R E S E A R C H A R T I C L E
Open Access
Role of the small RNA RyhB in the Fur regulon in
mediating the capsular polysaccharide
biosynthesis and iron acquisition systems in
Klebsiella pneumoniae
Su-Hua Huang
1†, Chien-Kuo Wang
1†, Hwei-Ling Peng
2, Chien-Chen Wu
2, Ying-Tsong Chen
3,4,5, Yi-Ming Hong
6and
Ching-Ting Lin
6*Abstract
Background: The capsular polysaccharide (CPS) and iron acquisition systems are important determinants of Klebsiella pneumoniae infections, and we have previously reported that the ferric uptake repressor (Fur) can play dual role in iron acquisition and CPS biosynthesis. In many bacteria, Fur negatively controls the transcription of the small non-coding RNA RyhB to modulate cellular functions and virulence. However, in K. pneumoniae, the role played by RyhB in the Fur regulon has not been characterised. This study investigated Fur regulation of ryhB transcription and the functional role of RyhB in K. pneumoniae.
Results: Deletion of fur from K. pneumoniae increased the transcription of ryhB; the electric mobility shift assay and the Fur-titration assay revealed that Fur could bind to the promoter region of ryhB, suggesting that Fur directly represses ryhB transcription. Additionally, in aΔfur strain with elevated CPS production, deletion of ryhB obviously reduced CPS production. The following promoter-reporter assay and quantitative real-time PCR of cps genes verified that RyhB activated orf1 and orf16 transcription to elevate CPS production. However, deletion of ryhB did not affect the mRNA levels of rcsA, rmpA, or rmpA2. These results imply that Fur represses the transcription of ryhB to mediate the biosynthesis of CPS, which is independent of RcsA, RmpA, and RmpA2. In addition, theΔfur strain’s high level of serum resistance was attenuated by the deletion of ryhB, indicating that RyhB plays a positive role in protecting the bacterium from serum killing. Finally, deletion of ryhB inΔfur reduced the expression of several genes corresponding to 3 iron acquisition systems in K. pneumoniae, and resulted in reduced siderophore production.
Conclusions: The regulation and functional role of RyhB in K. pneumoniae is characterized in this study. RyhB participates in Fur regulon to modulate the bacterial CPS biosynthesis and iron acquisition systems in K. pneumoniae.
Keywords: RyhB, Fur, Capsular polysaccharide, Iron acquisition system, Klebsiella pneumoniae
* Correspondence:[email protected]
†Equal contributors 6
School of Chinese Medicine, China Medical University, Taichung 40402, Taiwan
Full list of author information is available at the end of the article
© 2012 Huang et al.; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Background
Klebsiella pneumoniae, a member of Enterobacteriaceae, is a rod-shaped gram-negative opportunistic pathogen. A common cause of nosocomial infection, it is also found in various community-acquired infections, including bacter-aemia, septicbacter-aemia, and urinary tract and respiratory infections, particularly in immunocompromised patients [1-4]. In Asian countries, especially Taiwan and Korea, K. pneumoniae is the predominant pathogen found in pyo-genic liver abscess in diabetic patients [2,3,5]. The rapid development of antimicrobial resistance in K. pneumoniae has further troubled the clinical choices for treatments [6,7]. Studies of the pathogenic mechanisms of K. pneu-moniae are, therefore, essential in identifying new targets for the development of antibacterial agents.
Multiple virulence factors have been identified to be involved in K. pneumoniae infection, which include capsular polysaccharide (CPS), lipopolysaccharides, fim-briae, iron-acquisition system, and antibiotic resistance. Among these factors, CPS is probably considered the major determinants of pathogenesis. The pyogenic liver abscess isolates often carry heavy CPS that could pro-tect the bacteria from phagocytosis and killing by serum factors [8,9]. Apart from the antiphagocytic function, Klebsiella CPS also helps the bacterial colonization and biofilm formation at the infection sites [10-12]. The capsular serotypes of K. pneumoniae have been classified as more than 77 recognized capsu-lar antigens [13,14]. In Taiwan, a high prevalence of K1 and K2 serotypes of K. pneumoniae was documen-ted in liver abscess of diabetes mellitus patients [15]. The cps gene clusters that are responsible for the syn-thesis of different serotypes of CPS have been deter-mined [16]. The K2 cps gene cluster of K. pneumoniae Chedid contains a total number of 19 open reading frames (ORFs) organized into three transcription units, orf1-2, orf3-15, and orf16-17 [16]. In the previous stud-ies, numerous regulatory systems were demonstrated to control the biosynthesis of CPS via regulating the cps transcriptions in K. pneumoniae, such as the Rcs system, RmpA, RmpA2, KvhR, KvgAS, and KvhAS [17-20]. Among these, ferric uptake regulator (Fur) represses the gene expression of rcsA, rmpA, and rmpA2 to decrease CPS biosynthesis [21,22]. Therefore, overlapping regulons governed the regulation of these assorted virulence genes in response to numerous stress conditions.
Bacterial cells are constantly challenged by various en-vironmental stresses from their natural habitats. Similar to many gastrointestinal (GI) pathogens, K. pneumoniae faces several challenges during infection and colonisa-tion of the human body. These include gastric acid, the immune system, and a limited supply of oxygen and nutrients [23,24]. Among these, the concentration of
iron in the environment is critical for the control of cel-lular metabolism. Limitation of iron abolishes bacterial growth, but high intracellular concentrations of iron may damage bacteria because of the formation of un-desired reactive oxygen species (ROS). Iron homeostasis maintained by the transport, storage, and metabolism of iron is tightly controlled by Fur in many gram-negative bacteria [25-27]. To regulate gene transcription, Fur pro-tein functions as a dimer with Fe2+as a cofactor to bind to a 19-bp consensus sequence, called the Fur box (GATAATGATwATCATTATC; w = A or T), in the pro-moters of downstream genes [28]. In several gram-negative pathogens, Fur represses the expression of genes involved in iron homeostasis and in the regulation of multiple cellular functions such as oxidative stress, energy metabolism, acid tolerance, and virulence gene production [29-32]. In K. pneumoniae, Fur plays a dual role in controlling CPS biosynthesis and iron acquisition [21]. Recently, we also found that type 3 fimbriae expres-sion and bacterial biofilm formation were also controlled by Fur and iron availability [33]. Therefore, the regula-tory mechanism of Fur in control of multiple cellular function and virulence factors in K. pneumoniae needs to be further investigated.
