嚴重再生不良性貧血病童之骨髓間質幹細胞研究:其增生與分化的潛能較差; Poor potential of proliferation and differentiation in bone marrow mesenchymal stem cells derived from children with severe aplastic anemia
46
0
0
全文
(2) 中文摘要 研究目的 特發性嚴重再生不良性貧血(idiopathic severe aplastic anemia; SAA)雖然罕見,但對病童卻有著生命的威脅;其發病的原因主要 是喪失骨髓造血的功能,但直至今日真正的致病機轉卻還未能瞭解 透徹。骨髓間質幹細胞(mesenchymal stem cells; MSCs)在骨髓造 血微環境中扮演舉足輕重的角色,可提供造血幹細胞(hematopoietic stem cells)生長分化成熟之所需。本研究的目的為探討嚴重再生不 良性貧血病童之骨髓間質幹細胞基本特質的缺陷。. 研究方法 分別自五位嚴重再生不良性貧血的病童及五位對照組兒童取 得骨髓抽出液,再將間質幹細胞分離並作體外培養。藉由觀察骨髓 間質幹細胞之細胞型態(morphology)、測定其免疫表面標誌的表 現(immunophenotyping) 、分析其增生能力(proliferative capacity) 和分化潛能(differentiation potential) ,比較兩組之間是否有差異。. 研究結果. i.
(3) 雖然嚴重再生不良性貧血病童及對照組兒童兩組之骨髓間質 幹細胞體外培養的細胞型態均呈現相似之紡錘狀,且免疫表面標誌 的表現均為 CD45、CD14、CD34 陰性和 CD105、CD73、CD44 陽 性;但增生能力的實驗發現,嚴重再生不良性貧血病童之骨髓間質 幹細胞的增生速度較慢且累積群體倍增殖(cumulative population doubling)較對照組小,分別為 1.83 ± 1.21 與 3.36 ± 0.87(p = 0.046) ,這意味著嚴重再生不良性貧血病童的骨髓間質幹細胞有著 較低的增生能力。經過骨分化誘導(osteogenic induction)之後, 嚴重再生不良性貧血病童之骨髓間質幹細胞表現較弱的鹼性磷酸 塩酶活性(alkaline phosphatase activity;1.46 ± 0.04 與 2.27 ± 0.32;p = 0.013) 、呈現較淡的 von Kossa 染色和較低的 core binding factor α1 基因表現(0.0015 ± 0.0005 與 0.0056 ± 0.0017;p = 0.013) 。經過脂肪分化誘導(adipogenic induction)之後,嚴重再生 不良性貧血病童之骨髓間質幹細胞呈現較淡染的 Oil red O 染色 (0.86 ± 0.22 與 1.73 ± 0.42;p = 0.013)和較低的 lipoproteinlipase 基因表現(0.0105 ± 0.0074 與 0.0527 ± 0.0254;p = 0.013) 。 以 real time-PCR 分析骨分化與脂肪分化之特定基因表現的結果和特 殊染色的發現一致,兩者都代表著嚴重再生不良性貧血病童之骨髓. ii.
(4) 間質幹細胞骨分化與脂肪分化的潛能較差。. 研究結論 本研究中,我們首次成功地顯示:嚴重再生不良性貧血病童之 骨髓間質幹細胞的增生與分化潛能較差。而骨髓間質幹細胞的改變 或許和骨髓造血能力的喪失有著密切的相關,這需要後續的研究進 一步來闡明骨髓間質幹細胞與嚴重再生不良性貧血兩者之間的相 關性。. iii.
(5) 英文摘要 Background Idiopathic severe aplastic anemia (SAA), characterized by failure of hematopoiesis, is rare and potentially life-threatening to children. However, the pathogenesis has not been completely understood, and insufficiency in the hematopoietic microenvironment can be an important factor. Mesenchymal stem cells (MSCs) play an important role in maintaining bone marrow microenvironment. Therefore, we aimed at the intrinsic defects of bone marrow MSCs derived from SAA children.. Materials and Methods Bone marrow MSCs were obtained from 5 SAA children and 5 controls. The morphology, immunophenotyping, proliferative capacity and differentiation potential of MSCs from SAA children were determined and compared with those of MSCs from controls.. Results MSCs of SAA and control group shared a similar spindle-shaped morphology in vitro. Both revealed a consistent immunophenotypic. iv.
(6) profile which was negative for CD45, CD14 and CD34, and positive for CD105, CD73, and CD44. However, SAA MSCs had slower expansion rate and smaller cumulative population doubling from passage 4 to 6 (1.83 ± 1.21 vs 3.36 ± 0.87; p = 0.046), indicating lower proliferative capacity. Besides, only 3 of 5 cultures of SAA group retained the ability to continue expansion till 80%-90% confluent cell layer beyond passage 6, suggesting earlier senescence of SAA MSCs. After osteogenic induction, SAA MSCs showed lower alkaline phosphatase activity (1.46 ± 0.04 vs 2.27 ± 0.32; p = 0.013), less intense von Kossa staining and lower gene expression of core binding factor α1 (0.0015 ± 0.0005 vs 0.0056 ± 0.0017; p = 0.013). Following adipogenic induction, SAA MSCs showed less intense Oil red O staining (0.86 ± 0.22 vs 1.73 ± 0.42; p = 0.013) and lower lipoproteinlipase expression (0.0105 ± 0.0074 vs 0.0527 ± 0.0254; p = 0.013). The results of real time-PCR analysis for the assessment of lineage-specific. genes. were. consistent. with. the. findings. of. histochemical stains, and both indicated that SAA MSCs had poor osteogenic and adipogenic potential.. Conclusions. v.
