緒 言
植 物 菌 質 體 ( p h y t o p l a s m a ), 原 名 為 似 菌 質 體 (mycoplasm-like organism, MLO),是一種重要的植物病原 細菌。目前已知全球有超過兩百種的植物病害是由植物菌
質體所引起( 4 ),其中包括了梨、花生、甘藷及泡桐等重要
經濟作物的病害,在農業上造成十分嚴重的損失。花生簇 葉病 (peanut witches' broom, PNWB) 在台灣首先於澎湖及 苗 栗 兩 地 被 發 現 , 其 病 原 經 由 4 ' , 6 ' d i a m i d i n o 2 -phenylindole hydrochloride (DAPI)螢光染色、電子顯微鏡 之檢定,及利用菟絲子 ( d o d d e r ) 與南斑浮塵子 (O ro s i u s o r i e n t a l i s M a t s u m u r a )傳播等方式鑑定後,確定是由植物
菌質體所引起之病害( 3 2 )。
原核生物具有三種 D N A 聚合瓷,分別為 D N A 聚合瓷 I (DNA polymerase I)、D N A 聚合瓷 II (DNA polymerase I I )、及 D N A 聚合瓷 III (DNA polymerase III)。D N A 聚合 瓷 I 為具有 5' to 3' DNA 聚合功能、3' to 5' 核酸外切瓷功能 以及 5' to 3' 核酸外切瓷功能( 3 1 )。D N A 聚合瓷 I I 與 D N A 聚 合瓷 I I I 皆具有 5' to 3' DNA 聚合功能及 3' to 5' 核酸外切瓷 功能,但 D N A 聚合瓷 I I 之活性較 D N A 聚合瓷 I I I 低,且 其突變株仍可存活。一般認為 D N A 聚合瓷 I I I 為原核生物 最重要之 D N A 複製瓷 ( r e p l i c a s e ),與生物體維持及繁衍 生 命 息 息 相 關 ( 1 3 )。 D N A 聚 合 瓷 I I I 之 完 全 酵 (holoenzyme) 共由十個次單位 ( s u b u n i t ) 所組成,其中α 單位與 D N A 之合成具直接相關性,其係由 p o l C 基因轉 而成( 2 0 )。本實驗室曾於 1 9 9 6 年嘗試選殖花生簇葉病菌 體之 p o l C 基因,期間獲得一選殖株 P D P B 5,其嵌入片 3 '端之核酸序列與 M. genitalium 及 M. pulmonis 之 p o l C 因的 3 ' 端相似,其 5 ' 端之互補股序列與 M. genitalium d n a N基因相似( 1 ),然而其完整之 ORF (open reading fram 及 p o l C 基因之功能區塊均未在該選殖片段中找到,故 時無法有完整之結論,因此,本實驗即在於利用不同之 殖策略再次從事花生簇葉病菌質體之 p o l C 基因之選殖 企圖能選殖出該基因全長,並與前述之實驗結果加以 對,以期能對其核酸序列及特性有更完整之探討。
材料與方法
試驗植物之繁殖與植物全 DNA (total DNA) 之純化 本實驗以日日春 (Catharanthus ro s e u s, periwrinkle) 為試驗植物來繁殖植物菌質體。利用菟絲子 ( C u s c u
花生簇葉病菌質體p o l C基因之選殖與分析
紀凱齡
1林長平
1 , 2 1 臺北市 國立台灣大學植物病理與微生物學系 2 聯絡作者,電子郵件c p l i n @ n t u . e d u . t w;傳真:+ 8 8 6 - 2 - 2 3 6 6 1 9 8 0 接受日期:中華民國 93 年 11月 29 日摘 要
紀凱齡、林長平. 2005. 花生簇葉病菌質體 p o l C 基因之選殖與分析. 植病會刊 14: 51-58. 本研究在於從事花生簇葉病菌質體 p o l C 基因之選殖,採用之選殖策略為先行收集其他革蘭氏陽 性細菌 p o l C 基因之核酸及胺基酸序列,進行比對分析,根據其高保守性區域設計引子對 F3-2/ R6-2, 再以感染花生簇葉病之日日春全 D N A 為模板進行 P C R 反應,藉此獲得一 729 bp 大小之 P C R 產物, 再將其選殖至 p C R崰I I - TO P O質體中,得到一重組質體 p F 3 - 2 - 1 5,針對 p F 3 - 2 - 1 5 之嵌入片段進行核酸 定序,並根據此序列設計專一性引子對 F4/ R6,經利用 PCR DIG 標識法針對 p F 3 - 2 - 1 5 之嵌入片段增 幅出 493 bp 之 D I G 標識片段,以作為 p o l C 基因篩選用核酸探針,從事花生簇葉病菌質體重組質體之 篩選,如此遂獲得一選殖株重組質體 p H 1 5 - 4 0。重組質體 p H 1 5 - 4 0 之嵌入片段長度為 5221 bp,其中 包含一個完整 O R F 及一個不完整的 O R F,分別稱之為 ORF1 (4479bp) 與 ORF2 (502 bp)。將由 O R F 1 核酸序列推衍所得之胺基酸序列進行比對分析後,發現該序列與其他細菌的 D N A 聚合瓷I I I 基因 p o l C 相似。此外,由 O R F 1 所推衍出之胺基酸序列在一些重要的胺基酸位點上的保守性也很高,而且在 O R F 1所推衍之胺基酸序列中,在第 4 0 0 個至第 5 6 2 個胺基酸之間可發現 exo domain,而在第 1 2 3 3 個 胺基酸至第 1 3 3 5 個胺基酸之間可發現 pol domain,因此推測 O R F 1 應為花生簇葉病菌質體的 p o l C 基 因。由 O R F 2 之核酸序列推衍所得之胺基酸序列進行比對分析後,發現該序列與其他細菌的 N A D H o x i d a s e基因 n o x 相似。以罹病日日春全 R N A 進行反轉錄聚合瓷連鎖反應的結果顯示 p o l C 基因有轉錄 情形發生,且選殖株 p H 1 5 - 4 0 的嵌入片段包含 p o l C 基因全長。 關鍵詞:D N A聚合瓷、花生簇葉病、p o l Ca u s t r a l i s, dodder)作為媒介,將花生病株上之植物菌質體 由花生傳入日日春健株上 ( 3 2 )。溫室中乃利用旁嫁接法 (side graft)繁殖。而植物全 D N A 抽取之方法主要乃參考 K o與 L i n 等人的方法( 1 9 ),稍作修改後進行。 花生簇葉病菌質體 p o l C 基因篩選用核酸探針 ( p ro b e ) 之製備 本研究之資料係由國家衛生研究院巨分子序列分析軟 體 (GCG, http://gcg.nhri. or g . t w / ) 基因資料庫及 N C B I (National Center for Biotechnology Information, h t t p : / / w w w. n c b i . n l m . n i h . g o v / )基因資料庫中收集細菌之 p o l C基因之核酸序列 (nucleic acid sequence) 與胺基酸序列 (amino acid sequence),以親緣關係較近之 M o l l i c u t e s 綱及 其他革蘭氏陽性細菌為主;另輔以革蘭氏陰性細菌如 Escherichia coli的資料。