Calcium influxes and mitogen-activated protein kinase
kinase activation mediate ethylene inducing ipomoelin
gene expression in sweet potato
YU-CHI CHEN1,2*, HSIN-HUNG LIN3* & SHIH-TONG JENG3
1Department of Horticulture and Biotechnology, Chinese Culture University, Taipei, Taiwan,2Institute of Biological Chemistry, Washington State University, Pullman, WA 99164 USA; and3Institute of Plant Biology and Department of Life Science, National Taiwan University, Taipei 106, Taiwan
ABSTRACT
The ipomoelin gene (IPO) was identified to be a wound-inducible gene from Ipomoea batatas, and its expression was stimulated by methyl jasmonate (MeJA) and hydrogen peroxide. IPO protein was also characterized as a defence-related protein, and it is also a carbohydrate-binding protein. In this study, the expression of IPO was used as a molecular probe to study the effects of Ca2+on the signal transduction of ethylene. A confocal microscope monitored the Ca2+within cells, and Northern blotting examined IPO expression. The presence of Ca2+channel blocker, including diltiazem, neomycin or ruthenium red, abolished the increase of cytosolic Ca2+, and reduced the IPO expression in the cells induced by ethylene. Furthermore, both Ca2+ influxes and IPO expression stimulated by ethylene were prohibited in the presence of 10 mMethylene glycol-bis(2-aminoethyl ether)-N, N, N⬘, N⬘-tetraacetic acid (EGTA). These results indicated that Ca2+influxes into the cytosol induced by ethylene are from both apoplast and organelles, and are required for activating IPO expression. However, in the presence of 1 mM EGTA, ethylene can still stimulate
IPO expression, but mechanical wounding failed to do it.
Therefore, Ca2+channels in the plasma membrane induced by ethylene have higher affinity to Ca2+than that stimulated by wounding. Moreover, the addition of A23187, an iono-phore, raised cytosolic Ca2+, but was unable to stimulate
IPO expression. These findings showed that IPO induction
did not solely depend on Ca2+, and Ca2+elevation in cytosol is necessary but not sufficient for IPO expression. The application of PD98059, a mitogen-activated protein kinase kinase (MAPKK) inhibitor, did not prevent Ca2+ from increasing in the cytosol induced by ethylene, but inhibited the IPO expression stimulated by staurosporine (STA), a protein kinase inhibitor. Conclusively, elevation of cytosolic Ca2+by ethylene may stimulate protein phosphatase and MAPKK, which finally activates IPO expression.
Key-words: calcium ion; ethylene signalling; membrane channel.
Abbreviations: CEPA, 2-chloroethyl phosphonic acid;
EGTA, ethylene glycol-bis(2-aminoethyl ether)-N, N, N′,
N′-tetraacetic acid; IPO, ipomoelin; MAPKK,
mitogen-activated protein kinase kinase; MeJA, methyl jasmonate; OKA, okadaic acid; STA, staurosporine.
INTRODUCTION
Ethylene is produced by most plants in response to devel-opmental stages, such as seed germination, tissue dif-ferentiation, root formation and elongation, lateral bud development, flower initiation and opening, fruit ripening and leaf abscission (Jackson & Osborne 1970; Yeang & Hillman 1981; Whalen & Feldman 1988; Abeles, Morgan & Saltveit 1992; Payton et al. 1996; Deikman 1997; Lelièvre et al. 1997; Zhu et al. 2006). It also serves as signal mediator responding to environmental stresses, including flooding, wounding and pathogen infection (O’Donnell et al. 1996; Grichko & Glick 2001; Oñate-Sánchez & Singh 2002). Eth-ylene also triggered programmed cell death via calcium
ion (Ca2+), reactive oxygen species and mitogen-activated
protein kinase (MAPK) in tomato suspension cells (de Jong et al. 2002). Furthermore, Ca2+was involved in the
ethylene-mediated pathogenesis response, and the
ethylene-dependent induction of chitinase accumulation was
inhibited by blocking Ca2+influxes (Raz & Fluhr 1992).
In eukaryotic cells, Ca2+is one of the most common
sec-ondary messengers responding to extracellular stimuli (Pozzan et al. 1994; Miedema et al. 2001). In plant cells, the
Ca2+concentration increased in cytosol either as a result of
uptake from apoplastic space or of release from internal stores including vacuoles and endoplasmic reticulum (ER) (Gilroy, Read & Trewavas 1990). External signals may lead
to the opening of Ca2+channels in the plasma membrane
and in the membrane of organelles (Allen & Sanders 1994;
Ward & Schroeder 1994). In animal system, the Ca2+
-permeable channels of the plasma membrane were grouped
into three different families: voltage-operated Ca2+
channels, which include L, N, T, P/Q and R subtypes; Correspondence: S.-T. Jeng. Fax: 886-2-23918940; e-mail:
*Contributed equally to this work.
receptor-operated Ca2+ channels; and the second messenger-operated channels (Rizzuto & Pozzan 2003).
Moreover, two different intracellular Ca2+channels located
in the organelles within the cells were reported. They are the inositol 1,4,5-trisphosphate (IP3) receptors and ryanod-ine receptors (Nucifora et al. 1996). On the other hand, calcium-permeable channels have been classified on the basis of their voltage dependence into
depolarization-activated, hyperpolarization-activated and
voltage-independent cation channels in the plasma membrane of plants (White et al. 2000; Miedema et al. 2001; Sanders et al.
2002). Plants also have Ca2+channels similar to the L-type
channels characterized in animal systems by patch clamp techniques (Thuleau et al. 1994; White et al. 2000). Several
highly selective Ca2+channels activated by cytosolic second
messengers, including IP3and cyclic adenosine
diphospho-ribose (cADPR), are also present in tonoplast (Allen & Sanders 1997; White et al. 2000; Sanders et al. 2002).
Never-theless, the interaction between ethylene and Ca2+influxes
within plant cells is still not clear.
Except Ca2+mediating the signal transduction, MAPKs
were also known as intracellular signal mediators. MAPKs form a family of protein kinases that are activated in response to extracellular stimuli in eukaryotic organisms. In plants, MAPKs have been demonstrated to be an important enzyme in different types of stresses, including cold, droughts and salt stresses (Seo et al. 1995; Jonak et al. 1996; Bogre et al. 1997). MAPKs were also involved in the defence-related signal transduction (Ichimura et al. 2000; Meskiene & Hirt 2000; Tena et al. 2001; Zhang & Klessig 2001), and the wound-inducible protein kinase (WIPK) was found to respond to wounding in tobacco (Seo, Sano & Ohashi 1999). Moreover, OsMSRMK2 gene, which was cloned and demonstrated to be included in a MAPK family, was modulated by MeJA, salicylic acid, ethylene, tempera-ture, ozone and sulfur dioxide in rice (Agrawal, Rakwal & Iwahashi 2002). In Arabidopsis, MAPKs were activated by the ethylene precursor, aminocyclopropane-1-carboxylic acid, to demonstrate that the MAPK pathway mediated part of the ethylene signalling in plants (Ouaked et al. 2003). These results suggested that MAPK cascade medi-ates stress signal transductions and plays an important role in regulating the physical reactions.