(7) In this study, we demonstrated that bone marrow MSCs from children with SAA had poor potential of proliferation and differentiation. These alterations in MSCs may contribute to the failure of hematopoiesis, and lead to the development of the disease. Further studies are needed to elucidate the relationship between MSCs and SAA.. vi.
(8) 誌謝辭 終於能在半工半讀的狀況下,兩年內順利地畢業! 能夠進入醫研所就讀並圓滿地完成實驗、修畢學分、完成碩士 論文,最最感謝的就是我的指導老師:巫康熙醫師;感謝他在百忙 中抽空與我討論、感謝他提供實驗的資源、感謝他一切有形及無形 的支持!還有實驗室友善的夥伴:欣貝、嬌玲姊、羅博士,幫我這 個實驗室新手順利上路,完成我自己以為是不可能任務的實驗。周 寬基教授、彭慶添副院長及韓鴻志教授給予我的指導,是我的碩士 論文完成的關鍵,更是進一步實驗很重要的方向。 能在如此愉悅的氣氛下完成學業,當然要感謝班上同學的齊心 合作,大家互相幫忙、提供諮詢、相互打氣,讓這兩年的生活雖然 忙碌卻是甜美的回憶。我的爸爸、媽媽、妹妹給我無窮的支持與鼓 勵,也讓我這段期間能無後顧之憂地奔波於學校、醫院,感謝他們! 他們是老天給我最寶貴的禮物! 感謝一路支持的所有長輩、朋友!感謝你們讓我的生命更美 好!. vii.
(9) 目錄 中文摘要 ------------------------------------------------------------------ i 英文摘要 ------------------------------------------------------------------ iv 誌謝辭 --------------------------------------------------------------------- vii 目錄 ------------------------------------------------------------------------ viii 圖目錄 --------------------------------------------------------------------- x. 論文正文 第一章 前言 1.1 研究背景 --------------------------------------------------- 1 1.2 研究目的 --------------------------------------------------- 3 第二章 研究方法 2.1 研究材料 --------------------------------------------------- 4 2.2 研究設計 --------------------------------------------------- 5 2.2.1 Cell culture 2.2.2 Immunophenotypic analysis 2.2.3 Proliferative capacity 2.2.4 Osteogenic and adipogenic potential 2.3 統計方法 --------------------------------------------------- 11. viii.
(10) 第三章 研究結果 3.1 Morphology and immunophenotypic profile ------------ 12 3.2 Proliferative capacity --------------------------------------- 13 3.3 Differentiation potential ------------------------------------ 14 第四章 討論 4.1 結果討論 --------------------------------------------------- 16 4.2 其他相關性討論 ------------------------------------------ 18 4.3 研究限制 --------------------------------------------------- 21 第五章 結論與建議 ---------------------------------------------------- 22 參考文獻 ------------------------------------------------------------------ 23 圖 --------------------------------------------------------------------------- 31. ix.
(11) 圖目錄 Figure 1.. Morphology and immunophenotyping. ---------------. 31. Figure 2.. Proliferative capacity. ------------------------------------ 32. Figure 3.. Differentiation potential. --------------------------------- 33. Figure 4.. Comparison of differentiation potential by histochemical stains. ------------------------------------------------------- 34. Figure 5.. Comparison of differentiation potential by lineage-specific gene expression. ------------------------------------------- 35. x.
(12) 第一章 前言 1.1 研究背景 Childhood acquired aplastic anemia, characterized by failure of hematopoiesis, is rare and potentially life-threatening with an annual incidence of 1-6 per million [1-3]. Severe aplastic anemia (SAA) is defined as profound bone marrow (BM) hypocellularity and marked peripheral blood pancytopenia. Despite of many putative etiologies, a specific cause cannot be identified in most children, and is termed “idiopathic SAA”. Significant advances have been made in the management. of. the. disease,. including. allogeneic. stem. cell. transplantation and immunosuppressive therapy [1, 3, 4]. However, the mechanism by which idiopathic SAA develops has not been completely elucidated. Although many studies demonstrated the association of immune-mediated pathogenesis, up to 30% of patients do not have detectable evidence for an underlying immune basis and not respond to immunosuppressive therapy [1]. Therefore, other mechanisms do exist. Mesenchymal stem cells (MSCs), first described by Friedenstein et al. [5], have the capacity of self-renewal and differentiation into mesenchyme-lineage cells. BM MSCs can interact with hematopoietic stem cells (HSCs) and secret cytokines and regulatory molecules [6-11].. 1.
(13) They play a crucial role in providing a specialized microenvironment for HSC survival and differentiation [6, 7, 9-11]. Therefore, MSC dysfunction may result in the impairment of hematopoiesis, and lead to the development of SAA.. 2.