Although Fur typically acts as a repressor, it also functions as a transcriptional activator for the gene ex-pression such as acnA, fumA, and sdhCDAB (tricarb-oxylic acid [TCA] cycle enzymes), bfr and ftnA (iron storage), and sodB (iron superoxide dismutase [FeSOD]) [34-38]. However, positive regulation by Fur is often indirect, mediated by Fur-dependent repression of a small non-coding RNA (sRNA), RyhB [39]. RyhB negatively regulates gene expression by base pairing with mRNAs to trigger their degradation via RNase E and RNase III [40]. In many bacteria, RyhB participates in Fur-mediated positive regulation of various import-ant cellular functions, including TCA cycle activity, resistance to oxidative stress, and iron homeostasis in Escherichia coli and Vibrio cholerae [35,39,41-43]; bio-film formation in V. cholerae [44]; and virulence in Shigella dysenteriae [45]. In E. coli, RyhB has been demonstrated to directly regulate more than 18 tran-scripts, encoding a total of 56 proteins, most of them involved in iron metabolism [35]. Although the signifi-cance of RyhB has been demonstrated in different spe-cies, to date, the regulatory relationship of RyhB and Fur, and functionality of RyhB in K. pneumoniae has not been studied.
In this study, the regulatory role of Fur in ryhB expres-sion in K. pneumoniae was investigated. A ryhB-deletion mutant in wild type (WT) and Δfur strains and the induced expression of ryhB in WT were generated to demonstrate the role of RyhB in mediating CPS biosyn-thesis and iron acquisition systems.
Results
Fur directly repressesryhB expression in K. pneumoniae
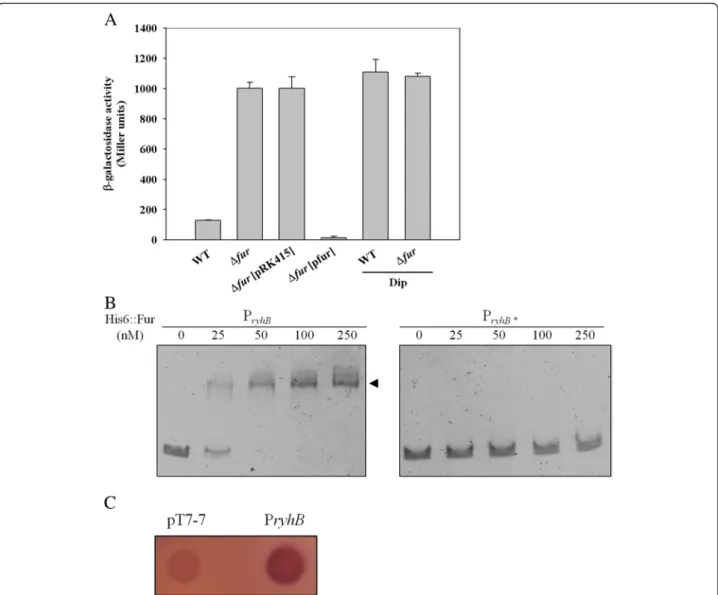
To determine whether K. pneumoniae ryhB is regulated by Fur, a LacZ reporter system was used. The ryhB pro-moter was cloned into the upstream region of a propro-moter- promoter-less lacZ gene in placZ15. The resulting plasmid pRyhB15 was then introduced into K. pneumoniae CG43S3ΔlacZ and ΔlacZΔfur. The bacterial β-galactosidase activity was measured to assess the expression level of ryhB. As shown in Figure 1A, the expression of ryhB was higher in ΔlacZΔfur than ΔlacZ. Introduction of the complement plasmid pfur, but not the empty vector control (pRK415),
intoΔlacZΔfur restored the Fur-deletion effect. Moreover, addition of the iron chelator 2, 2-dipyridyl (Dip) to the growth medium increased ryhB promoter activity, sug-gesting that a Fur-Fe(II) complex influences ryhB expres-sion. To verify that Fur directly regulates the expression of ryhB, an electrophoretic mobility shift assay (EMSA) was performed. As shown in Figure 1B, purified recombinant His6-Fur protein was able to bind the upstream region of
ryhB(PryhB), but not the PryhB* fragment, whose putative
Fur-box was deleted. In addition, the binding ability was abolished by the addition of 200μM EDTA to the reaction mixture (data not shown). Furthermore, E. coli H1717,
Figure 1 Fur directly represses the expression ofryhB. (A) The β-galactosidase activities of the K. pneumoniae CG43S3ΔlacZ strain and the isogenic fur deletion mutant carrying pRyhB15 (PryhB::lacZ) were determined from overnight cultures grown in LB with or without Dip. The plasmids pRK415 (vector control) and pfur were introduced intoΔfur to observe the complement effect. The average of triplicate experiments is shown. Error bars indicate standard deviations. (B) EMSA of the recombinant His6::Fur and the ryhB promoter regions, as indicated in the margin. DNA was incubated with an increasing amount of His6::Fur for 30 min, and then loaded onto a 5% non-denaturing polyacrylamide gel. The gel was stained with SYBR Green EMSA stain and photographed. PryhB* indicates deletion of the fur box in PryhB. (C) Assessment of the binding of Fur to the ryhB promoter by using the FURTA. E. coli H1717 strains carrying the vector control, pT7-7, or the P1 region harboured on pT7-7 are indicated. A red colony (Lac+) is considered to have a FURTA-positive phenotype.
when harbouring a plasmid containing K. pneumoniae PryhB, also showed a Fur titration assay (FURTA)-positive
phenotype (Figure 1C). The results suggest that, in an iron dependent manner, Fur suppresses ryhB promoter activity in K. pneumoniae by direct interaction with the Fur-box region upstream of ryhB.
RyhB activates CPS biosynthesis
In K. pneumoniae CG43, we found that the deletion of fur resulted in elevated CPS production [21,22]. To in-vestigate if RyhB participates in Fur-regulated CPS bio-synthesis, the CPS amount was assessed using measuring glucuronic acid content, which served as an indicator for KlebsiellaK2 CPS [46], in K. pneumoniae strains, includ-ing WT,ΔryhB, Δfur, and ΔfurΔryhB, was quantified. As shown in Figure 2A, although the deletion of ryhB alone did not change on the amount of K2 CPS production, the elevated CPS amount inΔfur cells was abolished by the deletion of ryhB when the bacteria were grown in LB medium. The result indicates that Fur regulates the ex-pression of RyhB to repress CPS biosynthesis. To confirm the RyhB expression could activate the CPS biosynthesis, the effect of RyhB induction on CPS amount was deter-mined using an IPTG-inducible vector, pETQ. As shown in Figure 2B, the induced expression of ryhB in K. pneu-moniaeCG43 increased CPS production, which confirms that RyhB positively regulates CPS biosynthesis.