於基因庫中所蒐集之資料包括 Bacillus subtilis ( X 5 2 11 6 )( 1 5 ), Mycoplasma genitalium ( L 4 3 9 6 7 )( 1 2 ), M. pneumoniae ( U 0 0 0 8 9 )( 1 7 ), M. pulmonis ( U 0 6 8 3 3 )( 7 ), U reaplasma ure a l y t i c u m ( A E 0 0 2 1 3 5 )( 1 4 ), Staphylococcus aur e u s ( A B 0 5 3 3 5 3 )( 1 8 ), S t re p t o c o c c u s p y o g e n e s ( A F 2 8 0 7 6 1 )(8) 及 Escherichia coli ( D 8 3 5 3 6 )(30) 等 細菌之 p o l C 基因之核酸及胺基酸序列共八筆。將收集到 之核酸及胺基酸序列先行輸入 C L U S TA L(16) 軟體進行比 對,找出此蛋白質所特有保守性較高且相距有數百鹼基 ( b a s e )的六個區域,依該等區域對應之核酸序列,分別設 計出長約 20 bp 大 小 之 簡 併 式 寡 核 啟 酸 ( d e g e n e r a t e o l i g o n u c l e o t i d e s )引子 F3-2: 5'- GGKCARCAY C C W G G K G G D AT- 3 '及 R6-2: 5'-RGCRT G K G C T T T M G G R A A - 3 '。聚合 瓷連鎖反應 (polymerase chain reaction) 中,兩引子濃度各 為 0 . 5μM、並以 50 ng 前述製備之罹病或健康植物全 D N A進行 3 5 個循環,反應條件如下: 9 4℃ 3 0 秒,4 7℃ 3 0秒, 7 2℃ 1 分鐘。反應產物經 1 % 瓊脂凝膠水平電泳 (agarose gel horizontal electrophoresis) 分析後再利用 Q I A q u i c k崰 gel extraction kit (QIAGEN GmbH, Hilden, G e r m a n y )與 TOPO TA cloning崰kit (Invitrogen Corporation, San Diego, CA)進行 P C R 產物之純化與選殖。所獲得之選 殖株重組質體嵌入片段之核酸序列定序分析工作委由明欣 生物科技有限公司 (Mission Biotech, Taipei, Ta i w a n ) 進行。 核 酸 探 針 之 標 識 反 應 採 用 Roche Molecular B i o c h e m i c a l s公司 (Mannheim, Germany) 所開發之 P C R DIG probe synthesis kit進行,在 P C R 非放射標識反應中, 引子則依上述選殖株重組質體核酸定序所得之部分 p o l C 基因片段序列,重新於其上適當位置設計專一性引子 F 4 : 5 ' - A C T T T G AT TA A G T T T T T G ATG-3', R6: 5'- CCAAT C AT T TA C AT TATGCTC -3' (引子 F 4 與 R 6 相距 493 bp)。以 此引子對 F 4 / R 6 對罹病日日春全 D N A 以 P C R 進行標識反 應,以獲得篩選 p o l C 基因之核酸探針。 南 方 氏 轉 漬 (Southern blotting) 及 雜 配 反 應 ( h y b r i d i z a t i o n ) 取 5μg 感染花生簇葉病之植物全 D N A,分別以 1 0 單 位之 B a mHI, B g lII, C l aI, E c oRI, H i ndIII, P s tI, S m aI, X b aI , Xh oI 核酸內限制瓷 (Roche Molecular Biochemicals) 進行酵 解;另外,取 5μg 健康日日春全 D N A 及 500 ng 用於製備
核酸探針之重組質體 D N A 分別以 1 0 單位之 H i nd I I I 與 E c oR I核酸內限制瓷進行酵解以作為對照組,進行6 小時之 酵解後,再以 0 . 8 % 瓊脂凝膠進行水平式電泳,並將 D N A 轉漬至尼龍濾膜 ( H y b o n dT M-N+, 0.45μm nylon transfer
membrane; Amersham Pharmacia Biotech, Buckinghamshire, E n g l a n d ),隨後進行雜配及呈色反應(9, 19)。 花生簇葉病菌質體基因體D N A 重組質體之構築與篩選 由於上述以 H i nd I I I 內限制瓷酵解罹病日日春全 D N A 之南方氏雜配反應結果顯示其略大於 5.0 kb 之酵解產物與 核酸探針有雜配訊號產生(見結果),而該 5.0 kb 之酵解 產物為可能含有完整 p o l C 基因之最小 D N A 片段,因此初 步選擇以 H i nd I I I 內限制瓷酵解罹病日日春全 D N A 以進行 選殖工作。首先取 10 μg 之罹病日日春全 D N A 以 1 5 單位 之 H i nd I I I 內限制瓷於 3 7℃下作用 6 小時,以 6 5℃ 處理 1 0 分鐘以終止反應,再以 0.8 % 瓊脂凝膠進行水平式電泳, 切下 5 至 5.5 kb 大小之酵解產物,以 Q I A G E N 公司所開發 生產之 Q I A q u i c k崰gel extraction kit進行純化,此即為擬選 殖嵌入之 D N A,並選用 pBluescriptII SK (-) phagemid (Stratagene, LA Jolla, CA)為載體,將其同樣以內限制瓷 H i nd I I I 酵 解 後 , 再 以 alkaline phosphatase ( C I A P ) ( S t r a t a g e n e ) 處 理 進 行 5 ' 端 去 磷 酸 化 反 應 ( d e p h o s p h o r y l a t i o n ),隨後將兩者以莫耳數比 1:1 (167 ng 擬嵌入之片段:100 ng 載體) 之比例混合放入微量離心管 中,加入 1 單位之 T4 DNA ligase (Promega Corporation, M a d i s o n , W I ),於 1 6℃ 水浴中作用 1 6 小時以進行黏結反 應。取 D N A 黏結反應混合液加入 2 0 0μl 以氯化鈣 ( C a C l2) 法( 11) 處理的 E. coli TO P 1 0 F ' 勝任細胞,進行轉形作用以 獲得轉形株 ( t r a n s f o r m a n t )。 以菌落雜配反應 (colony hybridization) 之方法大量地 篩選轉形株,其步驟為將尼龍濾膜 ( H y b o n dT M- N + ;
