.
[42] Plevy, S. E., Gemberling, J. H., Hsu, S., Dorner, A. J. and Smale, S. T. Multiple control elements mediate activation of the murine and human interleukin 12 p40 promoters:evidence of functional synergy between
C/EBP and Rel proteins. Mol.Cell. Biol. 17, 4572-4588, 1997. [43] Amy, C. M., Caleph, B. W., Manujendra, R. and Pamela, H. C.
Macrophage-Stimulating protein, the ligand for the stem cell-derived tyrosine kinase/RON receptor tyrosine, inhibits IL-12 production by primary peritoneal macrophages stimulated with IFN-γ and
lipopolysaccharide. J. Immunol. 172, 1825-1832, 2004.
[44] Brendan, A. H., Nicola, M., Lingyun, X., Jing, S., Salah-Eddine, L. C., Hsiou, C. L., Christopher, H. and Youhai, C. Critical role of c-Rel in autoimmune inflammation and helper T cell differtiation. J. Clin. Inverst.
110, 843-850, 2002.
[45] Manson, N., Aliberti, J., Caamano, J. C., Liou, H. C. and Hunter, C. A. Cutting edge: Identifivation of c-Rel-dependent and independent pathways of IL-12 rpoduction during infectious and inflammatory stimuli. J. Immunol.
168, 2590-2594, 2002.
[46] Hogan, P. G., Chen, L., Nardone, J. and Rao, A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes & Dev. 17, 2205 – 2232, 2003. [47] Masumi, A., Tamaoki, S., Wang, I. M., Ozato, K. and Komuro, K.
IRF-8/ICSBP and IRF-1 cooperatively stimulate mouse IL-12 promoter activity in macrophages. FEBS Letters 531, 348-353, 2002.
[48] Zhu, C., Gagnidze, K., Gemberling J. H., and Plevy, S. E. Charaterization of an activation protein-1-binding site in the murine interleukin-12 p40 promoter. J. Biol. Chem. 276, 18519-18528, 2001.
[49] Luo, Q., Ma, X., Wahl, S. M., Bieker, J. J. Crossley, M. and Montaner, L. J. Activation and repression of interleukin-12 p40 transcription by erythroid kruppel-like factor in macrophages. J. Biol. Chem. 279, 18451-18456, 2004.
[50] Zhang, J. S., Feng, W. G., Li, C. L., Wang, X. Y., and Chang, Z. L. NF-κB regulates the LPS-induced expression of interleukin 12 p40 in murine peritoneal macrophages: roles of PKC, PKA, ERK, p38 MAPK, and
proteasome. Cell. Immunol. 204, 38-45, 2000.
[51] Cowdery, J. S., Boerth, N. J., Norian, L. A., Myung, P. S. and Koretzky, G. A. Differential regulation of the IL-12 p40 promoter and of p40 secretion by CpG DNA and lipopolysaccharide. J. Immunol. 162, 6770-6775, 1995. [52] Li, J. J. The development of NF-IL6 and NF-κB based drug screening
system and its application on the study of Chinese traditional herbs. A thesis
Submitted to National Chiao Tung university for the Degree of Master in Science. 2001.
[53] Xu, X., Sumita, K., Feng, C., Xiong, X., Shen, H., Maruyama, S., Kanoh, M. and Asano, Y. Down-regulation of IL-12 p40 gene in Plasmodium
berghei-infected mice. J. Immunol. 167, 235-241, 2001.
[] 現代中藥彙整,柯達製藥公司編輯小組,柯達製藥股份有限公司,桃園, 民國八十四年。