Chimeric Polypeptide of Thymidine Kinase and Thymidylate Kinase of Shrimp White Spot
Syndrome Virus: Thymidine Kinase Activity of the Recombinant Protein
Expressed in a Baculovirus/Insect Cell System
Huey-Fen Tzeng,*,§ Zee-Fen Chang,† Shao-En Peng,* Chung-Hsiung Wang,‡ Jung-Yaw Lin,† Guang-Hsiung Kou,*,1and Chu-Fang Lo*,1
*Department of Zoology, †Institute of Biochemistry and Molecular Biology, and ‡Department of Entomology, National Taiwan University, Taipei,
Taiwan, Republic of China, and §Department of Applied Chemistry, National Chi Nan University, Puli, Nantou, Taiwan, Republic of China Received October 29, 2001; returned to author for revision December 18, 2001; accepted February 4, 2002
The unique chimeric organization of the white spot syndrome virus (WSSV) tk-tmk gene encodes a protein which has significant homology to both cellular-type thymidine kinase (TK) and cellular-type thymidylate kinase (TMK), but the functional activity of this protein has not been demonstrated. Because TK is usually expressed only at very low levels in host cells, in this study, the coding region of WSSV tk-tmk was expressed in an insect/baculovirus expression system. The His-tagged recombinant WSSV TK-TMK was purified by affinity chromatography, and its enzyme activity was characterized by steady-state kinetics. The recombinant WSSV TK-TMK catalyzed the phosphorylation of thymidine to form thymidine monophosphate (TMP), but we found no evidence that it was able to catalyze the further phosphorylation of TMP to form thymidine diphosphate (or thymidine triphosphate). This TK activity is sensitive to feedback inhibition by thymidine triphosphate. In addition to thymidine, of the nine other substrates tested, including acyclovir, ganciclovir, and 5-(2-bromovinyl)-2 ⬘-deoxyuri-dine, only 2⬘-deoxyuridine and 5-bromo-2⬘-deoxyuridine could also serve as substrates. These data suggest that the enzymatic characteristics of the recombinant WSSV TK-TMK are similar to those of the eukaryotic cytosolic TKs. We also found that TK activity increased as infection advanced in the integument and gills of experimentally infected shrimp, suggesting its functional involvement during WSSV infection. © 2002 Elsevier Science (USA)
Key Words: Penaeus monodon; white spot syndrome virus; WSSV; thymidine kinase activity.
INTRODUCTION
White spot syndrome virus (WSSV), an economically significant virus that has impacted the shrimp industry worldwide (Inouye et al., 1994; Chou et al., 1995; Flegel, 1997; Peng et al., 2001) is a large DNA virus (Wang et al., 1995; Wongteerasupaya et al., 1995) with genome size ranging from 292 to 307 kb (van Hulten et al., 2001; Chen
et al., 2002). Its morphological characteristics (Wang et al., 1995, 2000; Wongteerasupaya et al., 1995), biological
properties (Lo et al., 1996, 1997; Lo and Kou, 1998), and molecular data (Tsai et al., 2000a,b; van Hulten et al., 2001; Liu et al., 2001; Chen et al., 2002) all strongly suggest that WSSV is a distinct virus (likely at the family level) that does not belong to any of the currently recog-nized virus families. Given the uniqueness of WSSV, a better understanding of its molecular biology is urgently needed to adequately interpret the nature of the virus and its molecular pathogenesis.