Amersham Pharmacia Biotech)貼於已於 3 7℃ 培養 1 6 小時 之轉形株菌盤上,並以 18 G 針頭在濾膜上打洞做記號 後,在 3 7℃ 培養箱中靜置 5 分鐘使濾膜吸附菌體。隨後 將此吸附有菌體之濾膜取出置於以 denaturation solution (0.5 N NaOH, 1.5 M NaCl)浸濕之 3 M M 濾紙上 5 分鐘,將 接觸 菌體 的一 面朝 上, 而 後取 出尼 龍濾 膜置 於以 neutralization solution (0.5 M Tris-HCl, pH 7.5; 3 M NaCl) 浸濕之 3 M M 濾紙上 5 分鐘,再取出濾膜置於以 2x SSC 浸 濕之 3 M M 濾紙上 1 5 分鐘,之後置於 3 M M 濾紙上風乾, 利用紫外線聯結器施以 1 . 2 x 1 05μCi/ cm 強度的 U V 照射濾 膜 2 0 秒,使 D N A 與尼龍濾膜聯結,其後進行之雜配與呈 色反應步驟同南方氏雜配反應。篩選所得之選殖株重組質 體嵌入片段之核酸序列定序分析工作係委由明欣生物科技 有限公司 (Mission Biotech) 進行。定序結果並以 L a s e rg e n e 軟體 ( D N A S TAR Inc., Madison, WI) 及連結 N C B I 基因資料 庫對嵌入 D N A 之核酸序列進行分析,設定 AT G 為起始密 碼, T G A、TA G 及 TA A 為終止密碼 ( 2 1 ),找出可能之 O R F。 反轉錄聚合瓷連鎖反應 ( reverse transcription PCR, RT- P C R ) 取 5μg 罹病或健康植株之全 R N A,加入 1μl (10
units) RNase-free DNase I (Roche Molecular Biochemicals) ,以 n u c l e a s e - f r e e 無菌水調整體積至 2 0μl。以 3 7℃ 處理 3 0分鐘,再經 6 8℃ 處理 1 0 分鐘以去除 DNase I 活性及 R N A之二級結構,並立即置於冰上待用。本實驗之反轉 錄聚合瓷連鎖反應是採用 S u p e r S c r i p tT M One-Step RT- P C R with Platinum崰Ta q套組 (Invitrogen Corporation)。實驗時 取 1μl 上述反應液 ( 0 . 5μg RNA),再依序加入 1 2 . 5μl 的 2x reaction mix、引子 (FH15-3/ RH15 or FH15-4/ RH15) (FH153: 5' AAATAT TA G A A ATTTTCC 3', FH154: 5' -C -C AT TA ATAGAAGTTGTGG -3', RH15: 5'- TTTTA AT T -C T GAAATGTCAAA -3')(FH15-3、FH15-4 皆位於 polC 上游、 R H 1 5位於 p o l C 之近 5 ' 端處,其相對位置見圖一) 各 1μl (使反應濃度為 0 . 8μM) ,加入 0 . 5μlRT/ Platinum崰 Ta q, 再加入 n u c l e a s e - f r e e 無菌水使反應總體積為 2 5μl。反應流 程為 5 0℃ 3 0 分鐘,9 4℃ 2 分鐘;再以 9 4℃ 3 0 秒鐘,5 6℃ 3 0秒鐘,7 2℃ 3 0 秒鐘進行 3 5 個循環。另外,分別取健康 或感染花生簇葉病之日日春全 D N A 各 50 ng、健康或感染 花生簇葉病之日日春全 R N A 各 0 . 5μg,加入無菌水(使反 應總體積為 2 5μl )、上述引子 ( 2 0μM ) 各 1μl(使反應濃度 各為 0 . 8μM ),再加入 1 . 2 5 單位之 Ta q DNA polymerase (Promega Corporation),進行 P C R 反應,反應程式為 9 4℃ 3 0秒,5 6℃ 3 0 秒,7 2℃ 3 0 秒進行 3 5 個循環,以做為聚 合瓷連鎖反應之對照組。反應完成後之產物於 1 % 瓊脂凝 膠進行水平電泳分析。
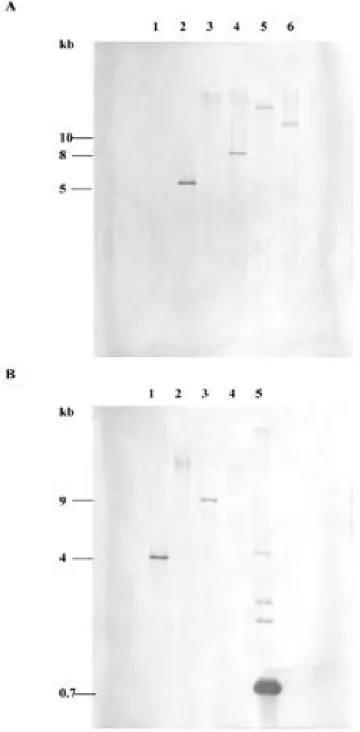
結 果
花生簇葉病菌質體 p o l C 基因篩選用核酸探針之製備 以簡併式寡核 啟酸引子對 F 3 - 2 / R 6 - 2 進行聚合瓷連鎖 反應之產物經純化與選殖後獲得一轉殖株重組質體 p F 3 2 -1 5,其嵌入片段大小為 729 bp。依重組質體 p F 3 - 2 - -1 5 嵌入 片段核酸定序所得之部分 p o l C 基因序列,重新於其上適 當位置設計專一性引子對 F 4 / R 6 對罹病日日春全 D N A 以 P C R進行 D I G 標識反應,即獲得 493 bp 之 p o l C 基因篩選 用核酸探針。 南方氏轉漬及雜配反應以核酸內限制瓷 B a mHI, B g lII, C l aI, E c oRI, H i nd I I P s tI, S m aI, X b aI及 X h oI 酵解感染花生簇葉病之日日春 D N A進行南方氏轉漬及雜配反應,並以 E c oR I 酵解之 組質體 p F 3 - 2 - 1 5 作為正反應對照組 (positive control)。 果顯示此探針對受花生簇葉病菌質體感染之日日春 D N A可產生雜配訊號,其中以 H i nd I I I 酵解者,可在約 於 5.0 kb 之 D N A 片段處產生雜配訊號;以 C l aI 酵解者 可在約 8.0 kb 之 D N A 片段處產生訊號;以 X b aI 酵解者 可在約 4.0 kb 之 D N A 片段處產生訊號;以 B g lI I 酵解者 可在約 9.0 kb 之 D N A 片段處產生訊號;而以其餘數種 酸內限制瓷酵解者皆在 10 Kb 以上之 D N A 片段處產生 配訊號,而正反應對照組也有專一性訊號產生;相對的 在健康日日春之全 D N A 則沒有任何雜配訊號 (圖二)。 花生簇葉病菌質體基因體 D N A 重組質體之篩選及 形株之特性分析 以菌落雜配反應進行篩選,得到一個有雜配訊息的 組質體轉形株 H 1 5 - 4 0。並以其製備大量的重組質體 p H 1 4 0之 D N A,並進行其嵌入片段全長之核酸定序分析。 酸序列之定序共進行兩次,確定無誤後再進行其核酸序 之後續分析。結果顯示重組質體 p H 1 5 - 4 0 包含 5221 bp 嵌入片段,其核酸組成為 1 4 . 2 % 的 G,3 7 . 3 % 的 A,3 8 . 2 的 T,1 0 . 3 % 的 C,A + T 值為 75.5 %,G + C 值為 2 4 . 5 % 符合植物菌質體基因體低 G + C 值的特性。接著 L a s e rg e n e軟體 ( D N A S TAR Inc.) 及連結至 N C B I 基因資 庫的 ORF Finder 針對此 5221 bp 之嵌入 D N A 核酸序列 行分析,找出可能之 O R F,結果在 p H 1 5 - 4 0 之嵌入片段 發現一個完整的 O R F 1 (nt 88-4566) 及一不完整之 ORF2 4 7 2 0 - 5 2 2 1 ) (圖一 )。本文中後續所述之序列已登錄 N C B I,登錄號碼為 GenBank AY 2 5 9 5 1 7,請參照。O R F 之核酸序列長度共有 4479 bp,可推衍出 1 4 9 2 個胺基酸 基因產物大小估計約為 174.9 kD 的蛋白質,轉譯起始密 為 AT G,終止密碼為 T G A。O R F 1 中 A + T 值為 7 5 . 4 2 % G + C值為 2 4 . 5 8 %,其核酸組成為 1 3 . 9 8 % 的 G,3 7 . 3 1 %
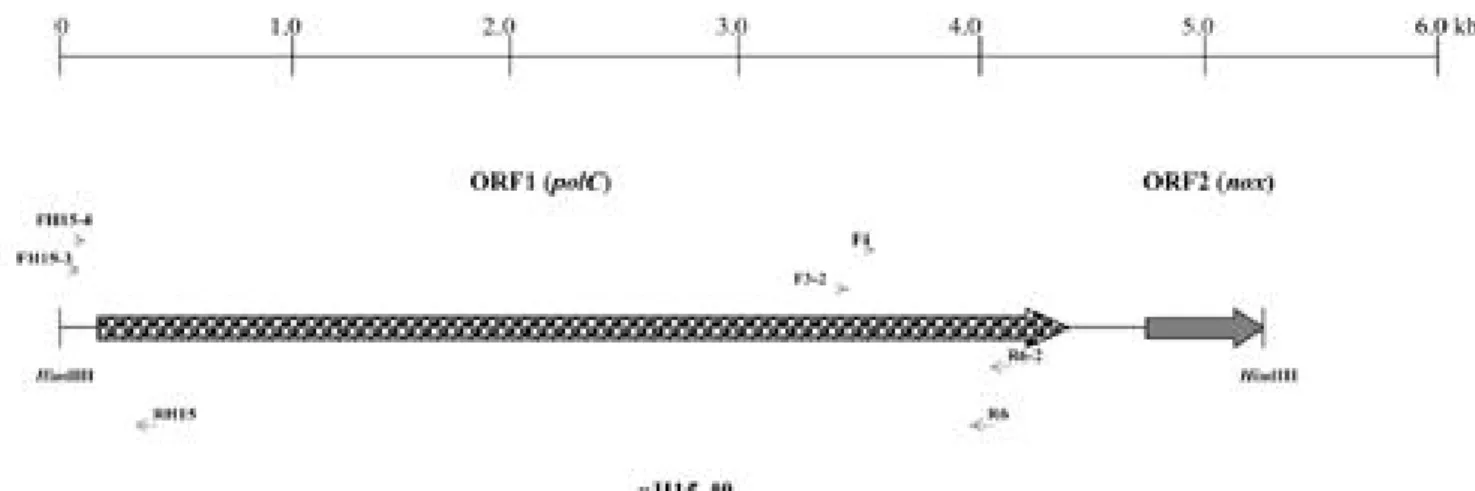
圖一、重組質體 pH15-40 所含之 ORF1 及 ORF2 示意圖及實驗中所用之引子相對位置。