RyhB increased the transcriptional level of the K2cps gene cluster
To investigate whether RyhB affects the expression of the three cps gene clusters, the mRNA levels of orf1, orf3, and orf16 inΔfur and ΔfurΔryhB strains were mea-sured by quantitative real-time PCR (qRT-PCR). As
shown in Figure 3A, compared to the mRNA levels in the Δfur strain, the mRNA levels of orf1 and orf16 were apparent decreased in the ΔfurΔryhB strain, and that of orf3also had a slight reduction in the ΔfurΔryhB strain. The result suggests that overexpression of RyhB acti-vated the cps gene expression. To confirm our hypoth-esis, the effect of ryhB induction on the mRNA levels of orf1, orf3, and orf16 was tested using an IPTG-inducible vector, pETQ. As shown in Figure 3B, the mRNA levels of orf1 and orf16 were higher in the pETQ-ryhB strain with IPTG induction than the pETQ mock strain, while no significant difference in orf3 expression was observed. To further investigate whether RyhB acts as a transcrip-tional activator for the promoter activity of orf1, orf3, and orf16, the reporter plasmids pOrf12 (Porf1-2::lacZ),
pOrf315 (Porf3-15::lacZ), and pOrf1617 (Porf16-17::lacZ),
each carrying a lacZ reporter gene transcriptionally fused to the putative promoter region of the K2 cps gene cluster [17], were used to transform the K. pneumoniae strains CG43S3ΔlacZΔfur and ΔlacZΔfurΔryhB. The promoter activity measurements shown in Figure 3C revealed that the deletion of ryhB in ΔlacZΔfur reduced activity of Porf1-2::lacZ by at least 50%, while no obvious
change was detected in the activity of Porf3-16::lacZ. The
activity of Porf16-17::lacZ was reduced by more than 75%
in ΔlacZΔfurΔryhB as compared to the ΔlacZΔfur strain. These results imply that RyhB enhances CPS bio-synthesis in K. pneumoniae by boosting the transcrip-tional level of the orf1 and orf16 gene clusters.
RyhB does not affect thercsA, rmpA2, and rmpA mRNA expression level
In previous studies, K. pneumoniae Fur was found to re-press the exre-pression of genes encoding the cps
Figure 2 RyhB activates CPS biosynthesis. (A) Comparison of CPS levels in WT,ΔryhB, Δfur, and ΔfurΔryhB strains. Bacterial strains were grown in LB medium at 37°C with agitation. After 16 h of growth, the bacterial glucuronic acid content was determined. *, P< 0.001 compared with WT. (B) WT strains carrying the vector control (pETQ) or pETQ-ryhB were grown in LB with 100μM IPTG to induce ryhB expression. *, P < 0.001 compared with WT strains carrying pETQ.
regulatory proteins RcsA, RmpA, and RmpA2 [21,22]. To investigate whether RyhB affects the expression of rcsA, rmpA, and rmpA2 to increase the orf1 and orf16 transcripts, the mRNA levels were measured by qRT-PCR after inducing the expression of ryhB in WT. How-ever, qRT-PCR results did not show a significant effect of ryhB on the mRNA levels of rmpA, rmpA2, and rcsA (Data not shown), suggesting that the activation of RyhB on the orf1 and orf16 expression is not via RmpA, RmpA2, and RcsA.
Deletion ofryhB attenuated the higher serum resistance inΔfur strain
In addition to the roles played by RyhB and Fur in regu-lating the CPS amount, we suggest that RyhB and Fur may also affect the ability of the strain to resist the bac-tericidal effects of serum. In a human serum resistance assay, we found that the deletion of fur in WT increased the survival rate in treatment with 75% normal human serum from 63.3% to 87.9% (Figure 4). However, the de-letion of ryhB in WT had no apparent effect on the sur-vival rate on treatment with 75% serum, and the higher serum resistance inΔfur cells was abolished by the dele-tion of ryhB. This result indicates that RyhB may partici-pate with Fur in regulating serum resistance in K. pneumoniae.
The regulatory role of RyhB in iron-acquisition systems
To assess whether RyhB affects iron-acquisition in K. pneumoniae, the Chrome azurol S (CAS) assay was used to measure siderophore secretions in Δfur and Δfur-ΔryhB strains (Figure 5). When bacteria were grown in M9 minimal medium (~2 μM iron) to mimic iron-limited condition, the deletion of ryhB in Δfur reduced the formation of the orange halo. However, this change
was not observed when bacteria were grown in LB
medium (~18 μM iron). Compared to M9 minimal
medium contains ~2μM iron, LB medium is considered an iron-repletion medium. Under iron-repletion, Fur is able to exert its repression on ryhB transcription. Thus, ryhB-deletion effect is difficult to observed under the growth condition that ryhB is poorly expressed. Our results suggest that in the regulation of iron-acquisition systems, RyhB plays a role downstream of Fur in K. pneumoniaeunder iron-limiting conditions.
To investigate the effects on downstream targets of RyhB in iron-acquisition regulons, the expression of genes corresponding to the eight putative iron-acquisition systems in K. pneumoniae CG43 was mea-sured in Δfur and ΔfurΔryhB by qRT-PCR (Table 1). In Figure 3 RyhB activates the transcriptional level of theorf1 and orf16. (A) qRT-PCR analyses of the expression of the K2 cps genes (orf1, orf3, and orf16) were measured inΔfur and ΔfurΔryhB strains. (B) WT strain carrying the IPTG inducible vector pETQ and pETQ-ryhB in response to 100μM IPTG. (C) The β-galactosidase activities of K. pneumoniae CG43S3ΔlacZΔfur and ΔlacZΔfurΔryhB carrying the reporter plasmid pOrf12 (Porf1-2::lacZ), pOrf315 (Porf3-15::lacZ) or pOrf1617 (Porf16-17::lacZ) were determined using log-phased cultures grown in LB broth. The results shown are an average of triplicate samples. Error bars indicate standard deviations.
Survival percentage (%) 0 20 40 60 80 100
WT ΔryhB Δfur Δfur
ΔryhB ΔgalU
Figure 4 Effect of Fur and RyhB on susceptibility to normal human serum. Survival percentage of WT,ΔryhB, Δfur, ΔfurΔryhB, andΔgalU (negative control) strains on treatment with 75% healthy human serum was determined, respectively. The results shown are an average of triplicate samples. Error bars indicate standard deviations.