DNA is composed of four deoxyribonucleoside
triphosphate precursors, which are provided in cells by
de novo and salvage pathways. The key enzyme of the de novo pathway is ribonucleotide reductase (RR) which
reduces all four common ribonucleoside diphosphates to produce deoxyribonucleotides (Thelander and Reichard, 1979; Reichard, 1988; Stubbe, 1990). The key enzymes of the deoxyribonucleoside salvage pathway are the deoxyribonucleoside kinases, and in mammalian cells there are four principal salvage enzymes, namely thymidine kinase 1 (TK1; cytosolic thymidine kinase), thymidine kinase 2 (TK2; mitochondrial thymidine ki-nase), deoxycytidine kinase (dCK), and deoxyguanosine kinase (dGK). Although there are two mammalian thymi-dine kinases, many studies refer simply to “thymithymi-dine kinase,” or “cellular-type thymidine kinase,” by which is generally meant the cytosolic thymidine kinase, TK1. Dif-ferences between vertebrate and other eukaryotic TKs are minor, at least for the cytosolic TKs (reviewed in Gentry 1992; Arner and Eriksson, 1995). The WSSV ge-nome is known to contain genes encoding the key en-zyme of the de novo pathway (RR) (Tsai et al., 2000a) and one of the salvage pathway key enzymes, i.e., TK, which is present as a TK-TMK chimeric protein (Tsai et al., 2000b).
Several viruses other than WSSV carry a gene for TK
1To whom correspondence and reprint requests should be
ad-dressed at Department of Zoology, National Taiwan University, Taipei 106, Taiwan, R.O.C. Fax: 886-2-23638179. E-mail: gracelow@ccms. ntu.edu.tw or [email protected].
doi:10.1006/viro.2002.1480
0042-6822/02 $35.00
© 2002 Elsevier Science (USA) All rights reserved.
(Gentry, 1992). Herpesvirus TKs have many features in common with the mammalian TK2 and dCK. The herpes-virus TKs have a relatively broad substrate specificity such that they can also phosphorylate deoxycytidine (dC) (Kit et al., 1974) and thymidine monophosphate (TMP) (Chen and Prusoff, 1978), and they are relatively insen-sitive to feedback inhibition by thymidine triphosphate (TTP) (Cheng, 1976). By contrast, poxvirus TKs are very similar to mammalian TK1: they have a strict substrate specificity for thymidine and are sensitive to feedback inhibition by TTP (Kit and Dubbs, 1965; Buller and Palumbo, 1991).
WSSV TK-TMK is a protein with significant homology to both cellular-type thymidine kinase (TK) and cellular-type thymidylate kinase (TMK) (Tsai et al., 2000b). However, this protein has a unique chimeric organization and its functional activity has hitherto not been demonstrated. Since expression levels of the protein in infected host cells are too low to be detected by antibodies (data not shown), to obtain sufficient purified protein for charac-terization, we therefore inserted the coding region of WSSV tk-tmk into an Autographa californica multiple nucleopolyhedrovirus (AcMNPV)-based recombinant vi-rus for the production of WSSV TK-TMK in insect cells, and then the recombinant protein (rWSSV-TK-TMK) was purified by affinity chromatography for activity assays. To date, our preliminary studies have failed to detect any TMKactivity of rWSSV-TK-TMK(data not shown); in the present article we therefore focused on characterizing its TKactivity. We found that the recombinant WSSV TK-TMK protein could catalyze the phosphate transfer from ATP to thymidine. Similar to the eukaryotic cytosolic TKs, the TKof WSSV TK-TMKhas a strict substrate specificity for thymidine and is sensitive to feedback inhibition by TTP. Further it can also utilize most nucleoside triphosphates (CTP, GTP, UTP, dATP, dCTP, dGTP, and dUTP, but not TTP) as phosphate donors for the phosphorylation of thymidine without affecting its functional activity. Finally, we measured TKactivity in artificially infected shrimp cell lysate and showed that the TKactivity level increased as the infection phase advanced. This is the first time that a recombinant WSSV enzyme has been shown to be func-tionally active, and this functional activity means that TK’s potential as a target molecule of a WSSV antiviral che-motherapy can now be further investigated. As dis-cussed below, however, there may be difficulties with this approach because the substrate specificity of WSSV TK-TMK is very similar to that of its hosts.