(14) 1.2 研究目的 However, data focusing on the role of MSCs in the pathophysiology of SAA are very limited [12-14]. Till now, no information. about. immunophenotyping,. the. characteristics. proliferative. capacity. of. morphology,. and. differentiation. potential of SAA MSCs has been reported. In order to clarify the pathophysiology of SAA and to identify the characteristic changes of SAA MSCs, we compared these basic properties of BM MSCs derived from SAA children and controls.. 3.
(15) 第二章 研究方法 2.1 研究材料 BM cells were obtained from iliac crest aspirates. Idiopathic SAA was defined as pancytopenia and hypocellular BM after excluding any other underlying diseases. To diagnose SAA, BM cellularity of less than 25% and at least two of the following criteria must be fulfilled: absolute neutrophil count less than 0.5 × 109/L, platelet count less than 20 × 109/L, and reticulocyte less than 1% [1, 3, 15]. Control subjects were patients who received BM examination for diseases other than hematological diseases with pathological proof of normal BM. All patients were previously untreated and aged less than 18 years old. The institutional review board of Tungs’ Taichung MetroHarbor Hospital approved this protocol, and written informed consents were obtained from the parents or legal guardians of the patients.. 4.
(16) 2.2 研究設計 2.2.1 Cell culture Mononuclear cells were isolated from BM aspirates by Ficoll-Paque. density. centrifugation. (1.077. g/ml;. Amersham. Biosciences, Uppsala, Sweden) and then seeded in low-glucose Dulbecco’s modified Eagle medium (DMEM; Gibco, Gaithersburg, MD) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% Antibiotic-Antimycotic (Gibco). Cells were incubated at 37°C with 5% CO2 in a humidified atmosphere. After 48 hours, medium with suspension of nonadherent cells was discarded and fresh medium was added. Thereafter medium was replaced twice a week. When reaching 80%-90% confluence, cells were detached with 0.25% trypsin-EDTA (Gibco) and replated at a concentration of 8.5 × 103/cm2 in 10-cm dishes.. 5.
(17) 2.2.2 Immunophenotypic analysis Cultured MSCs (passage 4) were detached, washed, and resuspended in phosphate-buffered saline (Gibco). After fixing and blocking, the cells were immunolabeled with the following mouse anti-human antibodies: fluorescein isothiocyanate-conjugated CD45 (FITC-CD45;. BD. phycoerythrin-conjugated. Biosciences, CD14. San. (PE-CD14;. Jose, BD. CA),. Biosciences),. FITC-CD34 (BD Biosciences), FITC-CD105 (Serotec, Oxford, UK), PE-CD73 (BD Pharmigen, San Diego, CA) and FITC-CD44 (BD Pharmigen). The nonspecific mouse IgG (BD Biosciences) served as isotype control. Data were analyzed by flow cytometry (FACSCalibur; BD Biosciences) with CellQuest software.. 6.
(18) 2.2.3 Proliferative capacity Yield of cells at each passage was enumerated using Trypan blue (Gibco) to exclude dead cells. The population doubling (PD) of cultured MSCs was calculated according to the equation: PD = log2 (the number of viable cells at harvest / the number of seeded cells). The cumulative PD was the sum of PD from passage 4 to 6.. 7.
(19) 2.2.4 Osteogenic and adipogenic potential To evaluate differentiation potential, cultured MSCs (third passage, at 80%-90% confluence) were subjected to osteogenic and adipogenic differentiation in vitro. Cells were detached from culture dishes and replated in 60-mm dishes for further studies. To promote osteogenic differentiation, cells were incubated in DMEM supplemented with 10% FBS, 10 mM β-glycerophosphate (Sigma, St Louis, MO), 0.1 μM dexamethasone (Sigma) and 0.2 mM ascorbic acid (Sigma) for 3 weeks. On day 21, cultures were stained for alkaline phosphatase (ALP; Sigma) activity and mineralized deposits were detected by von Kossa stain (Cedarlane, Ontario, Canada). To quantify ALP activity, 2 ml of 0.05 N NaOH in ethanol was added to each dish after ALP activity stain and the extraction was measured by spectrophotometry (Ultrospec 1100 pro; Amersham Biosciences) at 550 nm. For induction of adipogenic differentiation, cells were grown in DMEM supplemented with 10% FBS, 1 μM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine (Sigma), 0.1 mM indomethacin (Sigma) and 10 μg/ml insulin (Novo Nordisk A/S, Bagsværd, Denmark) for 2 weeks. On day 14, adipogenic differentiation was demonstrated by intracellular accumulation of lipid droplets stainable with oil red O 8.
(20) (Sigma). The dye content was eluted by ethanol and quantified spectrophotometrically. Differentiation was verified further by real-time polymerase chain reaction (Q-PCR) for the assessment of lineage-specific genes as core binding factor α1 (Cbfa1) for osteocytes and lipoprotein lipase for adipocytes. MSCs cultured in osteogenic and adipogenic induction medium were harvested on day 21 and 14, respectively. Total RNA was extracted with Trizol reagent (Invitrogen, Carlsbad, CA) following the manufacturer’s instruction. Concentration of the RNA samples was estimated spectrophotometrically at OD 260/280, and cDNA was synthesized. using. MMLV. reverse. transcriptase. (Epicentre. Biotechnologies, Madison, WI) in the presence of oligo-dT primer (Promega, Madison, WI). The sequences of PCR primers were as follows: Cbfa1 [16], sense 5’-CATGGCGGGTAACGATGAA-3’ and antisense 5’-CGGCCCACAAATCTCAGATC-3’; lipoprotein lipase [17], sense 5’-ATGGAGAGCAAAGCCCTGCTC-3’ and antisense 5’-TACAGGGCGGCCACAAGTTTT-3’. The expression of β-actin (sense. 5’-TGTGGATCAGCAAGCAGGAGTA-3’. and. antisense. 5’-CAAGAAAGGGTGTAACGCAACTAAG-3’) was used as an internal control to normalized specific gene expression in each sample [16]. Q-PCR was performed using cDNA samples with SYBR Green 9.
(21) PCR master mix (Applied Biosystems, Foster City, CA) and carried out in the ABI 7300 Real-Time PCR system (Applied Biosystems).. 10.
(22) 2.3 統計方法 Data analysis was performed using SPSS 14.0 for Windows. Results are presented as mean ± standard deviation and 95% confidence interval. Kolmogorov-Smirnov Z test was used for comparison of the two groups. Statistical value of p < 0.05 was considered significant.. 11.
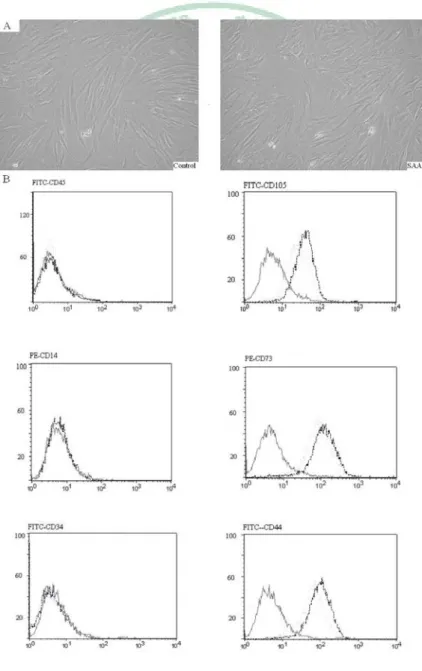
(23) 第三章 研究結果 3.1 Morphology and immunophenotypic profile A total of 10 children, 5 SAA patients and 5 controls, were enrolled in the study. The average age was 11.9 and 11.6 years old, respectively. In vitro, MSCs of SAA and control group shared a similar spindle-shaped morphology (Fig. 1A). Both revealed a consistent immunophenotypic profile which was negative for CD45, CD14 and CD34, and positive for CD105, CD73, and CD44 (Fig. 1B). No significant difference was noted in the expression of any single surface marker between the two groups.. 12.
(24) 3.2 Proliferative capacity To prevent hematopoietic cell contamination, which might be present in earlier passages, or the presence of senescent or differentiating MSCs in later passages, we used cells from passage 4 to 6 for the study of growth kinetics. SAA group had slower expansion rate than control group, shown as average PD of each passage (Fig. 2A). Two of five cultures of SAA group stopped proliferating at passage 5 and 6, respectively; whereas all cultures of control group continued to grow well. Lower proliferation potential of SAA MSCs was also demonstrated by smaller cumulative PD (1.83 ± 1.21 vs 3.36 ± 0.87; p = 0.046) (Fig. 2B).. 13.
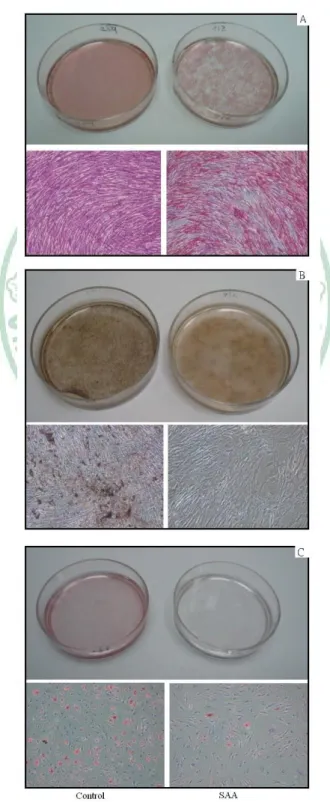
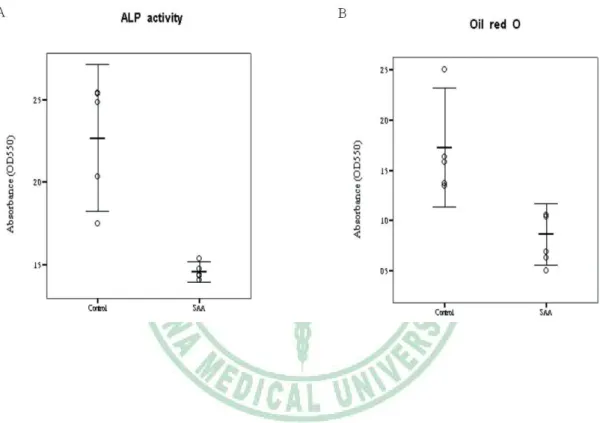
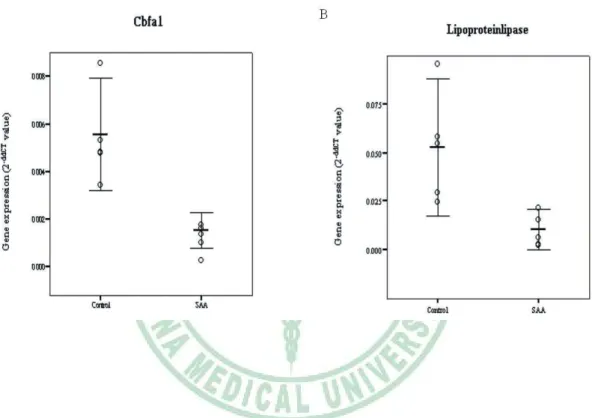
(25) 3.3 Differentiation potential When exposed to osteogenic induction medium, MSCs from SAA children had less robust osteogenic differentiation than MSCs from controls as shown by lower ALP activity (Fig. 3A). Greater extent of mineralization in control group was also demonstrated by more intense von Kossa stain (Fig. 3B). Under adipogenic condition, SAA MSCs gave rise to less lipid-containing cells. The intracytoplasmic vacuoles of neutral fat can be identified by Oil red O stain (Fig. 3C). The droplets of fat were more and larger within a single adipocyte in control group. Quantitation of dye content of ALP activity stain in the osteogenic cultures (1.46 ± 0.04 vs 2.27 ± 0.32; p = 0.013) and Oil red O stain in the adipogenic cultures (0.86 ± 0.22 vs 1.73 ± 0.42; p = 0.013) confirmed the histochemical observations (Fig. 4). Results of Q-PCR analysis presented in Figure 5 show the assessment of lineage-specific genes of osteogenesis and adipogenesis. After osteogenic induction, MSCs of SAA group expressed lower level of Cbfa1 than MSCs of control group (0.0015 ± 0.0005 vs 0.0056 ± 0.0017; p = 0.013). Following adipogenic induction, SAA MSCs showed lower lipoproteinlipase expression (0.0105 ± 0.0074 vs 0.0527 ± 0.0254; p = 0.013). The results of Q-PCR analysis were. 14.
(26) consistent with histochemical findings, and both strongly suggested that BM MSCs from SAA children had poor osteogenic and adipogenic potential.. 15.
(27) 第四章 討論 4.1 結果討論 MSCs can be characterized by a panel of surface markers, by their in vitro growth pattern and subsequent expansion and by their multilineage differentiation potential [10, 11, 18]. Many studies used these basic properties as indicators to identify MSCs from origins other than BM [8, 16, 17, 19-27]. We aimed at these properties of BM MSCs from SAA children and found that SAA MSCs had poor potential of proliferation and differentiation. The alterations in the characteristics of MSCs may contribute to failure of hematopoiesis and lead to the development of the disease. Although the morphology and surface marker expression of cultured MSCs did not change, SAA MSCs had slower expansion rate and smaller cumulative PD, indicating lower proliferation potential than normal MSCs. Besides, only 3 of 5 cultures of SAA group retained the ability to continue expansion till 80%-90% confluent cell layer beyond passage 6, suggesting earlier senescence of SAA MSCs. We can not answer the aging mechanisms here. However the heterogeneity in pathogenesis of the disease seems to explain the variations in MSC expansion potential.. 16.
(28) The significant decrease in osteogenic and adipogenic potential of SAA MSCs was demonstrated in our study. Even the 3 cultures which can proliferate beyond passage 6 showed lower expression of lineage-specific genes after differentiation under permissive conditions. The results of histochemical stains supported the gene expression study. These findings provided strong evidence that defects in BM MSCs of SAA children do exist.. 17.
(29) 4.2 其他相關性討論 The immune-mediated HSC destruction for the pathogenesis of idiopathic SAA has been widely accepted and many studies have devoted to the role of T cells in this disease [1, 4, 28-30]. Dubey et al. found elevated levels of interferon-gamma and tumor necrosis factor-alpha (TNF-α) in BM plasma of SAA patients [28], and these cytokines can induce apoptosis of CD34+ BM cells [31]. Hara et al. demonstrated excessive production of TNF-α by BM T cells and higher sensitivity of HSCs to TNF-α in patients with SAA [30]. However, up to 30% of patients have no immune-associated evidence and respond poorly to immunosuppressive therapy [1]. Pathogenesis of SAA remains to be Determined. Primary HSC deficiency, including decrease in number and dysfunction, has also been proposed to account for the development of SAA [32-35]. In vitro long-term BM culture provided evidence for primary HSC dysfunction in the regenerative capacity and in the response to various cytokine stimuli [32, 33]. Abnormal telomere shortening of HSCs was found in some patients with SAA [34]. However allogeneic HSC transplantation cannot cure all patients, suggesting that other mechanisms exist.. 18.