M9 minimal medium, the expression of genes (iucA, fepA, fepB, entC, fecA, and fecE) corresponding to three iron-acquisition systems (aerobactin, enterobactin, and ferric citrate) was decreased by half in the ΔfurΔryhB strain (ΔfurΔryhB/Δfur < 0.5). However, the expression of fhuA and sitA was significantly increased more than two-fold (ΔfurΔryhB/Δfur > 2.0). These results imply
that RyhB activates the expression of iucA, fepA, fepB, entC, fecA, and fecE, but represses the expression of fhuAand sitA.
Discussion
In this study, we provide an initial characterisation of K. pneumoniae RyhB. In K. pneumoniae, sequence com-parison indicated that the nucleotide sequence of the ryhBgene (91 bp) is 92.3% identical to the E. coli version (90 bp). However, the promoter sequence of K. pneumo-niae ryhBis only 72.4% identical to that of E. coli. In this study, we found that the expression of ryhB in K. pneu-moniaeis directly repressed by Fur-Fe(II), as is the case in E. coli (Figure 1).
In addition, structure of the genomic neighbourhood of ryhB differs between the 2 species. In the E. coli gen-ome, ryhB is found between yhhX and yhhY. In the K. pneumoniae genome, ryhB is flanked by yhhY and a hypothetical ORF. By Pfam search, the hypothetical ORF was found to contain a bactofilin domain (E-value = 3.7 e-24), which belongs to a new class of polymer-forming proteins that serve as versatile molecular scaffolds in a variety of cellular pathways [47]. Even though the func-tion of this hypothetical protein in K. pneumoniae has not yet been investigated, we found that RyhB could strongly repress the expression of this hypothetical pro-tein (unpublished data). This result suggests that RyhB could participate in a variety of cellular pathways in K. pneumoniae.
We previously showed in K. pneumoniae, Fur
represses CPS biosynthesis via regulation of RmpA, RmpA2, and RcsA. In addition to these 3 regulators, one or more regulators may be involved in the Fur-mediated control of cps transcription [21]. In this study, we found that RyhB also participates in Fur-regulated CPS biosyn-thesis via activation of orf1 and orf16 transcription and is independent of the 3 regulators, RmpA, RmpA2, and RcsA (Figure 2 and 3). We want to further analyse whether any potential transcriptional regulator-binding motifs exist in the promoter sequences of orf1 and orf16. We noted that a binding site typical of IscR, a transcrip-tional repressor that controls Fe–S biosynthesis [48], was located 172 bp upstream of the translation start site of GalF (encoded by orf1, 5′-ATAACCTGAACGAAAA TAAGATTAT-3′). The predication indicated that IscR could participate in control of orf1 expression. Further-more, a previous study reported that RyhB promotes the degradation of iscSUA transcripts, resulting in an in-crease in the ratio of apo-IscR/holo-IscR [48]. Whether RyhB activates CPS biosynthesis via regulation of the ratio of apo-IscR/holo-IscR in K. pneumoniae awaits fur-ther analysis. However, the regulatory mechanism of cps transcription is more complex than expected; whether another unknown transcriptional regulator is involved in Figure 5 Deletion ofryhB decreases K. pneumoniae Δfur
siderophore production assessed using CAS assay. Each of the strains,Δfur and ΔfurΔryhB, was grown overnight in LB medium or M9 minimal medium, and then 5μl each of cultures respectively was added onto a CAS agar plate. The orange halos formed around the colonies correspond to the iron-chelating activity of the siderophores in bacteria.
Table 1 qRT-PCR analyses of the expression of iron-acquisition genes inK. pneumoniae Δfur ΔryhB and Δfur strains
Systems Gene RNA expression ratioa
ΔfurΔryhB/Δfur Fe3+ Ferrichrome fhuA 2.62 ± 0.07 Aerobactin iucA 0.19 ± 0.06 Enterobactin fepA 0.36 ± 0.01 fepB 0.33 ± 0.05 entC 0.46 ± 0.02
Ferric citrate fecA 0.19 ± 0.02
fecE 0.34 ± 0.03
Salmochelin iroB 0.52 ± 0.05
Heme hmuR 0.69 ± 0.01
Fe2+
Ferrous iron feoB 0.55 ± 0.18
sitA 2.81 ± 0.08
a
activation of RyhB’s effect on orf16 transcription needs to be investigated. In addition, CPS is considered the major determinant that can protect the bacteria from phagocytosis and killing by serum factors [8,9]. In this study, higher serum resistance was found in Δfur, but this higher serum resistance was attenuated by further deletion of ryhB (Figure 4). We suggest the protective role of RyhB against serum killing is due to the activa-tion of CPS biosynthesis.
In E. coli, RyhB plays a positive role in control of the intracellular iron concentration via the degradation of nonessential iron-using proteins or an increase in sidero-phore production [49-51]. In this study, we also found the deletion of ryhB inΔfur decreased siderophore pro-duction on the CAS plate under iron-limiting condition (Figure 5). Consistent with E. coli [51], RyhB in K. pneu-moniae regulates siderophore production by activating the expression of enterobactin system genes (entC, fepA, and fepB). In addition, we found that RyhB may activate iucA and fecA expression. Since sRNA may positively regulate its target mRNAs via an anti-antisense mechan-ism to disrupt an intrinsic inhibitory structure in the 5′ mRNA region that sequesters the ribosome-binding site and the first translation codon [52,53], the 5′-untranslated regions of the iuc and fec operons were analysed for sequences complementary to RyhB by pre-diction with the bioinformatics application RNAhybrid [54] (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/ submission.html). However, no apparent base pairing was found in the 5′-untranslated region of the iuc or fec operons, suggesting that the activation of iucA and fecA by RyhB is not a result of direct interaction. Further-more, RyhB was found to repress the expression of fhuA and sitA in K. pneumoniae. In E. coli, RyhB represses the expression of fhuA, which also corresponds to our results [35]. A possible paring between RyhB with the adjacent sequence of translational start site of fhuA and sitA was also predicted by the RNAhybrid algorithm. Alignment of the protected residues predicts that RyhB forms a 7 + 4 + 4 bp RNA duplex with the sitA mRNA (Additional file 1: Figure S1), but no apparent base pairing was found between RyhB and fhuA. However, the direct interaction of RyhB with the sitA mRNA remains to be confirmed. In E. coli, RyhB has been shown to repress several genes that are involved in iron-binding, which may increase the intracellular iron con-centration, thereby allowing a better usage of iron and more complete Fur repression of these genes [35,55]. Nevertheless, this possibility in K. pneumoniae needs to be proven by careful experiments. In this study, the coordinated action of Fur and RyhB was found to regu-late the expression of the iron acquisition systems for maintaining intracellular iron homeostasis in K. pneumoniae.