RESULTS AND DISCUSSION
Expression and purification of recombinant WSSV TK-TMK
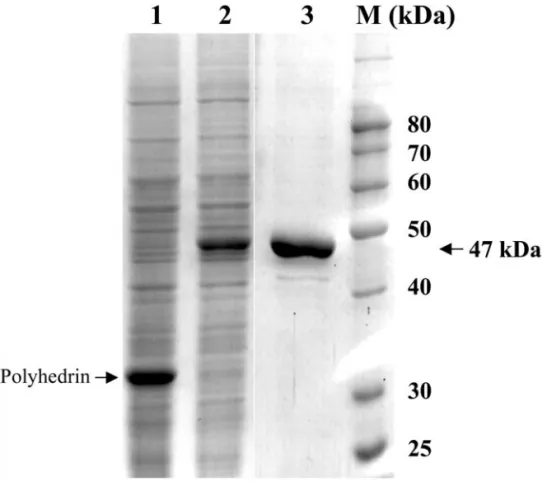
Recombinant WSSV TK-TMK (rWSSV-TK-TMK) was ex-pressed in Sf9 insect cells infected with the recombinant AcMNPV-WSSV-TK-TMK virus (Fig. 1). The recombinant
AcMNPV-WSSV-TK-TMK virus encoded 35 additional amino acids, including the hexahistidine-tag appended to the N-terminus of WSSV TK-TMK. This allowed purifi-cation of the recombinant protein on a nickel chelate column and resulted in a recombinant protein with a higher apparent molecular mass of about 47 kDa on SDS–PAGE (Fig. 1). This is about twice as large as the native cellular TK(20–25 kDa). Although some additional bands can be seen in Fig. 1, Lane 3, there was no contaminating band in this size range. Wild-type AcM-NPV does not contain a tk gene. Typically, 0.1 mg purified rWSSV-TK-TMK was recovered from cells grown in each 25-cm2 culture flask after passing through the Ni-NTA affinity column, and the enzyme was more than 90% pure as estimated from SDS–PAGE (Fig. 1, Lane 3). This puri-fied rWSSV-TK-TMK was used for the subsequent mea-surements of TKactivity.
Kinetic analysis for TK activity of rWSSV-TK-TMK A previous sequence analysis of WSSV TK-TMK has shown that WSSV TK-TMK contains the functional motifs of both TKand TMK(Tsai et al., 2000b). Therefore the purified rWSSV-TK-TMKwas examined for TKand TMK activity. No TMKactivity was detected (data not shown) by the enzyme-coupled TMKactivity assay (Chenal-Fran-cisque et al., 1999). Further investigation with more sen-sitive assay methods will be needed to determine whether or not the rWSSV-TK-TMK does in fact exhibit TMKactivity.
The kinetic parameters of TKactivity were determined by initial velocity (inmol/mg/min) measurements using tritium-labeled thymidine in the presence of ATP. The relationships between the substrate concentrations of thymidine and ATP and the initial velocity of the TK activity in rWSSV-TK-TMK are shown in double reciprocal plots (Fig. 2). The results indicated that rWSSV-TK-TMK was able to catalyze the phosphorylation of thymidine, and the kinetics of this TK-catalyzed reaction were hy-perbolic, as shown in Fig. 2. Steady-state kinetic param-eters of the TKactivity in rWSSV-TK-TMKwere also de-termined: the Michaelis constants (Km) of thymidine and
ATP were 4.1⫾ 0.5 and 76 ⫾ 6M, respectively, and Vmax was 1.5⫾ 0.1mol/mg/min.
In conclusion, rWSSV-TK-TMKhad TKactivity, and the thymidine affinity of this recombinant protein (Km4.1M)
was of the same order of magnitude as that reported for human TK1 (Km 0.5–3.0 M) (Eriksson et al., 1991;
Munch-Petersen et al., 1991; Berenstein et al., 2000). However, the velocity (Vmax 1.5 mol/mg/min) was 10 times lower than that of human TK1 (Vmax 16–17 mol/ mg/min) (Berenstein et al., 2000).
