國 立 交 通 大 學
生化工程研究所
碩 士 論 文
酸鹼值與脫落酸在台灣山櫻花種子解除
休眠上的影響
The effect of pH and abscisic acid on the seed
dormancy-breaking of Prunus campanulata Maxim.
研究生:吳秀華
指導教授:楊裕雄 教授
酸鹼值與脫落酸在台灣山櫻花種子解除休眠上的影響
The effect of pH and abscisic acid on the seed dormancy-breaking of Prunus
campanulata Maxim.
研 究 生:吳秀華 Student: Hsiu-Hua Wu
指導教授:楊裕雄 教授 Advisor: Prof. Yuh-Shyong Yang
國 立 交 通 大 學
生化工程研究所
碩 士 論 文
A Thesis
Submitted to Institute of Biochemical Engineering College of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biochemical Engineering July 2008
Hsinchu, Taiwan, Republic of China
i
酸鹼值與脫落酸在台灣山櫻花種子解除休眠上的影響
研究生:吳秀華 指導教授:楊裕雄 教授 國立交通大學生化工程研究所碩士班 摘要 種子休眠為植物最重要的生理現象之一,然而其生化的調節機制至今仍不明朗。 台灣山櫻花種子具有深度休眠的特性,並且其休眠的程度會大大地受到外覆組織的影 響,該性質使得其相當適合用於研究種子休眠的調節。在山櫻花種子解除休眠的過程 中,可以觀察到大量亞硫酸基水解酵素(sulfatase)的催化活性,因此可能產生酸性的亞硫酸根離子(HSO3-)。過去的研究已在紅米(red rice)證實酸鹼值具有影響種子解除休
眠的能力,並且亦有其他文獻指出脫落酸(abscisic acid)在葉細胞與細胞間隙之間的分 布,會受到酸鹼值梯度的影響,且未解離的脫落酸為主要可以穿透組織的形式。然而, 細胞如何調節本身的酸鹼值去回應環境中的刺激,以及參與其中的生化調節因子至今 仍未被詳細記載。首先,在本篇研究中發現,在解除休眠的過程中,台灣山櫻花種子 組織中的酸鹼值有隨著時間下降的趨勢。酸鹼值的不同亦會影響種子的發芽率與胚根 的生長,這兩種生理特性可作為判斷種子是否解除休眠的指標。此外,將真種子(true seed)培養於含有外源性脫落酸的低酸鹼值環境下,將會明顯地降低種子的發芽率與胚 根的生長。最後,利用二維電泳分析法評估在解除休眠過程中,酸鹼值對於種子蛋白 質模式的影響,並闡述扁桃腈裂解酶前驅物((R)-mandelonitrile lyase 1)和洋李(prunin) 與種子解除休眠之間的關係。
ii
The effect of pH and abscisic acid on the seed dormancy-breaking of
Prunus campanulata Maxim.
Student: Hsiu-Hua Wu Advisor: Prof. Yuh-Shyong Yang Institute of Biochemical Engineering, College of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan, ROC
ABSTRACT
The biochemical basis of the regulation of seed dormancy, one of the most pivotal physiological phenomena of the plants, is still poorly understood. Prunus campanulata Maxim. seeds are deeply dormant and completely imposed by covering layers (testa and endocarp), which makes them a suitable model for studying the regulation of seed dormancy. The significant sulfatase activity was observed during the seed dormancy-breaking of P. campanulata, which produced acidic hydrogen sulfate. It has been demonstrated that dormancy-breaking activity was pH-dependent in red rice. Abscisic acid (ABA) is found to distribute between leaf cells and free space according to pH gradients, with the un-dissociated ABA (acidic form) being the main penetrating species. However, how the cells adjust their pHs in response to the environmental stimulation and what are their biochemical regulations have not been studied in detail. In this study, we found that the pH of P. campanulata seed tissue declined during the dormancy-breaking process. The variations of pHs also affected germination percentage and radical growth, which was used as indictors resulted from the seed dormancy-breaking. In addition, the presence of exogenous ABA obviously decreased the germination percentage and radical length of the true seeds at low pH values. Finally, the effects of pH on the protein patterns of the seeds during the dormancy-breaking process was evaluated by two-dimension (2-D) electrophoretic analysis, and (R)-mandelonitrile lyase 1 precursor and prunin were identified in relation to the seed dormancy-breaking.
iii ACKNOWLEDGEMENT 時光荏苒,歲月流逝,轉眼間兩年的旅程已接近終點,回首過去,LEPE依然是 最佳的選擇。首要感謝的是深知灼見的 楊裕雄教授,自主管理的領導優勢,亦師亦 友的肺腑建言,加上首屈一指的研究資源,讓學生在這兩年中受益良多且收穫豐盛。 再者,感謝林業試驗所 簡慶德博士於實驗上的協助,以及論文研究上的指導與 勉勵,使此研究內容得以更加充實及完備。感謝口試蒞臨指導的 林彩雲教授與 李家 維 教授,您們深入且整體性的指教建議,使得論文與研究更臻完美。此外,感謝郁 吟學姊、青辰學長與小志學長於實驗及論文上的教授及指導;謝謝政哲、程允、淵仁 學長提供不同領域的看法與經驗,怡宏、陸宜、江平學長、小米、美春學姐所給予的 諸多意見,以及家煒、小胖、若芬、咏馨、音汝等的陪伴。希望大家在LEPE這樣良 好的資源環境可以做出更卓越的研究。 最後,由衷感激撫育秀華二十幾年的雙親,感謝您們不辭辛勞的養育之恩與默默 的支持和鼓勵,讓秀華得以無後顧之憂地向目標邁進。此外,好姐妹們,您們時而溫 柔時而嚴厲的鼓勵與督促,亦是促使秀華向上的一大動力。親愛的家人,擁有您們的 愛,是秀華最大的幸福。還有亦師亦友的老大嘟嘟,謝謝您在秀華茫然時所給予的諸 多建言,在您身邊真的是獲益良多,受益匪淺。 走過不算少的年頭,經歷過不算多的風風雨雨,籃球場上嬉鬧追逐的身影、意見 分歧時的暴戾狠樣都將成為我倆心中珍貴的回憶,八年來,有歡笑當然一定有淚水, 但更多的是幸福與感激,感激你的肯定與激勵,讓我可以勇敢的突破自己向前邁進, 衷心期盼這緣分可以直至永『衡』。
iv CONTENTS Chinese abstract………...i English abstract………..ii Acknowledgement………iii Contents………... iv Table contents………...v Figure contents………...vi Introduction………1 Experimental procedures………...6 Results………..12 Discussion………....19 Conclusion………....23 References………....41 Appendix………46 Autobiography………..47
TABLE CONTENTS
TABLE I The pH value of citrate buffer upon addition of various ABA concentrations...24 TABLE II Changes in the proteins of P. campanulata true seeds following various treatments...25 TABLE III Identified proteins of P. campanulata true seeds……….………...26
FIGURE CONTENTS
Figure 1 Longitudinal section through a seed of P. campanulata………...……27
Figure 2 Germination percentage of intact P. campanulata seeds following warm or/and cold stratification………...….28
Figure 3 Tissue pHs and germination percentage of P. campanulata true seeds during incubation at 30/20℃ for 20 days……….29
Figure 4 Determination of optimal buffer concentration for the radical growth of true seeds……….……...30
Figure 5 Germination of true seeds responded to the citrate buffer with different pHs.………..………...………....31
Figure 6 Effects of different pH conditions on germination and radical growth of true seeds..………32
Figure 7 Determination of optimal buffer concentration for the radical growth of isolated embryos………33
Figure 8 Effects of different pH conditions on radical growth of isolated embryos………...………34
Figure 9 Radical growth of true seeds responded to different concentrations of exogenous ABA………...35
Figure 10 The exogenous ABA effects on germination of true seeds under the buffer treatments at different pHs.………...36
Figure 11 The exogenous ABA effects on radical growth of true seeds under the buffer treatments at different pHs..………..……….37
Figure 12 ABA contents of the fresh and treated true seeds……….38
Figure 13 Protein patterns of embryos of true seeds with different treatments……….39
Figure 14 Protein patterns of testae of true seeds with different treatments……….40
1
1. Introduction 1.1 Seed dormancy
Seed dormancy and germination, complex adaptive traits of higher plants, are influenced by a large number of environmental factors. Seed dormancy could be defined as the failure of an intact viable seed to complete germination under favorable conditions (Bewley, 1997). The role of seed dormancy is across all species to spread germination across time but in synchrony with seasonal cycle to avoid unfavorable weather to maximize competitive advantage and ensure the establishment of plants (Baskin and Baskin, 1998). Dormancy may be due to certain properties of the seed coat, mobilization of reserve components, hormone levels, or the joint action of several of these factors (Nicolas et al., 1996). And dormancy release is regulated by a combination of environmental and endogenous signals with both synergistic and competing effects. The breaking of seed dormancy is greatly affected by environmental factors such as temperature, moisture, oxygen, and light (Geneve, 2003). In addition, the balance of abscisic acid (ABA)/gibberellin (GA) levels and sensitivity is a major, but not the sole, regulator of dormancy status (Finklelstein et al. 2008). But many key factors involved in the mechanism of dormancy are still not well known. Studies with model species that have comparatively shallow dormancy have identified a number of genes involved in the regulation of dormancy and germination (Koornneef et al., 2002; Bewley, 1997). However, there have been very few molecular studies in more deeply dormant species in which moist seeds require prolonged exposure to low temperature to release dormancy and induce germination (Stephen et al., 2003).
2
1.2 Seed dormancy of Prunus campanulata Maxim
Prunus campanulata Maxim. (Taiwan cherry) is a common forest tree in the mountains of Taiwan, growing to 10 m in height. It distributes from Japan’s Ryukyu Islands and Taiwan to southern China, and is a deciduous and outstanding flowering tree with dense branching (Hiroyoshi, 1993). The trees grow on mountains at elevations between 400 and 2,500 m. They produce red, campanulate flowers from January to March and dark red fruits from April to May, making it a valuable ornamental plant. The mature fruits of P. campanulata exhibit a dark red or dark purple color (Chien et al., 2002; Chien, 1996). P. campanulata seeds are deeply dormant and known for their morphological and physiological dormancy which can be broken by a combination of warm and cold stratification (Chien et al., 2002). The dormancy mechanism in Prunus, like those of other species of deeply dormant seeds, appears to be complex.
In the first report of a proteomic analysis of dormancy and dormancy breaking in woody plant seeds (Lee et al., 2006), significant changes in P. campanulata seed proteins were observed by using 2-D gel electrophoresis before and after the breaking of seed dormancy and protein patterns of the testa (endosperm plus seed coat) significantly differed from those of the embryo. Besides, the reduction of ABA level during warm or cold stratification coincided with the decrease of dehydrin level. The prunin digestion and dehydrin disappearance provide evidence that the germination results from a combination of complex regulatory networks are possibly associated with GA and ABA. Furthermore, studies on P. campanulata seeds also show that a decrease of ABA content of the covering layers and an increase of embryonic GA4 content go along with dormancy-break
progression (Chen et al., 2007). Several studies have shown that seed covers play an important role in dormancy in genus of Prunus since embryos isolated from the seeds germinate readily (Chen et al., 2007; Mehanna et al., 1985; Zielinski, 1958; Tukey and Barrett, 1936; Giersbach and Crocker, 1932). Thus it is possible that the surrounding
3
structures (endosperm, seed coat and endocarp) impose the seed dormancy (coat-enhanced dormancy).
4
1.3 The relationship between sulfatase, pH, ABA and seed dormancy
During the progress of dormancy-breaking in P. campanulata seeds, dramatic change of sulfatase activity was found in warm/cold stratification. The sulfatase activity exhibited maximum in only warm-stratified period, and gradually decreased to a stable level in following stratification period in covering tissues, endocarp and testa (including seed coat and endosperm) (Chiu thesis 2006). There was less sulfatase activity in the embryo (including cotyledons, epicotyl, hypocotyl, and radicle), neither in warm-stratified period nor in cold stratified one. In biology systems, hydrogen sulfate (HSO3-) is released from
sulfate ester (R-OSO3-) and the reaction is catalyzed by sulfatase. The low pKa of HSO3-
(pKa=1.92) made it a candidate to supply acidic cellular environment. It was speculated
that the hydrolysis product of desulfation, hydrogen sulfate, might play an important role of modulating the cellular pH to change ABA distribution which was related to seed dormancy.
In previous literatures concerning seeds, there are several reports regarding the changes in seed tissue pH during the dormancy-breaking process and germination. During cold stratification, embryo pH decreases in dormant seeds of Crataegus gloriosa (Eckerson, 1913), Acer saccharinum (Jones, 1920), and Juniperus virginiana (Pack, 1921) and in whole seeds of Tilia americana (Rose, 1919). Embryo pH also decreased during germination (Pack, 1921; Rose, 1919; Eckerson, 1913). Furthermore, in Jerusalem artichoke tubers, the internal pH was found to be higher in dormant versus non-dormant buds (Gendraud and Lafleuriel, 1983). And the pH-dependence of the dormancy-breaking activity has been demonstrated later (Cohn et al., 1987).
There are many researches focused on the relationship between ABA and dormancy. In barley, it has been suggested that synthesis, catabolism, or removal of ABA, as well as sensitivity to ABA, concur to determine seed dormancy (Wang et al., 1995). The chemical inhibition of ABA synthesis also provokes germination of previously dormant seeds
5
(Kermode, 2005). In addition, ABA is distributed between leaf cells and free space according to pH gradients, it has been postulated in 1981 that the un-dissociated ABA is the main penetrating species (Kaiser and Hartung, 1981). The pH values of incubation medium that favor formation of the protonated species resulted in the highest germination percentages (Cohn et al., 1987). Recently, Gianinetti and Vernieri have confirmed that the pH of the incubation medium shows a relevant effect on the accumulation of exogenous ABA (Gianinetti and Vernieri, 2007). Indeed, weak acids (like ABA) have low ability to penetrate through the seed coats at pH values higher than their pKa because the movement
of the un-dissociated form of weak acids into seeds is favored.
In this study, we used P. campanulata seeds which are deeply dormant and germinate poorly before a period of dormancy breaking for determing the role of pH and ABA on dormancy-breaking and germination process. Both the individual effect and relationship of pH and ABA were considered. However, the emergence of proteomics has been inspired by the realization that the final product of a gene is inherently more complex and closer to its function than the gene itself. Proteomic tools offer other ways to analyze net-works of proteins that control important physiological reactions involved in seed dormancy-breaking/germination (Zhu et al., 2003). The effects of pH on the protein patterns of the seeds during the dormancy-breaking process were evaluated by 2-D electrophoretic analysis.
6
2. Experimental procedures 2.1 Materials
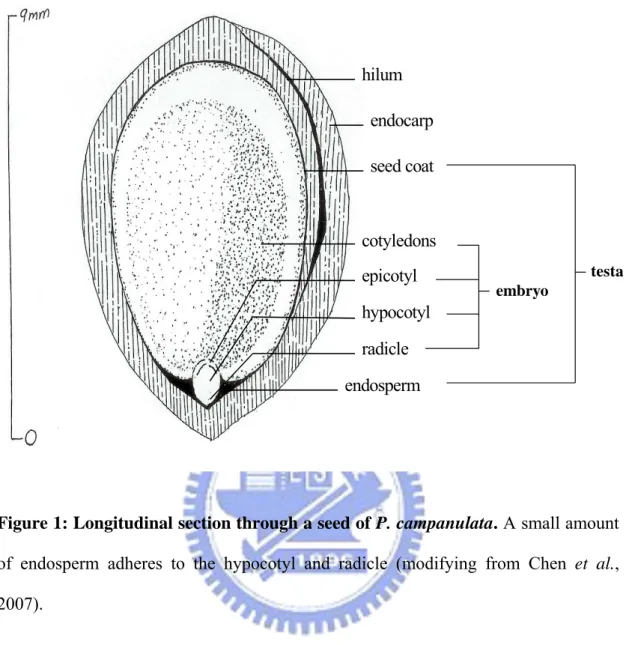
Fruits of P. campanulata were collected in the field when they were mature, i.e. dark red or dark purple in color. More than 18,000 mature fruits were collected from about five trees at Alishan, Chiayi County, central Taiwan at 2,000 m elevation in early May 2007. Seeds were extracted by removing the pulp (exocarp + mesocarp) in water, and the clean sunken seeds (with endocarp) were used for subsequent treatments. Seeds of P. campanulata are 8–9 mm in length, and consist of a large embryo and a thin seed coat adhered to a small amount of endosperm. The seed is surrounded by a hard endocarp (Figure 1). Thus, the word ‘seed’ in this study refers to the true seed plus endocarp, which is the germination unit in Prunus. Then the seeds were saved in cooling requirements at 4℃ to keep them fresh, alive and be at our disposal. By fresh seeds we mean that the seeds didn’t process any stratification.
The IPG strips (13 cm, pH 3–10, nonlinear), Pharmalytes, the protein marker kit, silver-staining kit, and glycerol were purchased from GE Healthcare (NJ, USA). The RC DC protein assay kit was obtained from BioRad (Richmond, CA, USA). Sequencing-grade modified trypsin was obtained from Promega (Mannheim, Germany). All other chemicals were the highest purity grade and were obtained commercially from Sigma-Aldrich (Deisenhofen, Germany).
7
2.2 Methods
2.2.1 The stratification processions and germination tests of intact P. campanulata seeds
2.2.1.1 The warm stratification processions
The stratification processions used in this study were prepared according to published procedures (Chien et al., 2002). Fresh P. campanulata seeds were intermingled with vaporous sphagnum in a polyethylene bag capable of sealing. In the warm stratification conditions, the sealed polyethylene bags which contained the seeds and moist sphagnum in a desired period were kept alternative between at 30℃ in the fluorescent light at the irradiance of 80–100 μmol/m2/s for 12 hours and 20℃ in the dark for 12 hours. The bags were opened for checking moisture weekly.
2.2.1.2 The cold stratification processions
In the cold stratification conditions, the sealed polyethylene bags which contained the seeds and moist sphagnum were kept at 4℃ in the dark in a desired period. The bags were opened for checking moisture weekly.
2.2.1.3 The germination tests of intact P. campanulata seeds in alternative stratifications or warm plus cold stratifications
The stratified seeds were placed on moist sphagnum in sealable polyethylene bags and germinated at 30/20℃ (day/night) with 12 hours of fluorescent light (80–100 μmol/m2/s) during the high temperature period of the regime for 12 weeks. Germination, judged by radical protrusion of at least 2 mm, was recorded weekly, and results were expressed as percent germination.
8
2.2.2 The tests of germination and radical growth of true seeds and isolated embryos after various pH buffer treatments
2.2.2.1 The tests of germination and radical growth of true seeds (seeds without endocarps)
The true seeds, the endocarps were removed from intact seeds, were cut down to half for the seed radicles to contact the liquids of Whatmen No. 1 filter paper (90 mm diameter) in plastic Petri dishes (90 mm diameter). Thirty true seeds were sowed on plastic Petri dishes with two sheets of filter paper and 4 ml of deionzation (DI) water or various pH buffers. The test was processed in the incubator at 30/20℃ (day/night) with 12 hours of fluorescent light (80–100 μmol/m2/s) during the high temperature period of the regime for 10 days. In addition, 2 ml DI water or various pH buffers was added to the dishes at 3 days after sowing (DAS) (the sowing day is 0 day), then the germination and the length of radicles were measured at 10 days by tweezers and section paper (1 mm, AI TE BRAND). The same experimental procedures as described above were used to determine sensitivity to exogenous ABA and the pH effects on ABA movement.
2.2.2.2 The tests of radical growth of isolated embryos (seeds without endocarps and testae)
The true seeds were soaked in the DI water or variant pH buffers for 1 hour. Then, the isolated embryos, the testae (endosperm plus seed coat) were removed from the true seeds, were cut down to half for the radicles to contact the liquids of the filter paper in Petri dishes. Thirty isolated embryos were sowed on plastic Petri dishes with two sheets of filter paper and 4 ml of DI water or variant pH buffers. In addition, 2 ml DI water or variant pH buffers were added to the dishes at 3 DAS (the sowing day was 0 day) and the radical length was measured at 5 days by tweezers and section paper.
9
2.2.3 Homogenate pH measurement of true seeds during dormancy-breaking and germination
For each pH determination, the incubated seeds were separated into two parts, testa and embryo, by tweezers. Testae and embryos were weighed, and homogenized with PRO 250 homogenizer (PRO Scientific, Oxford, CT, USA) in 3 and 5 ml DI water, respectively. The pH value of each tissue homogenate was measured immediately (to avoid alkalinization by uptake of atmospheric carbon dioxide) with a pH electrode. The pH value was recorded when the electrode registered a stable pH (<0.01 units min-1) which was generally achieved within 3 or 5 min.
2.2.4 2-D gel electrophoresis (2-DE) and protein identification by mass spectroscopy (MS)
2.2.4.1 Sample preparation and 2-DE
True seeds were separated into two parts, testa and embryo, by tweezers. Extracts were homogenized with the homogenizer containing 125 mM sucrose, 10% (v/v) glycerol, 1 mM dithiothreitol (DTT), 1 mM (ethylenedinitrilo)tetraacetic acid (EDTA), 1 mM phenylmethylsulfonyl fluoride (PMSF), and 10 mM Tris-HCl at pH 7.4. The suspension was centrifuged for 30 min at 10,000 g at 4℃, and the resultant supernatant liquid was centrifuged for 120 min at 22,000 g at 4℃. The final supernatant liquid was used for 2-DE. Protein concentrations of extracts were estimated on the basis of an RC DC protein assay kit from BioRad (Richmond, CA, USA). 2-DE was performed by a commercially available Ettan IPGphor IEF system and Hoefer SE600 Ruby (gel size 13 cm × 15 cm) from GE Healthcare (Uppsala, Sweden). Proteins from the various extracts were separated using gel strips and formed an immobilized nonlinear pH gradient from 3 to 10 (Immobiline DryStrip, pH 3–10 nonlinear, 13 cm; GE Healthcare). Analytical IPG strips were rehydrated for 12 hours at 20℃ with 250 μL of the rehydration buffer including 200 μg of
10
sample proteins. Isoelectric focusing (IEF) was performed at 20℃ in the Ettan IPGphor system (GE Healthcare) for 1 hour at 500 V, 1 hour at 1,000 V, 1 hour at 4,000 V, and 3 hours at 8,000 V. Prior to the second dimension, the strips were equilibrated for 2 × 15 min in equilibration solution containing 6 M urea, 30% (v/v) glycerol, 2% (w/v) SDS, 0.5 M Tris-HCl (pH 8.8). DTT (1% w/v) was added to the first equilibration solution and 2.5% (w/v) iodoacetamide was added to the second one. For the second dimension, the strips were transferred onto sodium dodecyl sulfate (SDS) polyacrylamide gels (12.5%) with a run of 50 mA per gel for 4–5 hours at 4℃.
2.2.4.2 Protein staining and analysis of 2-DE gels
After electrophoresis, proteins were visualized by a modified silver-staining kit (Yan et al., 2000). Digital images of the gels were obtained by using an ImageScanner and were analyzed using ImageMaster 2D elite software (GE Healthcare). The spots were detected and the background was subtracted (mode: average on boundary), and the 2-DE gels were aligned and matched. A quantitative determination of the spot volumes was performed (mode: total spot volume normalization). Specific spots were described during different treatments when their volumes significantly differed (at least ten-fold in relative abundance). The interesting proteins were identified by matrix-assisted laser desorption inoization-time of flight mass spectrometry (MALDI-TOF-MS) or electrospray ionisation tandem mass spectrometry (ESI-MS/MS) analysis.
2.2.4.3 Protein identification by MS
For MS analysis, protein spots were excised from the gel and digested with trypsin according to published procedures (Shevchenko et al., 1996). Proteins were identified by searching the protein databases (Swiss-Prot, MSDB and NCBInr) using MASCOT (http://www.matrixscience.com). To denote a protein as unambiguously identified, the
11
Mowse scoring algorithms were used. Only proteins whose score exceeded the significance threshold (probability, which can be ambiguous if smaller than 0.05) were discussed.
2.2.5 The measurement of ABA contents of true seeds
The treatment of true seed was described above and the gas chromatography-mass spectrometry (GC-MS-SIM) analysis of ABA was according to the procedures of Chen et al. (2007). The high performance liquid chromatography (HPLC) fractions were dried using a SpeedVac and derivatized by adding ethereal diazomethane, then dried with nitrogen. ABA was further trimethysilylated with 50 ml pyridine plus 100 ml of N,N’-bis(trimethylsilyl)trifluoroacetamide (BSTFA) with 1% trimethylchlorosilane (TMCS) (Macherey-Nagel, USA) at 90℃ for 30 min. The derivatized samples were analysed using a HP 6890 GC and 5973 MSD with a DB-1 capillary column (30 m × 0.25 mm internal diameter, 0.25 μm film thickness, J&W Scientific, USA). Operating conditions of GC-MS were similar to those described by Nakayama et al. (2001). The carrier gas was helium at a flow rate of 1 ml min-1, and the interface temperature of the ion source was 250℃. The MS source temperature was 200 , and the electron energy was 70 ℃ eV. The split/splitless injector was used in the splitless mode at 250 . The oven ℃ temperature was programmed to begin at 60 for 2 min, then raised to 210 at a rate of ℃ ℃ 30 min℃ -1 and to 280 at a rate of 2℃ ℃ min-1, where it remained for 5 min. The sample was dissolved in 10 ml of hexane, and 1 ml of it was injected. The m/z ratios of 190/194 was used for quantification of ABA.
12
3. Results
3.1 Germination percentages of intact P. campanulata seeds following various stratifications
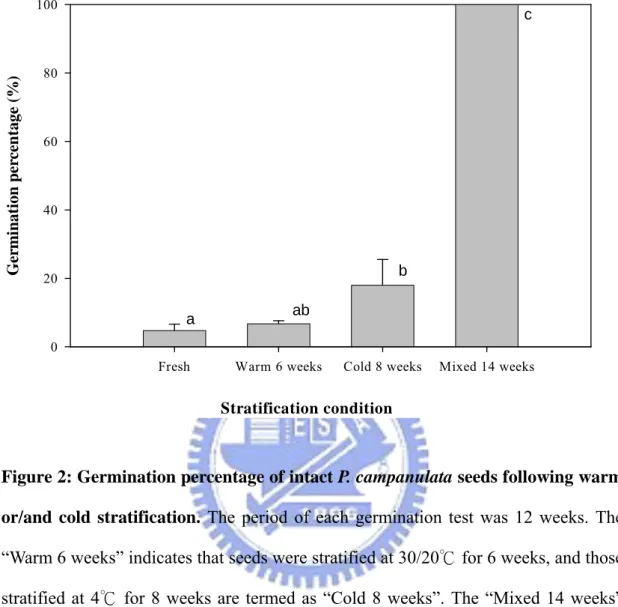
As shown in Figure 2, the germination percentage of fresh intact seeds from mature fruits was less than 5%, and the seeds treated with warm stratification for 6 weeks merely germinated to 6.7%. Therefore, warm stratification did not affect germination of P. campanulata seeds directly. Seeds that were treated with cold stratification for 8 weeks germinated to 18%, while those treated with warm stratification for 6 weeks followed by cold stratification for 8 weeks germinated to 100%. Thus, the dormancy of intact P. campanulata seeds could be completely broken after 6 weeks warm stratification plus 8 weeks cold stratification. The stratification of either warm or cold alone could not completely break dormancy, although the effect of cold stratification was greater than that of warm stratification.
13
3.2 The pH effects on dormancy-breaking and germination of P. campanulata seeds 3.2.1 The pH effects on true seeds
3.2.1.1 Testae and embryos acidification during dormancy-breaking and subsequent germination
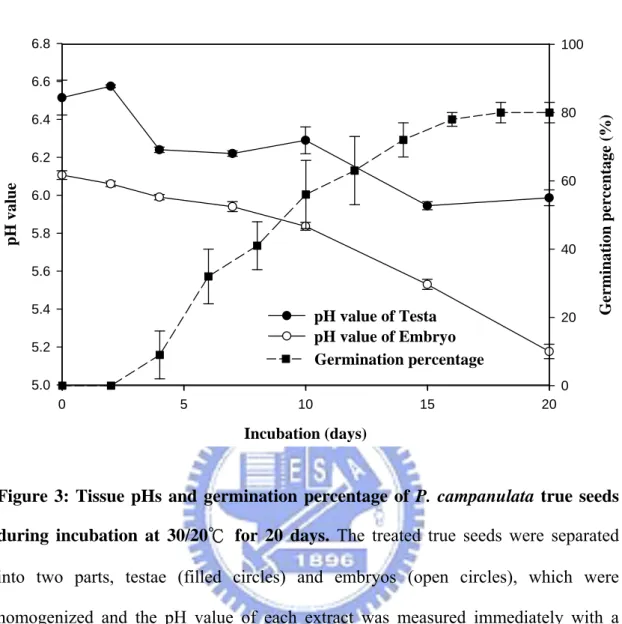
The dormancy of P. campanulata true seeds that were cut a half and incubated with DI water at 30/20℃ was broken, and the treated seeds began to germinate at 3 days after sowing. The germination percentages were gradually raised and reached 80% finally (filled square, Figure 3). The pH values of testae (filled circle) and embryos (open circle) both declined during dormancy-breaking and germination (Figure 3). In addition, the pH values of the embryos changed significantly, and the pH difference between testae and embryos became greater with time.
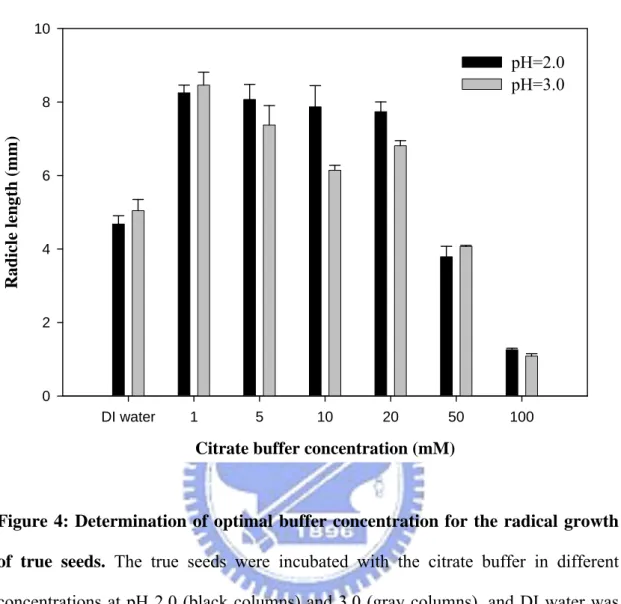
3.2.1.2 Selection of optimal buffer concentration for radical growth of true seeds Figure 4 showed that ionic strength effects of citrate buffer on the radical growth of
true seeds. The radical growth of true seeds decreased with the increase of the buffer concentration. In high concentration, poor radical growth was found and it was almost suppressed as the buffer concentration up to 100 mM. There were similar tendency in different pH buffer treatments (black and gray columns), and 5 mM buffer concentration was determined as the optimal condition and used it for subsequent pH tests.
3.2.1.3 Response of germination and radical growth of true seeds to different pHs
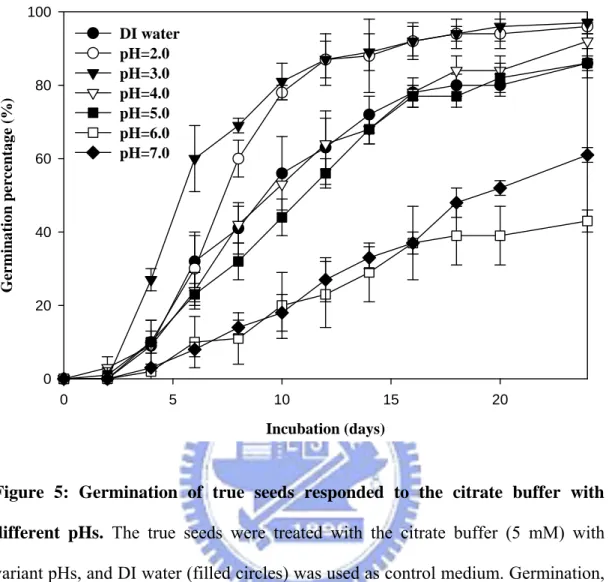
The true seeds were treated with citrate buffer (5 mM) at various pHs (from pH 2.0 to 7.0), and the obvious differences in germination percentages were found (Figure 5). The seeds which were incubated with low pH values, pH 2.0 (open circle) and pH 3.0 (filled triangle), began to germinate at 3 DAS, and the germination percentages were up to near 100% till the 24th day. Comparatively, the seeds germinated at the 5th day and the
14
germination percentages merely reached 60% at pH 6.0 (open square) and pH 7.0 (filled diamond) as shown in Figure 5. The germination tendencies of true seeds after the buffer treatments, pH 4.0 (open triangle) and pH 5.0 (filled square), were similar to DI water control. The results revealed that true seeds which were incubated with low pH buffer germinated faster and reached higher germination percentages than those in close-to-neutral pH conditions.
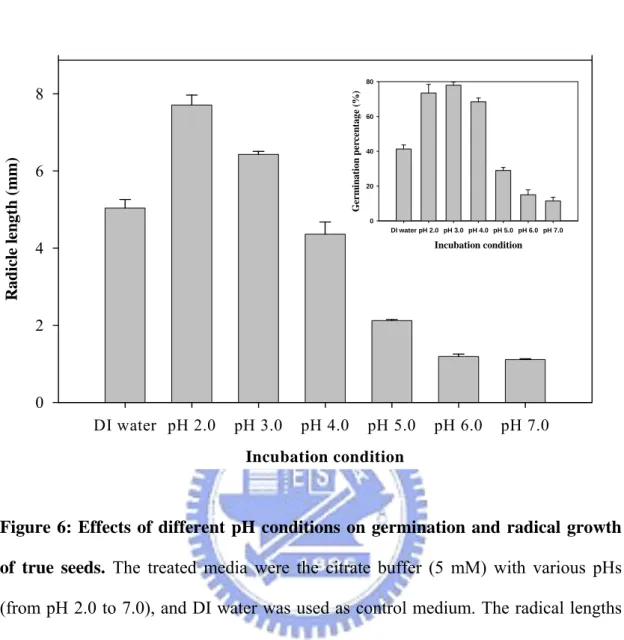
Furthermore, the pH effect on dormancy-breaking and germination through the observation of the radical growth of true seeds in different pH buffers were identified. The seeds were treated with citrate buffer (5 mM) at pH values from 2.0 to 7.0, and the radical growth was better in low pH environments than neutral ones as shown in Figure 6. The germination percentages (Figure 6, inset) of true seeds were recorded at the 10th day, and the phenomenon which was similar to radical growth under variant pH incubations was found.
3.2.2 The pH effects on isolated embryos
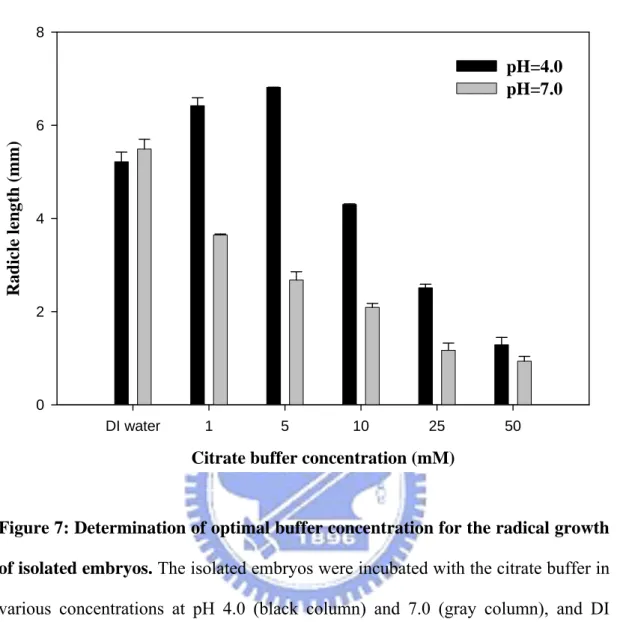
3.2.2.1 Selection of optimal buffer concentration for radical growth of isolated embryos
As shown in Figure 7, the radical growth of embryos was affected by citrate buffer concentration. The radicles of isolated embryos decreased with the increase of the buffer concentration. In high concentration, poor radical growth was found and it was almost suppressed as the buffer concentration up to 50 mM. There were similar tendencies in different pH buffer treatments (black and gray columns), and 5 mM buffer concentration was determined as the optimum and used it for subsequent pH tests.
3.2.2.2 Radical growth of isolated embryos responded to different pHs.
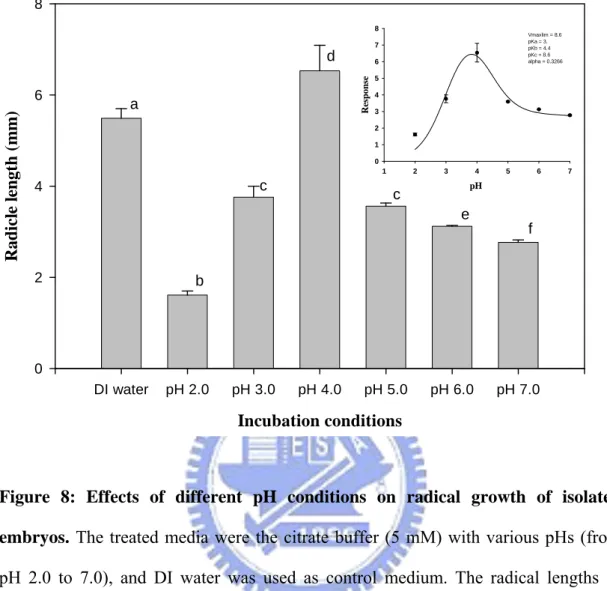
15
2.0 to 7.0) in the period of 5 days, and obvious differences in radical growth were observed (Figure 8). The optimal condition of radical growth of isolated embryos was the pH 4.0, which allowed the radical length grow up to 6.5 mm compared with 5.5 mm in DI water control. In low pH environments, pH 2.0 and pH 3.0, the radical growth of isolated embryos was inhibited appreciably and the radical lengths were only 1.6 and 3.7 mm, respectively. The seed radicles also grew poorly with the increase of pH from 5.0 to 7.0. According to the phenomenon as described above and previous literatures, we speculated that ABA might be involved in the pH effects on the radical growth of the seeds by its dissociation under different pH conditions. The pKa value which was evaluated using
nonlinear regression with SigmaPlot 2001, Version 7.0 and Enzyme Kinetics Module, Version 1.1 (SPSS Inc., Chicago, IL) was 4.4, and it was considerably closed to the theoretical value of ABA (pKa=4.8) as shown in Figure 8 (inset).
16
3.3 The ABA effects on dormancy-breaking and germination of P. campanulata true seeds under various pH environments
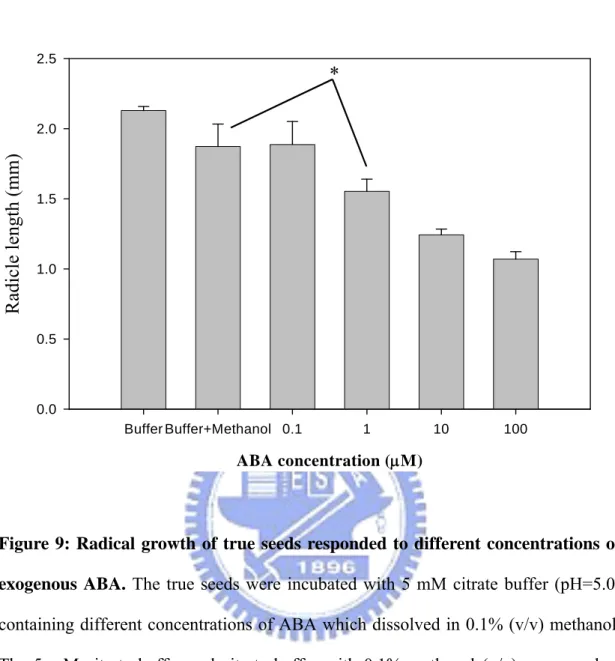
3.3.1 Determination of the concentration of ABA which could affect radical growth of true seeds
Constant pH values of citrate buffers (5 mM) remained upon the addition of ABA. As shown in TABLE I, the citrate buffers exhibited their buffered capacities to maintain the constant pH values in the presence of ABA, even up to 100 μM. The true seeds were then incubated with various ABA concentrations from 0.1 to 100 μM in citrate buffer (5 mM) at pH 5.0, and it was worth to note that there was a significant decrease in 1μM ABA compared with the 0.1% methanol control (Figure 9). Thus, 1 μM ABA concentration was selected as the treated condition and used it for subsequent pH tests.
3.3.2 The sensitivity of germination and radical growth of true seeds to exogenous ABA under various pH treatments
The minimal level of ABA (1 μM) that in the previous trial showed the least effect was used in this experiment. Significant effects of exogenous ABA on the germination (Figure 10) and radical growth (Figure 11) in low pH environments (from pH 2.0 to 4.0) were observed. On the contrary, the abilities of ABA to suppress germination and radical growth were strongly reduced at near neutral pH values.
3.3.3 Determination of ABA contents of the fresh and treated true seeds
We utilized GC-MS to measure ABA concentrations of true seeds under the treatments of DI water or citrate buffer with different pH values (pH 2.0 and 7.0) to verify the pH effects on the penetration of ABA. The control, fresh seed, contained a great amount of ABA (3,960 pg seed-1) which was approximately 3.6-fold higher than that of treated seeds. Comparison of the true seeds which were incubated with DI water for 2 and
17
10 days, decreased ABA contents with the dormancy-breaking or germination processes were observed (Figure 12). In order to validate the pH effects on seed dormancy-breaking, ABA concentrations were measured in true seeds which were un-germinated after 2-day incubation of different pH buffers. However, the present data indicated that there was no significant difference in ABA concentration between pH 2.0 and 7.0.
18
3.4 The pH effects on protein expressions of the dormant true seeds
Proteins were extracted from the various true seed samples (freshly harvested, DI water treated for 2 days, 5 mM citrate buffer (pH 2.0) treated for 2 days, and 5 mM citrate buffer (pH 7.0) treated for 2 days) and analyzed by 2-DE as described in Section 2.2.4. As shown in Figures 13 (embryos) and 14 (testae), there were some protein spots with significant changes (of at least ten-fold) which were obtained by comparing protein patterns of fresh seeds and treated seeds, and 45 and 70 dramatic changes of embryos and testae in abundances were observed, respectively. These protein spots were summarized in
TABLE II. A total of 115 interesting spot proteins were excised from preparative 2-DE
gels. After trypsin in-gel digestion, the proteins were analyzed by MALDI-TOF-MS or ESI-MS/MS. Among the changed proteins, three spots showed significant matches. MS data including peptide masses and identified amino acid sequences for spots of interest were summarized in TABLE III. Protein No. 8, 21, and 25 were identified as Prunus serotina (R)-mandelonitrile lyase 1 precursor, Vitis vinifera hypothetical protein, and Prunus dulcis prunin, respectively. It seemed important to discuss the functions of the identified proteins and related metabolic pathways involved in dormancy-breaking.
19
4. Discussion
In the germination experiments (Figure 2), low germination percentages of freshly harvested intact seeds of P. campanulata were observed, and most of seeds did not germinate under either warm or cold stratification, but warm plus cold stratification increased seed germination significantly. It indicated that a combination of warm and cold stratification is necessary for releasing dormancy completely. According to the previous researches (Chen et al., 2007), cold stratification alone stimulated a high percentage of the seeds to germinate, up to 70% for 8 weeks treatments, and it has been widely used for breaking seed dormancy and promoting the maximum percentage and rate of germination (International Seed Testing Association, 1999; Schopmeyer, 1974). However, our results (Figure 2) showed that small amounts of seeds germinated after cold stratification, and the germination percentage was only 18%. The differences in the germination of seeds in response to stratification were linked to fruit maturity which depended upon the year and the location of the seed harvested (Chen et al., 2007). Nevertheless, we could confirm that the seeds were alive for subsequent experiments because their germination percentages reached 100% through warm stratification for 6 weeks plus cold stratification for 8 weeks.
In this research, the true seeds which still kept their dormancy characteristics (Chen et al., 2007) were used as the experimental materials for studying the mechanism of P. campanulata seed dormancy. With red rice and several other species, embryo pH decreased during dormancy-breaking process and subsequent germination (Footitt and Cohn, 1992). In true seeds of P. campanulata, pH values of testae and embryos declined as dormancy-breaking and germination were also found (Figure 3). The pH change of embryos was more rapid than it of testae. In 1992, Footitt and Cohn mentioned that multicellular systems of the plant and animal kingdoms seem to be a decrease in internal pH upon activation. Our results also presented that the tissues, testae and embryos, acidification was a prerequisite for the termination of dormancy.
20
We applied different pH environments with citrate buffer to induce the dormancy-breaking of the seeds. Figure 5 indicated that true seeds which were incubated with low pH buffers germinated faster and reached higher germination percentages than those in near neutral pH conditions. The radical growth of the true seeds was further examined to understand pH effects on other plant physiological phenomenon based on the results of the pH effects on dormancy-breaking and germination. As shown in Figure 6, the radicles of true seeds favored to grow in low pH environments instead of near neutral ones. Previous studies on seed dormancy of red rice showed that dormancy-breaking activity was pH-dependent. Incubation medium pH values that favored formation of the protonated species resulted in the highest germination percentages (Cohn et al., 1987). Consequently, low pH treatments effectively promote seed dormancy-breaking and induce its radical growth.
We also used different type of the seeds, isolated embryos, to identify the pH effects on P. campanulata seeds. As shown in Figure 8, the seed radicles favored to grow in weak acidic environments, and there were similar characteristics in true seeds. It should be noted that pH 4.0 was the best condition for radical growth of isolated embryos, possibly resulting from the pH effects on ABA movement according to the previous studies (Kaiser and Hartung, 1981). In this situation (pH < pKa), un-dissociated species would be the
major forms, and the main penetrating species between the cells and free space has been proven as un-dissociated ABA (Kaiser and Hartung, 1981). The pKa which was calculated
as described in Section 3.2.2.2 was 4.4, and it was considerably closed to the theoretical value of ABA (pKa=4.8) as shown in Figure 8 (inset). Therefore, the seed radicles grew
well in low pH values (pH≦4.0) because the un-dissociated ABA was easy to penetrate the tissues and move outside. But, poor radical growth of isolated embryos in acidic conditions, pH 2.0 and 3.0, was observed, and we considered that isolated embryos lacked testae protections resulted in this phenomenon. The radical growth of isolated embryos
21
(Figure 7) which was more sensitive to high concentration of the buffers than true seeds (Figure 4), and this may be due to isolated embryos lacked the protections of testae.
Figures 10 and 11 showed the effects of exogenous ABA under various pH
conditions on the progress of dormancy-breaking and germination through the observation of seed germination and radical growth. Incubation in the presence of exogenous ABA obviously decreased the germination percentage (Figure 10) and radical length (Figure 11) of the true seeds at low pH values (from pH 2.0 to 4.0). The low pH of the incubation medium might produce a relevant effect on the accumulation of exogenous ABA. At the more acidic pH, the greater total amount of exogenous ABA entered the seed and favored to accumulate into the embryo and endosperm even above the levels of the untreated control in red rice (Gianinetti and Vernieri, 2007). By contrast, the ability of ABA to suppress germination and radical growth was strongly reduced at near neutral pH values. Indeed, weak acids (like ABA) have low ability to penetrate through the seed coats at pH values higher than their pKa because the movement of the un-dissociated form of weak
acids into seeds is favored (Cohn et al., 1987). In ABA quantification experiments (Figure
12), we found the difference between dormant and dormancy-breaking seeds which
incubated with DI water for 2 and 10 days, respectively. There were similar ABA concentrations in dormant seeds even through DI water or different pH buffer treatments. Thus, ABA should not be the key modulator and there might be other regulatory factors involved in pH effects on the dormancy-breaking of true seeds.
Studies of plant proteomic analyses used so far have been reported (Lee et al., 2006; Cánovas et al., 2004). Proteomic tools offer the ways to analyze net-works of proteins that control important physiological reactions involved in seed dormancy-breaking/germination. As results, the pH treatments effectively affected the dormancy-breaking of P. campanulata true seeds, and the change in proteins following this treatment might be important factors in seed dormancy. According to the result as shown in Figure 5, true
22
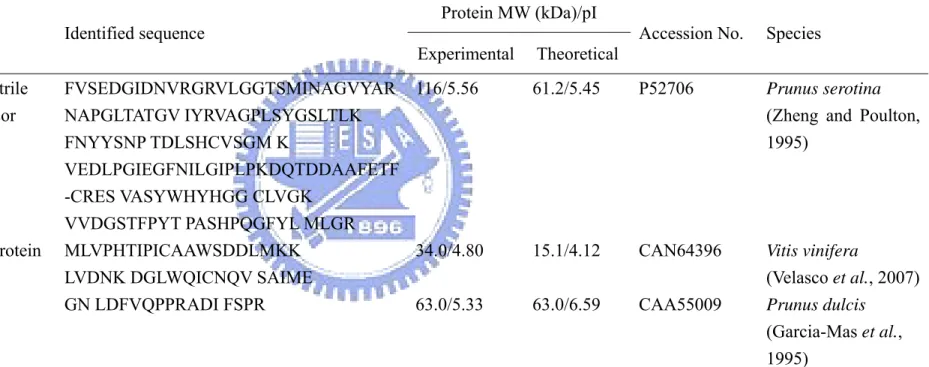
seeds were treated for 2 days to study the pH effects on the process of the seed dormancy-breaking. The labeled protein spots identified by MS were listed in TABLE III. In embryos, three proteins which exhibited dramatic changes after DI water or different pH buffer treatments were identified as (R)-mandelonitrile lyase 1 precursor and prunin which belonged to the genus Prunus except hypothetical protein (Garcia-Mas et al., 1995).
Amygdalin is hydrolyzed and generated hydrogen cyanide gas (HCN) and benzaldehyde by several enzymes including (R)-mandelonitrile lyase (MDL) (Zhou et al., 2002). Prunus species (Rosaceae) are a rich source of the cyanogenic diglucoside (R)-amygdalin [the β-gentiobioside of (R)-mandelonitrile] and its catabolic enzymes which accumulate in leaves and immature fruits (Zheng and Poulton, 1995). Also, changes in proteins probably indicate biochemical activities of the seed after the uptake of water (Lee et al., 2006). Here, (R)-mandelonitrile lyase 1 precursor exhibited dramatic decrease after DI water treatment because the dormancy of true seeds might be broken.
In embryos, the other identified proteins were Vitis vinifera hypothetical protein whose function was unknown at present (Velasco et al., 2007), and Prunus dulcis prunin, was known as globulin of the genus Prunus, which comprise the main family of storage proteins synthesized in seeds during embryogenesis (Garcia-Mas et al., 1995). The protein which was identified as prunin exhibited dramatic changes after the treatment at pH 2.0 that accelerated the dormancy-breaking of true seeds. Previous articles indicated that the fragmentation of prunin may possibly be used as an index of seed germination (Lee et al., 2006), and the GA contents of P. campanulata seeds increased during dormancy-breaking process (Chen et al., 2007). In other words, degradation of prunin which occurred during dormancy-breaking is probably related to GA induction. It will be the important evidences to study the effect of the degradation of prunin during the breaking of seed dormancy.
23
5. Conclusion
P. campanulata seeds were suitable experimental materials for studying the mechanism of dormancy-breaking resulted from their dormancy characteristics. We confirmed that low pH treatments effectively promoted seed dormancy-breaking and induce radical growth. Results of our analysis provided evidences of the involvement of MDL and prunin in response to the dormancy-breaking of true seeds. The study of the pH effects on the proteomes of tree seeds will contribute to an understanding of the molecular basis of seed dormancy.
TABLE I
The pH values of citrate buffer upon addition of various ABAa concentrations
ABA concentrationb (μM) 5 mM citrate buffer pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 - 2.12±0.015 3.06±0.007 4.01±0.011 5.05±0.049 6.04±0.026 7.09±0.014 1 2.13±0.010 3.06±0.020 4.01±0.035 5.08±0.030 6.07±0 7.00±0.042 10 2.14±0.014 3.05±0.023 4.02±0.021 5.06±0.056 6.06±0.020 7.00±0.015 100 2.11±0.017 3.03±0.025 4.00±0.026 5.01±0.011 6.01±0.015 6.99±0.005
aThe pH value of each solution was measured immediately (to avoid alkalinization by uptake of atmospheric carbon dioxide) with a normal pH
electrode. And the pH value was recorded when the electrode registered a stable pH (<0.01 units min-1); this was generally achieved between 3
and 5 min. Values were means ± SD of three repeats independently.
bABA was dissolved in methanol whose final concentration was 0.1% (v/v).
TABLE II
Changes in P. campanulata true seed proteins following various treatmentsa
Treatments
Increased protein spots numbers Decreased protein spots numbers
Embryo Testa Embryo Testa
DI water 4, 6, 9, 12, 14, 15, 16, 20, 21 47, 50, 52, 55, 57, 63, 66, 68, 69, 72, 75, 76, 77 3, 8, 10, 11, 13, 17, 18, 19 46, 48, 49, 51, 53, 54, 56, 58, 59, 60, 61, 62, 64, 65, 67, 70, 71, 73, 74 pH 2.0 24, 28, 29, 31, 32, 33, 34 47, 82, 86, 89, 92, 94 23, 25, 26, 27, 30, 35, 36 53, 58, 78, 79, 80, 81, 83, 84, 85, 87, 88, 91, 93, 95, 96, 97 pH 7.0 1, 37, 45 100, 101, 103, 106, 108, 109, 110, 111, 112, 113, 114 2, 5, 7, 19, 22, 38, 39, 40, 41, 42, 43, 44 48, 78, 81, 90, 98, 99, 102, 104, 105, 107, 115
aSpot changes were obtained by comparing protein patterns of fresh seeds and treated seeds (DI water, 5 mM citrate buffer (pH 2.0), and 5 mM
citrate buffer (pH 7.0) for 2 days).
26
TABLE III
Identified proteins of P. campanulata true seedsa
Spot No. Protein name Identified sequence
Protein MW (kDa)/pI
Accession No. Species Experimental Theoretical 8 (R)-mandelonitrile lyase 1 precursor FVSEDGIDNVRGRVLGGTSMINAGVYAR NAPGLTATGV IYRVAGPLSYGSLTLK FNYYSNP TDLSHCVSGM K VEDLPGIEGFNILGIPLPKDQTDDAAFETF -CRES VASYWHYHGG CLVGK VVDGSTFPYT PASHPQGFYL MLGR 116/5.56 61.2/5.45 P52706 Prunus serotina
(Zheng and Poulton, 1995)
21 Hypothetical protein MLVPHTIPICAAWSDDLMKK LVDNK DGLWQICNQV SAIME
34.0/4.80 15.1/4.12 CAN64396 Vitis vinifera
(Velasco et al., 2007) 25 Prunin GN LDFVQPPRADI FSPR 63.0/5.33 63.0/6.59 CAA55009 Prunus dulcis
(Garcia-Mas et al., 1995)
aProtein spots labeled with the same number in different gels indicate the same protein by analysis of ImageMaster 2D elite software. Except for
spot No. 8, 21, and 25, other spots showed no good matches. Spot No. 8 and 21 were identified by MALDI-TOF-MS; spots No. 25 was identified by ESI-MS/MS.
hilum endocarp seed coat cotyledons epicotyl hypocotyl endosperm radicle embryo testa
Figure 1: Longitudinal section through a seed of P. campanulata. A small amount
of endosperm adheres to the hypocotyl and radicle (modifying from Chen et al., 2007).
Stratification condition
Fresh Warm 6 weeks Cold 8 weeks Mixed 14 weeks
Ger m in at ion per cen tage ( % ) 0 20 40 60 80 100 a ab b c
Figure 2: Germination percentage of intact P. campanulata seeds following warm or/and cold stratification. The period of each germination test was 12 weeks. The
“Warm 6 weeks” indicates that seeds were stratified at 30/20℃ for 6 weeks, and those stratified at 4℃ for 8 weeks are termed as “Cold 8 weeks”. The “Mixed 14 weeks” indicates that the seeds were stratified at 30/20℃ for 6 weeks followed by 4 for 8 ℃ weeks. Columns with the same letter were not significantly different (P≦0.05).
Incubation (days) 0 5 10 15 20 pH v a lue 5.0 5.2 5.4 5.6 5.8 6.0 6.2 6.4 6.6 6.8 pH value of Testa pH value of Embryo Ge rmination perc ent age ( % ) 0 20 40 60 80 100 Germination percentage
Figure 3: Tissue pHs and germination percentage of P. campanulata true seeds during incubation at 30/20℃ for 20 days. The treated true seeds were separated
into two parts, testae (filled circles) and embryos (open circles), which were homogenized and the pH value of each extract was measured immediately with a normal pH electrode. The germination (filled squares) was recorded every two days, and results were expressed as percent germination.
Citrate buffer concentration (mM) DI water 1 5 10 20 50 100 Ra d icle le ng th (m m) 0 2 4 6 8 10 pH=2.0 pH=3.0
Figure 4: Determination of optimal buffer concentration for the radical growth of true seeds. The true seeds were incubated with the citrate buffer in different
concentrations at pH 2.0 (black columns) and 3.0 (gray columns), and DI water was used as control medium. The radical lengths of true seeds were recorded at 10 days after sowing (DAS).
Incubation (days) 0 5 10 15 20 Germina tio n percentag e (% ) 0 20 40 60 80 100 DI water pH=2.0 pH=3.0 pH=4.0 pH=5.0 pH=6.0 pH=7.0
Figure 5: Germination of true seeds responded to the citrate buffer with different pHs. The true seeds were treated with the citrate buffer (5 mM) with
variant pHs, and DI water (filled circles) was used as control medium. Germination, judged by radical protrusion of at least 2 mm, was recorded every two days.
Incubation condition DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 Ra d icl e l en g th ( mm) 0 2 4 6 8 Incubation condition DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 G erm ina tion per cen tag e (% ) 0 20 40 60 80
Figure 6: Effects of different pH conditions on germination and radical growth of true seeds. The treated media were the citrate buffer (5 mM) with various pHs
(from pH 2.0 to 7.0), and DI water was used as control medium. The radical lengths and germination percentages (inset) of true seeds were recorded at the 10th day after sowing.
Citrate buffer concentration (mM) DI water 1 5 10 25 50 Radicl e le ngth (mm) 0 2 4 6 8 pH=4.0 pH=7.0
Figure 7: Determination of optimal buffer concentration for the radical growth of isolated embryos. The isolated embryos were incubated with the citrate buffer in
various concentrations at pH 4.0 (black column) and 7.0 (gray column), and DI water was used as control medium. The radical lengths of isolated embryos were collected at 5 DAS.
Incubation conditions DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 Rad icl e l en gt h ( m m ) 0 2 4 6 8 pH 1 2 3 4 5 6 7 Re sp o n se 0 1 2 3 4 5 6 7 8 Vmaxlim = 8.6 pKa = 3. pKb = 4.4 pKc = 8.6 alpha = 0.3266 a b c c d f e
Figure 8: Effects of different pH conditions on radical growth of isolated embryos. The treated media were the citrate buffer (5 mM) with various pHs (from
pH 2.0 to 7.0), and DI water was used as control medium. The radical lengths of isolated embryos were recorded at 5 DAS by tweezers and section paper. Columns with the same letter were not significantly different (P≦0.05). The pKa (inset) was
calculated using nonlinear regression with SigmaPlot 2001, Version 7.0 and Enzyme Kinetics Module, Version 1.1 (SPSS Inc., Chicago, IL).
ABA concentration (μM) Buffer Buffer+Methanol 0.1 1 10 100 Radicle length (mm) 0.0 0.5 1.0 1.5 2.0 2.5
*
Figure 9: Radical growth of true seeds responded to different concentrations of exogenous ABA. The true seeds were incubated with 5 mM citrate buffer (pH=5.0)
containing different concentrations of ABA which dissolved in 0.1% (v/v) methanol. The 5 mM citrate buffer and citrate buffer with 0.1% methanol (v/v) were used as control solutions. The radical lengths of true seeds were recorded at the 10th day after sowing. The differences of the radical growth of true seeds were analyzed by t-test using SigmaPlot 2001, Version 7.0 (SPSS Inc., Chicago, IL), and the star symbol indicated P<0.05.
pH values of 5 mM citrate buffers DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 Ge rm ina ti n pe rc ent a ge (% ) 0 20 40 60 80 w/o ABA w/ 1 μM ABA
pH values of 5 mM citrate buffers
DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 G erm in a tio n pe rce nta g e (% ) 0 10 20 30 40 50 Difference
Figure 10: The exogenous ABA effects on germination of true seeds under the buffer treatments at different pHs. The seeds were treated with 5 mM citrate buffer
from pH 2.0 to 7.0 either alone or in the presence of 1 μM ABA which were dissolved in 0.1% methanol (v/v). In this experiment, DI water was used as a control solution. The germination percentages of true seeds were recorded at 10 DAS. The differences of the germination percentages between the seeds which incubated with (w/, gray column) and without (w/o, black column) ABA in the same pH environments were shown as inset.
pH values of 5 mM citrate buffers DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 Rad icl e l en g th ( m m ) 0 2 4 6 8 w/o ABA w/ 1 μM ABA
pH values of 5 mM citrate buffers
DI water pH 2.0 pH 3.0 pH 4.0 pH 5.0 pH 6.0 pH 7.0 Radicl e le ng th (m m ) 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 Difference
Figure 11: The exogenous ABA effects on radical growth of true seeds under the buffer treatments at different pHs. The seeds were treated by 5 mM citrate
buffer from pH 2.0 to 7.0 either alone or in the presence of 1 μM ABA which were dissolved in 0.1% methanol (v/v). In this experiment, DI water was used as a control solution. The radical lengths of true seeds were recorded at 10 DAS. The differences of the radical length between the seeds which incubated with (w/, gray column) and without (w/o, black column) ABA in the same pH environments were shown as inset.
ABA concentrations (pg seed-1) 0 1000 2000 3000 4000 T reate d conditions pH 7.0 (2 days) pH 2.0 (2 days) DIW (10 days) DIW (2 days) Fresh
Figure 12: ABA contents of the fresh and treated true seeds. The seeds were
incubated with either DI water for 2 and 10 days or citrate buffers (5 mM) for 2 days. The ABA concentrations of different treated seeds were measured with GC-MS as described in Section 2.2.5. Results were presented as means ± SD from 2 replicates of 100 seeds each.
(A)
(B)
(D)
(C)
3
pH
10 3
pH
10
Mass
(kDa) 116 66.2 45.0 35.0 25.0 18.4 14.4 66.2 45.0 35.0 25.0 18.4 14.4 116Figure 13: Protein patterns of embryos of true seeds with different treatments.
The embryos of fresh (A, untreated), DI water treated (B), 5 mM citrate buffer (pH 2.0) treated (C), and 5 mM citrate buffer (pH 7.0) treated (D) true seeds were isolated for 2-D electrophoretic analysis. Labeled spots of the gels were those differentially expressed by at least ten-fold during the various treatments.
(A)
(B)
(D)
(C)
3
pH
10 3
pH
10
Mass
(kDa) 66.2 45.0 35.0 25.0 18.4 14.4 116 66.2 45.0 35.0 25.0 18.4 14.4 116Figure 14: Protein patterns of testae of true seeds with different treatments.
The testae of fresh (A, untreated), DI water treated (B), 5 mM citrate buffer (pH 2.0) treated (C), and 5 mM citrate buffer (pH 7.0) treated (D) true seeds were isolated for 2-D electrophoretic analysis. Labeled spots of the gels were those differentially expressed by at least ten-fold during the various treatments.