Conclusions
In this study, we provide an initial characterisation of K. pneumoniae RyhB. Our results suggest that RyhB plays an important role in the Fur regulon, which modulates the CPS biosynthesis and iron acquisition systems in K. pneumoniae, both of which contribute to the infectivity and survival of the bacterium.
Methods
Bacterial strains, plasmids, and media
Bacterial strains and plasmids used in this study are listed in Table 2. Primers used in this study are list in Additional file 2: Table S1. Bacterial were routinely cul-tured at 37°C in Luria-Bertani (LB) medium or M9 min-imal medium supplemented with appropriate antibiotics. The antibiotics used include ampicillin (100 μg/ml), kanamycin (25 μg/ml), streptomycin (500 μg/ml), and tetracycline (12.5μg/ml).
Construction of the gene-deletion mutants
Specific gene deletion was introduced into K. pneumo-niaeCG43S3 using an allelic exchange strategy as previ-ously described [57]. The pKAS46 system was used in the selection of the mutants [59], and the mutations were respectively confirmed by PCR and Southern hybridization (data not shown).
Measurement of promoter activity
The promoter region of ryhB was PCR-amplified with primer pair pGT44/pGT45, and the amplicons were then cloned into placZ15 [63]. The promoter-reporter plasmids, pRyhB15, pOrf12, pOrf315, and pOrf1617, were individually mobilized into K. pneumoniae strains by conjugation from E. coli S17-1 λpir. The bacteria were grown to logarithmic phase in LB broth with or without 200 μM Dip (OD600 of 0.7), and the
β-galactosidase activity was measured as previously described [63].
EMSA
Recombinant K. pneumoniae Fur protein was expressed in E. coli and purified as previously described [22]. DNA fragment of the putative promoter region of ryhB was respectively PCR amplified by using specific primer sets (Table 2). The purified His6-Fur was incubated with
10-ng DNA in a 15μl solution containing 50 mM Tris–HCl (pH 7.5), 100 mM NaCl, 100 mM dithiothreitol, 200μM MnCl2, and 1 μg/μl BSA at room temperature for
20 min. The samples were then loaded onto a native gel of 5% nondenaturing polyacrylamide containing 5% gly-cerol in 0.5× TB buffer (45 mM Tris–HCl, pH 8.0, 45 mM boric acid). Gels were electrophoresed with a 20-mA current at 4°C and then stained with SABR safe Gel stain (Invitrogen).
FURTA
FURTA was performed according to the method described by Stojiljkovic et al. [64]. DNA sequences con-taining a putative Fur box were PCR amplified with spe-cific primer sets and then cloned into pT7-7. The resulting plasmids were introduced into the E. coli strain H1717, and the transformants were plated onto MacConkey-lactose plates containing 100μg/ml ampicil-lin and 30 μM Fe(NH4)2(SO4)2. The indicator strain
H1717 contained a chromosomal fhuF::lacZ fusion, and a low affinity Fur box has been demonstrated in the fhuF promoter. The introduction of pT7-7 derived plasmids carrying Fur-binding sequences could thus cause the re-moval of Fur from the fhuF Fur box [60]. H1717 harbor-ing pT7-7 was used as a negative control. Colony
phenotype was observed after incubation at 37°C for 10 h. Red colony (Lac+) denoted a FURTA-positive phenotype and indicated the binding of Fur to the DNA sequence cloned into the pT7-7 plasmid.
Extraction and quantification of CPS
CPS was extracted and quantified as previously described [65]. The glucuronic acid content, represents the amount of K. pneumoniae K2 CPS, was determined from a standard curve of glucuronic acid (Sigma-Aldrich) and expressed as micrograms per 109CFU [46].
qRT-PCR
Total RNAs were isolated from early-exponential-phase grown bacteria cells by use of the RNeasy midi-column
Table 2 Bacterial strains and plasmids used in this study
Strains or plasmids
Descriptions Reference or source
K. pneumoniae
CG43S3 CG43 Smr [56]
ΔlacZ CG43S3ΔlacZ [17]
Δfur CG43S3Δfur [22]
ΔlacZΔfur CG43S3ΔlacZΔfur [22]
ΔryhB CG43S3ΔryhB This study
ΔfurΔryhB CG43S3ΔfurΔryhB This study
ΔlacZΔfurΔryhB CG43S3ΔlacZΔfurΔryhB This study
ΔgalU CG43S3ΔgalU [57]
E. coli
DH5α supE44ΔlacU169 (f80 lacZΔμ15) hsdR17 recA1 endA1 gyrA96 thi-1 relA1 [58]
BL21-RIL F-ompT hsdS
B[rB-mB-]gal dcm [DE3] Laboratory stock
S17-1λ pir H1717 hsdR recA pro RP4-2 [Tc::Mu; Km::Tn7] [λpir] araD139ΔlacU169 rpsL150 relA1 flbB5301 deoC1 ptsF25 rbsR aroB fhuF::λ placMu
[59,60]
Plasmids
pKAS46 Positive selection suicide vector, rpsL AprKmr [59]
yT&A TA cloning vector Yeastern
pRK415 Broad-host-range IncP cloning vector, Tcr [61]
pT7-7 Cloning vector, Apr [62]
pETQ Kmr, protein expression vector [61]
placZ15 Cmr, promoter selection vector, lacZ+ [17]
pfur Tcr, 0.8-kb fragment containing a fur allele cloned into pRK415 [22]
pET30c-Fur Kmr, 450-bp fragment encoding full-length Fur cloned into pET30c [22]
pRyhB04 2.0 kb fragment containing an internal ~70-bp deletion in ryhB cloned into pKAS46 This study pRyhB15 Cmr, 178-bp fragment containing the region upstream of ryhB cloned into placZ15 This study
pOrf12 Cmr, 500-bp fragment containing the region upstream of Klebsiella K2 cps orf1-orf2 cloned into placZ15 [17]
pOrf315 Cmr, 900-bp fragment containing the region upstream of Klebsiella K2 cps orf3-orf15 cloned into placZ15 [17]
pOrf1617 Cmr, 300-bp fragment containing the region upstream of Klebsiella K2 cps orf16-orf17 cloned into placZ15 [17]
pT7-7-pryhB 178-bp fragment containing the putative ryhB promoter, cloned into pT7-7 This study
(QIAGEN) according to the manufacturer’s instructions. RNA was DNase-treated with RNase-free DNase I (MoBioPlus) to eliminate DNA contamination. RNA of 100 ng was reverse-transcribed with the Transcriptor First Strand cDNA Synthesis Kit (Roche) using random primers. qRT-PCR was performed in a Roche LightCy-clerW 1.5 Instrument using LightCycler TaqMan Master (Roche). Primers and probes were designed for selected target sequences using Universal ProbeLibrary Assay De-sign Center (Roche-applied science) and listed in Add-itional file 2: Table S1. Data were analyzed using the real time PCR software of Roche LightCyclerW 1.5 Instru-ment. Relative gene expressions were quantified using the comparative threshold cycle 2-ΔΔCT method with 23S rRNA as the endogenous reference.