WSSV TK is a cellular-type (cytosolic) TK
Our previous sequence analysis also suggested that WSSV TKis a cellular-type TK(Tsai et al., 2000b). The
purified rWSSV-TK-TMK was therefore investigated for its ability to utilize phosphate donors, its nucleoside sub-strate specificity, and its end-product inhibition by TTP. Phosphate utilization for TK activity of rWSSV-TK-TMK
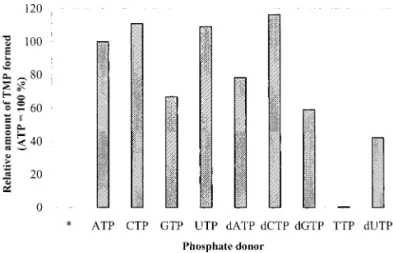
To test if rWSSV-TK-TMK can utilize triphosphates other than ATP as phosphate donors, ATP in the reaction buffer of the TKactivity assay was replaced by other NTPs and dNTPs. CTP, GTP, UTP, dATP, dCTP, dGTP, and dUTP could all substitute for ATP as the phosphate donor. The TKactivity of rWSSV-TK-TMKfell by about 20–60% when ATP was replaced by GTP, dATP, dGTP, or dUTP in the assay reaction, and TTP could not substitute for ATP (Fig. 3).
Other studies have shown that pure TK1 can also use most nucleoside triphosphates except TTP and CTP as phosphate donors (Eriksson et al., 1991; Munch-Petersen
et al., 1991). Thus in terms of flexibility in its use of
phosphate donors, WSSV TKis similar to TK1 and also to other eukaryotic cytosolic TKs. Unlike TK1, however, WSSV TKcan use CTP as the phosphate donor. In this, WSSV TKresembles the herpesvirus TKs and mitochon-drial TK(TK2), which are also able to use both ATP and CTP as phosphate donors (Kit et al., 1974; Arner and Eriksson 1995).
Nucleoside substrate specificity of rWSSV-TK-TMK for TK activity
For the nucleoside substrate specificity assay, the tested nucleosides and their derivatives were added into the TKassay solution as competitors to thymidine. If WSSV TKis able to catalyze the phosphorylation of any nucleoside substrates other than thymidine, then lower amounts of TMP should be formed in the presence of these competing nucleoside substrates. Nine other nucleosides and their derivatives were tested: 2 ⬘-de-oxyadenosine (dA), 2⬘-deoxycytidine (dC), 1--D -arabino-furanosylcytosine (AraC), 2⬘-deoxyguanosine (dG), 2⬘-de-oxyuridine (dU), 5-bromo-2⬘-deoxyuridine (BrdU), 5-(2-bromovinyl)-2⬘-deoxyuridine (BVdU), acyclovir (ACV), and ganciclovir (GCV). As shown in Fig. 4, of all the tested competing substrates only 2⬘-deoxyuridine (dU) and 5-bromo-2⬘-deoxyuridine (BrdU) influenced the formation of TMP, which suggests that only dU and BrdU can serve as nucleoside substrates for rWSSV-TK-TMK. The narrow nucleoside substrate specificity shown here by rWSSV-TK-TMK is also a characteristic of the cytosolic TKs.