Fig. 1. Diagram showing the relative positions of ORF1 and ORF2 in the insert DNA of recombinant plasmid pH15-40 and annealing sites of primers applied in this study. Large arrows refer to the direction of transcription, and small arrows refer to 5' to 3' direction of oligonucleotide primers.
A,3 8 . 11 % 的 T,1 0 . 6 0 % 的 C;其密碼利用性 ( c o d o n u s a g e )如表一所示,密碼利用率最高為 A A A ( 9 . 5 7 % ),其 次為 A U U ( 7 . 9 % ),密碼第三位核酸為 A 或 U 者佔所有密 碼之 8 8 . 0 7 %,密碼第一位核酸為 A 或 U 者佔所有密碼之 6 2 . 7 5 %。O R F 2 為一不完整的 O R F,其核酸序列長度為 502 bp,轉譯成胺基酸序列後共有 1 6 7 個胺基酸,轉譯起 始密碼為 AT G,未發現終止密碼。 O R F 2 中 A + T 值為 71.66 %,G + C 值為 2 8 . 3 4 %,其核酸組成為 1 8 . 3 6 % 的 G, 3 3 . 9 3 %的 A,3 7 . 7 3 % 的 T,9 . 9 8 % 的 C。以 O R F 1 之核酸 及胺基酸序列連結至 N C B I 資料庫,利用其中 B L A S T 軟 體進行比對,發現 O R F 1 與其他革蘭氏陽性細菌之 p o l C 基 因 相 似 , 在 胺 基 酸 序 列 上 有 5 0 % 以 上 之 相 似 度 ( s i m i l a r i t y ) (表二),例如與 B. subtilis 之 D N A 聚合瓷 I I I 之 相似度達 5 7 %,因此推測 O R F 1 可能為花生簇葉病原菌質 體之 p o l C 基因。根據前人之研究,革蘭氏陽性細菌之 D N A聚合瓷 I I I 包含兩個重要之功能區塊,如 B. subtilis 之 D N A聚合瓷 I I I,分別為位於第 4 1 5 至第 5 7 0 個胺基酸具 有 3' to 5' 核酸外切瓷功能的 exo domain;與位於第 11 7 5 至第 1 2 7 3 個胺基酸具有 5' to 3' 之 D N A 聚合功能的 p o l d o m a i n(6, 23),此革蘭氏陽性細菌的 D N A 聚合瓷 I I I 所特有 的兩個功能區塊在本研究中之花生簇葉病菌質體之 P o l C 蛋白上都有發現,其中 exo domain 位於花生簇葉病菌質體 P o l C蛋白的第 4 0 0 至第 5 6 2 個胺基酸之間;pol domain 位 於花生簇葉病菌質體 P o l C 蛋白的第 1 2 3 3 至第 1 3 3 5 個胺基 酸之間。此外,花生簇葉病菌質體的 P o l C 蛋白的胺基酸 序列在 D N A 聚合瓷 I I I 重要的胺基酸位點上之保守性很 高,如 exo domain 中的 A s p4 0 9、G l u4 11、G l y414 及 A s p4 9 9 (相對於 B. subtilis D N A 聚合 瓷 I I I 的 A s p4 2 5、G l u4 2 7、 G l y4 3 0及 A s p5 1 0)( 6 , 2 7 );pol domain 中的 S e r1 2 3 3、L e u1 2 3 5、 P h e1326 及 Va l1335 (相對於 B. subtilis D N A 聚合 瓷 I I I 的 S e r11 7 5、L e u11 7 7、P h e1 2 6 4及 Va l1 2 7 3)(5, 6, 15, 26),及分別位於 pol domain上游及下游的 Cys988 與 Ser1420(相對於 B. subtilis DNA聚合瓷 III 的 Cys940 與 Ser1360)(15)。在其起始密碼 ATG
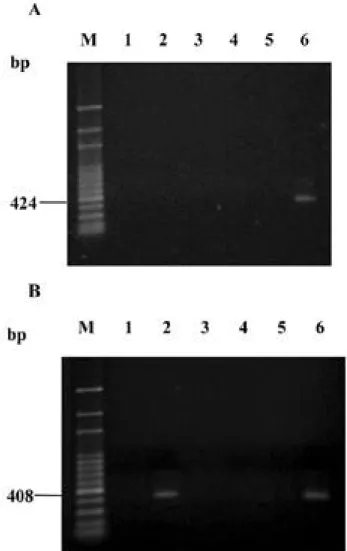
之上游約 9 至 2 7 個鹼基間,有兩個與植物菌質體(22) 及一
般原核生物的 16S rRNA 3' 端的予列互補,類似 S h i n e -D a l g a r n o序列 (Shine-Dalgarno sequence),亦即核糖體鍵結 位置 (ribosomal binding site, RBS) 以及啟動子 (promoter)(29)
。而以 O R F 2 之核酸及胺基酸序列連結至 N C B I 資料庫, 利用其中 B L A S T 軟體進行比對,發現與其他革蘭氏陽性 細菌之 NADH oxidase 基因 n o x 之 5 ' 端相似,在胺基酸序 列上有 6 1 % 以上之相似度 (表三),在其起始密碼 AT G 之 上游約 7 至 1 9 個鹼基間,亦發現兩個可能的核糖體鍵結 位置。 反轉錄聚合瓷連鎖反應 以花生簇葉病菌質體感染之日日春全 R N A 進行 P C R 反應,結果並沒有任何產物產生,顯示 D N a s e I 作用完 全,R N A 樣本內無 D N A 殘留。而以花生簇葉病菌質體感 染之日日春全 R N A 進行 RT- P C R 的結果,以 F H 1 5 - 3 / R H 1 5引子進行 RT- P R C 反應,無增幅出任何產物 (圖三 A );而以 FH15-4/ RH15 引子對進行 RT- P C R 反應,則可對 罹病日日春全 R N A 分別增幅出如預期大小 (408 bp) 之產 物,而以健康日日春全 R N A 為模版則沒有任何產物出現 (圖三B ),顯示所選殖到之花生簇葉病菌質體 p o l C 基因在 圖二、健康及受花生簇葉病菌質體感染之日日春全 D N A 以不同內限制 瓷酵解後以 polC 基因核酸探針進行南方氏 雜配分析。
Fig. 2. A, Southern blot analysis of total DNA (5μg per lane) prepared from healthy periwinkle digested with restriction enzyme H i ndIII (lane 1) and from diseased periwinkle infected with PNWB-phytoplasma digested with restriction enzymes H i ndIII (lane 2), B a mHI (lane 3), C l aI (lane 4), E c oRI (lane 5), and P s tI (lane 6). B, Southern blot analysis of total DNA from diseased periwinkles infected with PNWB-phytoplasma digested with restriction enzymes X b aI (lane 1), X h oI (lane 2), B g lII (lane 3), S m aI (lane 4) and DNA from recombinant plasmid pF3-2-15 digested with restriction enzyme E c oRI (lane 5). Hybridizations were conducted at 68 ℃ using the PCR DIG-labeled p o l C gene probes. Sizes (in kb) of the hybridization signals are indicated.