Bacterial survival in serum
Normal human serum, pooled from healthy volunteers, was divided into equal volumes and stored at −70°C be-fore use. Bacterial survival in serum was determined with minor modifications [57]. First, The bacteria were grown to log phase in LB broth and the viable bacterial concentration was adjusted to 1 × 106 colony forming units/ml. 1 ml of the cultures was washed twice by using phosphate-buffered saline (PBS) and resuspended in 1 ml PBS. The mixture containing 250μl of the cell sus-pension and 750 μl of pooled human serum was incu-bated at 37°C for 60 min. The number of viable bacteria was then determined by plate counting. The survival rate was expressed as the number of viable bacteria treated with human serum compared to the number of pre-treatment. The assay was performed triple, each with triplicate samples. The data from one of the representa-tive experiments are shown and expressed as the mean and standard deviation from the three samples. The 0% survival of K. pneumoniae CG43S3ΔgalU served as a negative control.
CAS assay
The CAS assay was performed according to the method described by Schwyn and Neilands [66]. Each of the bac-terial strain was grown overnight in M9 minimal medium, and then 5μl of culture was added onto a CAS agar plate. After 24 hr incubation at 37°C, effects of the bacterial siderophore production could be observed. Sid-erophore production was apparent as an orange halo around the colonies; absence of a halo indicated the in-ability to produce siderophores.
Statistical method
An unpaired t-test was used to determine the statistical significance and values of P< 0.001 were considered significant. The results of CPS quantification and qRT-PCR analysis were derived from a single experiment
representative of three independent experiments. Each sample was assayed in triplicate and the mean activity and standard deviation are presented.
Additional files
Additional file 1: Figure S1. RyhB pairs with sitA. The file contains supplemental figure S1 that the potential base pairing in RyhB/sitA mRNA in this study.
Additional file 2: Table S1. Primers used in this study. The file contains supplemental Table S1 that the detailed information of primer sets used in this study.
Competing interests
The authors declare that they have no competing interests. Acknowledgements
The work is supported by the grants from National Science Council (NSC 97-2314-B-039-042-MY2 and NSC 99-2320-B-039-002-MY3) and China Medical University (CMU98-ASIA-01 and CMU99-ASIA-07).
Author details
1
Department of Biotechnology, Asia University, Taichung 41354, Taiwan.
2Department of Biological Science and Technology, National Chiao Tung
University, Hsin Chu 30068, Taiwan.3Institute of Genomics and Bioinformatics, National Chung Hsing University, Tai Chung City 40227, Taiwan.4Biotechnology Center, National Chung Hsing University, Tai Chung City 40227, Taiwan.5Institute of Molecular and Genomic Medicine, National
Health Research Institutes, Miaoli County 35053, Taiwan.6School of Chinese Medicine, China Medical University, Taichung 40402, Taiwan.
Authors’ contributions
SHH, CKW, HLP, and CTL made substantial contributions to design and conduct the experiments. YMH performed qRT-PCR and growth experiments. SHH and CKW performed the bioinformatics analyses and interpretation of data. CCW, YTC, and HLP contributed to the writing and editing of the manuscript. CTL coordinated the study and performed manuscript editing. All authors have read and approved this work.
Received: 15 May 2012 Accepted: 9 July 2012 Published: 24 July 2012
References
1. Chou FF, Kou HK: Endogenous endophthalmitis associated with pyogenic hepatic abscess. J Am Coll Surg 1996, 182(1):33–36.
2. Han SH: Review of hepatic abscess from Klebsiella pneumoniae. An association with diabetes mellitus and septic endophthalmitis. West J Med 1995, 162(3):220–224.
3. Lau YJ, Hu BS, Wu WL, Lin YH, Chang HY, Shi ZY: Identification of a major cluster of Klebsiella pneumoniae isolates from patients with liver abscess in Taiwan. J Clin Microbiol 2000, 38(1):412–414.
4. Peng HL, Wang PY, Wu JL, Chiu CT, Chang HY: Molecular epidemiology of Klebsiella pneumoniae. Zhonghua Min Guo Wei Sheng Wu Ji Mian Yi Xue Za Zhi 1991, 24(3):264–271.
5. Yang YS, Siu LK, Yeh KM, Fung CP, Huang SJ, Hung HC, Lin JC, Chang FY: Recurrent Klebsiella pneumoniae liver abscess: clinical and
microbiological characteristics. J Clin Microbiol 2009, 47(10):3336–3339. 6. Paterson DL: Resistance in gram-negative bacteria: enterobacteriaceae.
Am J Med 2006, 119(6 Suppl 1):S20–S28. discussion S62-70.
7. Lagamayo EN: Antimicrobial resistance in major pathogens of hospital-acquired pneumonia in Asian countries. Am J Infect Control 2008, 36(4 Suppl):S101–S108.
8. Sahly H, Podschun R, Oelschlaeger TA, Greiwe M, Parolis H, Hasty D, Kekow J, Ullmann U, Ofek I, Sela S: Capsule impedes adhesion to and invasion of epithelial cells by Klebsiella pneumoniae. Infect Immun 2000,
68(12):6744–6749.
9. Lin JC, Chang FY, Fung CP, Xu JZ, Cheng HP, Wang JJ, Huang LY, Siu LK: High prevalence of phagocytic-resistant capsular serotypes of
Klebsiella pneumoniae in liver abscess. Microbes Infect 2004, 6(13):1191–1198.
10. Boddicker JD, Anderson RA, Jagnow J, Clegg S: Signature-tagged mutagenesis of Klebsiella pneumoniae to identify genes that influence biofilm formation on extracellular matrix material. Infect Immun 2006, 74 (8):4590–4597.