Taken together, in terms of TKsubstrate specificity, WSSV-TK-TMK is similar to the cytosolic TKs, which are the most restricted of all the key salvage enzymes. Cy-tosolic TKs accept only thymine or uracil as a base with minor substitutions at the 5-position (e.g., a halogen or
FIG. 1. SDS–PAGE of total proteins in the lysate of Sf9 insect cells infected for 3 days with AcMNPV (Lane 1) and AcMNPV-WSSV-TK-TMK (Lane 2). Lane 3 shows recombinant WSSV TK-TMK protein purified by Ni-NTA resin. A protein ladder (lane M) was used as the molecular mass standard.
an ethyl group, but not bulky substitutions such as a bromovinyl group) or at the 3⬘-position of the sugar (e.g., hydrogen, halogens, or an azido group, but not arabino-furanosy- or acyclic sugars). In view of this, it is not surprising that similar to human cytosolic TK, WSSV TK is not affected by ACV, GCV, or BVdU (Fig. 4). Unfortunately, the fact that WSSV TKhas similar substrate preferences to the other cytosolic TKs means that it may be relatively difficult to develop anti-WSSV chemopeutic chemicals on the basis of nucleoside analogs. (Compare for instance the situation for herpesviruses, where the three nucleo-side analogs, ACV, GCV, and BVdU, are all preferential substrates for herpesvirus TKs and are thus potent and selective compounds against herpesviruses; see Elion et
al., 1977; Schaeffer et al., 1978; De Clercq et al., 1979;
Smith et al., 1982; De Clercq, 1983.)
TTP feedback inhibition of WSSV TK activity
The addition of TTP to the TKassay solution caused the WSSV TKactivity of rWSSV-TK-TMKto drop
dramati-cally. In the presence of 10M, 100 M, and 1 mM TTP, the TKactivity decreased to 85, 57, and 17%, respectively; no significant change was observed in the presence of 1 mM CTP (Fig. 5). These data suggest that, similar to cytosolic TKs (but unlike herpesvirus TK; see Cheng, 1976), the TKactivity of rWSSV-TK-TMKis sensitive to feedback inhibition by TTP.
TK activity in WSSV-infected shrimp organs
A previous study has shown that the transcription levels of the WSSV tk-tmk gene increased over the first 18 h postinfection (Tsai et al., 2000b). To test whether TK activity is coupled with WSSV tk-tmk transcription during infection, crude extracts were prepared from the integu-ment, gills, and hearts of artificially infected Penaeus
monodon shrimp for a TKactivity assay. [Note that these
FIG. 3. Relative amounts of TMP formed with different phosphate donors (as labeled) or without any phosphate donor (*) in the TKactivity assay for rWSSV-TK-TMK. TK activity was determined at 37°C and pH 8.0 with 46M of thymidine and 5 mM of NTP or dNTP. Data are mean values (n⭌ 3).
FIG. 4. Relative amounts of TMP formed (TKactivity) in the presence of competing nucleoside substrates. TKactivity was determined in the presence or absence of various competing nucleosides at 37°C and pH 8.0 with 46M of thymidine (T) and 4.8 mM of ATP. Data are mean values (n⭌ 3).
FIG. 2. Substrate concentration vs initial velocity of the recombinant WSSV TK. TK activity was determined at 37°C and pH 8.0 with (A) ATP at 4.8 mM and thymidine varying from 1 to 150M; (B) thymidine at 46 M and ATP varying from 0.03 to 5 mM. Data are mean values (n ⭌ 3).
extract were prepared from organs that had been stored in liquid nitrogen, because our pilot studies suggested that WSSV TKactivity does not decrease significantly during storage (data not shown).] By 60 h p.i. at both 37 and 25°C, activity increased markedly in the integument and to a lesser extent in the gill (Fig. 6). By contrast, in the heart, the muscle cells of which are not usually targeted by WSSV increases in TKactivity over time were only very slight. TKactivity was also generally higher at 37°C than at 25°C (cf. Figs. 6A and 6B).
Viral TKis often regarded as a virulence factor of viruses in vivo, even though viruses that are deficient in TKcan often multiply readily in cell culture (i.e., where resources are abundant) (Tenser, 1991; Buller et al., 1985; Buller and Palumbo, 1991). Both TKand dCKare impor-tant because they phosphorylate thymidine and dC, re-spectively, and if pyrimidine nucleosides such as thymi-dine and dC are not phosphorylated, they could other-wise cross the cell membrane and leak out into the surrounding medium (Plagemann and Erbe, 1972; Bian-chi et al., 1987; Hoglund et al., 1988; Gehring and Schro¨-der, 1991, Gentry, 1992). So far, dCKhas not been found in WSSV, which suggests that the virus is probably using host cell dCKto prevent the leakage of dC. In most cell types, dCKis expressed constitutively and is thus avail-able throughout the cell cycle; by contrast, TKactivity is strictly S phase correlated (Arner and Eriksson, 1995). Thus by expressing its own TKin infected cells, WSSV provides itself with a continuously available mechanism for preventing wastage of the thymine-containing com-pounds that have been derived from the TMP pool through dephosphorylation or DNA degradation. WSSV’s probable inability to express dCK, on the other hand, may help to account for the tissues/cell types that WSSV targets. In mammals, some cell types (notably brain, muscle, and colon cells) do not express dCK. It is not yet known whether dCKis expressed in shrimp muscle cell, but if it is not, then this may explain why WSSV attacks almost every other cell type derived from ectoderm and mesoderm (Wongteerasupaya et al., 1995), but does not target shrimp muscle cells.
MATERIALS AND METHODS Virus and plasmid clone
The WSSV plasmid libraries used in this study were constructed by Wang et al. (1995). These libraries were based on the WSSV Taiwan isolate (Lo et al., 1999), which was originally isolated from a batch of WSSV-infected P. monodon collected in Taiwan in 1994 (Wang et
al., 1995). The pms library (where pm indicates P. mon-odon and s indicates SalI fragments) contains a WSSV
genomic DNA fragment (pms94) with an ORF that shows high homology to TKand TMK.
Expression of rWSSV-TK-TMK
rWSSV-TK-TMK was expressed in Sf9 insect cells. The coding region of WSSV-TK-TMK was amplified from the plasmid clone containing the pms94 fragment. The 5⬘ primer used for the polymerase chain reaction (PCR) was ATG CAA CTC GAG CTT TCT CAT CAT CTA, and the 3⬘ primer was TAC CCC GGG TTA TTC TTC AAC AAT ATT; the underlined bases indicate the restriction sites. The 5⬘ primer was designed to insert an XhoI site immediately 5⬘ to nt ⫺24, and the 3⬘ primer was designed to insert a
SmaI site immediately 3⬘ to nt 1167. Twenty-five cycles consisting of 45 s at 94°C, 45 s at 50°C, 80 s at 72°C were run using 50 ng pms94, 0.25g of each primer, 200
FIG. 6. TKactivity of desalted total soluble proteins from various organs of WSSV infected Penaeus monodon shrimps at different times postinfection. TKactivity was determined at pH 8.0 with 46M of thymidine and 4.8 mM of ATP (A) at 37°C and (B) at room temperature (25°C). Data are mean values for the triplicated assays. Bars show standard deviations.
FIG. 5. TTP inhibition of recombinant WSSV TKactivity. TKactivity was determined at 37°C and pH 8.0 with 46M of thymidine and 4.8 mM of ATP in the presence of CTP (E) or TTP (F). Data are mean values (n⭌ 3).
M of each dNTP, and 1 unit of TaKaRa Ex Taq DNA polymerase. The PCR product was cloned into pGEM-T easy according to the manufacturer’s instructions (Pro-mega, Madison, WI). The resulting construct was desig-nated pGEM-T-TK-TMK. The plasmids pGEM-T-TK-TMK and pAcSG His NT-A (Pharmingen, San Diego, CA) were digested with the restriction endonucleases XhoI and
SmaI. The oligonucleotides were purified by 1% agarose
gel electrophoresis and then extracted with the QIAquick gel extraction kit (Qiagen, Hilden, Germany). The TK insert was ligated with XhoI- and SmaI-digested pAcS-GHisNTA. The resulting transfer construct, designated pAcSGHisNTA-TK-TMK, was purified by QIAprep spin miniprep kit (Qiagen) and quantified by spectrophotom-eter at 260 nm.
The transfer plasmid pAcSGHisNTA-TK-TMK was co-transfected with triple-cut baculovirus AcMNPV DNA into Sf9 insect cells according to the manufacturer’s instruc-tions (Novagen, Madison, WI). The resulting recombinant virus AcMNPV-TK-TMK was confirmed by PCR, limiting dilution, and dot-hybridization. The Sf9 insect cells (3⫻ 106) were infected by AcMNPV-TK-TMK virus solution (0.3 ml) with 5–10 m.o.i. for 3 days at 27°C, and then the cells were harvested by centrifugation at 1000 g for 15 min at 4°C and stored at⫺20°C until use.
Purification of rWSSV-TK-TMK
The cell pellet from one 25-ml flask was resuspended in 0.4 ml binding buffer (20 mM Tris–HCl at pH 8.0, 0.3 M sodium chloride, and 10 mM imidazole), and the suspen-sion was sonicated with three 10-s bursts at 80% power with the microtip of a dr.hielscher UP-50H. The lysate was centrifuged at 20,000 g for 20 min, and the super-natant was mixed with 0.2 ml Ni-NTA resin (Qiagen) that had been equilibrated with binding buffer. The resin was washed with 1 ml of binding buffer twice and then with 1 ml of wash buffer (20 mM Tris–HCl at pH 8.0, 0.3 M sodium chloride, and 40 mM imidazole) twice. The pro-tein was eluted with 0.3 ml elution buffer (20 mM Tris– HCl at pH 8.0, 0.3 M sodium chloride, and 0.25 M imida-zole), and dialyzed against 20 mM Tris–HCl at pH 8.0, 50 mM sodium chloride, and 25 mM imidazole. Then glyc-erol was added to 30% and the resulting solution was stored at ⫺20°C until use. The concentration of total protein was determined using Bio-Rad protein assay solution (Bio-Rad, Hercules, CA) with bovine serum al-bumin as the standard.
Preparation of crude extracts from various organs of WSSV-infected shrimps
WSSV inoculum was prepared and P. monodon were artificially infected with WSSV as previously described (Liu et al., 2001). From three WSSV-infected P. monodon shrimps (one specimen at 0 h p.i., one at 18 h p.i., and the third at 60 h p.i.) the integument, gills, and heart were
separately removed, frozen, and stored in liquid nitrogen until use. These organs were then washed twice with an extraction buffer containing 20 mM HEPES, pH 7.5, 400 mM NaCl, 1 mM DTT with complete protease inhibitor cocktail from Roche Diagnostics GmbH (Mannheim, Ger-many). Each washed organ was then placed in a micro-centrifuge tube and an aliquot (0.2 ml) of fresh extraction buffer was added. After homogenization with a wooden splint, the mixture was sonicated with 10-s bursts and centrifuged at 20,000 g for 20 min. To ensure that there was no interference from endogenous nucleotides such as TTP, the supernatant was loaded onto a Sephadex G-25 column (⬃2 ml) equilibrated with the extraction buffer. The crude extracts were collected from the G-25 columns by centrifugation, and their concentrations were determined using Bio-Rad protein assay solution. Note that a general (as opposed to an optimized) extraction buffer was used here, but the extraction procedure was verified by mixing purified rWSSV-TK-TMK with unin-fected tissues, and after the desalting step,⬃95% of the TKactivity was recovered. The crude extracts (⬃74g) then served as the sources of TKfor the enzyme assays (50 l per assay; each assay performed in triplicate) at 37°C or at room temperature (25°C) as described below.