In the previous study, IPO protein was characterized as a defence-related protein by the insect-feeding experiment, and it reduced both silkworm survival rate and weight gain. Based on its sequence, IPO protein is a jacalin-like lectin, and its C-terminus is highly homologous with those of mannose-binding lectin, jacalin and agglutinin (Chen et al. 2005). Lectin was known as a carbohydrate-binding protein and is widely existed in certain species. In the previous
carbohydrate-binding assay, IPO protein recognized
monosaccharides, such as glucose, galactose and mannose, rather than polygalacturonic acid and sucrose. Additionally, IPO protein strongly bound methyl glucoside
includ-ing methyl a-d-mannopyranoside and methyl
a-d-glucopyranoside. However, IPO protein cannot bind sugar alcohols like mannitol and sorbitol (Chen et al. 2003). IPO
in sweet potato was also reported to be a wound-inducible gene. The IPO mRNA was also accumulated after the stimulation of sweet potato by ethylene, MeJA and
H2O2 (Imanishi et al. 1997; Chen et al. 2003; Jih, Chen &
Jeng 2003). The signal transduction model for IPO expression in sweet potato was suggested by Chen et al. (2003). Following the stimulation by wounding, the
cytoso-lic Ca2+quickly increased, and further enhanced the release
of Ca2+ from organelles. The elevation of cytosolic Ca2+
stimulated the biosynthesis of MeJA through the octade-canoid pathway. Then, MeJA may induce the expression of IPO via dephosphorylated proteins. On the other
hand, ethylene caused the opening of Ca2+ channels in
the vacuolar membrane, and thus resulted in the activation of protein phosphatases (Chen et al. 2003). In addition, hydrogen peroxide and nitric oxide were indicated to be involved in the signal transduction of IPO in sweet potato (Jih et al. 2003). However, the signal transduction of the IPO expression induced by ethylene is not known in detail.
We report here that the influxes of cytosolic Ca2+induced by
ethylene was from the apoplastic space via the L-type Ca2+
channel in plasma membrane, and then more Ca2+released
from intracellular organelles was stimulated. Consequently,
the accumulation of Ca2+in cytosol induced the expression
of IPO via dephosphorylated protein(s) and MAPKK cascade.
MATERIALS AND METHODS
Plant materials and assay conditions
Sweet potato (Ipomoea batatas cv. Tainung 57) plants were grown in a controlled environment (16 h/25 °C day;
8 h/22 °C night; humidity 70%; light 100 mmol
photons m-2s-1). Plants with six to eight fully developed
leaves were used in this study. The second and third fully expanded leaves counted from the terminal bud were excised, and their petiole cuts were immersed in water or 1¥ Murashige–Skoog (MS) (3 mm CaCl2, 1.25 mm KH2PO4, 0.1 mm Na2EDTA, 0.1 mm FeSO4, 0.1 mm MnSO4, 0.037 mm ZnSO4, 0.1 mm CuSO4, 18.8 mm KNO3, 0.1 mm CoCl2, 30 mm
NH4Cl, 1.5 mm MgSO4and 5 mm KI) (Murashige & Skoog
1962) (pH 5.8) with the final concentration of 1, 5 or 10 mm EGTA; 0.1 mm neomycin; 0.1 mm diltiazem; 50 mm ruthe-nium red; 0.1 mm PD98059; or 1 mm A23187 under 100 mmol
photons m-2s-1for 16 h at 25 °C. Leaves with their petiole
in water or 1¥ MS without any chemical treatment were used as controls. The effect of wounding caused by the separation of leaves from the mother plants should be reduced during this long incubation period in water or MS solution. Then, the leaves were stimulated by 1 mm CEPA or 1 mm STA for another 2 h before Northern assay. The CEPA releases ethylene (Min & Bartholomew 1996), while STA inhibits protein kinase (Barwe, Sathiyabama & Jaya-baskaran 2001). Leaves were otherwise wounded by twee-zers, and Northern assays were subsequently applied 2 h later. All chemicals were purchased from Sigma (St Louis, MO, USA).
RNA isolation and analysis
Total RNA was isolated from liquid N2-ground leaves with Trizol reagent (Gibco BRL, Carlsbad, CA, USA) according to the procedure suggested by the manufacturer. The quality of RNA was analysed by agarose gel with formal-dehyde, while its quantity was estimated by a spectropho-tometer. Ten micrograms of total RNA was separated on formaldehyde–agarose gels, and the digital images of RNA gels were taken by the CCD camera with Scion Visicapture software (Scion Corporation, Frederick, MD, USA) after
gels were stained with 2 mg mL-1 ethidium bromide for
10 min. Then, the RNA gels were transferred to nylon mem-branes, and hybridized with radiolabelled probes following standard laboratory procedures (Sambrook, Fritsch & Maniatis 1989). The radiolabelled probe for IPO mRNA was produced by the PCR using the template of the full-length IPO cDNA isolated from the cDNA library in a previous study (Chen et al. 2003). Prehybridization was
per-formed in 5¥ SSPE [0.05 m NaH2PO4(pH 6.8), 0.9 m NaCl
and 5 mm ethylenediaminetetraacetic acid (EDTA)], 0.5% sodium dodecyl sulphate (SDS), 5¥ Denhardt’s solution (0.1% Ficoll, 0.1% bovine serum albumin, 0.1% polyvinyl pyrrolidine) at 60 °C for 1 h, and upon the addition of the radiolabelled probe, hybridization was performed under the same conditions for another 16 h. Blots were washed twice in 0.1¥ SSPE and 0.1% SDS at 60 °C for 15 min. Radioactive blots were displayed on the Molecular Dynam-ics Phosphoimage (Molecular DynamDynam-ics, Sunnyvale, CA, USA). Images of Northern assays were produced by Phos-phage as TIFF-formatted files, while those of ethidium-bromide-stained gels were produced by CCD camera as TIFF-formatted files as mentioned earlier. The amounts of IPO mRNA fragments and 28S rRNA in Northern and ethidium-bromide-stained gels, respectively, were then quantified by software Zero-Dscan 1.1 (BD Biosciences, Rockville, MD, USA), and their ratios of IPO mRNA to 28S rRNA, the relatively normalized mRNA values, are indi-cated in Figs 3–5.
Ca2+detection by confocal
scanning microscope
Leaves of sweet potato were cut into 0.5 ¥ 0.5 cm pieces, and floated abaxial upwardly in 1¥ MS solution with the final concentration of 1 or 10 mm EGTA, 0.1 mm neomycin, 0.1 mm diltiazem, 50 mm ruthenium red or 0.1 mm PD98059 for 16 h. The leaf pieces were then transferred to 0.55 m mannitol, which maintained the osmotic pressure of plant
cells, with fluorescent dye fluo-3 AM (Sigma F-6142) at a final concentration of 1 mm for 1 h. Fluo-3 AM, which binds
Ca2+specifically, is highly lipophilic, and easily crosses the
plasma membrane (Tsien 1981; Kao, Harootunian & Tsien 1989). The extra fluo-3 AM was removed by washing the leaf pieces with 0.55 m mannitol for 15 min (three times). Then, the addition of 1 mm CEPA or 1 mm A23187 was performed, or the mechanical wounding was applied to
ini-tiate Ca2+ influxes into cytosol. During this treatment
period, inhibition reagents always existed at a final concen-tration as described earlier. Leaf pieces without inhibition reagents were also added with 1 mm CEPA or 1 mm A23187 as control reactions. Leica TCS-SP2 confocal laser scann-ing microscope (Leica Lasertechnik GmbH, Heidelberg, Germany) with an excitation wavelength at 488 nm and emission wavelength between 500 and 535 nm was used to investigate the leaf pieces with fluo-3 AM. The focal plane of the confocal microscope in this study was always set at the epidermis that includes guard cells and epidermal cells. The images were taken every 20 s or 1 min after the images of leaf pieces were focused, and then their fluorescence intensities were recorded. Data were presented as fluores-cence intensities, whose values were directly from micro-scope (Fig. 1), or were indicated as relative fluorescences, whose values were normalized to their initial fluorescence at time zero (Fig. 2). It always took 90 s for the application of CEPA, A23187 or mechanical wounding before the leaf pieces were analysed by the confocal microscope.
RESULTS
Induction of A23187 and ethylene on the accumulation of cytosolic Ca2+
IPO in sweet potato was stimulated by mechanical wound-ing (Chen et al. 2003), and its expression induced by ethyl-ene was further studied here. The induction of ethylethyl-ene on
the accumulation of cytosolic Ca2+within the cells stained
with fluo-3 AM was investigated by a confocal scanning
microscope. To test the working system, Ca2+ influxes of
leaves, which were abaxial upwardly floated on the medium, was artificially increased by the application of ionophore A23187, which is widely used in biological systems to study
the regulatory roles of Ca2+(Rasmussen & Goodman 1997).
The fluorescence of fluo-3AM increased right after the addition of A23187 (Fig. 1a), indicating the occurrence of
Ca2+influxes within cytosol. The same working condition
was performed to study the effects of ethylene on the
accu-mulation of cellular Ca2+(Fig. 1b), and leaves treated with
Figure 1. The induction of cytolic Ca2+by A23187, ethylene and PD98059. The 0.5 ¥ 0.5 cm leaf pieces were abaxial upwardly floated in 1¥ Murashige–Skoog (MS) solution without or with PD98059 (C) for 16 h, and then the final concentration of 1 mm fluo-3 AM in 0.55 m mannitol was added for 1 h. Leaf pieces were then added with 1 mm A23187 (a) or 1 mm CEPA (b & c), and investigated by a confocal scanning microscope to analyse the appearance of cytosolic Ca2+. The focal plane of the confocal microscope was always set at the epidermis that includes guard cells and epidermal cells. The fluorescence intensity was recorded every 20 s, and the serial pictures shown here were taken at 0, 3, 6, 9, 12, 15 and 18 min after the images of cells were focused. It took 90 s for reagent application and image focusing. The fluorescence intensities read directly from microscope of this observation view versus time were showed in the last panel. Bar = 80 mm.
20 15 10 5 0 3 6 9 12 15 18 Time (min) Fluor escence intensity 20 15 10 5 0 3 6 9 12 15 18 Time (min) Fluor escence intensity 20 15 10 5 0 3 6 9 12 15 18 Time (min) Fluor escence intensity © 2007 The Authors
CEPA releasing ethylene showed a similar fluorescence
profile as that of Fig. 1a. Therefore, the elevation of Ca2+in
cytosol was rapidly induced by ethylene, and that the Ca2+
increment disappeared soon.
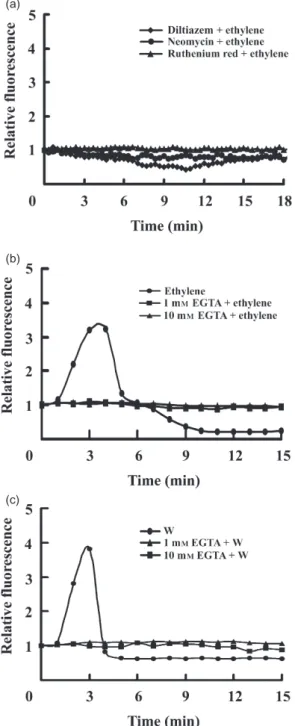
Effects of diltiazem, neomycin and ruthenium red on the accumulation of cytosolic Ca2+
induced by ethylene
Diltiazem, neomycin and ruthenium red are different types
of Ca2+ channel blockers, and they inhibit L-type Ca2+
channel in plasma membrane, IP3-mediated Ca2+channels
in organelles and cADPR-mediated Ca2+ channels in
organelles, respectively (Tsien et al. 1987; Hess 1988; Frankling-Tong et al. 1996; Durner, Wendehemme & Klessig 1998). The small pieces of sweet potato leaves pre-treated with diltiazem, neomycin or ruthenium red in the 1¥ MS buffer were probed by fluo-3 AM. The leaves were then added with 1 mm CEPA to release ethylene, and the images of these leaves were taken by a confocal scanning
micro-scope. The change of fluorescence intensity with each Ca2+
channel blocker was not significant during the investigation, demonstrating that diltiazem, neomycin and ruthenium red
inhibited ethylene inducing Ca2+ accumulation (Fig. 2a).
Therefore, Ca2+ from both apoplast and organelles is
involved in the signal transduction of ethylene. Further-more, fluorescence variations in this study were simplified into two patterns: an increase of fluorescence intensity
indi-cated the accumulation of Ca2+in cytosol, while the
fluores-cence intensity remained unchanged pointed out that Ca2+
did not enter into cytosol.
Effects of EGTA on the accumulation of cytosolic Ca2+
The influxes of intercellular Ca2+induced by ethylene were
blocked by the presence of diltiazem (Fig. 2a), indicating
the involvement of apoplastic Ca2+in ethylene stimulation.
The roles of apoplastic Ca2+during the stimulation of
eth-ylene and wounding were further studied separately in the presence of EGTA, a cation chelator. The leaf pieces of sweet potato were floated in 1¥ MS buffer with the final concentration of 1 mm or 10 mm EGTA for 16 h. After being probed by fluo-3 AM, the leaf pieces were treated with CEPA to release ethylene (Fig. 2b) or were wounded by tweezers (Fig. 2c). The emission of fluorescences caused
by the influxes of Ca2+was recorded by the confocal
micro-scope. In the absence of EGTA, the treatment of ethylene (Fig. 2b) or mechanical wounding (Fig. 2c) resulted in the
increment of Ca2+in cytosol. However, in the presence of
1 mm or 10 mm EGTA in which sweet potato looked
healthy, the increase of Ca2+in cytosol was not observed in
either ethylene or wounding treatments (Fig. 2b,c),
demon-strating that EGTA prevented apoplastic Ca2+from
enter-ing into the cells that were treated with ethylene or
wounding. Furthermore, apoplastic Ca2+is involved in the
signal tansduction of ethylene and wounding.
Involvement of Ca2+from organelles and
apoplast in IPO expression
Ethylene was able to induce the expression of IPO, and
the action of ethylene may require Ca2+. The operation of
Ca2+ channels in both plasma membrane and organelles
was required for ethylene to increase Ca2+within cytosol
(Fig. 2a,b). However, whether the expression of IPO
induced by ethylene was influenced by Ca2+ from Ca2+
channels was not clear. The petiole cuts of the excised
leaves were placed in 1¥ MS with Ca2+channel blockers
including neomycin, diltiazem or ruthenium red for 16 h before the CEPA was added to release ethylene. After another 2 h, the total RNAs of leaves were analysed by Northern blotting. The presence of ethylene alone enhanced the IPO expression (Fig. 3), but the presence of
any Ca2+signal blocker including neomycin, diltiazem or
ruthenium red significantly abolished the expression of IPO based on the relative amount of IPO mRNA (Neo, Dilt and RR in Fig. 3). These results showed that ethylene
inducing IPO expression required Ca2+released from Ca2+
channels, which were located in both organelles and plasma membrane.
Additionally, the ionophore A23187 was used to increase
the intracellular Ca2+concentration artificially and to study
the regulatory role of Ca2+in cells. The cytosolic Ca2+
con-centration was induced by A23187 within several minutes (Fig. 1a), but the application of A23187 did not induce the expression of IPO (A23 in Fig. 3). These results point out
that the induction of IPO did not solely depend on the Ca2+
increase in cytosol, and moreover the increase of Ca2+in
cytosol is necessary but not sufficient for inducing IPO expression.
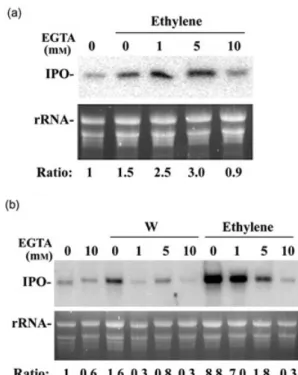
Effects of EGTA on IPO expression
To further understand the effects of apoplastic Ca2+on IPO
expression, the petiole cuts of the excised sweet potato leaves were placed in 1¥ MS or water with 1, 5 or 10 mm EGTA for 16 h, and then CEPA was added to release eth-ylene or mechanical wounding was applied with tweezers. After another 2 h, their total RNAs were analysed by Northern blotting. Figure 4a shows that in the presence of 10 mm EGTA, the expression of IPO induced by ethylene was similar to the background reaction without EGTA. Surprisingly, the activation of IPO was not inhibited in the condition with 1 or 5 mm EGTA, although the
ethylene-stimulated Ca2+influxes did not occur in the presence of
1 mm EGTA (Fig. 2b). Therefore, these results demon-strated that only high concentration of EGTA is able to
chelate the Ca2+that is participated in ethylene activating
the expression of IPO.
Solution 1¥ MS contains Ca2+, which might interfere
EGTA in chelating apoplastic Ca2+during the 16 h
incuba-tion. MS solution with 0, 1, 5 and 10 mm EGTA contains 92.68, 67.50, 7.86 and 2.65% free calcium ion, respectively, calculated by GEOCHEM PC software. Therefore, water © 2007 The Authors
instead of 1¥ MS solution was used to perform the ethylene induction and mechanical wounding assays. Again, the expression of IPO induced by ethylene was still observed in the presence of 1 or 5 mm EGTA, and the addition of 10 mm EGTA was high enough to inhibit IPO expression (Fig. 4b). Hence, high concentration of EGTA even in water is still
needed to chelate Ca2+involved in the expression of IPO
induced by ethylene. However, the presence of 1 mm EGTA was already able to abolish the expression of IPO induced by wounding (Fig. 4b), and also prohibited the increase of
Ca2+within the cytosol of the wounded cells (Fig. 2c). These
results may point out that the mechanisms of apoplastic
Ca2+entering into cytosol under the induction of ethylene
and wounding are different, and also that Ca2+affinity of the
Ca2+channels induced by ethylene is higher than that of
Ca2+channels stimulated by mechanical wounding.
Effects of PD98059 on IPO expression
PD98059 is an MAPKK inhibitor, which was widely used in biological analysis (Burnett et al. 2000), and was used to study whether the MAPK pathway was involved in the activation of IPO induced by ethylene. It was added into the incubation medium with leaf pieces for 16 h before CEPA application to release ethylene, and the confocal scanning microscope was then used to investigate the leaf pieces with
fluo-3 AM. The fluctuation of cytosolic Ca2+was observed
after the addition of CEPA in the presence of PD98059
(Fig. 1c). The relative concentration pattern of Ca2+ with
time course of the PD98059-treated leaf pieces was similar (a)
Diltiazem
(b)
(c)
Figure 2. Effects of Ca2+channel blockers and EGTA on cytosolic Ca2+induced by ethylene and mechanical wounding. The 0.5 ¥ 0.5 cm leaf pieces were abaxial upwardly floated in 1¥ Murashige–Skoog (MS) solution without or with a channel blocker (a), including 0.1 mm diltiazem, 0.1 mm neomycin or 50 mm ruthenium red, or with 1 mm (b & c) or 10 mm (b & c) EGTA for 16 h, and then transferred into 0.55 m mannitol with 1 mm fluo-3 AM for 1 h. Leaf pieces were then added with 1 mm CEPA (a & b) to release ethylene or wounded by tweezers (c) before they were investigated by the confocal microscope. The focal plane of the confocal microscope in this study was always set at the epidermis. The intensities of fluorescence were scanned by confocal microscope every 20 s (a) or 1 min (b & c). It took 90 s for reagent application and image focusing. The relative fluorescences, whose values were normalized to their initial fluorescence at time zero, versus time were shown.
Figure 3. Effects of Ca2+channel blockers and A23187 on the expression of IPO. The petiole cuts of the second and third fully expanded leaves from the terminal bud were immersed in 1¥ Murashige–Skoog (MS) with an inhibitor, including 0.1 mm neomycin (Neo), 0.1 mm diltiazem (Dilt), 50 mm ruthenium red (RR) or 0.1 mm PD98059 (PD). CEPA (1 mm) was added to release the ethylene 16 h later. In addition, leaves were put in 1¥ MS without any inhibitor for 16 h, and A23187 (A23), an ionophore, was added. Neomycin, diltiazem and ruthenium red are Ca2+channel blockers, while PD98059 is a MAPKK inhibitor. For another 2 h, the total RNAs of these leaves were analysed by Northern blotting to detect IPO mRNA. Ethidium-bromide-stained agarose gel presents ribosome RNA (rRNA) as a loading control. The ratios of IPO mRNA to rRNA in each reaction were calculated, and the ratio of the reaction treated with ethylene alone was treated as a value of one for determining the relative ratios of other reactions.
to that treated with CEPA alone (Fig. 1b). This result indi-cated that the MAPKK inhibitor, PD98059, cannot affect
the cytosolic Ca2+increment induced by ethylene.
Interest-ingly, the comparison among Fig. 1a–c also shows that the
concentration of Ca2+ in the guard cells treated with
PD98059 increased significantly once the addition of ethyl-ene (Fig. 1c).
However, the IPO expression induced by ethylene was inhibited in the presence of PD98059 (PD in Fig. 3). This result implied that PD98059 interfered with the expression
of IPO in sweet potato, while it did not prevent Ca2+from
releasing in the presence of ethylene. Therefore, MAPKK is involved in the IPO expression pathway, but the regulatory role of MAPKK in ethylene transduction pathway is in the
downstream of Ca2+accumulation in cytosol.
Signal transduction of MAPKK and dephosphorylated proteins in the IPO expression induced by ethylene
The application of OKA, a protein phosphatase inhibitor, and STA, a protein kinase inhibitor (Barwe et al. 2001), had
the opposite effects on the accumulation of IPO mRNA induced by wounding and ethylene (Chen et al. 2003). OKA inhibited the IPO expression, while STA induced it. These results indicated that IPO gene induced by wounding and ethylene was activated by dephosphorylated proteins, but inhibited via protein phosphorylation (Chen et al. 2003). Because both dephosphorylated proteins and MAPKK are important for stimulating IPO, the order of these factors participating in the signal transduction of ethylene was studied here. The leaf petiole cuttings were first placed in PD98059 solution for 16 h, then STA was added to the PD98059 solution for another 2 h before IPO mRNA was measured. The induction of IPO mRNA by STA in the presence of PD98059 was inhibited (Fig. 5). Therefore, the presence of PD98059 interfered the function of STA in stimulating IPO expression. These results also indicated that the position of MAPKK in the signal transduction of ethylene is downstream of the dephosphorylated pro-tein(s). In addition, the signal induced by ethylene is trans-ferred to dephosphorylated protein(s) first, and then passes to MAPKK to cause the activation of IPO (Fig. 6).
DISCUSSION
In this study, the signal transduction of ethylene inducing the expression of IPO was investigated. Right after the
stimulation of ethylene, Ca2+ from both apoplast and
organelles entered into cytosol in minutes. The involvement
of apoplastic Ca2+ was identified by the application of
channel blocker diltiazem, which inhibited the influxes of
Ca2+from apoplast (Fig. 2a) and decreased IPO expression
(Fig. 3). The application of cation chelator EGTA in cells
(Fig. 4a,b) also indicated the involvement of apoplastic Ca2+
Figure 4. Effects of EGTA on the expression of IPO stimulated by ethylene and wounding. The petiole cuts of the second and third fully expanded leaves from the terminal bud were immersed in 1¥ Murashige–Skoog (MS) (a) or water (b) without or with 1, 5 or 10 mm EGTA for 16 h. Leaves were added with 1 mm CEPA to release ethylene (Ethylene) or wounded by tweezers (W). The control leaves without or with 10 mm EGTA were not wounded or not added with CEPA. The total RNAs of these leaves were analysed by Northern blotting to detect IPO mRNA. Ethidium-bromide-stained agarose gel presents ribosome RNA (rRNA) as a loading control. The ratios of IPO mRNA to rRNA in each reaction were calculated, and the ratio of the reaction without any treatment was treated as a value of one for determining the relative ratios of other reactions.
Figure 5. Effects of PD98059 on the expression of IPO stimulated by STA. The petiole cuts of the second and third fully expanded leaves from the terminal bud were immersed in 0.1 mm PD98059, an MAPKK inhibitor, for 16 h. The final concentration of 1 mm STA, a protein kinase inhibitor, was then added for another 2 h. The control leaves were incubated separately in 1 mm STA or 0.1 mm PD98059 alone. Their total RNAs were analysed by Northern blotting to detect IPO mRNA (IPO). Ethidium-bromide-stained agarose gel presents ribosome RNA (rRNA) as a loading control. The ratios of IPO mRNA to rRNA in each reaction were calculated, and the ratio of the reaction treated with STA alone was treated as a value of one for determining the relative ratios of other reactions.
during ethylene activation. Neomycin and ruthenium red
blocking IP3-mediated and cADPR-mediated Ca2+channels
in organelles, separately, also inhibited the entry of Ca2+
from organells (Fig. 2a) and decreased the expression of IPO (Fig. 3), indicating the involvement of organelles’ Ca2+ in the ethylene signal transduction for inducing IPO. The
accumulation of Ca2+in the cytosol induced by ethylene
may stimulate protein dephosphorylation, which further activated MAPKK (Fig. 5), and then induced the expres-sion of IPO (Fig. 6).
IPO protein is a defence-related protein by the insect-feeding experiment, and it decreased silkworm survival rate. IPO protein is a lectin based on its sequence, and its C-terminus is highly homologous with those of mannose-binding lectin, jacalin and agglutinin (Chen et al. 2003). In the previous study, IPO protein is a carbohydrate-binding protein. However, IPO protein cannot bind sugar alcohols like mannitol and sorbitol (Chen et al. 2003). Regarding the gene expression, accumulation of IPO mRNA appeared 2 h, peaked at 6 h and then continuously declined up to 16 h after wounding the sweet potato leaves (Chen et al. 2003). The expression of IPO was also induced by the stimulus of MeJA (Imanishi et al. 1997; Chen et al. 2003). The influences
of Ca2+, dephosphorylated proteins, H2O2and nitric oxide
on IPO expression were also studied (Chen et al. 2003; Jih
et al. 2003). This study further investigates the signal trans-duction of ethylene in inducing the expression of IPO in sweet potato.
During the signal transduction induced by external
stimuli, Ca2+is always considered to be the first response
messenger. Monitoring the quick accumulation of cytosolic
Ca2+ by confocal microscope is a convenient method to
investigate the activation of cells. However, the increase of
Ca2+in cytosol is very fast, and it did take 90 s for sample
treatment and microscope focusing in order to record the
change of Ca2+by confocal microscope. Consequently, the
increase of Ca2+in cells may not be recorded thoroughly,
and only the second half of the increasing peak was
docu-mented by the confocal microscope (Fig. 1a–c). The Ca2+
rising pattern of the cells treated with A23187, which is an
ionophore allowing Ca2+to influxes into cytosol, was a
half-peak shape monitored by confocal microscope (Fig. 1a).
Therefore, the Ca2+rising pattern of the cells induced by
ethylene showed the same half-peak shape, and was
consid-ered to be the consequence of Ca2+ influxes into cells
(Fig. 1b). However, the full Ca2+ induction peak was
observed in Fig. 2b, indicating Ca2+accumulation in the cells
stimulated by ethylene initiated at the different time points. The variations of plant tissues used in each assay were used to explain this phenomenon. Conclusively, fluorescence variations in this study were simplified into two patterns.
The increase in fluorescence indicates an increment of Ca2+
in cytosol, while the unchanged fluorescence intensity with
time demonstrates that the concentration of Ca2+in cytosol
is unaltered.
Antagonists to Ca2+channels have been frequently used
to characterize transport processes. To understand the
effects of Ca2+in the signal transduction of ethylene, Ca2+
channel blockers and chelators were used in this study.
Ruthenium red and neomycin were used to block the Ca2+
release from internal ER and vacuoles. Ruthenium red was treated as a cADPR-mediated channel blocker in this study. The synthesis of the second messenger cADPR was induc-ible by nitric oxide in animal cells, and cADPR activated
Ca2+release from the specific subset of membrane vesicles
(Wendehenne et al. 2001). In addition, cADPR in plant cells
was able to induce Ca2+ release from ER and vacuoles
(Allen, Muir & Sanders 1995; Leckie et al. 1998). In tobacco cells, cADPR induced PR-1 and PAL gene expression, and the induction of cADPR was inhibited by ruthenium red (Durner et al. 1998). On the other hand, neomycin was able to inhibit phospholipase C (Frankling-Tong et al. 1996), which broke down phosphatidyl inositol 4,5-bisphosphate
into IP3 that was responsible for the release of Ca2+from
intracellular stores (Berridge 1984). Neomycin was used
previously as a phospholipase C inhibitor to study Ca2+
metabolism in plant cells (Frankling-Tong et al. 1996; Knight, Trewavas & Knight 1997). Hence, neomycin in this study was used as a channel blocker, which inhibited IP3-mediated channel. In addition, diltiazem was used as an inhibitor to block L-type channel, which is one of the
voltage-operated Ca2+channels in plasma membrane (Tsien
et al. 1987; Hess 1988).
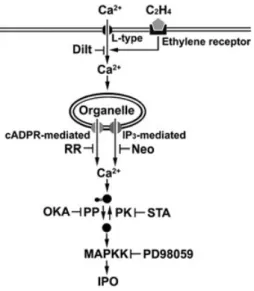
Figure 6. Model for the signal transduction of ethylene activating IPO in sweet potato. Ethylene binding to its receptor allows the influxes of apoplastic Ca2+into cytosol through the L-type voltage Ca2+channel in the plasma membrane, and then activates the release of Ca2+via inositol 1,4,5-trisphosphate (IP3)-mediated and cyclic adenosine diphosphoribose (cADPR)-mediated Ca2+channels in organelle membrane. The elevation of cytosolic Ca2+may stimulate the protein
dephosphorylation, and then the signal was passed through MAPKK to induce the IPO expression. Diltiazem (Dilt) blocks the L-type voltage-operated Ca2+channel, neomycin (Neo) blocks the IP3-mediated Ca2+channel, whereas ruthenium red (RR) blocks the cADPR-mediated Ca2+channel. STA inhibits protein kinase (PK), OKA inhibits protein phosphatase (PP) and PD98059 inhibits MAPKK. The arrows represent induction, while the blunted lines indicate repression.
The cation chelator EGTA, which bound apoplastic
Ca2+, was applied to the reaction in order to further
confirm the roles of apoplastic Ca2+during ethylene
acti-vation. In the previous study, IPO treated by mechanical
wounding induced both the influxes of apoplastic Ca2+and
the release of organelles’ Ca2+. Nevertheless, the
ethylene-dependent IPO expression was reported to be regulated
by the release of Ca2+ from intracellular stores only
because neomycin but not EGTA (1 mm) inhibited the expression of IPO induced by ethylene (Chen et al. 2003). The same results showed in this study that in the presence of 1 mm EGTA, ethylene can still induce IPO expression
(Fig. 4a,b), but the increase of cytosolic Ca2+ was not
detected by confocal microscope (Fig. 2b). However, in
the same concentration of 1 mm EGTA, both Ca2+influxes
(Fig. 2c) and IPO expression (Fig. 4b) were inhibited in the cells treated with mechanical wounding. The high con-centration of EGTA (10 mm) in both ethylene- and
wounding-stimulated cells was needed to prevent Ca2+
from entering into cytosol (Fig. 2b,c), and reduced the expression of IPO (Fig. 4a,b). Hence, both signal transduc-tion pathways induced by ethylene and wounding did
require apoplastic Ca2+to activate IPO, but the nature of
Ca2+channels induced by ethylene and wounding was
dif-ferent. The possible reason for 1 mm EGTA not inhibiting the ethylene-induced IPO but repressing the wound-induced IPO is that ethylene and wounding triggered the
expression of IPO via different plasma membrane Ca2+
channels that may have different affinity with Ca2+.
Because only high concentration of EGTA can retard the
activation of ethylene, Ca2+channels induced by ethylene
may have higher affinity to Ca2+than those stimulated by
wounding. Therefore, after harvesting of plants, high con-centration of EGTA is required to delay the action of eth-ylene. At the same time, the presence of 1 mm EGTA can block the induction of mechanical wounding, but it still allows the stimulation of ethylene.
Ca2+ influxes into cytosol are mainly from apoplastic
space and internal organelles, but Ca2+ from different
source for activating genes was reported. Ca2+antagonists,
including EGTA (up to 10 mm), lanthanum, verapamil and ruthenium red, were used to test the involvement of the
apoplastic and the organelles’ Ca2+ in the ethylene
responses of Pisum sativum (Petruzzelli et al. 2003). Results
indicated that Ca2+release from organelles, but not
apoplas-tic space, was involved in activating PS-ACO1 gene, and
also that both the apoplastic and the organelles’ Ca2+were
required for negatively regulating PS-ACS2 gene and posi-tively activating class 1 b-1,3-glucanase (Petruzzelli et al.
2003). However, our study indicated that Ca2+from both
apoplast and organelles was involved in the activation of IPO induced by ethylene or wounding (Figs 3 & 4).
Additionally, Ca2+is an important secondary messenger
and regulates the complicated gene expression in both animals and plants. The concentration and localization
of Ca2+ were responsible for the specific regulation of
Ca2+between stimuli and responses (Sanders et al. 2002).
The Ca2+ channel was also regulated by Ca2+ feedback
regulation. The cADPR-dependent Ca2+channels opened
at physiological trans-tonoplast voltages, but repressed
when the Ca2+ concentration was greater than 600 nm
(Leckie et al. 1998). Therefore, even though the cytosolic
Ca2+was increased largely by the application of A23187,
the expression of IPO did not occur (Fig. 3). This result
further indicated that the unsuitable increase of Ca2+
cannot turn on transcription, at least the transcription of IPO gene.
MAPK plays an important role in signal transcription in all eukaryotic cells, and it represents the convergence point for many signalling pathways and modulates a variety of cellular events. Mechanical manipulation of Arabidopsis leaves induced the gene expressions of MAPKKK and MAPK (Bogre et al. 1996, 1997). MAPK and other protein kinases were induced by touch, cold and water stress within 30 min in Arabidopsis (Mizoguchi et al. 1996). Wounding alfalfa leaves specifically induces the transient activation of
the p44MMK4 kinase, which belongs to the MAPK family
(Bogre et al. 1997). These evidences displayed MAPK pathway mediating the plant stress signal, and thus it was worth finding out the regulation of MAPKs within the eth-ylene transduction pathway inducing the IPO expression in sweet potato. PD98059, a MAPKK inhibitor, blocked the expression of IPO induced by ethylene (Fig. 3), but it did
not inhibit the influxes of Ca2+in the cells treated by
ethyl-ene (Fig. 1c). These results showed that MAPKK was involved in the ethylene transduction activating IPO, and
its action was located after the Ca2+accumulation. STA, a
protein kinase inhibitor, was demonstrated to induce IPO expression (Fig. 5), while OKA, a protein phosphatase inhibitor, decreased the IPO expression activated by ethyl-ene (Chen et al. 2003). However, leaves of sweet potato were treated with PD98059 before STA was added, and its IPO expression was still reduced as those treated with PD98059 alone (Fig. 5). These results indicated that MAPKK was involved in the ethylene-induced signal pathway, and the reaction order of MAPKK in the ethylene transduction pathway was behind the dephosphorylated proteins (Fig. 6).
Conclusively, the ethylene transduction pathway induc-ing the expression of IPO is shown in Fig. 6. Followinduc-ing
ethylene stimulation, Ca2+ from the intercellular space
quickly enters into cytosol via L-type voltage-operated Ca2+
channel in the plasma membrane , and it further activates
the IP3-mediated and cADPR-mediated Ca2+channels in
organelles to release Ca2+to enter into cytosol. The
eleva-tion of cytosolic Ca2+may stimulate the protein
dephospho-rylation, which further activates MAPKK, and finally, IPO gene is induced.
ACKNOWLEDGMENTS
This work was supported by the National Science Council under grant 95-2313-B-002-011 to S.-T.J. We wish to thank Technology Commons, College of Life Science, National Taiwan University for technical support.
REFERENCES
Abeles F.B., Morgan P.W. & Saltveit M.E., Jr (1992) Ethylene in Plant Biology. Academic Press, San Diego, CA, USA. Agrawal G.K., Rakwal R. & Iwahashi H. (2002) Isolation of novel
rice (Oryza sativa L) multiple stress responsive MAP kinase gene, OsMSRMK2, whose mRNA accumulates rapidly in response to environmental cues. Biochemical and Biophysical Research Communications 294, 1009–1016.
Allen G.J. & Sanders D. (1994) Two voltage-gated, calcium-release channels coreside in the vacuolar membrane of broad bean guard cells. Plant Cell 6, 685–694.
Allen G.J. & Sanders D. (1997) Vacuolar ion channels in higher plants. Advances in Botanical Research 25, 217–252.
Allen G.J., Muir S.R. & Sanders D. (1995) Release of Ca2+from individual plant vacuoles by both InsP3 and cyclic ADP-ribose. Science 268, 735–737.
Barwe S.P., Sathiyabama M. & Jayabaskaran C. (2001) Induction of chitinase activity by exogenous cytokinins in excised dark-grown cucumber cotyledons: involvement of Ca2+and staurosporine-sensitive protein kinase(s) in cytokinin signaling. Journal of Plant Physiology 158, 1–7.
Berridge M.J. (1984) Inositol triphosphate and diacylglycerol as secondary messengers. Biochemical Journal 220, 345–360. Bogre L., Ligterink W., Heberle-Bors E. & Hirt H. (1996)
Mecha-nosensors in plants. Nature 383, 489–490.
Bogre L., Ligterink W., Meskienne I., Barker P.J., Heberle-Bors E., Huskisson N.S. & Hirt H. (1997) Wounding induces the rapid and transient activation of a specific MAP kinase pathway. Plant Cell 9, 75–83.
Burnett E.C., Desikan R., Moser R.C. & Neill S.J. (2000) ABA activation of an MBP kinase in Pisum sativum epidermal peels correlates with stomatal responses to ABA. Journal of Experi-mental Botany 51, 197–205.
Chen Y.C., Tseng B.W., Huang Y.L., Liu Y.C. & Jeng S.T. (2003) Expression of the ipomoelin gene from sweet potato is regulated by dephosphorylated proteins, calcium ion, and ethylene. Plant, Cell & Environment 26, 1373–1383.
Chen Y.C., Chang H.S., Lai H.M. & Jeng S.T. (2005) Characteriza-tion of the wound-inducible protein ipomoelin from sweet potato. Plant, Cell & Environment 28, 251–259.
Deikman J. (1997) Molecular mechanisms of ethylene regulation of gene transcription. Physiologia Plantarum 100, 561– 566.
Durner J., Wendehemme D. & Klessig D.F. (1998) Defense gene induction in tobacco by nitric oxide, cyclic GMP and cyclic ADP-ribose. Proceedings of the National Academy of Sciences of the United States of America 95, 10328–10333.
Frankling-Tong V.E., Droback B.K., Allan A.C., Watkins P.A.C. & Travawas A.J. (1996) Growth of pollen tubes of Papaver rhoeas is regulated by a slow-moving calcium wave propagated by inosi-tol 1,4,5-triphosphate. Plant Cell 8, 1305–1321.
Gilroy S., Read N.D. & Trewavas A.J. (1990) Elevation of cyto-plasma calcium by caged calcium or caged inositol triphosphate initiate stomatol closure. Nature 346, 769–771.
Grichko V.P. & Glick B.R. (2001) Ethylene and flooding stress in plants. Plant Physiology and Biochemistry 39, 1–9.
Hess P. (1988) Elementary properties of cardiac calcium channels: a brief review. Canadian Journal of Physiology and Pharmacol-ogy 66, 1218–1223.
Ichimura K., Mizoguchi T., Yoshida R., Yuasa T. & Shinozaki K. (2000) Protein phosphorylation and dephosphorylation in envi-ronmental stress responses in plants. Advances in Botanical Research 32, 355–377.
Imanishi S., Kito-Nakamura K., Matsuoka K., Morikami A. & Nakamura K. (1997) A major jasmonate-inducible protein of
sweet potato, ipomoelin, is an ABA-independent wound-inducible protein. Plant & Cell Physiology 38, 643–652. Jackson M.B. & Osborne D.J. (1970) Ethylene, the natural
regula-tor of leaf abscission. Nature 225, 1019–1022.
Jih P.J., Chen Y.C. & Jeng S.T. (2003) Involvement of hydrogen peroxide and nitric oxide in expression of the ipomoelin gene from sweet potato. Plant Physiology 132, 381–389.
Jonak C., Kiegerl S., Ligterink W., Barker P.J., Huskisson N.S. & Hirt H. (1996) Stress signaling in plants: a MAP kinase pathway is activated by cold and drought. Proceedings of the National Academy of Sciences of the United States of America 93, 11274– 11279.
de Jong A.J., Yakimova E.T., Kapchina V.M. & Woltering E.J. (2002) A critical role for ethylene in hydrogen peroxide release during programmed cell death in tomato suspension cells. Planta
214, 537–545.
Kao J.P.Y., Harootunian A.T. & Tsien R.Y. (1989) Photochemically generated cytosolic calcium pulses and their detection by fluo-3. Journal of Biological Chemistry 264, 8179–8184.
Knight H., Trewavas A.J. & Knight M.R. (1997) Calcium signaling in Arabidopsis thaliana responding to drought and salinity. Plant Journal 12, 1067–1078.
Leckie C.P., McAinsh M.R., Allen G.J., Sanders D. & Hetherington A.M. (1998) Abscisic acid-induced stomatal closure mediated by cyclic ADP-ribose. Proceedings of the National Academy of Sciences of the United States of America 95, 15837–15842.
Lelièvre J.M., Latché A., Jones B., Bouzayen M. & Pech J.C. (1997) Ethylene and fruit ripening. Physiologia Plantarum 101, 727– 739.
Meskiene I. & Hirt H. (2000) MAP kinase pathways: molecular plug-and-play chips for the cell. Plant Molecular Biology 42, 791–806.
Miedema H., Bothwell J.H.F., Brownlee C. & Davies J.M. (2001) Calcium uptake by plant cells: channels and pumps acting in concert. Trends in Plant Science 6, 514–519.
Min X.J. & Bartholomew D.P. (1996) Effect of plant growth regu-lators on ethylene production, 1-aminocyclopropane-1-carboxylic acid oxidase activity, and initiation of inflorescence development of pineapple. Journal of Plant Growth Regulator
15, 121–128.
Mizoguchi T., Irie K., Hayashida N., Yamaguchi-Shinozaki K., Mat-sumoto K. & Shinizaki K. (1996) A gene encoding a mitogen-activated protein kinase and S6 ribosomal protein kinase by touch, cold, and water stress in Arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America 93, 765–769.
Murashige T. & Skoog F. (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15, 473–497.
Nucifora F.C., Jr, Sharp A.H., Milgram S.L. & Ross C.A. (1996) Inositol 1,4,5-trisphosphate receptors in endocrine cells: localiza-tion and associalocaliza-tion in hetero- and homotetramers. Molecular Biology of the Cell 7, 949–960.
O’Donnell P.J., Calvert C., Atzorn R., Wasternack C., Leyser H.M.O. & Bowles D.J. (1996) Ethylene as a signal mediating the wound response of tomato plants. Science 274, 1914–1917. Oñate-Sánchez L. & Singh K.B. (2002) Identification of
Arabidop-sis ethylene-responsive element binding factors with distinct induction kinetics after pathogen infection. Plant Physiology
128, 1313–1322.
Ouaked F., Rozhon W., Lecourieux D. & Hirt H. (2003) A MAPK pathway mediates ethylene signaling in plants. EMBO Journal
22, 1282–1288.
Payton S., Fray R.G., Brown S. & Grierson D. (1996) Ethylene receptor expression is regulated during fruit ripening, flower © 2007 The Authors
senescence and abscission. Plant Molecular Biology 31, 1227– 1231.
Petruzzelli L., Sturaro M., Mainieri D. & Leubner-Metzger G. (2003) Calcium requirement for ethylene-dependent responses involving 1-aminocyclopropane-1-carboxylic acid oxidase in radicle tissues of germinated pea seeds. Plant, Cell & Environ-ment 26, 661–671.
Pozzan T., Rizzuto R., Volpe P. & Meldolesi J. (1994) Molecular and cellular physiology of intracellular calcium stores. Physiological Reviews 74, 595–636.
Rasmussen H. & Goodman D.B.P. (1997) Relationships between calcium and cyclic nucleotides in cell activation. Physiological Reviews 57, 421–509.
Raz V. & Fluhr R. (1992) Calcium requirement for ethylene-dependent response. Plant Cell 4, 1123–1130.
Rizzuto R. & Pozzan T. (2003) When calcium goes wrong: genetic alterations of a ubiquitous signaling route. Nature Genetics 34, 135–141.
Sambrook J., Fritsch E.F. & Maniatis T. (1989) Molecular Cloning: A Laboratory Manual, 2nd edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, USA.
Sanders D., Pelloux J., Brownlee C. & Harper J. (2002) Calcium at the crossroads of signaling. Plant Cell 14, S401–S417.
Seo S., Okamoto M., Seto H., Ishizuka K., Sano H. & Ohashi Y. (1995) Tobacco MAP kinase: a possible mediator in wound signal transduction pathway. Science 270, 1988–1992.
Seo S., Sano H. & Ohashi Y. (1999) Jasmonate-based wound signal transduction requires activation of WIPK, a tobacco mitogen-activated protein kinase. Plant Cell 11, 289–298.
Tena G., Asai T., Chiu W.L. & Sheen J. (2001) Plant mitogen acti-vated protein kinase signaling cascades. Current Opinion in Plant Biology 4, 392–400.
Thuleau P., Ward J.M., Ranjeva R. & Schroeder J.I. (1994)
Voltage-dependent calcium-permeable channels in the plasma membrane of a higher plant cell. EMBO Journal 13, 2970– 2975.
Tsien R.Y. (1981) A non-disruptive technique for loading calcium buffers and indicators into cells. Nature 290, 527–528.
Tsien R.W., Hess E., McCleskey E.W. & Rosenberg R.L. (1987) Calcium channels: mechanism of selectivity, permeation, and block. Annual Review of Biophysics and Biophysical Chemistry
16, 265–290.
Ward J.M. & Schroeder J.I. (1994) Calcium activated K+channels and calcium-induced calcium release by slow vacuolar ion chan-nels in guard cell vacuoles implicated in the control of stomatal closure. Plant Cell 6, 669–683.
Wendehenne D., Pugin A., Klessig D.F. & Durner J. (2001) Nitric oxide: comparative synthesis and signalling in animal and plant cells. Trends in Plant Science 6, 177–183.
Whalen M.C. & Feldman L.J. (1988) The effect of ethylene on root growth of Zea mays seedlings. Canadian Journal of Botany 66, 719–723.
White P.J., Pineros M., Tester M. & Ridout M.S. (2000) Cation permeability and selectivity of a root plasma membrane calcium channel. Journal of Membrane Biology 174, 71–83.
Yeang H.Y. & Hillman J.R. (1981) Control of lateral bud growth in Phaseolus vulgaris L. by ethylene in the apical shoot. Journal of Experimental Botany 32, 395–404.
Zhang S. & Klessig D.F. (2001) MAPK cascades in plant defense signaling. Trends in Plant Science 6, 520–527.
Zhu C., Gan L., Shen Z. & Xia K. (2006) Interactions between jasmonates and ethylene in the regulation of root hair develop-ment in Arabidopsis. Journal of Experidevelop-mental Botany 57, 1299– 1308.
Received 17 July 2007; accepted for publication 10 September 2007