11. Moranta D, Regueiro V, March C, Llobet E, Margareto J, Larrate E, Garmendia J, Bengoechea JA: Klebsiella pneumoniae capsule polysaccharide impedes the expression of beta-defensins by airway epithelial cells. Infect Immun 2010, 78(3):1135–1146.
12. Favre-Bonte S, Joly B, Forestier C: Consequences of reduction of Klebsiella pneumoniae capsule expression on interactions of this bacterium with epithelial cells. Infect Immun 1999, 67(2):554–561.
13. Fung CP, Hu BS, Chang FY, Lee SC, Kuo BI, Ho M, Siu LK, Liu CY: A 5-year study of the seroepidemiology of Klebsiella pneumoniae: high prevalence of capsular serotype K1 in Taiwan and implication for vaccine efficacy. J Infect Dis 2000, 181(6):2075–2079.
14. Pan YJ, Fang HC, Yang HC, Lin TL, Hsieh PF, Tsai FC, Keynan Y, Wang JT: Capsular polysaccharide synthesis regions in Klebsiella pneumoniae serotype K57 and a new capsular serotype. J Clin Microbiol 2008, 46 (7):2231–2240.
15. Fung CP, Chang FY, Lee SC, Hu BS, Kuo BI, Liu CY, Ho M, Siu LK: A global emerging disease of Klebsiella pneumoniae liver abscess: is serotype K1 an important factor for complicated endophthalmitis? Gut 2002, 50 (3):420–424.
16. Arakawa Y, Wacharotayankun R, Nagatsuka T, Ito H, Kato N, Ohta M: Genomic organization of the Klebsiella pneumoniae cps region responsible for serotype K2 capsular polysaccharide synthesis in the virulent strain Chedid. J Bacteriol 1995, 177(7):1788–1796.
17. Lin CT, Huang TY, Liang WC, Peng HL: Homologous response regulators KvgA, KvhA and KvhR regulate the synthesis of capsular polysaccharide in Klebsiella pneumoniae CG43 in a coordinated manner. J Biochem (Tokyo) 2006, 140(3):429–438.
18. Majdalani N, Gottesman S: The Rcs phosphorelay: a complex signal transduction system. Annu Rev Microbiol 2005, 59:379–405.
19. Gottesman S, Stout V: Regulation of capsular polysaccharide synthesis in Escherichia coli K12. Mol Microbiol 1991, 5(7):1599–1606.
20. Stout V: Regulation of capsule synthesis includes interactions of the RcsC/RcsB regulatory pair. Res Microbiol 1994, 145(5–6):389–392. 21. Lin CT, Wu CC, Chen YS, Lai YC, Chi C, Lin JC, Chen Y, Peng HL: Fur
regulation of the capsular polysaccharide biosynthesis and iron-acquisition systems in Klebsiella pneumoniae CG43. Microbiology 2011, 157(Pt 2):419–429.
22. Cheng HY, Chen YS, Wu CY, Chang HY, Lai YC, Peng HL: RmpA regulation of capsular polysaccharide biosynthesis in Klebsiella pneumoniae CG43. J Bacteriol 2010, 192(12):3144–3158.
23. De Champs C, Sauvant MP, Chanal C, Sirot D, Gazuy N, Malhuret R, Baguet JC, Sirot J: Prospective survey of colonization and infection caused by expanded-spectrum-beta-lactamase-producing members of the family Enterobacteriaceae in an intensive care unit. J Clin Microbiol 1989, 27 (12):2887–2890.
24. Markowitz SM, Veazey JM Jr, Macrina FL, Mayhall CG, Lamb VA: Sequential outbreaks of infection due to Klebsiella pneumoniae in a neonatal intensive care unit: implication of a conjugative R plasmid. J Infect Dis 1980, 142(1):106–112.
25. Ernst JF, Bennett RL, Rothfield LI: Constitutive expression of the iron-enterochelin and ferrichrome uptake systems in a mutant strain of Salmonella typhimurium. J Bacteriol 1978, 135(3):928–934.
26. Hantke K: Regulation of ferric iron transport in Escherichia coli K12: isolation of a constitutive mutant. Mol Gen Genet 1981, 182(2):288–292. 27. Achenbach LA, Yang W: The fur gene from Klebsiella pneumoniae:
characterization, genomic organization and phylogenetic analysis. Gene 1997, 185(2):201–207.
28. Griggs DW, Konisky J: Mechanism for iron-regulated transcription of the Escherichia coli cir gene: metal-dependent binding of fur protein to the promoters. J Bacteriol 1989, 171(2):1048–1054.
29. Hassett DJ, Sokol PA, Howell ML, Ma JF, Schweizer HT, Ochsner U, Vasil ML: Ferric uptake regulator (Fur) mutants of Pseudomonas aeruginosa demonstrate defective siderophore-mediated iron uptake, altered aerobic growth, and decreased superoxide dismutase and catalase activities. J Bacteriol 1996, 178(14):3996–4003.
30. Ochsner UA, Vasil ML: Gene repression by the ferric uptake regulator in Pseudomonas aeruginosa: cycle selection of iron-regulated genes. Proc Natl Acad Sci USA 1996, 93(9):4409–4414.
31. Bijlsma JJ, Waidner B, Vliet AH, Hughes NJ, Hag S, Bereswill S, Kelly DJ, Vandenbroucke-Grauls CM, Kist M, Kusters JG: The Helicobacter pylori homologue of the ferric uptake regulator is involved in acid resistance. Infect Immun 2002, 70(2):606–611.
32. van Vliet AH, Stoof J, Poppelaars SW, Bereswill S, Homuth G, Kist M, Kuipers EJ, Kusters JG: Differential regulation of amidase- and formamidase-mediated ammonia production by the Helicobacter pylori fur repressor. J Biol Chem 2003, 278(11):9052–9057.
33. Wu CC, Lin CT, Cheng WY, Huang CJ, Wang ZC, Peng HL: Fur-dependent MrkHI regulation of type 3 fimbriae in Klebsiella pneumoniae CG43. Microbiology 2012, 158(Pt 4):1045–1056.
34. Hantke K: Iron and metal regulation in bacteria. Curr Opin Microbiol 2001, 4(2):172–177.
35. Masse E, Vanderpool CK, Gottesman S: Effect of RyhB small RNA on global iron use in Escherichia coli. J Bacteriol 2005, 187(20):6962–6971.
36. Andrews SC, Harrison PM, Guest JR: Cloning, sequencing, and mapping of the bacterioferritin gene (bfr) of Escherichia coli K-12. J Bacteriol1989, 171 (7):3940–3947.