花生簇葉病菌質體有轉錄產物的產生。因此推測花生簇葉 病菌質體 p o l C 基因的轉錄起始位置應位於 F H 1 5 - 3 引子黏 合位置與 F H 1 5 - 4 引子黏合位置之間,故無法利用 F H 1 5 - 3 / R H 1 5引子對以 RT- P C R 增幅出任何產物。
討 論
由於 D N A 聚合瓷 I I I 為原核生物 D N A 複製過程中最 重要的酵素之一,因此一般認為解譯出此蛋白的 p o l C 基 因在演化的變異過程中累積相當的保守性,尤其是其中的 exo domain以及 pol domain。但鑑於革蘭氏陽性細菌與革 蘭氏陰性細菌之 D N A 聚合瓷 I I I 之基因序列、功能等特性 上仍有較大的差異存在(7, 8, 12, 14, 15, 17, 18, 20, 23, 30),因此本實 驗採用之基因選殖策略為利用相同基因在胺基酸序列上應 具有較高保守性之特性,收集多種革蘭氏陽性細菌 p o l 基因之核酸及胺基酸序列,搜尋其保守性區段與高保守 胺 基 酸 序 列 所 相 對 應 之 核 酸 密 碼 , 進 行 簡 併 ( d e g e n e r a t e ) P C R引子設計。引子對 F3-2/ R6-2,可增幅 具專一性大小為 729 bp 的單一 P C R 產物。將該 P C R 產 進行選殖後,獲得一選殖株重組質體 p F 3 - 2 - 1 5,其嵌入 段包含了花生簇葉病菌質體 p o l C 基因之一部分片段, 進而利用其序列設計專一性 P C R 引子,以獲得 p o l C 基 篩選用探針,此乃植物菌質體基因選殖之最關鍵步驟。 本 實 驗 乃 將 花 生 簇 葉 病 菌 質 體 D N A 構 築 pBluescriptII SK (-) phagemid中,以期得到具有花生簇 病菌質體 p o l C 基因之選殖株。 O t t 等人在研究中提到 s u b t i l i s的 D N A 聚合瓷 I I I 會抑制 E. coli 的生長( 2 4 ),因此 人於篩選重組質體轉形株時即未添加基因表現誘導 表一、花生簇葉病菌質體 p o l C 基因之密碼應用性Table 1. Codon usage in the PNWB-phytoplasma p o l C g e n e
Codon with 2n db a s e
U C A G
1s tb a s e A m i n o No. of A m i n o No. of A m i n o No. of A m i n o No. of 3r db a s e A c i d o c c u r r e n c e A c i d o c c u r r e n c e A c i d o c c u r r e n c e A c i d o c c u r r e n c e
U Phe (F) 1 0 4 Ser (S) 4 2 Tyr (Y) 6 8 Cys (C) 1 3 U
Phe (F) 4 Ser (S) 5 Tyr (Y) 4 Cys (C) 1 C
Leu (L) 1 0 4 Ser (S) 1 4 T E R1 0 T E R 1 A
Leu (L) 2 7 Ser (S) 5 T E R 0 Trp (W) 1 2 G
C Leu (L) 1 2 Pro (P) 3 6 His (H) 3 2 A rg (R) 1 3 U
Leu (L) 0 Pro (P) 1 His (H) 1 A rg (R) 5 C
Leu (L) 7 Pro (P) 11 Gln (Q) 5 6 A rg (R) 3 A
Leu (L) 1 Pro (P) 4 Gln (Q) 7 A rg (R) 2 G
A Ile (I) 11 8 Thr (T) 3 4 Asn (N) 9 6 Ser (S) 1 6 U
Ile (I) 11 Thr (T) 4 Asn (N) 5 Ser (S) 2 C
Ile (I) 3 4 Thr (T) 1 5 Lys (K) 1 4 3 A rg (R) 1 3 A Met (M) 2 2 Thr (T) 3 Lys (K) 1 2 A rg ( R ) 5 G G Val (V) 4 7 Ala (A) 3 6 Asp (D) 9 7 Gly (G) 3 1 U
Val (V) 0 Ala (A) 5 Asp (D) 5 Gly (G) 7 C
Val (V) 1 3 Ala (A) 5 Glu (E) 8 5 Gly (G) 1 6 A
Val (V) 4 Ala (A) 4 Glu (E) 7 Gly (G) 3 G
1
TER, termination codon
表二、花生簇葉病菌質體p o l C基因之胺基酸序列與多種細 菌之p o l C基因之胺基酸序列相似度及相同度之分析 Table 2. The percentage of similarities and identities of the amino acid sequence of PNWB-phytoplasma putative PolC protein compared with those of the PolC proteins of various o rg a n i s m s O rg a n i s m s S i m i l a r i t i e s I d e n t i t i e s Bacillus subtilis 5 7 % 4 0 % S t reptococcus pyogenes 5 5 % 3 8 % Staphylococcus aure u s 5 4 % 3 7 % Mycoplasma penetrans 5 4 % 3 5 % Mycoplasma genitalium 5 2 % 3 4 % U reaplasma ure a l y t i c u m 5 2 % 3 3 % Mycoplasma pneumoniae 5 1 % 3 4 % Mycoplasma pulmonis 5 0 % 3 4 % 表三、花生簇葉病菌質體 n o x基因之胺基酸序列與多種 菌之n o x基因之胺基酸序列相似度及相同度之分析 Table 3. The percentage of similarities and identities of amino acid sequence of PNWB-phytoplasma putative NAD oxidase compared with those of the NADH oxidases various org a n i s m s O rg a n i s m s S i m i l a r i t i e s I d e n t i t i e s E n t e rococcus faecalis 7 5 % 5 4 % Lactobacillus bre v i s 7 5 % 5 3 % Mycoplasma pulmonis 7 0 % 4 8 % Mycoplasma pneumoniae 6 3 % 3 7 % S t reptococcus pyogenes 6 1 % 3 9 % Mycoplasma genitalium 6 1 % 3 8 %
(inducer) IPTG,故乃直接利用菌落雜合反應篩選選殖 株,以避免重組質體之嵌入片段中 p o l C 基因表現而抑制 轉形株生長,徒增轉形株篩選之困難。 實驗中所篩選獲得之選殖株 H 1 5 - 4 0,其內之重組質 體 p H 1 5 - 4 0 包含兩個 O R F s,其 A + T 值分別高達 7 5 . 4 2 % 與 7 1 . 6 6 %,符合植物菌質體基因體之低 G + C 值特性,將其 核酸及胺基酸序列利用 N C B I 資料庫中 B L A S T 軟體進行 比對,發現 O R F 1 所推衍之胺基酸序列與其他革蘭氏陽性 細菌之 D N A 聚合瓷 I I I 相似,其大小 ( 1 4 9 2 個胺基酸) 亦與 其他革蘭氏陽性細菌之 D N A 聚合瓷 I I I 符合,而與 D N A 聚合瓷 I、I I 除了大小不同 (M. gentalium 之 D N A 聚合瓷 I 大小為 2 9 1 個胺基酸;Thermoplasma acidophilum 之 D N A 聚合瓷 I I 大小為 7 9 1 個胺基酸)(12, 25) 之外,其序列之比對 結果亦不相似(未展示結果);而不完整之 O R F 2 則與 n o x基 因之 5 ' 端相似。 在 O R F 1 核酸序列中共發現兩個起始密碼 AT G,分別 位於 O R F 1 的第 1 個至第 3 個鹼基 (nt 88-90, pH15-40) 與第 2 2 0個至第 2 2 2 個鹼基 (nt 307-309, pH15-40),若由前者開 始轉譯,則可轉譯成 1 4 9 2 胺基酸,略大於其他革蘭氏陽 性細菌之 D N A 聚合 瓷 I I I (含有 1 4 4 3 個至 1 4 5 1 個胺基 酸);但若由後者開始轉譯,則轉譯成 1 4 1 9 個胺基酸,略 小於其他革蘭氏陽性細菌之 D N A 聚合瓷 I I I。以正股引子 之黏合位置位於起始密碼 (第 1 個至第 3 個鹼基) 上游 3 2 個 鹼基之引子對 FH15-3/ RH15 無法經 RT- P C R 對感染花生簇 葉病病原植物菌質體之日日春全 R N A 增幅出任何產物, 而以正股引子之黏合位置位於起始密碼上游 1 5 個鹼基的 引子對 FH15-4/ RH15 進行 RT- P C R 反應,則可對罹病日日 春全 R N A 增幅出預期之產物,因此 O R F 1 的轉錄之起始 位置應位於 F H 1 5 - 3 引子與 F H 1 5 - 4 引子黏合位置之間,也 就是位於重組質體 p H 1 5 - 4 0 的第 3 9 個至第 7 5 個鹼基之 間,故推測 O R F 1 轉譯起始位置應位於 O R F 1 的第一個鹼 基;至於確切之轉錄作用起始鹼基位置則仍需進一步研究 證實。在轉譯起始密碼的上游亦發現一個可能的啟動子以 及兩個可能的核糖體鍵結位置,而核糖體鍵結位置一般位 於轉譯之起始密碼上游約 5 個至 9 個鹼基的位置,故推測 離轉譯起始密碼最接近的核糖體鍵結位置為最有可能的核 糖體鍵結位置。此外,由於前人研究發現植物菌質體密碼 應用上沒有以 U G A 為 t r y p t o p h a n 密碼的情形 (2, 3, 33), 故推測 O R F 1 之終止密碼應為位於 O R F 1 第 4 4 7 7 個鹼 基 (nt 4564, pH15-40) 之 T G A,而非位於其下游 5 2 個 鹼基 (nt 4618, pH15-40) 之 TA A。 革蘭氏陽性細菌的 D N A 聚合 瓷 I I I 所特有的 e x o domain與 pol domain 兩個功能區塊在吾人研究之花生簇葉 病菌質體之 P o l C 蛋白上都有發現,花生簇葉病菌質體的
PolC蛋白的胺基酸序列在 DNA 聚合瓷 III 重要的胺基酸位
點上之保守性很高,前人研究證實 exo domain 中的
A s p4 0 9、G l u4 11、G l y414 及 A s p4 9 9,單一個胺基酸發生改變 就會影響 3 ' to 5' 核酸酵解的功能,造成 D N A 複製時錯誤
率提高(6, 27);pol domain 中的 S e r1 2 3 3、L e u1 2 3 5、P h e1326 及
Val1335,其中一個發生改變就會影響 5' to 3' DNA 聚合的功
能,可能會導致 DNA 聚合瓷 III 對 arylhydrazino pyrimidine 藥劑產生抗性或 DNA 複製受到抑制(5, 6, 15, 26, 27),以及分別 位於 pol domain 上游及下游的 C y s988 與 S e r1 4 2 0,若其發生 改變會造成 DNA 聚合瓷 III 對高溫敏感(15)。由於吾人所選 殖到的花生簇葉病菌質體 p o l C 基因,其所轉譯之 P o l C 蛋 白的一級結構 (primary structure) 與其他革蘭氏陽性細菌的 D N A聚合瓷 I I I 的特性相同,因此進一步支持此基因即為 花生簇葉病菌質體之 p o l C 基因,且其產物與革蘭氏陽性 細菌的 D N A 聚合瓷 I I I 應具有相同的功能。 圖三、以反轉錄聚合瓷連鎖反應分析 p o l C基因之轉錄產 物。
Fig. 3. Transcript of the p o l C gene was verified by using RT-PCR. A, Primer FH15-3, located in the upstream region of the p o l C gene and primer RH15, in the 5' region of the p o l C g e n e were used. B, Primer FH15-4, located in the upstream region of the p o l C gene and primer RH15, in the 5' region of the p o l C gene were used. RT-PCR products were amplified by using RNA templates extracted from healthy periwinkle (lanes 1, A and B), and from periwinkle infected with PNWB-phytoplasma (lanes 2, A and B). PCR products were amplified by using RNA templates extracted from healthy periwinkle (lanes 3, A and B), and from periwinkle infected with PNWB-phytoplasma (lanes 4, A and B) and by using DNA templates extracted from healthy periwinkle (lanes 5, A and B), and from periwinkle infected with PNWB-phytoplasma (lanes 6, A and B). RT-PCR and PCR products were separated in a 1% agarose gel. M, 100 bp DNA ladder as molecular weight standards. Sizes of RT-PCR and PCR products are indicated on the left.
在一般生長期的原核生物細胞內,其 D N A 聚合瓷 I I I 之含量並不高,直到進入對數生長期,細胞中的 D N A 聚 合瓷 I I I 含量才迅速累積,此時的 D N A 聚合瓷 I I I 含量與 之前相比提高了五至八倍,每個細胞中約含有 5 0 個 D N A 聚合瓷 I I I 分子,直到將進入穩定生長期 (stationary phase) 時,D N A 聚合瓷 I I I 含量又迅速降低( 1 0 ),因此推測菌體內 p o l C基因之表現受到良好而嚴謹的調控。吾人所選殖的花 生簇葉病菌質體 p o l C 基因在其上游發現可能的核糖體鍵 結位置以及啟動子 TATA box,然而並未發現如 B. subtilis 之 p o l C 基因上游的終止子 ( t e r m i n a t o r )、反向重複序列 (inverted repeats)及 DnaA box 等與基因表現調控可能相關
的序列 ( 2 8 ),但是以上與基因表現調控之可能相關序列於 其他革蘭氏陽性細菌之 p o l C 基因上亦無發現,因此推測 花生簇葉病菌質體 p o l C 基因的調控可能是採取與 B . s u b t i l i s不同之方式,然而有關 p o l C 基因之調控機制之相 關研究目前仍然不多。 在本研究中,吾人亦從日日春罹病植株中抽取其全 R N A,做為研究花生簇葉病菌質體 p o l C 基因表現與否之 材料。利用反轉錄聚合 瓷連鎖反應,由黏合位置位於 p H 1 5 - 4 0嵌入片段 5 ' 端之引子對 FH15-3/ RH15 無法從日日 春罹病植株全 R N A 中增幅出產物,而利用 FH15-4/ RH15 引子對進行增幅,則均可增幅出預期大小之產物,顯示本 研究所選殖到之 p o l C 基因有轉錄產物的產生,且轉錄作 用起始位置應位於 F H 1 5 - 3 引子與 F H 1 5 - 4 引子黏合位置之 間,而選殖株 p H 1 5 - 4 0 之嵌入片段中的 O R F 1 即為 p o l C 基 因全長。
引用文獻
1 . 唐天仁 1996. 花生簇葉病病原菌質體核酸聚合瓷 I I I 基 因的選殖 . 國立台灣大學植物病蟲害學研究所碩士論 文. 2. 莊景光 2000. 花生簇葉病病原菌質體 g y r B 和 g y r A 基因 之選殖. 國立台灣大學植物病理學研究所碩士論文. 3. 黃俊霖 1996. 絲瓜簇葉病植物菌質體可能的 A B C轉運 系統基因之分離及特性分析. 國立台灣大學植物學研究 所碩士論文.4. Agrios, G. N. 1997. Plant diseases caused by mollicutes: phytoplasmas and spiroplasmas. Pages 457-470 in: Plant p a t h o l o g y, 4t hed. Academic Press, San Diego, CA. 5. Barnes, M. H., Hammond, R. A., Foster, K. A.,
M i t c h e n e r, J. A., and Brown, N. C. 1989. The cloned p o l C gene of Bacillus subtilis: characterization of the azp12 mutation and controlled in vitro synthesis of active DNA polymerase III. Gene 85: 177-186.
6. Barnes, M. H., Hammond, R. A., Kennedy, C. C., Mack, S. L., and Brown, N. C. 1992. Localization of the exonuclease and polymerase domains of Bacillus subtilis DNA polymerase III. Gene 111: 43-49.
7. Barnes, M. H., Tarantino, P. M., Jr., Spacciapoli, P. , Brown, N. C., Yu, H., and Dybvig, K. 1994. DNA polymerase III of Mycoplasma pulmonis: isolation and characterization of the enzyme and its structural gene,
p o l C. Mol. Microbiol. 13: 843-854.
8. Bruck, I. and O'Donnell, M. 2000. The DNA replicat machine of a Gram- positive organism. J. Biol. Che 275: 28971-28983.
9. Chen, M. F. and Lin, C. P. 1997. DNA probes and P primers for the detection of a phytoplasma associa with peanut witches' broom. Eur. J. Plant Pathol. 1 1 3 7 - 1 4 5 .
10. Ciarrocchi, G., Attolini, C., Cobianchi, F., Riva, S., a Falaschi, A. 1977. Modulation of deoxyribonucleic a polymerase III level during the life cycle of B a c i l l s u b t i l i s. J. Bacteriol. 131: 776-783.
11. Dagert, M. and Ehrlich, S. D. 1979. Prolonged incubat in calcium chloride improves the competence Escherichia coli cells. Gene 6: 23-28.
12. F r a s e r, C. M., Gocayne, J. D., White, O., Adama, M. Clayton, R. A., Fleischmann, R . D . , B u l t , C. Kerlavage, A. R., Sutton, G. G., Kelley, J. M., Fritchm J. L., Weidman, J. F., Small, K. V., Sandusky, M Fuhrmann, J. L., Nguyen, D. T., Utterback, T., Saudek, M., Phillips, C. A., Merrick, J. M., Tomb, J., Doughe B. A., Bott, K. F., Hu, P. C., Lucier, T. S., Peterson, S. Smith, H. O., and Ve n t e r, J. C. 1995. The minimal ge complement of Mycoplasma genitalium. Science 27 3 9 7 - 4 0 3 .
1 3 . G e f t e r, M. L., Hirota, Y., Kornberg, T., We c h s l e r, J. and Barnoux, C. 1971. Analysis of DNA polymerases and III in mutants of Escherichia coli Thermosensitive DNA Synthesis. Proc. Natl. Acad. Sci. USA 68: 315 3 1 5 3 .
14. Glass, J. I., Lefkowitz, E. J., Glas, J. S., Heiner, C. Chen, E. Y., and Cassell, G. H. 2000. The compl sequence of the mucosal pathogen U re a p l a s m u re a l y t i c u m. Nature 407: 757-762.
15. Hammond, R. A., Barnes, M. H., Mack, S. L., Mitchen J. A., and Brown, N. C. 1991. Bacillus subtilis D N polymerase III: complete sequence, overexpression, a characterization of the p o l C gene. Gene 98: 29-36. 16. Higgins, D. G. and Sharp, P. M. 1989. CLUSTA L :
package for performing multiple sequence alignment o m i c r o c o m p u t e r. Gene 73: 237-244.
17. Himmelreich, R., Hilbert, H., Plagens, H., Pirkl, E., Li, C., and Herrmann, R. 1996. Complete sequence analy of the genome of the bacterium Mycoplasma pneumoni Nucleic Acids Res. 24: 4420-4449.
18. Inoue, R., Kaito, C., Tanabe, M., Kamura, K., Akimit N., and Sekimizu, K. 2001. Genetic identification of t distinct DNA polymerases, DnaE and PolC, that essential for chromosomal DNA replication Staphylococcus aure u s. Mol. Genet. Genomics 66: 56 5 7 1 .
19. Ko, H. C. and Lin, C. P. 1994. Development a application of cloned DNA probe for a mycoplasmal o rganism associated with sweet potato witches'-broo
Phytopathology 84: 468-473.
20. K o r n b e rg, A. and Baker, T. A. 1992. Prokaryotic DNA polymerases other than E. coli Pol I. Pages 165-196 i n: DNA replication, 2n d ed. W. H. Freeman and Company, San Francisco, USA
21. Kozak, M. Y. 1983. Comparison of initiation of protein synthesis in prokaryotes, eucaryotes, and org a n e l l e s . Microbiol. Rev. 47: 1-45.
22. Lim, P. O. and Sears, B. B. 1989. 16S rRNA sequence indicates that plant- pathogenic mycoplasmalike o rganisms are evolutionarily distinct from animal mycoplasmas. J. Bacteriol. 171: 5901-5906.
23. L o w, R. L., Rashbaum, S. A., and Cozzarelli, N. R. 1976. Purification and characterization of DNA polymerase III from Bacillus subtilis . J. Biol. Chem. 251: 1311 - 1 3 2 5 . 24. Ott, R. W., Goodman, L. E., and Ganesan, A. T. 1987.
Expression of the Bacilllus subtilis p o l C gene in Escherichia coli. Mol. Gen. Genet. 207: 335-341.
25. Ruepp, A., Graml, W., Santos-Martinez, M. L., Koretke, K. K., Vo l k e r, C., Mewes, H. W., Frishman, D., Stocker, S., Lupas, A. N., and Baumeister, W. 2000. The genome sequence of the thermoacidophilic scavenger Thermoplasma acidophilum. Nature 407: 508-513. 26. Sanjanwala, B. and Ganesan, A. T. 1989. DNA
polymerase gene of Bacillus subtilis. Proc. Natl. Acad.
Sci. USA 86: 4421-4424.
27. Sanjanwala, B. and Ganesan, A. T. 1991. Genetic structure and domains of DNA polymerase III of B a c i l l u s s u b t i l i s. Mol. Gen. Genet. 226: 467-472.
28. Sanjanwala, B. and Ganesan, A. T. 1993. Leader region of the gene encoding DNA polymerse III of Bacillus subtilis. Mol. Gen. Genet. 236: 374- 378.
29. Shine, J. and Dalgarno, L. 1974. The 3'-terminal sequence of Escherichia coli 16S ribosomal RNA: complementarity to nonsense triplets and ribosome binding sites. Proc. Natl. Acad. Sci. USA 71: 1342-1346.
30. Tomasiewics, H. H. and McHenry, C. S. 1987. Sequence analysis of the Escherichia coli d n a E gene. J. Bacteriol. 169: 5735-5744.
31. We a v e r, R. F. 1999. DNA replication I: basic mechanism and enzymology. Pages. 633-611 i n: Molecular Biology, 1s ted. WCB/ McGraw-Hill, Boston, USA
32. Yang, I. L. 1988. Witches' broom diseases of sweet potato and peanut in Taiwan. Ph. D. Thesis. Hokkaido Univ. , J a p a n .
33. Yu, Y. L., Yeh, K. W., and Lin, C. P. 1998. An antigenic protein gene of a phytoplasma associated with sweet potato witches' broom. Microbiology 144: 1257-1262.
A B S T R A C T
Chi, K. L.1, and Lin, C. P.1 , 22005. Cloning and analysis of p o l C gene of phytoplasma associated with peanut witches' broom. Plant Pathol. Bull. 14:51-58. (1 .Department of Plant Pathology and Microbiology, National Taiwan University, Taipei, Taiwan 106, R. O. C.;2 Corresponding author, E-mail: [email protected]; Fax: + 8 8 6 - 2 - 2 3 6 6 1 9 8 0 )
To clone and analyze the p o l C gene of peanut witches' broom- phytoplasma (PNWB-phytoplasma), a pair of degenerate oligonucleotide PCR primer F3-2/ R6-2 were designed according to the conserved sequences of the p o l C gene of various Gram-positive bacteria. The total DNA from a periwinkle infected with PNWB-phytoplasma was prepared for PCR reaction to amplify a 729-bp PCR product. The PCR product was then cloned into the vector pCR崰I I - TOPO to obtain a recombinant plasmid pF3-2-15. A 493-bp DIG-labeled PCR product was amplified by using the primers F4/ R6 designed according to the sequence of the insert DNA of pF3-2-15 and was used as a nucleic acid probe for the screening of recombinant plasmids of PNWB-phytoplasma DNA. The recombinant plasmid pH15-40 thus obtained comprises an insert DNA of 5221 nucleotides and encompasses a complete ORF of 4479 nucleotides and an incomplete ORF of 502 nucleotides, referred to as ORF1 and ORF2, respectively. The alignment of the primary structures of ORF1 and the DNA polymerase IIIs of other Gram-positive bacteria reveals strong conservation of long segments of sequences and perfect conservation of amino acid residues in the exo domain (400t h- 5 6 2n damino acid) and the pol domain (1233r d - 1 3 3 5t hamino acid). The analyses of nucleic acid sequence and amino acid sequence suggest that ORF1 is the putative p o l C gene of PNWB-phytoplasma. Moreover, the sequence of ORF2 is homologous to the NADH oxidase gene, n o x, of other organisms. The results of RT-PCR reveal that the p o l C gene was transcribed when the total RNA from PNWB-phytoplasma infected periwinkle was applied as the template and the recombinant plasmid pH15-40 encompasses the entire p o l C gene of PNWB-p h y t o PNWB-p l a s m a .