(30) Another important concept proposed in the context of SAA is related to the deficiency or dysfunction of BM microenvironment. MSCs. are. an. essential. component. of. the. hematopoietic. microenvironment which is appropriate for HSCs to survive, proliferate and differentiate [6, 7, 9-11]. MSCs provide an appropriate scaffold and a complex network of cytokines, adhesion molecules, and extracellular matrix proteins that are crucial for hematopoiesis [6-11]. Many studies have reported the promotive effect of MSCs for HSC expansion in vitro [7, 36-39]. In 2000, Koc et al. found rapid hematopoietic recovery after coinfusion of autologous MSCs at the time of HSC transplantation [40]. Accordingly, Lazarus et al. presented a multicenter trial of 46 patients receiving allogeneic HSCs and MSCs from HLA-identical siblings, and found prompt hematopoietic recovery in most patients, suggesting that the beneficial effect of MSCs on engraftment may relate to their supportive role in the hematopoiesis [41]. Only several studies have been conducted on the relationship between MSCs and SAA [12-14]. Bacigulupo et al. found that BM MSCs of SAA patients were deficient in their ability to suppress T cell proliferation and cytokine release, suggesting the lack of MSC immunoprotection in SAA BM [12]. Wu et al. investigated GATA gene expression of MSCs from chronic aplastic anemia patients, and. 19.
(31) proposed that aberrant expression of these genes in BM MSCs may influence. the. BM. microenvironment. and. lead. to. abnormal. hematopoietic regulation [13]. In current study, we observed poor potential of proliferation and differentiation in BM MSCs derived from SAA children. The above studies provide strong evidence for MSC defects in SAA BM. Several investigations have found that MSCs can not be acquired efficiently from umbilical cord blood of full-term infants, suggesting that MSCs are sparse or absent in cord blood [19-21]. Rubinstein et al. reported 562 recipients of cord blood HSC transplantation and found successful engraftment reduced significantly among SAA patients [42]. This can be explained, at least in part, by the insufficiency of BM hematopoietic microenvironment in SAA patients resulting from MSC defects and the few amount of MSCs provided by cord blood during transplantation. Therefore, cotransplant of MSCs and HSCs could be a potential strategy to treat SAA patients.. 20.
(32) 4.3 研究限制 Our study was of course limited by the small number of patients and the diseases of controls.. 21.
(33) 第五章 結論與建議 According to our results, BM MSCs derived from children with SAA had poor potential of proliferation and differentiation and these alterations may be important in the pathogenesis of the disease. Our findings. should. be. useful. in. further. understanding. of. the. pathophysiology of SAA and consequently lead to the development of novel treatment modalities. Further studies are needed to elucidate the relationship between MSCs and SAA.. 22.
(34) 參考文獻 1. Davies JK, Guinan EC (2007) An update on the management of severe idiopathic aplastic anaemia in children. Br J Haematol 136:549-564 2. Montane E, Ibanez L, Vidal X, Ballarin E, Puig R, Garcia N, Laporte J-R, Catalan Group for Study of Agranulocytosis and Aplastic Anemia (2008) Epidemiology of aplastic anemia: A prospective multicenter study. Haematologica 93:518-523 3. Kurre P, Johnson FL, Deeg HJ (2005) Diagnosis and treatment of children with aplastic anemia. Pediatr Blood Cancer 45:770-780 4. Young NS, Calado RT, Scheinberg P (2006) Current concepts in the pathophysiology. and. treatment. of. aplastic. anemia.. Blood. 108:2509-2519 5. Friedenstein AJ, Piatetzky-Shapiro II, Petrakova KV (1966) Osteogenesis in transplants of bone marrow cells. J Embryol Exp Morphol 16:381-390 6. Gordon MY (1988) Extracellular matrix of the marrow microenvironment. Br J Haematol 70:1-4 7. Verfaillie CM (1993) Soluble factor(s) produced by human bone marrow stroma increase cytokine-induced proliferation and maturation. 23.
(35) of primitive hematopoietic progenitors while preventing their terminal differentiation. Blood 82:2045-2053 8. Sorrentino A, Ferracin M, Castelli G, Biffoni M, Tomaselli G, Baiocchi M, Fatica A, Negrini M, Peschle C, Valtieri M (2008) Isolation and characterization of CD146+ multipotent mesenchymal stromal cells. Exp Hematol 36:1035-1046 9. Tocci A, Forte L (2003) Mesenchymal stem cell: Use and perspectives. Hematol J 4:92-96 10. Deans RJ, Moseley AB (2000) Mesenchymal stem cells: Biology and potential clinical uses. Exp Hematol 28:875-884 11. Lazennec G, Jorgensen C (2008) Concise review: Adult multipotent stromal cells and cancer: Risk or benefit? Stem Cells 26:1387-1394 12. Bacigalupo A, Valle M, Podesta M, Pitto A, Zocchi E, De Flora A, Pozzi S, Luchetti S, Frassoni F, Van Lint MT, Piaggio G (2005) T-cell suppression mediated by mesenchymal stem cells is deficient in patients with severe aplastic anemia. Exp Hematol 33:819-827 13. Wu X, Li Y, Zhu K, Wang Z, Chen S, Yang L (2007) GATA-1, -2 and -3 genes expression in bone marrow microenviroment with chronic aplastic anemia. Hematology 12:331-335 14. Scopes J, Ismail M, Marks KJ, Rutherford TR, Draycott GS, Pocock C, Gordon-Smith EC, Gibson FM (2001) Correction of stromal. 24.
(36) cell defect after bone marrow transplantation in aplastic anaemia. Br J Haematol 115:642-652 15. Camitta BM, Thomas ED, Nathan DG, Santos G, Gordon-Smith EC, Gale RP, Rappeport JM, Storb R (1976) Severe aplastic anemia: A prospective study of the effect of early marrow transplantation on acute mortality. Blood 48:63-70 16. Chang YJ, Shih DT, Tseng CP, Hsieh TB, Lee DC, Hwang SM (2006) Disparate mesenchyme-lineage tendencies in mesenchymal stem cells from human bone marrow and umbilical cord blood. Stem Cells 24:679-685 17. Lu LL, Liu YJ, Yang SG, Zhao QJ, Wang X, Gong W, Han ZB, Xu ZS, Lu YX, Liu D, Chen ZZ, Han ZC (2006) Isolation and characterization of human umbilical cord mesenchymal stem cells with hematopoiesis-supportive function and other potentials. Haematologica 91:1017-1026 18. Tocci A, Forte L (2003) Mesenchymal stem cell: Use and perspectives. Hematol J 4:92-96 19. Wexler SA, Donaldson C, Denning-Kendall P, Rice C, Bradley B, Hows JM (2003) Adult bone marrow is a rich source of human mesenchymal 'stem' cells but umbilical cord and mobilized adult blood are not. Br J Haematol 121:368-374. 25.
(37) 20. Yu M, Xiao Z, Shen L, Li L (2004) Mid-trimester fetal blood-derived. adherent. cells. share. characteristics. similar. to. mesenchymal stem cells but full-term umbilical cord blood does not. Br J Haematol 124:666-675 21. Secco M, Zucconi E, Vieira NM, Fogaca LL, Cerqueira A, Carvalho MD, Jazedje T, Okamoto OK, Muotri AR, Zatz M (2008) Multipotent stem cells from umbilical cord: Cord is richer than blood! Stem Cells 26:146-150 22. Wagner W, Wein F, Seckinger A, Frankhauser M, Wirkner U, Krause U, Blake J, Schwager C, Eckstein V, Ansorge W, Ho AD (2005) Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue, and umbilical cord blood. Exp Hematol 33:1402-1416 23. Wang H-S, Hung S-C, Peng S-T, Huang C-C, Wei H-M, Guo Y-J, Fu Y-S, Lai M-C, Chen C-C (2004) Mesenchymal stem cells in the Wharton's jelly of the human umbilical cord. Stem Cells 22:1330-1337 24. in 't Anker PS, Noort WA, Scherjon SA, Kleijburg-van der Keur C, Kruisselbrink AB, van Bezooijen RL, Beekhuizen W, Willemze R, Kanhai HH, Fibbe WE (2003) Mesenchymal stem cells in human second-trimester bone marrow, liver, lung, and spleen exhibit a similar immunophenotype but a heterogeneous multilineage differentiation. 26.
(38) potential. Haematologica 88:845-852 25. Campagnoli C, Roberts IA, Kumar S, Bennett PR, Bellantuono I, Fisk NM (2001) Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 98:2396-2402 26. Baksh D, Yao R, Tuan RS (2007) Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 25:1384-1392 27. Kim JW, Kim SY, Park SY, Kim YM, Kim JM, Lee MH, Ryu HM (2004) Mesenchymal progenitor cells in the human umbilical cord. Ann Hematol 83:733-738 28. Dubey S, Shukla P, Nityanand S (2005) Expression of interferon-gamma and tumor necrosis factor-alpha in bone marrow T cells and their levels in bone marrow plasma in patients with aplastic anemia. Ann Hematol 84:572-577 29. Hirano N, Butler MO, Von Bergwelt-Baildon MS, Maecker B, Schultze JL, O'Connor KC, Schur PH, Kojima S, Guinan EC, Nadler LM (2003) Autoantibodies frequently detected in patients with aplastic anemia. Blood 102:4567-4575 30. Hara T, Ando K, Tsurumi H, Moriwaki H (2004) Excessive. 27.
(39) production of tumor necrosis factor-alpha by bone marrow T lymphocytes is essential in causing bone marrow failure in patients with aplastic anemia. Eur J Haematol 73:10-16 31. Maciejewski J, Selleri C, Anderson S, Young NS (1995) Fas antigen expression on CD34+ human marrow cells is induced by interferon gamma and tumor necrosis factor alpha and potentiates cytokine-mediated. hematopoietic. suppression. in. vitro.. Blood. 85:3183-3190 32. Marsh JC, Chang J, Testa NG, Hows JM, Dexter TM (1990) The hematopoietic defect in aplastic anemia assessed by long-term marrow culture. Blood 76:1748-1757 33. Scopes J, Daly S, Atkinson R, Ball SE, Gordon-Smith EC, Gibson FM (1996) Aplastic anemia: Evidence for dysfunctional bone marrow progenitor. cells. and. the. corrective. effect. of. granulocyte. colony-stimulating factor in vitro. Blood 87:3179-3185 34. Ball SE, Gibson FM, Rizzo S, Tooze JA, Marsh JC, Gordon-Smith EC (1998) Progressive telomere shortening in aplastic anemia. Blood 91:3582-3592 35. Rizzo S, Scopes J, Elebute MO, Papadaki HA, Gordon-Smith EC, Gibson FM (2002) Stem cell defect in aplastic anemia: Reduced long term culture-initiating cells (LTC-IC) in CD34+ cells isolated from. 28.
(40) aplastic anemia patient bone marrow. Hematol J 3:230-236 36. Wang J-F, Wang L-J, Wu Y-F, Xiang Y, Xie C-G, Jia B-B, Harrington J, McNiece IK (2004) Mesenchymal stem/progenitor cells in human umbilical cord blood as support for ex vivo expansion of CD34(+) hematopoietic stem cells and for chondrogenic differentiation. Haematologica 89:837-844 37. Li N, Feugier P, Serrurrier B, Latger-Cannard V, Lesesve J-F, Stoltz J-F, Eljaafari A (2007) Human mesenchymal stem cells improve ex vivo expansion of adult human CD34+ peripheral blood progenitor cells and decrease their allostimulatory capacity. Exp Hematol 35:507-515 38. Zhang Y, Li C, Jiang X, Zhang S, Wu Y, Liu B, Tang P, Mao N (2004) Human placenta-derived mesenchymal progenitor cells support culture expansion of long-term culture-initiating cells from cord blood CD34+ cells. Exp Hematol 32:657-664 39. Van Overstraeten-Schlogel N, Beguin Y, Gothot A (2006) Role of stromal-derived factor-1 in the hematopoietic-supporting activity of human mesenchymal stem cells. Eur J Haematol 76:488-493 40. Koc ON, Gerson SL, Cooper BW, Dyhouse SM, Haynesworth SE, Caplan AI, Lazarus HM (2000) Rapid hematopoietic recovery after coinfusion of autologous-blood stem cells and culture-expanded. 29.
(41) marrow mesenchymal stem cells in advanced breast cancer patients receiving high-dose chemotherapy. J Clin Oncol 18:307-316 41. Lazarus HM, Koc ON, Devine SM, Curtin P, Maziarz RT, Holland HK, Shpall EJ, McCarthy P, Atkinson K, Cooper BW, Gerson SL, Laughlin MJ, Loberiza FR, Jr., Moseley AB, Bacigalupo A (2005) Cotransplantation. of. HLA-identical. sibling. culture-expanded. mesenchymal stem cells and hematopoietic stem cells in hematologic malignancy patients. Biol Blood Marrow Transplant 11:389-398 42. Rubinstein P, Carrier C, Scaradavou A, Kurtzberg J, Adamson J, Migliaccio AR, Berkowitz RL, Cabbad M, Dobrila NL, Taylor PE, Rosenfield RE, Stevens CE (1998) Outcomes among 562 recipients of placental-blood transplants from unrelated donors. N Engl J Med 339:1565-1577. 30.
(42) Figure 1. MSCs of controls and SAA children shared a similar spindle-shaped morphology (A, ×100 magnification) and a consistent immunophenotypic profile which was negative for CD45, CD14 and CD34, and positive for CD105, CD73, and CD44 (B). Black and dotted lines indicate cultured cells from controls and SAA children stained with antibodies, respectively. The respective isotype controls are shown as grey lines.. 31.
(43) Figure 2. Proliferative capacity. (A) Average PD of passage 4 to 6 of control and SAA group. (B) Comparison of cumulative PD, shown as mean and 95% confidence interval. Each circle represents a subject studied.. 32.
(44) Figure 3. Differentiation potential. Osteogenic differentiation was demonstrated by ALP activity (A) and von Kossa stain (B) after 3-week induction. Adipogenic differentiation was demonstrated by Oil red O stain (C) after 2-week induction. Magnification of micrographs: ×100.. 33.
(45) Figure 4. Comparison of osteogenic and adipogenic potential by quantification of ALP activity (A) and Oil red O (B) stain spectrophotometrically, respectively. Mean and 95% confidence interval are illustrated. Each circle represents a subject studied.. 34.
(46) Figure 5. Comparison of differentiation potential by lineage-specific gene expression, Cbfa1 for osteogenesis (A) and lipoproteinlipase for adipogenesis (B). Mean and 95% confidence interval are illustrated. Each circle represents a subject studied.. 35.
(47)
數據


+3
Outline
相關文件
11[] If a and b are fixed numbers, find parametric equations for the curve that consists of all possible positions of the point P in the figure, using the angle (J as the
• 是細胞不正常增生,且這些增生的細胞可
In the context of the Hong Kong school curriculum, STEM education is promoted through the Science, Technology and Mathematics Education Key Learning Areas (KLAs) in primary
唇音 b巴 p趴 m媽 f花 舌尖音 d打 t它 n拿 l啦 舌葉音 z渣 c茶 s沙 j也 舌根音 g家 k卡 ng牙. 圓唇音 gw瓜
唇音 b巴 p趴 m媽 f花 舌尖音 d打 t它 n拿 l啦.. 舌葉音 z渣 c茶 s沙 j也 舌根音 g家
They could also successfully form single-cell derived colonies (Fig.. Pyogenic granuloma-derived cells. A) After the initial seeding, many cells attached to the plates in an
This implies that f is an open mapping which maps interior points (of X) to interior points (of Y ) and f −1 is an open mapping from Y onto X... Is this convergence uniform
Figure 6 shows the relationship between the increment ∆y and the differential dy: ∆y represents the change in height of the curve y = f(x) and dy represents the change in height