37. Gruer MJ, Guest JR: Two genetically-distinct and differentially-regulated aconitases (AcnA and AcnB) in Escherichia coli. Microbiology 1994, 140(Pt 10):2531–2541.
38. Niederhoffer EC, Naranjo CM, Bradley KL, Fee JA: Control of Escherichia coli superoxide dismutase (sodA and sodB) genes by the ferric uptake regulation (fur) locus. J Bacteriol 1990, 172(4):1930–1938.
39. Masse E, Gottesman S: A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc Natl Acad Sci USA 2002, 99(7):4620–4625.
40. Masse E, Escorcia FE, Gottesman S: Coupled degradation of a small regulatory RNA and its mRNA targets in Escherichia coli. Genes Dev 2003, 17(19):2374–2383.
41. Dubrac S, Touati D: Fur positive regulation of iron superoxide dismutase in Escherichia coli: functional analysis of the sodB promoter. J Bacteriol 2000, 182(13):3802–3808.
42. Davis BM, Quinones M, Pratt J, Ding Y, Waldor MK: Characterization of the small untranslated RNA RyhB and its regulon in Vibrio cholerae. J Bacteriol 2005, 187(12):4005–4014.
43. Argaman L, Elgrably-Weiss M, Hershko T, Vogel J, Altuvia S: RelA protein stimulates the activity of RyhB small RNA by acting on RNA-binding protein Hfq. Proc Natl Acad Sci USA 2012, 109(12):4621–4626.
44. Mey AR, Craig SA, Payne SM: Characterization of Vibrio cholerae RyhB: the RyhB regulon and role of ryhB in biofilm formation. Infect Immun 2005, 73 (9):5706–5719.
45. Murphy ER, Payne SM: RyhB, an iron-responsive small RNA molecule, regulates Shigella dysenteriae virulence. Infect Immun 2007,
75(7):3470–3477.
46. Blumenkrantz N, Asboe-Hansen G: New method for quantitative determination of uronic acids. Anal Biochem 1973, 54(2):484–489. 47. Kuhn J, Briegel A, Morschel E, Kahnt J, Leser K, Wick S, Jensen GJ,
Thanbichler M: Bactofilins, a ubiquitous class of cytoskeletal proteins mediating polar localization of a cell wall synthase in Caulobacter crescentus. EMBO J 2010, 29(2):327–339.
48. Desnoyers G, Morissette A, Prevost K, Masse E: Small RNA-induced differential degradation of the polycistronic mRNA iscRSUA. EMBO J 2009, 28(11):1551–1561.
49. Masse E, Salvail H, Desnoyers G, Arguin M: Small RNAs controlling iron metabolism. Curr Opin Microbiol 2007, 10(2):140–145.
50. Jacques JF, Jang S, Prevost K, Desnoyers G, Desmarais M, Imlay J, Masse E: RyhB small RNA modulates the free intracellular iron pool and is essential for normal growth during iron limitation in Escherichia coli. Mol Microbiol 2006, 62(4):1181–1190.
51. Salvail H, Lanthier-Bourbonnais P, Sobota JM, Caza M, Benjamin JA, Mendieta ME, Lepine F, Dozois CM, Imlay J, Masse E: A small RNA promotes siderophore production through transcriptional and metabolic remodeling. Proc Natl Acad Sci USA 2010, 107(34):15223–15228.
52. Frohlich KS, Vogel J: Activation of gene expression by small RNA. Curr Opin Microbiol 2009, 12(6):674–682.
53. Prevost K, Salvail H, Desnoyers G, Jacques JF, Phaneuf E, Masse E: The small RNA RyhB activates the translation of shiA mRNA encoding a permease
of shikimate, a compound involved in siderophore synthesis. Mol Microbiol 2007, 64(5):1260–1273.
54. Rehmsmeier M, Steffen P, Hochsmann M, Giegerich R: Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10(10):1507–1517. 55. Salvail H, Masse E: Regulating iron storage and metabolism with RNA: an
overview of posttranscriptional controls of intracellular iron homeostasis. Wiley Interdiscip Rev RNA 2012, 3(1):26–36.
56. Lai YC, Peng HL, Chang HY: Identification of genes induced in vivo during Klebsiella pneumoniae CG43 infection. Infect Immun 2001, 69(11):7140–7145. 57. Lai YC, Peng HL, Chang HY: RmpA2, an activator of capsule biosynthesis in Klebsiella pneumoniae CG43, regulates K2 cps gene expression at the transcriptional level. J Bacteriol 2003, 185(3):788–800.
58. Hanahan D: Studies on transformation of Escherichia coli with plasmids. J Mol Biol 1983, 166(4):557–580.
59. Skorupski K, Taylor RK: Positive selection vectors for allelic exchange. Gene 1996, 169(1):47–52.
60. Hantke K: Selection procedure for deregulated iron transport mutants (fur) in Escherichia coli K 12: fur not only affects iron metabolism. Mol Gen Genet 1987, 210(1):135–139.
61. Keen NT, Tamaki S, Kobayashi D, Trollinger D: Improved broad-host-range plasmids for DNA cloning in gram-negative bacteria. Gene 1988, 70 (1):191–197.
62. Tabor S, Richardson CC: A bacteriophage T7 RNA polymerase/promoter system for controlled exclusive expression of specific genes. Proc Natl Acad Sci USA 1985, 82(4):1074–1078.
63. Lin CT, Huang TY, Liang WC, Peng HL: Homologous response regulators KvgA, KvhA and KvhR regulate the synthesis of capsular polysaccharide in Klebsiella pneumoniae CG43 in a coordinated manner. J Biochem 2006, 140(3):429–438.
64. Stojiljkovic I, Baumler AJ, Hantke K: Fur regulon in gram-negative bacteria. Identification and characterization of new iron-regulated Escherichia coli genes by a fur titration assay. J Mol Biol 1994, 236(2):531–545.
65. Domenico P, Schwartz S, Cunha BA: Reduction of capsular polysaccharide production in Klebsiella pneumoniae by sodium salicylate. Infect Immun 1989, 57(12):3778–3782.
66. Schwyn B, Neilands JB: Universal chemical assay for the detection and determination of siderophores. Anal Biochem 1987, 160(1):47–56.
doi:10.1186/1471-2180-12-148
Cite this article as: Huang et al.: Role of the small RNA RyhB in the Fur regulon in mediating the capsular polysaccharide biosynthesis and iron acquisition systems in Klebsiella pneumoniae. BMC Microbiology 2012 12:148.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit