行政院國家科學委員會專題研究計畫 成果報告
以具基因特異性之蛋白體學方法研究訊息傳導
計畫類別: 個別型計畫 計畫編號: NSC92-3112-B-002-003- 執行期間: 92 年 05 月 01 日至 93 年 04 月 30 日 執行單位: 國立臺灣大學醫學院臨床醫學研究所 計畫主持人: 周祖述 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 93 年 9 月 4 日
TABLE OF CONTENTS
Progress Report... 1
1. Response to previous reviewers’ critiques... 1
2. Specific
Aims ... 2
3. Progress
Summary... 3
4. Projected Timeline & Brief Summary of Plans for Next Year... 19
5. Personnel ... 20
6. Publications and/or Patents ... 21
6a. Publications ... 21
6b. Patents ...錯誤! 尚未定義書籤。
1
Progress Report
1. Response to previous reviewers’ critiques
Please describe the previous reviewers’ critiques and how based on the critiques
,
you
made modifications to specific aims, experimental design, or resource allocation etc.
1) In aim1, Dr. Jou propose to use the dual regulatory expressing system (Ecdysone and tetracycline inducible system) …….using constitutively active Rac1 and dominant negative IkB as a study pair. This aim is well thought through, and several alternative approaches were described. One minor comment is that the limitation of 2-D gel is not discussed, and the new ICAT and /or 2-D LC system is not mentioned as alternative approaches.
We have set up a collaboration with Dr. Chung-Lin Liao in Academia Sinica using ICAT as an alternative approach to 2DGE for identifying candidates in relaying the signaling from Rac1 to NFkB (please see the following section).
2) In aim 2, Dr. Jou proposed to construct stable cell lines,……… However, it is hard to know whether these candidates are biologically relevant. This is an intrinsic problem for all proteomic approaches.
With those candidate genes identified by microarray assay, we would then use quantitative RT-PCR and Western blotting analysis to confirm the roles they play in Rho family proteins signaling.
3) and 4) In aim 3, Dr. Jou proposed to perform proteomic approaches to discover target molecules linking RhoA family small GTPases after differentially expressing RhoA/Rac1/Cdc42 mutant genes…... With very good controls and carefully designed experiments, he should be able to produce very interesting results. In aim 4, Dr. Jou proposed to apply these systems to study medically important issues. ……..
2
2. Specific Aims
Our primary goal, which has not been modified so far, is to generate double inducible gene expression system in a simplistic and efficient way, and apply this system for dissecting signalling transduction pathways involving Ras related small GTPases using experimental approaches, such as DNA microarray and proteomic studies, based upon comparison of global differences in gene expression profiles.
Specific Aim I. Using the dual regulatory expressing system (Ecdysone and tetracycline inducible
system) already set up in HEK293 cell lines to demonstrate the feasibility of conjugating this double regulated expression system with proteomic studies for dissecting complex signalling transduction pathways; using constitutively active Rac1 and dominant negative IkB as a study pair.
We would like to catagorize the downstream signaling molecules of Rac1, according to the global gene expression patterns, into three groups:
Group X- molecules downstream of Rac1and upstream of NFkB Group Y- molecules downstream of NFkB and further downstream of Rac1
Group Z- molecules downstream of Rac1and unrelated to NFkB signaling pathway
Specific Aim II. Construct stable cell lines, using inducible expression systems set up in MDCK
cells, expressing candidate genes identified during the experimental approaches aimed at specific aim I to confirm the finding and also explore the relevant biological meaning
Specific Aim III. Performing proteomic approaches to discover target molecules linking RhoA
family small GTPases after differentially expressing RhoA/Rac1/Cdc42 mutant genes in either dual regulatory HEK293 or MDCK cells.
Specific Aim IV. Apply the achievement that would be made to characterize genes involved in
hepatocellular oncogensis after successfully carrying out the researches mentioned for specific aim I to III.
Specific Aim IV(A). Construct dual regulatory cell lines in differentiated rat liver cell line,
WB, and transformed hepatic cell line, such as HepG2, Hep3B, and Huh7.
Specific Aim IV(B). Generate stable cell lines expressing viral genes or other
oncogenesis-related candidate genes in cell line set up under Specific Aim IV (A), and then apply comparative proteomic studies to dissect the signalling pathways leading to hepato-oncogenesis .
3
3. Progress Summary
To pursue
specific aim 1
of our proposal, we had used the following three different approaches in the past 8 months:(a) Two-dimensional gel electrophoresis (2-DGE) analysis- both (1a) conventional and
(1b) DIGE (Difference Gel Electrophoresis), which is using the commercially available CyDye system (Amersham/Pharmacia) to differentially label different protein samples, to resolve our samples in gel.
(b) Isotope-Coded Affinity Tag (ICAT) technique- a collaboration between Dr. Chung-Lin Liao's group in the Institute of Chemistry in the Academia Sinica.
(c) DNA Microarray analysis
Before we present our data in the later section, we would like to first define our experimental settings and use E1~E4 as the abbreviations to facilitate communication.
The lysates or mRNAs from HEK293 cells differentially expressing either constitutive Rac1 (Rac1V12) or dominant negative IkB(IkB-DN) mutant were collected in the following settings:
Experimental condition 1 (E1)- neither Rac1V12 nor IkB-DN was expressed (Basal condition) Experimental condition 2 (E2)- Only Rac1V12 was induced (all group X, Y, and Z protein
expressions should be elicited)
Experimental condition 4 (E4)- Both Rac1V12 and IkB-DN were expressed (group Y proteins
should return to near basal condition, group Z should not be affected, while group X proteins might be further changed)
Experimental condition (E3) was not performed, because it was not related to our goal, in which only IkB-DN was expressed, and therefore group Y proteins might be down-regulated.
Two-dimensional gel electrophoresis (2-DGE) analysis
While we were using the conventional 2DGE to resolve the protein lysates from experimental condition 1, 2, and 4 (E1, E2, and E4), we met some problems at first including inadequate sonication of the cell pellets or inadequate use of protease inhibitors, just to name a few of them. We eventually solved out what would be the most optimal condition to get the proteins well separated on 2D gel and stained by Sybro Ruby staining for protein quantification (Fig. 1).
4
In the meantime, we also tried the DIGE system advocated by Amersham/Pharmacia. Although we could see many differentially labeled spots on the gels, but many are just artifacts caused by so-called reflection errors (the single arrows in Fig. 2A, 2B, and 2C), which are characterized by the double-vision like, vertically overlapped images. We noticed they were happening no matter which combination of protein samples were subjected for the DIGE analysis. There were also some other types of differentially labeled spots visible on the gel, which were well separated and horizontally arranged (the double arrows in Fig. 2A, 2B, and 2C). Initially, we thought these might be the ones which we sought to pursue. However, we found they were still happening without any correlation to the ways how protein samples were combined; i.e., no matter whether Rac1V12 or dominant negative IkB was induced or not, they were there. Furthermore, we also found if we expressed different pair of transgenes, such as Rac1V12 and RhoAV14, they were still there with the same kind of labeling pattern (data not shown). We were puzzled by this finding and decided to isolate the protein spots for mass spectrometrical analysis to make sure the identities of those mysterious proteins.
Due to the intrinsic problems associated with DIGE, we decided to take conventional approach for 2DGE despite it is tedious and challenging to get three gels (E1, E2, and E4 on three separate gels) run at the same condition. Nevertheless, we did catagorize the differential expression pattern under these three different conditions (E1, E2, and E4) into group X, Y, and Z patterns. The examples are given in Fig. 3.
We have so far identified 7, 7, and 5 spots with group X, Y, and Z expression patterns, respectively, and would be analyzed by MS them when we get enough proteins.
Isotope-Coded Affinity Tag (ICAT) technique
Collaborating with Dr. Chung-Lin Liao in the Institute of Chemistry, Academia Sinica (Genomics/Proteomics Center), we also had our protein samples undergo ICAT approach. Dr. Liao developed a new ICAT reagent, which labels the NH2- group, instead of SH-group, of proteins. This results in higher chance of labeling in proteins containing few cysteines. After trypsinization of the protein samples prepared in E1, E2, and E4 conditions, the peptides were differentially labeled with hydrogen (D0, light) or deuterium (D4, heavy) containing reagents, and undergoing multiple dimensional liquid chromatography (MDLC) and MS analysis. Initial protein identification revealed about one thousand proteins in the samples, and further manual MS spectrum inspection, which took totally three weeks' effort because there was no commercially available software could reliably substitute this part of work, confirmed about 200 peptide pairs between E1/E2 and E2/E4 experimental conditions. A selective list of proteins and their quantification ratios in E2/E1 and E4/E2 conditions are shown in Table 1 and Table 2.
5
So far, we haven't identified a protein significantly increasing in E2 vs. E1 conditions, and also in E4 vs. E2 conditions. Our hypothesis for this phenomenon would be discussed later after we presented the data from our microarray analysis.
DNA Microarray analysis
We initially performed microarray assays at our own lab using the protocol and DNA arrays from the Third Core Laboratory in National Taiwan University Hospital, which is also sponsored by the
National Research Program in Genomic Medicine. However, we got inconsistent data which
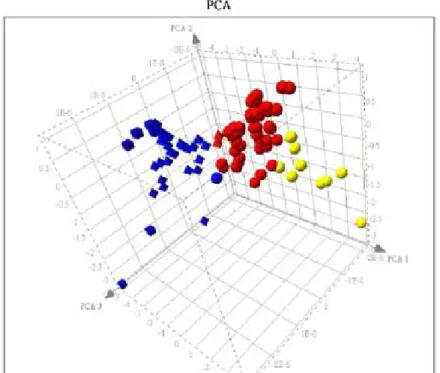
were difficult to analyze. Later, we shifted to glass type DNA array using fluorescent probe instead of alkaline phosphatase based colorimetric probe. We think we have finally got much more reliable data (Table 3A-C) because we could identify Rac1, the transgene regulated by the tetracycline inducible system, gene up-regulated in experimental condition E2 vs. E1, and the Rac1 mRNA level is the same between experimental condition E2 and E4 (group Z expression pattern in Table 3C). Furthermore, after we used several algorithms to analyze the expression pattern we got, we could get pretty nice clustering of the gene expression patterns (Fig. 4A and 4B).
There are many interesting genes and expression patterns from the microarray assay. Taking the genes classified as group X expression pattern for examples, ARRH (RhoH) is an atypical Rho family GTPase, which doesn't have endogenous GTPase activity and is demonstrated to have a dominant negative effect of the endogenous RhoA protein. It has been previously hypothesized that Rac1 has an antagoisitic effect on RhoA, but the detailed mechanism has been elusive. If RhoH is activated by Rac1, which might be the reason why Rac1 activation might downregulate RhoA activity.
Another gene belonging to group X expression pattern, FNBP1 (formin binding protein 1), has been demonstrated to be a Cdc42 interacting partner and a GEF for Cdc42. This protein may act as a link between cdc42 signaling and regulation of the actin cytoskeleton. If FNBP1 is transcriptionally regulated by Rac1, this could provide a mechanistic basis for the interaction between Rac1 and Cdc42 on actin cytoskeleton.
6
Mismatch between the data from microarray and ICAT assays, and the possible solutions to reconcile these differences in the future
Although we are satisfied with the result coming out of microarray assay, the protein expression patterns between the result from microarray do not match with each other. Our explanation is that we overload the MS capacity with a protein sample of very high complexity. The ICAT reagent we used is different from the conventional one in that the former doesn't contain biotin modification and we therefore do not enrich the ICAT labeled tryptic peptides with streptavidin beads. Basically, all the proteins, after trypsin digestion, are entering the MDLC-MS analysis, and it is very likely that the MS spectra of low abundance proteins are masked by those of high abundance proteins. If the expression products of those genes identified by microarray are low in abundance, it is very likely they could be missed from our current ICAT assay.
To solve this problem, we are planning to fractionate our protein samples first into membranous, microsomal, cytosolic, nuclear, and cytoskeletal compartments, followed by reverse phase LC separation of the protein samples of each sub-fractionation before going to the ICAT procedure. Preliminary data from Dr. Liao's group show this might be helpful in decreasing the complexity of proteins going into the ICAT analysis.
In the meantime, we are going to isolate the protein complex by immunoprecipitating either the myc-epitope tagged Rac1V12 or the inhibitor of nuclear factor kappa B (IkB) under experimental conditions differentially expressing constitutively active Rac1V12 or dominant negative IkB. We hypothesize the associated signaling molecules to either Rac1V12 IkB would be different when the signaling pathways are manipulated by Rac1 and IkB mutant proteins. This might sound a desperate approach, but could be easily tested.
Specific Aim II. Construct stable cell lines, using inducible expression systems set up in MDCK
cells, expressing candidate genes identified during the experimental approaches aimed at specific aim I to confirm the finding and also explore the relevant biological meaning
This part of the project awaits the identities of those candidate genes being disclosed after experiments related to Specific Aim I would be completed.
7
Specific Aim III. Performing proteomic approaches to discover target molecules linking RhoA
family small GTPases after differentially expressing RhoA/Rac1/Cdc42 mutant genes in either dual regulatory HEK293 or MDCK cells.
To search for possible connections between different members of Rho family GTPases, we have focused in the past one year on the connections between Cdc42 and Rac1. We had then demonstrated a signaling cascade ignaling cascade Cdc42 → Rac1 → PI3K in modulating
detachment induced apoptosis (anoikis). This part of work has been written up as a submitted manuscript, and now the manuscript has been revised and re-evaluated by Experimental Cell
Research (please refer to the manuscript attached in appendix 2).
Specific Aim IV(A). Construct dual regulatory cell lines in differentiated rat liver cell line, WB,
and transformed hepatic cell line, such as HepG2, Hep3B, and Huh7.
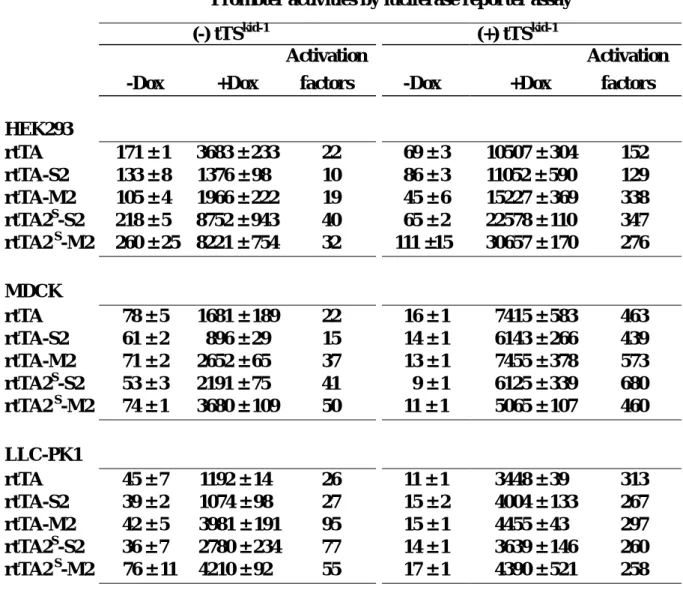
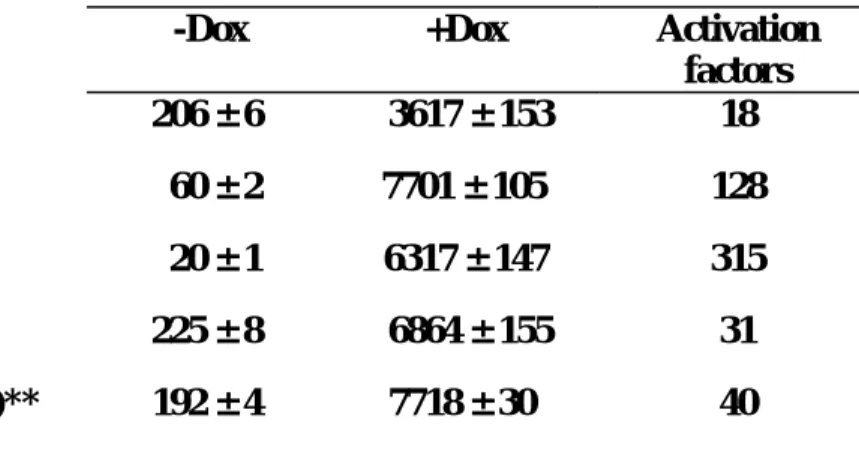
We have so far generated Tetracycline inducible expression system in three of the above three cell lines, using the strategy we have recently published (Am J Physiol Cell Physiol 2003; 285:C711-719). Taking the Huh7 cell line as example, after three rounds of positive FACS based selection and one round of negative selection, we have enriched inducible clones with an inducibility of about 100 folds.
While we were generating the dual regulatory expression system, we accidentally found doxycycline, previously thought to be an inert chemical compound, might differentially regulate the expression level of reverse type tetracycline regulated tranactivator (rtTA). We studied the mechanistic basis of this phenomenon and wrote up a manuscript which had been submitted to
Journal of Gene Medicine (please appendix 3).
For HepG2 and Hep3B, after similar approaches, we have inducible cell populations with inducibility of about 60 and 80 folds.
As for the differentiated WB cell line, we would conduct similar selection scheme very soon. Once we finish establishing tetraycline controlled expression system in all these four cell lines, we would set up ecdysone inducible expression system in them to complete the making of dual regulatory expression systems in four cell lines.
8
Table 1
Proteins identified to be up-regulated in E2 vs E1 condition by ICAT analysisMascot No.
Accession No. Protein Name E2/E1
23 gi|4885375 H1 histone family, member 2; histone H1d [Homo sapiens] 1.541
373 gi|27498927 similar to CG5882-PA [Homo sapiens] 1.474
422 gi|4506583 replication protein A1, 70kDa; replication protein A1 (70kD) [Homo sapiens]
1.429
396 gi|37538778 similar to dJ753D5.2 (novel protein similar to RPS17 (40S ribosomal protein S17)) [Homo sapiens]
1.426
186 gi|444021 sub2.3 [Homo sapiens] 1.409
947 gi|6424942 ALG-2 interacting protein 1 [Homo sapiens] 1.397
506 gi|243123 cytochrome c oxidase subunit VIIa='liver-type' isoform {EC 1.9.3.1} [human, skeletal muscle, Peptid
1.394
360 gi|4506681 ribosomal protein S11; 40S ribosomal protein S11 [Homo sapiens] 1.383
559 gi|9022435 ribosomal protein S3; RPS3 [Homo sapiens] 1.378
233 gi|6841566 HSPC173 [Homo sapiens] 1.370
448 gi|4757900 calreticulin precursor; Sicca syndrome antigen A (autoantigen Ro; calreticulin); autoantigen Ro [Ho
1.370
474 gi|5032087 splicing factor 3a, subunit 1, 120kDa; pre-mRNA splicing factor SF3a (120 kDa subunit) [Homo sapien
1.336
471 gi|36142 ribosomal protein homologous to yeast S24 [Homo sapiens] 1.335
230 gi|478813 nonhistone chromosomal protein HMG-1 - human 1.318
183 gi|14250148 Ribosomal protein L3 [Homo sapiens] 1.313
8 gi|1136741 KIAA0002 [Homo sapiens] 1.298
623 gi|5901922 CDC37 homolog [Homo sapiens] 1.293
122 gi|2136253 TCP1 ring complex protein TRiC5 - human 1.281
194 gi|1203969 filamin [Homo sapiens] 1.278
46 gi|4507677 tumor rejection antigen (gp96) 1; Tumor rejection antigen-1 (gp96); glucose regulated protein, 94 k
1.276
28 gi|28614 aldolase A [Homo sapiens] 1.273
295 gi|178663 medium tumor antigen-associated 61-kD protein 1.272
84 gi|7433799 fatty-acid synthase (EC 2.3.1.85) (version 2) - human 1.267
75 gi|115206 C-1-tetrahydrofolate synthase, cytoplasmic (C1-THF synthase) [Includes: Methylenetetrahydrofolate d
1.262
192 gi|423123 tpr protein - human 1.250
141 gi|553640 ribosomal protein S13 [Homo sapiens] 1.249
539 gi|20987810 MRPS27 protein [Homo sapiens] 1.214
18 gi|5174735 tubulin, beta, 2 [Homo sapiens] 1.213
147 gi|5174447 guanine nucleotide binding protein (G protein), beta polypeptide 2-like 1; protein homologous to ch
9
Table 2
Proteins identified to be up-regulated in E4 vs E2 condition by ICAT analysisMascot No.
Accession No. Protein Name 4H/2L
ratio 102 gi|37542903 similar to 60 kDa heat shock protein, mitochondrial precursor (Hsp60)
(60 kDa chaperonin) (CPN60) (
8.29
350 gi|37538336 KIAA1856 protein [Homo sapiens] 2.99
1090 gi|14591909 ribosomal protein L5; 60S ribosomal protein L5 [Homo sapiens] 1.33 59 gi|4758756 nucleosome assembly protein 1-like 1; HSP22-like protein interacting
protein; NAP-1 related protein
1.31 469 gi|2135244 chromosome segregation protein smc1 [similarity] - human 1.28
62 gi|6005854 repressor of estrogen receptor activity; B-cell associated protein [Homo sapiens]
1.22 71 gi|7433799 fatty-acid synthase (EC 2.3.1.85) (version 2) - human 1.21 589 gi|4884564 vitamin D3 receptor interacting protein [Homo sapiens] 1.21
41 gi|180555 creatine kinase-B 1.19
93 gi|105294 alternative splicing factor ASF-2 - human 1.18
142 gi|20521660 KIAA0788 protein [Homo sapiens] 1.14
186 gi|438069 thiol-specific antioxidant protein [Homo sapiens] 1.14 177 gi|2559008 chaperonin containing t-complex polypeptide 1, delta subunit;
CCT-delta [Homo sapiens]
1.10 79 gi|4503297 DEAH (Asp-Glu-Ala-His) box polypeptide 9 isoform 1; DEAD/H
(Asp-Glu-Ala-Asp/His) box polypeptide 9
1.08 90 gi|4506607 ribosomal protein L18; 60S ribosomal protein L18 [Homo sapiens] 1.07 513 gi|8923579 hypothetical protein FLJ20625 [Homo sapiens] 1.07
805 gi|4759196 symplekin [Homo sapiens] 1.06
841 gi|25136577 ELYS transcription factor-like protein TMBS62 [Homo sapiens] 1.04 96 gi|4506671 ribosomal protein P2; 60S acidic ribosomal protein P2; acidic
ribosomal phosphoprotein P2 [Homo sap
1.04 249 gi|4506691 ribosomal protein S16; 40S ribosomal protein S16 [Homo sapiens] 1.02 74 gi|9802306 DNA-binding protein TAXREB107 [Homo sapiens] 1.00
170 gi|5031635 cofilin 1 (non-muscle) [Homo sapiens] 1.00
49 gi|337424 poly(ADP-ribose) synthetase 0.99
8 gi|7106439 tubulin, beta 5 [Mus musculus] 0.99
254 gi|135538 T-complex protein 1, alpha subunit (TCP-1-alpha) (CCT-alpha) 0.99
313 gi|136066 TRIOSEPHOSPHATE ISOMERASE (TIM) 0.99
220 gi|4506209 proteasome 26S ATPase subunit 2; proteasome 26S subunit, ATPase, 2; mammalian suppressor of sgv-1 o
0.98 305 gi|1431788 Chain A, Cyclophilin A Complexed With Cyclosporin A (Nmr, 22
Structures)
0.98 456 gi|36142 ribosomal protein homologous to yeast S24 [Homo sapiens] 0.98 283 gi|18848326 Similar to GDP dissociation inhibitor 2 [Homo sapiens] 0.97
10
Table 3A. Genes displaying group X expression pattern (expression ratio under E2/E1 experimental conditions is more than 2, while expression ratio under E4/E2 experimental conditions is more than 1.45) by DNA microarray assay
CLID NAME E2-E1 E4-E2
IMAGE:704274 transcriptional regulating protein 132 20.549 2.540
IMAGE:1306637 tumor protein D52 2.382 2.227
IMAGE:1285720 formin binding protein 1 2.133 1.925
IMAGE:454475 phosphorylase kinase, alpha 2 (liver) 2.035 1.557 IMAGE:302591 ras homolog gene family, member H 8.846 1.491 IMAGE:1307809 Similar to interferon-gamma receptor alpha chain 2.208 1.467
Table 3B. Genes displaying group Y expression pattern (expression ratio under E2/E1 experimental conditions is more than 2, while expression ratio under E4/E2 experimental conditions is less than 0.5) by DNA microarray assay
CLID NAME E2-E1 E4-E2
IMAGE:809901 collagen, type XV, alpha 1 19.213 0.032
IMAGE:789369 inhibitor of DNA binding 4, dominant negative helix-loop-helix protein
8.462 0.083 IMAGE:196992 aldo-keto reductase family 1, member C1 8.369 0.077 IMAGE:1309018 uncharacterized hypothalamus protein HCDASE 7.863 0.062 IMAGE:430038 FYN binding protein (FYB-120/130) 6.797 0.141 IMAGE:669310 mitogen-activated protein kinase-activated protein
kinase 5
6.440 0.259
IMAGE:712341 ribonuclease 6 precursor 5.996 0.128
IMAGE:323181 fibroblast activation protein, alpha 5.856 0.241
IMAGE:21655 5'-nucleotidase, ecto (CD73) 5.680 0.181
IMAGE:223176 MAX dimerization protein 1 5.389 0.206
IMAGE:2477598 secretory leukocyte protease inhibitor 5.216 0.499 IMAGE:1271368 SH3-domain binding protein 1 4.803 0.368
IMAGE:784928 sushi-repeat protein 4.721 0.372
IMAGE:461727 phenylalanine hydroxylase 4.624 0.151
IMAGE:1302646 chromosome 13 open reading frame 9 4.579 0.363 IMAGE:814251 signaling lymphocytic activation molecule family
member 1
4.569 0.460 IMAGE:753346 aminolevulinate, delta-, synthase 2 4.341 0.477 IMAGE:1240974 Rho GDP dissociation inhibitor (GDI) beta 4.284 0.360
IMAGE:2448778 cathepsin F 4.067 0.265
IMAGE:2508044 haptoglobin 4.059 0.241
IMAGE:1287536 zinc finger protein, subfamily 1A, 1 (Ikaros) 4.006 0.250 IMAGE:1470151 GRB2-associated binding protein 2 3.926 0.168
11
Table 3B. Continued
CLID NAME E2-E1 E4-E2
IMAGE:74537 alpha-fetoprotein 3.880 0.208
IMAGE:204541 asialoglycoprotein receptor 1 3.853 0.266 IMAGE:685022 diacylglycerol kinase, epsilon 64kDa 3.829 0.395
IMAGE:712278 c-fos 3.769 0.245
IMAGE:428083 basic transcription factor 2 3.630 0.368 IMAGE:487418 filamin A, alpha (actin binding protein 280) 3.620 0.274 IMAGE:2028959 dopa decarboxylase (aromatic L-amino acid decarboxylase) 3.615 0.371 IMAGE:1335809 hypoxia-inducible factor 1, alpha subunit 3.470 0.394 IMAGE:243174 serine (or cysteine) proteinase inhibitor, clade F (alpha-2
antiplasmin member 2
3.425 0.392
IMAGE:2353365 deoxyribonuclease I-like 1 3.371 0.290
IMAGE:704815 Inhibitor of DNA binding 2 3.274 0.239
IMAGE:70692 serine (or cysteine) proteinase inhibitor, clade B (ovalbumin)
3.182 0.247 IMAGE:284022 Rho guanine nucleotide exchange factor (GEF) 10 3.141 0.423 IMAGE:565319 mal, T-cell differentiation protein 2 3.076 0.301
IMAGE:324492 matrix metalloproteinase 3 3.072 0.433
IMAGE:612576 3-oxoacid CoA transferase 3.055 0.402
IMAGE:2449395 aldo-keto reductase family 1, member C2 3.044 0.310 IMAGE:825645 activation-induced cytidine deaminase 3.013 0.392
IMAGE:1534853 centaurin, beta 1 3.008 0.394
IMAGE:325145 pentaxin-related gene, rapidly induced by IL-1 beta 2.932 0.456 IMAGE:2449786 tumor necrosis factor receptor superfamily, member 18 2.882 0.393
IMAGE:50604 **androgen-induced 1 2.834 0.425
IMAGE:1241180 zinc finger protein, subfamily 1A, 1 (Ikaros) 2.670 0.299
IMAGE:1334310 CGI-109 protein 2.661 0.332
IMAGE:50503 integrin, beta 2 2.650 0.232
IMAGE:840844 heat shock 70kDa protein 5 2.623 0.264 IMAGE:1656636 UDP glycosyltransferase 2 family, polypeptide B4 2.583 0.312
IMAGE:704084 centaurin, delta 1 2.567 0.341
IMAGE:487429 collagen, type VI, alpha 1 2.491 0.271
IMAGE:212188 apolipoprotein H (beta-2-glycoprotein I) 2.485 0.283
IMAGE:1306275 mutL homolog 3 (E. coli) 2.224 0.328
IMAGE:2298080 kynureninase (L-kynurenine hydrolase) 2.213 0.345 IMAGE:1681489 serine (or cysteine) proteinase inhibitor, clade E (nexin,
plasminogen activator inhibitor type 1), member 1
2.210 0.326
IMAGE:31093 cadherin 13, H-cadherin (heart) 2.155 0.205 IMAGE:1241157 tripartite motif-containing 22 2.151 0.338
IMAGE:668182 zinc finger protein 193 2.120 0.326
IMAGE:768443 microsomal glutathione S-transferase 1 2.104 0.326 IMAGE:2458975 inhibin, beta A (activin A, activin AB alpha polypeptide) 2.089 0.227
12
Table 3C. Genes displaying group Z expression pattern (expression ratio under E2/E1 experimental conditions is more than 2, while expression ratio under E4/E2 experimental conditions is between 0.5 and 1.45) by DNA microarray assay
CLID NAME E2-E1 E4-E2
IMAGE:1369976 ecotropic viral integration site 2A 9.547 0.564
IMAGE:143523 collagen, type V, alpha 1 5.049 1.438
LCP:138 methionine-tRNA synthetase 4.800 1.217
IMAGE:2117981 47-kD autosomal chronic granulomatous disease protein 4.332 1.250 IMAGE:703774 **mitogen-activated protein kinase 8 interacting protein 3 4.332 0.699
IMAGE:1338277 RAC1 4.190 0.812
IMAGE:2491247 solute carrier family 21 (organic anion transporter), member 9
3.761 0.692 IMAGE:827132 ras-related C3 botulinum toxin substrate 2 3.489 1.426 IMAGE:781105 AHA1, activator of heat shock 90kDa protein ATPase
homolog 2 (yeast)
3.437 1.154
IMAGE:825590 xylosyltransferase 3.401 1.301
IMAGE:1307643 phosphatidylinositol transfer protein, membrane-associated 1
3.265 0.529
IMAGE:712279 kelch-like 6 (Drosophila 3.211 0.641
LCP:1 deoxynucleotidyltransferase, terminal 3.180 1.053 IMAGE:284263 myelin-associated oligodendrocyte basic protein 3.134 0.777
IMAGE:1019777 butyrylcholinesterase 3.130 0.851
IMAGE:704154 epidermodysplasia verruciformis 1 3.065 1.113
IMAGE:563574 FSHD region gene 1 2.986 0.527
IMAGE:701572 NADP-dependent retinol dehydrogenase/reductase 2.963 0.923 IMAGE:1184934 purinergic receptor P2Y, G-protein coupled, 8 2.914 0.723 IMAGE:683257 EST from selenoprotein P promoter region 2.866 0.954 IMAGE:358842 runt-related transcription factor 3 2.805 1.171 IMAGE:685210 T-cell activation GTPase activating protein 2.572 1.306
IMAGE:127636 **zuotin related factor 1 2.530 1.281
IMAGE:686274 peptidylprolyl isomerase (cyclophilin)-like 2 2.519 1.163
IMAGE:1543346 transketolase-like 1 2.474 1.221
CLID NAME E2-E1 E4-E2
IMAGE:701018 transcription factor Dp-2 (E2F dimerization partner 2) 2.302 1.340 IMAGE:1306711 ceroid-lipofuscinosis, neuronal 6, late infantile, variant 2.299 1.324 IMAGE:260200 myeloid cell nuclear differentiation antigen 2.201 1.401 IMAGE:80915 succinate dehydrogenase complex, subunit A 2.124 1.437 IMAGE:1286006 centrosomal kinesin-like protein 2.108 1.442 IMAGE:1670870 unc-93 homolog B1 (C. elegans) 2.062 1.429
13
Figure 1
14
Figure 2A
E1 lysate- neither Rac1V12 nor IkB-DN was expressed, labeled in red
15
Figure 2B
E2 lysate- Only Rac1V12 was induced, labeled in red
16
Figure 2C
E1 lysate- neither Rac1V12 nor IkB-DN was expressed, labeled in red
E4 lysate- Both Rac1V12 and IkB-DN were expressed, labeled in green
17
patterns; the left upper quadrant in each composite image shows the quantified intensities of the index spots as a histogram in the order of E1, E2, and E4. The gels were stained with Sybro Ruby stain, scanned by Typhoon 9200 (Amersham/Pharmacia) and quantified by using the PDQuest7.1.1 software (Bio-Rad).
18
Figure 4A
Hierarchical clustering result of group X (red), Y (green) , and Z (blue) genes.Figure 4B
Non-hierarchical clustering result of group X (yellow), Y (blue), and Z (red) genes.19
4. Projected Timeline & Brief Summary of Plans for Next Year
Specific Aim I. (From 2004-01-01 to 2004-4-30)
Using the combined approaches by conventional 2DGE, ICAT, and microarray assay to dissect the complex signalling transduction pathways linking constitutively active Rac1 and nuclear factor kappa B (NFkB), we would eventually track down the possible candidates.
As we found the complexity of our protein samples might be too high for the ICAT assay, we plain to sub-fractionate the protein samples according to their distribution in different cellular compartment, and the fractionated protein samples would be further separated by reverse phase liquid chromatography. We would use quantitative real-time RT-PCR and western blotting analysis to confirm the expression patterns of several candidate genes we identified by microarray, notably those involved in Rho family protein signalling, such as RhoH and FNBP1(formin binding protein 1). We also plan to summarize our microarray data in a small methodology paper.
Specific Aim II. (From 2004-5-01 to 2004-08-31)
Construct stable cell lines, using inducible expression systems set up in MDCK cells, expressing candidate genes identified during the experimental approaches aimed at specific aim I to confirm the finding and also explore the relevant biological meaning
Specific Aim III. (From 2004-01-01 to 2004-12-31)
Given the experience we obtained in the past 8 months or so, we think we could start using the combined proteomic and microarray approach to discover target molecules linking Rho family small GTPases after differentially expressing RhoA/Rac1/Cdc42 mutant genes in either dual regulatory HEK293 or MDCK cells. Since we have already written up a manuscript, which is under second revision after being sent to Experimental Cell Research, describing the signaling cascade Cdc42 → Rac1 → PI3K in modulating detachment-induced apoptosis (anoikis) (please
refer to the manuscript in appendix 2). We would propably start from the constitutively active Cdc42 and dominant negative Rac1 double mutant cells first.
Specific Aim IV. (From 2004-01-01 to 2004-06-30)
Specific Aim IV(A). Finish constructing dual regulatory cell lines in differentiated rat liver
20
5. Personnel
Summarize the
personnel involved in the project during the grant period
. List
the personnel in accordance to the following categories: (1) senior investigators,
including visitors; (2) postdoctoral fellows; (3) graduate students; (4) technicians or
research assistants. Specify for each individual the period of involvement and the
percentage commitment of effort.
Name In Chinese In English Position Title Education Degree % of personal effort on this project Job Description or Responsibilities
周祖述
鄭子琳
鄭心遠
蔡舒聿
林育誼
Tzuu-Shuh Jou Tzu-Ling Cheng Hsin-Yuan Cheng Su-Yu Tsai Yu-Yi Lin Assistant Professor (Principal Investigator) 1st year technician 1st year technician 1st year technician 3rd Year Resident in internal medicine MD;Ph.D Master degree Master degree Master degree M.D. 50% (with another NSC sponsored project at hand) 100% 100% 100% 50%Coordinating the works and planning the experiments associated with this project Writing proposals,
manuscripts, and rebuttals for the publication
DNA plasmid constrtuction, Cell Culturing, and
Making stable clones
. cell biology and biochemical assay; carrying the project related to the submitted paper No-1
cell biology and biochemical assay; carrying the project related to the submitted paper No-2
Protein extraction, Running 2D gel
FACS sorting
Microarray assay and data analysis
21
6. Publications and/or Patents
6a. Publications
List the title and complete references (author(s), journal or book, year, page number)
of all publications
directly resulting from studies supported by the project (i.e.,
with citation of this grant in the acknowledgement section)
. List the publications
for the project in accordance to the following categories: (1) manuscripts published
and accepted for publications; (2) manuscripts submitted; and (3) conference
proceedings. Provide one copy of each publication not previously reported to the
National Science Council in the Appendix.
1.
Lai JF, Juang SH, Hung YM, Cheng HY, Cheng TL, Mostov KE, Jou TS.
Related Articles, Links
An ecdysone and tetracycline dual regulatory expression system for studies on Rac1 small GTPase-mediated signaling.
Am J Physiol Cell Physiol. 2003 Sep;285(3):C711-9. Epub 2003 May 07.
2. Tzu-Ling Cheng, Marc Symons, Tzuu-Shuh Jou Regulation of Anoikis by Cdc42 and Rac1
(Submitted to Experimental Cell Research, revised and resubmitted)
3. Jen-Feng Lai, Hsin-Yuan Cheng, Tzu-Ling Cheng, Yu-Yu Lin, Li-Chieh Chen, Mau-Ting Lin, Tzuu-Shuh Jou
Doxycycline and Tetracycline Regulated Transcriptional Silencer Enhance the Expression Level and Transactivating Performance of rtTA
22
Appendix 1
Lai JF, Juang SH, Hung YM, Cheng HY, Cheng TL, Mostov KE, Jou TS. An ecdysone
and tetracycline dual regulatory expression system for studies on Rac1 small GTPase-mediated signaling. Am J Physiol Cell Physiol. 2003 Sep;285(3):C711-9. Epub 2003
May 07 (To save pages, please log onto
23
Appendix 2 (Submitted to Experimental Cell Research, revised and
resubmitted).
Regulation of Anoikis by Cdc42 and Rac1
Tzu-Ling Cheng, Marc Symons*, Tzuu-Shuh Jou§
Department of Internal Medicine, National Taiwan University Hospital and National Taiwan University College of Medicine
No. 7, Chung-Shan S. Road Taipei, 100 Taiwan
* Center for Oncology and Cell Biology, North Shore-LIJ Research Institute, 350 Community Drive, 11030, Manhasset, NY, USA
§To whom reprint should be addressed. Tel: 8862-23123456 ext.7258
Fax: 8862-23709820
E-mail: [email protected]
Manuscript Information: 41 pages of text, and 8 figures in the paper.
24
ABSTRACT
Ras family small GTPases play a critical role in malignant transformation, and Rho subfamily
members contribute significantly to this process. Anchorage-independent growth and the ability to
avoid detachment-induced apoptosis (anoikis) are hallmarks of transformed epithelial cells. In this
study, we have demonstrated that constitutive activation of Cdc42 inhibits anoikis in Madin Darby
canine kidney (MDCK) epithelial cells. We showed that activated Cdc42 could stimulates the
ERK, JNK and p38 MAPK pathways in suspension condition; however, inhibition of these
signalling does not affect Cdc42-stimulated cell survival. However, we demonstrated that
iInhibition of phosphatidylinositol 3-kinase (PI3K) pathway however abolishesd the protective
effect of Cdc42 on anoikis. Taking advantage of a double regulatory expression system, we also
showed that Cdc42-stimulated cell survival in suspension condition is, at least in part, mediated by
Rac1. The consequence of Rac1 activation initiates aWe also provide evidence for a positive
regulatory feedback loop between involving Rac1 and PI3K. In addition, we show that the
survival functions of both constitutively active Cdc42 and Rac1 GTPases are abrogated by
Latrunculin B, an actin filament-depolymerizing agent, implying an important role for the actin
cytoskeleton in mediating survival signaling activated by Cdc42 and Rac1. Together, our results
suggest indicate a role for Cdc42 in anchorage-independent survival of epithelial cells. We also
propose conclude that this survival function depends on a positive feedback loop involving Rac1
25
26
INTRODUCTION
The interaction between cells and the surrounding matrix is a major determinant of cellular
behavior, modulating gene expression, cell growth and differentiation, cell migration, and overall
tissue architecture [1]. Anchorage-dependent survival is also an important consequence of
cell-matrix interaction [2]. Epithelial cells, endothelial cells, and muscle cells undergo
programmed cell death when they are deprived of the contact with extracellular matrix [3].
Apoptosis induced by disruption of the interaction between epithelial cells and extracellular
matrix has been termed as "anoikis", which means homelessness in Greek [4]. Anoikis plays an
essential role in regulating tissue homeostasis in normal epithelial tissues. When keratinocytes
and colonic epithelial cells migrate from the dividing basal layer toward the outer lining layer [5],
the cells lose the ability to divide and eventually exofoliate from the monolayer. Anoikis also
regulates involution of mammary glands [6], and is an important step in the first cavitation of
embryogenesis [7]. Anoikis also modulates many pathological conditions. An important
characteristic of transformed cells is the loss of anchorage-dependent growth control, thereby
disrupting an essential surveillance mechanism that prevents cells from colonizing elsewhere
when they are detached from their normal residence [8][9]. The capability of escaping anoikis
27
Initially identified as major players relaying the signalling from lipid and growth factor
components in serum to actin cytoskeleton organization, Rho family GTPases have been
demonstrated to regulate a large number of biological processes in response to cell-cell and
cell-substratum adhesion [10, 11]. Rho family members regulate distinct actin cytoskeleton-based
structures; namely, Cdc42 induces filopodia, Rac1 stimulates lamellipodia formation, while RhoA
regulates the formation of stress fibers and focal adhesions [12-14]. There is considerable cross
talk between members of the Rho family, the details of which appear to depend on the cell type and
observation conditions [15]. Notably however, in most circumstances, Cdc42 appears to act
upstream of Rac1 [14].
Rho family GTPases play an important role in cell transformation [16]. Expression of
constitutively active (hydrolysis-defective) Rac1 in Rat1 fibroblasts induces serum- and
anchorage-independent growth and is tumorigenic in nude mice [17]. Cdc42 regulates
anchorage-independent growth and dominant negative Cdc42 N17 inhibits Ras focus formation
and anchorage-independent growth [18]. We have also shown that constitutively active Rac1
protects MDCK cells from anoikis, while dominant negative Rac1 potentiates anoikis in MDCK
cells [19]. In this paper, we address the role of Cdc42 in the regulation of anoikis, and examine its
28
Experimental procedures
Plasmids
Coding sequences expressing constitutively active and dominant negative Cdc42 were
amplified by PCR using pCMVneoMYC-Cdc42V12 and pCMVneoMYC-Cdc42N17 (gifts of Dr.
Arie Abo and Matt Hart, Onyx Pharmaceuticals) as a template respectively. The PCR primers
were designed so that the amplified products were tagged with a 5' EcoRI and a 3' XhoI site. The
PCR products were cloned into the EcoRI and XhoI restricted pCMV-Tag2B (Stratagene) to tag a
FLAG epitope at their amino terminals, and resulted in two intermediate plasmids,
pCF-Cdc42V12 and pCF-Cdc42N17. The FLAG-tagged Cdc42V12 coding sequence was
released from pCF- Cdc42V12 by NotI and XhoI digestion, and cloned into similarly restricted
vector, pIND(SP1), to generate pISF-Cdc42V12. By a similar strategy, pISF-Cdc42N17 was also
made. pISF-Cdc42V12 and pISF-Cdc42N17 were confirmed to express FLAG-tagged
constitutively active or dominant negative Cdc42 in an ecdysone responsive manner by
immunoblotting and immunofluorescence after they were transiently transfected into MDCK and
HEK293 cell clones which had been stably transfected with heterodimeric ecdysone receptors
system [20]. pVgRXR (Invitrogen) is used to express the heterodimeric ecdysone receptors and
confer zeocin (Invitrogen) resistance during drug selection. All the engineered plasmids were
made according to standard molecular biological techniques and were confirmed by DNA
29
Stable cell lines construction
Madin-Darby canine kidney (MDCK) cells were grown in DMEM containing 10% fetal
bovine serum at 37oC in a humidified atmosphere containing 5% CO2. Transfection was
performed using lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's
instruction. Our ultimate goal is to establish dual regulatory cell lines expressing different small
GTPase genes independently under tetracycline and ecdysone inducible systems. Therefore, we
started with a well characterized MDCK cell line expressing dominant negative Rac1 (Rac1N17)
gene under tetracycline repressible system [21, 22], and established ecdysone inducible expression
system by zeocin selection and a FACS enrichment strategy [20] in this cell line which we named
Rac1N17.1. We co-transfected 6 x 105 Rac1N17.1 cells in a p-35 well with 3.8 µg
pISF-Cdc42V12 or pISF-Cdc42N17 respectively and 0.2 µg pUB-Bsd (Invitrogen), which carried
blasticidin-S selection marker. Six hours after transfection, the cells were trypsinized and splitted
onto 6 p-100 wells with the addition of 20 ng/ml of doxycycline to suppress the expression of the
originally established (stably integrated) Rac1N17 transgene. The day after transfection, cell
culture medium was replete with 10 µg/ml of blasticidin-S for selection of stably transfected
clones. Individual candidate clones expressing FLAG-tagged Cdc42V12 or Cdc42N17 were
identified by immunofluorescence and Western blotting using anti-FLAG antibody (M2) from
Sigma. In parallel, empty pIND(SP1) vector was cotransfected and selected in the same way to
establish mock transfectant control (used to generate data described in Figure 1C). To induce the
30
concentration of ponasterone (Stratagene), and an aliquot of cells were added with the same
volume of vehicle (alcohol) as a control. To establish MDCK cell line expressing constitutively
active PI3K mutant in the background of tetracycline repressible Rac1N17 transgene, we followed
the same transfection and selection protocol as described above except we co-transfected 6 x 105
Rac1N17.1 cells in a p-35 well with 3.8 µg plasmid expressing CMV promoter driven PI3K
constitutively active mutant (p110*) and 0.2 µg pVgRXR (Invitrogen), which carried zeocin
selection marker.
Antibodies and inhibitors
Immunoblotting was performed with one of the following primary antibodies: rabbit
anti-Cdc42 and anti-p110 catalytic domain of PI3K (Santa Cruz), rabbit anti-total ERK p42/44,
rabbit anti-phosphorylated ERK p42/44 (T202/Y204), rabbit anti-total and anti- phosphorylated
JNK, rabbit anti-total and anti- phosphorylated JNK, anti- phosphorylated GSK-3 (the above
antibodies were from Cell Signaling Technology), mouse anti Rac1 (UBI), anti-myc (purified
from 9E10 hybridoma) or anti-FLAG tag (M2, Sigma). The secondary antibody used for Western
blotting analyis was either an anti-rabbit or anti-mouse HRP-linked antibody at 1:3000 dilution
(Amersham). The following small molecule inhibitors were dissolved in DMSO and used for
dissecting Cdc42V12 mediated survival signalling pathway taken by suspended cells: MEK
inhibitor U0126 (Promega), JNK inhibitor SP600125 (Tocris), p38 inhibitor SB203580, PI3K
31
used, the same volume of DMSO was also added to the control wells.
Western blot analysis
Adherent or suspended cells were washed in PBS, and cell extracts were prepared by lysing
cells in boiling SDS sample buffer (2% SDS, 150 mM NaCl, 62.5 mM Tris-HCl (pH 6.8), 10%
glycerol, 50 mM dithiothreitol, 0.01% bromophenol blue). The protein samples were separated by
SDS-PAGE and transferred to nitrocellulose membrane (Schleicher & Schuell) and the
membranes were blocked in 5% non-fat milk in PBST (0.1% Tween-20 in PBS) followed by
immunoblotting analysis. Blots were developed with ECL reagent (Amersham-Pharmacia Biotech)
for Western blotting analysis of total lysate or SuperSignal West Femto substrate (Pierce) for
GST-PAK pull-down assay.
P38 and PI 3-kinase activity assay
Immobilized phospho-p38 (Thr180/Tyr182) monoclonal antibody (Cell Signaling
Technology) and immobilized phospho-Akt (Ser473) monoclonal antibody (1G1) (Cell Signaling
Technology) were used to determine p38 and Akt activities of MDCK cells in suspension.
Extracts were made from 6 x106 cells that were induced to express transgenes for 24 hours under
adherent condition and further cultured in suspension for 2 hours. The cells were then extracted
using lysis buffer (20 mM Tris pH 7.5, 150 mM NaCl, 1mM EDTA, 1mM EGTA, 1% Triton, 2.5
mM sodium pyrophosphate, 1 mM β-glycerol phosphsate, 1 mM Na3VO4, and 1 µg/ml leupeptin)
32
overnight and for 3 hours at 4oC respectively. Immunoprecipitates of activated Akt were washed
twice with lysis buffer and twice with kinase buffer (25 mM Tris pH 7.5, 5 mM β-glycerol
phosphsate, 2 mM DTT, 0.1 mM Na3VO4, and 10 mM MgCl2). Then kinase reactions were carried
out at 30oC for 30 minutes in the presence of 200 µM ATP and 1 µg of GSK-3 to assay PKB
activity. Reactions were terminated by adding 2X SDS sample buffer and the boiled samples were
loaded onto a SDS-PAGE gel. Immunoprecipitates of activated p38 were washed three times with
lysis buffer and 2X SDS sample buffer was added before loading onto a SDS-PAGE gel. Western
blotting analysis was processed as described above and incubated with rabbit anti-phosphorylated
GSK-3 or anti-phosphorylated p38 antibody to assess the activities of Akt and p38 respectively.
p38 activity assay
Immobilized phospho-p38 (Thr180/Tyr182) monoclonal antibody (Cell Signaling
Technology) was used to determine p38 activities of MDCK cells in suspension. Extracts were
made from 6 x106 cells that were induced to express transgenes for 24 hours under adherent
condition and further cultured in suspension for 2 hours. The cells were then extracted using lysis
buffer (20 mM Tris pH 7.5, 150 mM NaCl, 1mM EDTA, 1mM EGTA, 1% Triton, 2.5 mM sodium
pyrophosphate, 1 mM β-glycerol phosphsate, 1 mM Na3VO4, and 1 µg/ml leupeptin) supplied by
the manufacturer. Immunoprecipitation of activated p38 was carried out overnight at 4oC.
Immunoprecipitates of activated p38 were washed three times with lysis buffer and 2X SDS
sample buffer was added before loading onto a SDS-PAGE gel. Western blotting analysis was
33
assess the activities of p38.
Akt activity assay
Immobilized phospho-Akt (Ser473) monoclonal antibody (1G1) (Cell Signaling
Technology) was used to determine Akt activities of MDCK cells in suspension. Extracts were
made from 6 x106 cells that were induced to express transgenes for 24 hours under adherent
condition and further cultured in suspension for 2 hours. The cells were then extracted using lysis
buffer (20 mM Tris pH 7.5, 150 mM NaCl, 1mM EDTA, 1mM EGTA, 1% Triton, 2.5 mM sodium
pyrophosphate, 1 mM β-glycerol phosphsate, 1 mM Na3VO4, and 1 µg/ml leupeptin) supplied by
the manufacturer. Immunoprecipitation of activated Akt were carried out for 3 hours at 4oC.
Immunoprecipitates of activated Akt were washed twice with lysis buffer and twice with kinase
buffer (25 mM Tris pH 7.5, 5 mM β-glycerol phosphsate, 2 mM DTT, 0.1 mM Na3VO4, and 10
mM MgCl2). Then kinase reactions were carried out at 30oC for 30 minutes in the presence of 200 µM ATP and 1 µg of GSK-3. Reactions were terminated by adding 2X SDS sample buffer and the boiled samples were loaded onto a SDS-PAGE gel. Western blotting analysis was processed as
described above and incubated with rabbit anti-phosphorylated GSK-3 to assess the activities of
Akt.
Immunofluorescence mMicroscopy
The procedure for morphological studies was the same as the one published previously [21].
The mouse anti-FLAG antibody (M2, Sigma) was used at 1 ng/ml for indirect
34
Anoikis induction and DNA fragmentation assays.
Mutant GTPases expressing MDCK cells were induced to express transgenes by adding the
indicated concentration of ponasterone (for ecdysone inducible transgenes) or removing
doxycycline (for tetracycline repressible transgenes) for 24 hours as a monolayer and subsequently
trypsinized and cultured in suspension on ultra low attachment plates (Costar) at a density of 5
x104 cells/ ml for 16-18 hours. Cells were then processed for assessing the level of DNA
fragmentation using the Cell Death ELISA kit (quantifying histone-associated DNA fragments)
using the protocol suggested by the manufacturer (Roche Molecular Biomedicals). Lysates
assayed were equivalent to 2.5 x104 cells. Each condition contained at least four independent
samples. Results were representative of at least three independent experiments, and shown as the
means with S.E.M.
Cell survival assay
10,000 MDCK cells expressing Cdc42V12 and control cells in 200 µl of culture medium
were maintained in suspension condition for 18 hours. Then, 40 µl of a modified MTT reagent,
MTS
(3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
mixed with an electron coupling reagent (phenazine ethosulfate) from Promega, was added to the
well with gentle pipetting. After incubation in the incubator at 37oC for 60 minutes, absorbance at 490 nm was recorded. In the meantime, 10,000 control cells were detached from plastic dish and
35
36
Rac1 activation assay
The procedure of preparing GST-PAK fusion protein and processing cellular lysates to
perform the pull down assay is essentially as described before [23]. In brief, the p21-binding
domain (PBD) of p21-activated kinase 1 (PAK1) was fused with glutathione-S-transferase to make
a recombinant protein (GST-PBD). The GST-PBD fusion protein was then expressed, purified,
and coupled to glutathione-sepharose beads (Amersham-Pharmacia Biotech). One twentieth of
the clarified cell lysate was immunoblotted for the corresponding GTPase specific antibody to
confirm the presence of equal loading, while the rest of the lysate was then incubated for 60 min at
37
Results
Constitutively activeRegulation of anoikis by Cdc42 suppresses, while inhibition
of Cdc42 activities enhances, MDCK cell anoikis
We choose MDCK cells, which readily undergo apoptosis when cultured in suspension, for
studying anoikis [4, 24]. In order to explore the role of Cdc42 in cell survival in anchorage
independent conditions, we generated stable MDCK cell lines that expressed constitutively active
Cdc42V12 or dominant negative Cdc42N17 under the control of an ecdysone-inducible promoter
[25]. We selected several lines for each mutant transgene, and examined the effect of regulated
expression of Cdc42V12 and Cdc42N17 on anoikis. Results were consistent among independent
clones, and were presented here with those generated from a representative clone. Addition of
ponasterone, a synthetic ecdysone analogue, induced expression of Cdc42V12 comparable to that
of endogenous Cdc42 (Fig 1A). Interestingly, the endogenous Cdc42 expression appeared to be
down-regulated by the over-expressed transgene, possibly reflecting a regulatory mechanism to
keep the activation status of Cdc42 in check (Fig. 1A).
Expression of Cdc42V12 by addition of ponasterone reduces the extent of apoptosis in
suspension conditions in a dose-dependent manner, as measured by the level of histone-associated
DNA fragments (Fig. 1B). In contrast, addition of the same concentration of ponasterone did not
have any effect on apoptosis in the parental cell line (Fig. 1B). To exclude the possibility of
38
similar doses of ponasterone into the medium of wild type MDCK, and mock transfectant controls.
No significant effect of ponasterone was observed on the anoikis of these cells (Fig. 1C), arguing
for the specificity of the effect elicited by Cdc42 activation on anoikis regulation.
To confirm that Cdc42 significantly enhances the survival potential of MDCK cells in
suspension, we determined the viability of control and Cdc42V12-expressing cells after keeping
them in suspension culture for 18 hours. We used a modified MTT
(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl) assay to quantify cell survival. Whereas only 40% of
the control cells survived in suspension conditions, up to 70% of the Cdc42V12-expressing cells
remained viable (Fig. 1D).
In contrast to the effect of Cdc42V12 on anoikis, ponasterone-regulated expression of
dominant negative Cdc42 very slightly, but reproducibly, increased apoptosis in suspension
conditions (Fig. 1ED). Inhibition of Cdc42 activity under adherent condition, however, did not
elicit any detectable difference in apoptosis in comparison to the control (Fig. 1ED). Since the
endogenous Cdc42 activity significantly declines after substratum detachment (unpublished data),
the marginal effect of Cdc42N17 on cell survival could result from the inhibition of the residual
Cdc42 activity in the suspended cells. The effects of Cdc42V12 and Cdc42N17 on the survival of
39
Cdc42 survival signaling is independent of p42/44 ERK activation
To investigate the signalling mechanisms that mediate Cdc42V12- stimulated cell survival
under anchorage-independent situation, we first examined the contribution of MAPK cascades.
Cdc42 and Rac1 have been demonstrated to synergize with Raf in activating ERKs [26, 27].
Recently, Cdc42 and Rac1 were shown to enhance association of ERK2 with MEK1 through the
activity of PAK in a fibronectin-dependent manner [28]. Furthermore, several reports indicate
activated ERK prevents anoikis [29, 30]; conversely, inhibition of Cdc42 activity has been shown
to trigger anoikis in murine fibroblasts which depend on an activated ERK activity [31].
To determine the role of ERK in Cdc42-stimulated survival signaling, we examined ERK
activity in attached and suspension conditions using the ecdysone-inducible
Cdc42V12-expressing cells. In control cells, ERK activity decreased significantly after cell
detachment, and subsequently gradually returned to baseline levels (Fig. 2A). At present, we do
not know which mechanism governs the restoration of the ERK activity after cell detachment.
Expression of Cdc42V12 did not significantly enhance ERK activiation in either attachment or
suspension conditions (Fig. 2A). In line with these observations, inhibition of MEK by U0126 did
not affect Cdc42-induced protection against anoikis, although ERK activation was nearly
abolished by the inhibitor (Fig. 2B). These results indicate that ERK activity does not play a
40
Cdc42 survival signaling is independent of JNK/SAPK pathway
The role of JNK in anoikis regulation remains controversial. Whereas JNK activation has
been shown to be required for the induction of anoikis in one study [32], this has been contradicted
by another study [24]. To investigate the role of JNK activation in the inhibition of anoikis caused
by Cdc42, we used SP600125, a reversible ATP-competitive inhibitor of JNK-1, -2, -3 [33].
Whereas SP600125 efficiently inhibited the Cdc42V12-induced JNK activation, indicated by the
diminished autophosphorylation of JNK (Fig. 3A) [34], the protective effect of Cdc42V12 against
anoikis was not affected (Fig. 3B). Notably, the application of SP600125 at 10 µM, which inhibits
JNK activation significantly, didn't have any effect on Cdc42V12 expression (Fig. 3A). We
therefore conclude the protective effect of Cdc42 on anoikis is independent of the JNK pathway
activation.
Cdc42 survival signaling is independent of p38
The role of p38 in anoikis regulation has also been controversial in that both pro-apoptotic
[35] and anti-apoptotic [36] effects have been reported. This controversy could reflect the
observation that distinct members of the p38 subfamily of mitogen-activated protein kinases may
have different functions in apoptosis [37], which needs further to be reconciled within different
cellular contexts [38]. Since p38 has been implicated in Cdc42-mediated survival signalling in
fibroblasts grown in suspension condition [36], we investigated the role of p38 in survival
41
compound specifically inhibiting p38 at micromolar concentration [39]. Since p38 can
autophosphorylate [40, 41], we examined the extent of p38 activation with a polyclonal serum
specific for phosphorylated p38, after immunoprecipitating the phosphorylated p38 with a mouse
monoclonal antibody. In the presence of 10 µM SB203580, p38 activation was significantly
suppressed, while Cdc42V12 expression still fully protected suspended MDCK cells from anoikis
(Fig. 4A and 4B). We therefor conclude that although Cdc42V12 can stimulate p38 in the absence
of cell attachment, p38 activation is dispensable for Cdc42-mediated MDCK cell survival in these
conditions.
Cdc42 mediates survival signaling in MDCK cells cultured in suspension by
activating PI3
We have shown that activated Rac1 protects MDCK cells against anoikis via activation of
PI3K [19]. To investigate whether PI3K/Akt signalling also plays a role in Cdc42-mediated
survival in suspension culture, we performed in vitro kinase assays for Akt, and observed that
Cdc42V12 significantly stimulates Akt in suspended MDCK cells (Fig. 5A). Furthermore,
LY294002, a pharmacological inhibitor of PI3K, strongly inhibited PI3K activity (Fig. 5A) as well
as the protective effect of Cdc42V12 (Fig. 5B). These results provide strong evidence for a
significant role for PI3K in the protective effect induced by Cdc42 activation in MDCK cells in
suspension.
42
Since both Cdc42 and Rac1 can inhibit anoikis via activation of PI3K, we next addressed the
question whether there is a signaling cascade from Cdc42 to Rac1 in suspended MDCK cells,
similar to the one observed in fibroblasts [14]. We used a Rac1N17 and Cdc42V12 double
expressing cell line and examined whether differential expression of these two transgenes could
affect apoptosis of MDCK cells kept in suspension condition. Fig. 6A shows the morphologyies of
this particular MDCK cell line under adherent conditions. When Cdc42V12 was induced, the cells
tended to be flatter than uninduced controls (panel b and a in Fig. 6A). This Cdc42V12 cell line
also displayed other morphological features similar to previously characterized MDCK cell lines
that express constitutively active Rac1 [21] (panel f in Fig. 6A). These included the formation of
lamellipodia and macropinocytotic vesicles [21]. Immunostaining of Cdc42V12-expressing
MDCK cells showed a spectrum of morphological characteristics from cells displaying abundant
filopodia to cells with broad lamellipodia (Fig. 6B). Between these two types of cellsextremes, we
observed cells with long protrusions embedded in sheet-like structures similar to those fibroblasts
reported to be microinjected with constitutively active Cdc42 recombinant protein [14]. The
Cdc42V12 microinjected fibroblasts first developed filopodia followed by lamellipodia
progressively growing between the pre-formed filopodia and finally coalescing into a web like
structure enclosing the majoritiesy of the filopodia [14]. When Rac1N17 was induced in addition
to the expression of Cdc42V12, the cells had an aggregated and contracted morphology similar to
the one when Rac1N17 was singly induced (panels d and c in Fig. 6A and [21]). The
43
viable, provided they are attached to substratum or embedded in a three dimensional collagen gel.
In addition, they can polarize marker proteins at specific membrane domains [21, 22].
The observations that induction of Cdc42V12 induction elicitings filipodia as well as
lamellipodia, a hallmark feature for Rac1 activation, and that dominant negative Rac1 expression
revertings these phenotypes, in MDCK cells prompted us to examine the possibility thatwhether
constitutively active Cdc42 could lead to activation of Rac1. GST-PAK fusion protein pull-down
assay demonstrated that Rac1 activity was higher when Cdc42V12 was induced in attached and
suspended MDCK cells (Fig. 6C). In addition, Cdc42 activated PI3K in a Rac1 dependent manner,
as indicated by the observation that dominant negative Rac1 abolished Akt activation by
Cdc42V12 (Fig. 6D). In addition, expression of Rac1N17 strongly inhibited the protective effect
of Cdc42V12 against anoikis (Fig. 6E). These results suggest that Rac1 acts downstream of Cdc42
in survival signalling in MDCK cells cultured in suspension.
PI3K-mediated protection against anoikis requires Rac1
While the survival signals stimulated by Rac1 in suspended cells clearly depend on PI3K
activity [19], there are many conditions where Rac1 functions downstream of PI3K [42-44]. To
further address this issue, we generate MDCK cell lines stably expressing constitutively active
PI3K activity in the background of a tetracycline repressible dominant negative Rac1 (Rac1N17)
44
catalytic sub-unit and the SH2 domain from the p85 regulatory sub-unit of PI3K. This chimeric
protein was demonstrated to activate PI3K signaling pathways independent of growth factor
stimulation [45]. Two independent MDCK clones were selected and demonstrated to express
p110* by Western blotting analysis (upper panel in Fig. 7A). It should be noted that the fusion of
p110 and the SH2 domain makes p110* migrate as a 140 kDa molecule on the PAGE, and also the
rabbit antibody we used for Western Blot could not recognize the endogenous canine form of PI3K.
Inducible expression of Rac1N17 did not affect the expression level of the CMV promoter driven
p110*, while the endogenous Rac1 expression levels seemed to be down-regulated by the
expression of Rac1N17 (Fig. 7A).
When these p110* expressing MDCK cells were cultured in suspension conditions, they were
more resistant to anoikis than the mock control (Fig. 7B) which was consistent with a previous
report [46]. This protective effect of constitutively active PI3K, however, was inhibited by the
inducible expression of Rac1N17 (Fig. 7B). This result indicates Rac1 could also function
downstream of PI3K in providing survival signals against anoikis.
Cdc42- and Rac1-mediated protection against anoikis requires an intact actin
cytoskeleton
Since both Cdc42 and Rac1 play major roles in controlling the organization of the actin
45
depends on an intact actin cytoskeleton. Latrunculin B, a potent actin polymerization inhibitor,
significantly inhibited the anoikis-protective effects of both Cdc42V12 and Rac1V12 (Fig. 87B),
suggesting an important role for Cdc42/Rac1-controlled actin polymerisation in survival
signalling by Cdc42 and Rac1. Interestingly however, the activation of PI3K by both Cdc42 and
Rac1 was not significantly affected by latrunculin B (Fig. 8A), indicating that an intact actin
cytoskeleton is largely dispensable for the Cdc42- and Rac1-induced PI3K activation.
46
Discussion
We have examined the role of Cdc42 in apoptosis induced by cell detachment from the
substratum. Our study shows that inducible expression of Cdc42V12 significantly inhibits anoikis
in MDCK cells. Our results also strongly suggest indicate that Cdc42-induced protection against
anoikis is mediated by the activation of Rac1,. In addition, our data support a model in which Rac1
and PI3K participate inwhich then initiates a positive regulatory feedback loop involving Rac1
and PI3K to inhibit anoikis. Furthermore, our data suggest that both Cdc42 and Rac1-stimulated
cell survival depends on an intact actin cytoskeleton.
Our previous [19] and present studies demonstrate that PI3K activity is critical for Rac1V12-
and Cdc42V12-mediated protection against anoikis. These observations are in line with several
studies showing that Rac1-regulated functions, including cell survival, can be inhibited by
pharmacological inhibitors of PI3K [47-49]. In addition, similar to the results that we obtained in
MDCK cells, expression of constitutively active Rac1 in hematopoietic cells can stimulate PKB in
a PI3K-dependent fashion [50]. Interestingly however, a number of earlier reports had indicated
that Rac1 could function downstream of PI3K [42-44]. It appears therefore that PI3K may function
both upstream and downstream of Rac1 (reviewed in [51]). In several cases, this could be
demonstrated in the same type of cells [50]. Based on these observations, a positive feedback loop
between Rac1 and PI3K has been postulated to function in the establishment of front-to-back