41
References
1. Baskin, C. C., Baskin, J. M. (1998) Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. San Diego, CA, USA: Academic Press.
2. Bewley, J. D. (1997) Seed germination and dormancy. The Plant Cell, 9, 1055-1066.
3. Cánovas, F. M., Dumas-Gaudot, E., Recorbet, G., Jorrin, J., Mock, H. P., Rossignol, M. (2004) Plant proteome analysis. Proteomics, 4, 285-298.
4. Chen, S. Y., Chien, C. T., Chung, J. D., Yang, Y. S., Kuo, S. R. (2007) Dormancy-break and germination in seeds of Prunus campanulata (Rosaceae): role of covering layers and changes in concentration of abscisic acid and gibberellins. Seed Science Research, 17, 21-32.
5. Chien, C.T., Chen, S.Y. and Yang, J.C. (2002) Effect of stratification and drying on the germination and storage of Prunus campanulata seeds. Taiwan Journal of Forest Science, 17, 413-420.
6. Chien, C. T., Lin. S.H., Hsu, P. J. (1996) The strategies enhancing germination of Prunus campanulata seeds. Taiwan Forest, 22, 10-11.
7. Chiu, C. W. (2006) The dramatic changes of sulfatase activities in Prunus campanulata Maxim. seeds from dormancy to germination. Thesis.
8. Cohn, M. A., Chiles, L. A., Hughes, J. A., Boullion, K. J. (1987) Seeddormancy in red rice. VI. Monocarboxylic acids: a new class of pH-dependent germination stimulants. Plant Physiology, 84, 716-719.
9. Eckerson, S. (1913) A physiological and chemical study of afterripening. Botanical Gazette, 55, 286-299.
10. Finkelstein, R., Reeves, W., Ariizumi, T., Steber, C. (2008) Molecular aspects of seed dormancy. Annual Review of Plant Biology, 59, 387-415.
42
Acidification during Dormancy-Breaking and Subsequent Germination. Plant Physiology, 100, 1196-1202.
12. Garcia-Mas, J., Messeguer, R., Arus, P., Puigdomenech, P. (1995) Molecular characterization of cDNAs corresponding to genes expressed during almond (Prunus amygdalus Batsch) seed development. Plant Molecular Biology, 27, 205-210.
13. Gendraud, M., Lafleuriel, J. (1983) Caracteristiques de l'absorptiondu saccharose et du tetraphenylphosphonium par lesparenchymes de tubercules de Topinambour, dormants et nondormants, cultives in vitro. Physiology of Vegetable, 21, 1125-1133.
14. Geneve, R. L. (2003) Impact of temperature on seed dormancy. Horticultural Science, 38, 336-341.
15. Gianinetti, A., and Vernieri, P. (2007) On the role of abscisic acid in seed dormancy of red rice. Journal of Experimental Botany, 58, 3449-3462.
16. Giersbach, J. and Crocker, W. (1932) Germination and storage of wild plum seeds. Contributions from the Boyce Thompson Institute, 4, 39-51.
17. Hiroyoshi, O. (1993) Editorial committee of the flora of Taiwan. Rosaceae, 3, 69-157.
18. International Seed Testing Association (ISTA) (1999) International rules for seed testing. Seed Science and Technology, 27, Supplement.
19. Jones, H. A. (1920) Physiological study of maple seeds. Botanical Gazette, 69, 127-152.
20. Kaiser, W. N. and Hartung, W. (1981) Uptake and release of abscisic acid by isolated photoautotrophic mesophyll cells, depending on pH gradients. Plant Physiology, 68, 202-206.
43
Growth Regulation, 24, 319-344.
22. Koornneef, M., Bentsink, L., Hilhorst, H. (2002) Seed dormancy and germination. Current Opinion in Plant Biology, 5, 33-36.
23. Lee, C. S., Chien, C. T., Lin, C. H., Chiu, Y. Y., Yang, Y. S. (2006) Protein changes between dormant and dormancy-broken seeds of Prunus campanulata Maxim. Proteomics, 6, 4147-4154.
24. Mehanna, H. T., Martin, G. C., Nishijima, C. (1985) Effects of temperature, chemical treatments and endogenous hormone content on peach seed germination and subsequent seedling growth. Scientia Horticulturae, 27, 63-73.
25. Nakayama, M., Koshioka, M., Matsui, H., Ohara, H., Mander, L. N., Leitch, S. K., Twitchin, B., Kraft-Klaunzer, P., Pharis, R. P., Yokota, T. (2001) Endogenous gibberellins in immature seeds of Prunus persica L.: identification of GA118, GA119, GA120, GA121, GA122 and GA126. Phytochemistry, 57, 749-758.
26. Nicolas, C., Nicolas, G., Rodrıguez, D. (1996) Antagonistic effects of abscisic acid and gibberellic acid on the breaking of dormancy of Fagus sylvatica seeds. Physiologia Plantarum, 96, 244-250.
27. Pack, D. A. (1921) After-ripening and germination of Juniperus seeds. Botanical Gazette, 71, 32-60.
28. Rose, R. C. (1919) After-ripening and germination of seeds of Tilia, Sambucus, and Rubus. Botanical Gazette, 67, 281-308.
29. Schopmeyer, C.S. (1974) Seeds of woody plants in the United States.US Department of Agriculture Forest Service. Agriculture Handbook No. 450. Washington, D. C.
30. Shevchenko, A., Wilm, M., Vorm, O., Mann, M. (1996) Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Analytical Chemistry, 68, 850-858.
44
31. Stephen, J. R., Dent, K. C., Finch-Savage, W. E. (2003) Molecular responses of Prunus avium (wild cherry) embryonic axes to temperatures affecting dormancy.
New Phytologist, 161, 401-413.
32. Tukey, H. B. and Barrett, M. S. (1936) Approximate germination test for non-after-ripened peach seed. Plant Physiology, 11, 629-633.
33. Velasco, R., Zharkikh, A., Troggio, M., Cartwright, D. A., Cestaro, A., Pruss, D., Pindo, M., Fitzgerald, L. M., Vezzulli, S., Reid, J., Malacarne, G., Iliev, D., Coppola, G., Wardell, B., Micheletti, D., Macalma, T., Facci, M., Mitchell, J. T., Perazzolli, M., Eldredge, G., Gatto, P., Oyzerski, R., Moretto, M., Gutin, N., Stefanini, M., Chen, Y., Segala, C., Davenport, C., Dematte, L., Mraz, A., Battilana, J., Stormo, K., Costa, F., Tao, Q., Si-Ammour, A., Harkins, T., Lackey, A., Perbost, C., Taillon, B., Stella, A., Solovyev, V., Fawcett, J. A., Sterck, L., Vandepoele, K., Grando, S. M., Toppo, S., Moser, C., Lanchbury, J., Bogden, R., Skolnick, M., Sgaramella, V., Bhatnagar, S. K., Fontana, P., Gutin, A., Van de Peer, Y., Salamini, F., Viola, R. (2007) A high quality draft consensus sequence of the genome of a heterozygous grapevine variety. PLoS ONE, 2, E1326.
34. Wang, M., Heimovaara-Dijkstra, S., Van Duijn, B. (1995) Modulation of germination of embryos isolated from dormant and nondormant barley grains by manipulation of endogenous abscisic acid. Planta, 195, 586-592.
35. Yan, J. X., Wait, R., Berkelman, T., Harry, R. A., Westbrook, J. A., Wheeler, C. H., H Dunn, M. J. (2000) A modified silver staining protocol for visualization of
proteins compatible with matrix-assisted laser desorption/ionization and electrospray ionization-mass spectrometry. Electrophoresis, 21, 3666-3672.
36. Zheng, L. and Poulton, J. E. (1995) Temporal and spatial expression of amygdalin hydrolase and (R)-(+)-mandelonitrile lyase in black cherry seeds. Plant
45
Physiology, 109, 31-39.
37. Zhou, J., Hartmann, S., Shepherd, B. K., Poulton, J. E. (2002) Investigation of the microheterogeneity and aglycone specificity-conferring residues of black cherry prunasin hydrolases. Plant Physiology, 129, 1252-1264.
38. Zhu, H., Bilgin, M., Snyder, M., (2003) Proteomics. Annual Review of Biochemistry, 72, 783-812.
39. Zielinski, Q. B. (1958) Some factors affecting seed germination in sweet cherries. Proceedings of the American Society for Horticulture Science, 72, 123-128.
46
Appendix
Equipment Company U-3300 UV/VIS Spectrophotometer Hitachi
High-speed centrifuge (himac CR 22G) Hitachi
550 Sonic Dismenbrator Fisher Scientific
pH meter Fisher Scientific
Ettan IPGphor IEF system GE Healthcare
Hoefer SE600 Ruby GE Healthcare
ImageScanner GE Healthcare
Orbital shaker DEHG YNG Instruments Co., LTD
PRO 250 homogenizer PRO Scientific
47
Autobiography
2006-2008: Institute of Biochemical Engineering, National Chiao Tung University, Hsinchu, Taiwan, ROC.
2000-2004: Department of Biological Resources, National Chiayi University, Chiayi, Taiwan, ROC.
1997-2000: National Taichung Wen-Hua Senior High School, Taichung, Taiwan, ROC.