Enzyme assays
The enzyme assays were performed at 37°C or at room temperature (25°C) for 20 min as previously de-scribed (Cheng and Ostrander, 1976) with slight modifi-cation. Unless otherwise specified, the standard thymi-dine kinase assay mixture contained 25 ng purified rWSSV-TK-TMK, 46 M [methyl-3H]thymidine (6.7 Ci/ mmol, from Perkin–Elmer Life Sciences, Boston, MA), 4.8 mM Mg2⫹ATP, 50 mM Tris–HCl (pH 8.0), 1 mg/ml of bovine serum albumin, 30 mM NaF, 0.5 mM PMSF, and 0.014%-mercaptoethanol in a final volume of 50 l. The reaction was stopped by adding ice-cold stop buffer (25 l of 9 mM thymidine) and spotting 30 l of the final mixture on a Whatman DE-81 filter paper. After air-drying, the filters were washed three times with 1 mM ammo-nium formate (5 ml/filter), twice with methanol (5 ml/ filter), and once with acetone (5 ml/filter). The washed filters were air-dried and inserted into vials containing 5 ml of a scintillation solution (FluoranSafe 2 Scintran; BDH Poole, Dorset, U.K.). Radioactivity counting was per-formed in a Wallac liquid scintillation counter LKB 1219 RACKBETA.
Kinetic analysis
The kinetic parameters, Kmand Vmax, were determined
from two sets of enzyme assays using the same methods as described above. The concentration of thymidine was varied from 1 to 150M at a fixed concentration of 4.8 mM ATP. The ATP concentration was varied from 0.03 to
5 mM at a fixed concentration of thymidine (46M). Data were applied to the equation v ⫽ Vmax[S]/(Km ⫹ [S]),
where [S] is the substrate concentration, by means of a computer program (Sigma Plot 5; Jandel Scientific, Sigma) that provides an iterative nonlinear least-squares fit to the best rectangular hyperbola.
ACKNOWLEDGMENTS
This work was supported by the Council of Agriculture Grant 89-BT-2.1-FID-01(13), the National Science Council Grants NSC89-2311-B-002-042 and NSC89–2316-B-260-001, and Education Administration Grant 89-B-FA01-1-4. We are indebted to Paul Barlow for helpful criticism of the manuscript.
REFERENCES
Arner, E. S. J., and Eriksson, S. (1995). Mammalian deoxyribonucleoside kinases. Pharmacol. Ther. 67, 155–186.
Berenstein, D., Christensen, J. F., Kristensen, T., Hofbauer, R., and Munch-Pertersen, B. (2000). Valine, not methionine, is amino acid 106 in human cytosolic thymidine kinase (TK1). J. Biol. Chem. 275, 32187– 32192.
Bianchi, V., Pontis, E., and Reichard, P. (1987). Regulation of pyrimidine deoxyribonucleotide metabolism by substrate cycles in dCMP deaminase-deficient V79 hamster cells. Mol. Cell. Biol. 7, 4218–4224. Buller, R. M. L., Smith, G. L., Cremer, K., Notkins, A. L., and Moss, B. (1985). Decreased virulence of recombinant vaccinia virus expres-sion vectors is associated with thymidine kinase-negative pheno-type. Nature 354, 520–522.
Buller, R. M. L., and Palumbo, G. J. (1991). Poxvirus pathogenesis.
Microbiol. Rev. 55, 80–122.
Chen, L. L., Leu, J. H., Huang, C. J., Chou, C. M., Chen, S. M., Wang, C. H., Lo, C. F., and Kou, G. H. (2002). Identification of a nucleocapsid protein (VP35) gene of shrimp white spot syndrome virus and char-acterization of the motif important for targeting VP35 to the nuclei of transfected insect cells. Virology 293, 44–53, doi:10.1006/ viro.2001.1273.
Chen, M. S., and Prusoff, W. H. (1978). Association of thymidylate kinase activity with pyrimidine deoxyribonucleoside kinase induced by her-pes simplex virus. J. Biol. Chem. 253, 1325–1327.
Chenal-Francisque, V., Tourneux, L., Carniel, E., Christova, P., de la Sierra, I. L., Baˆrzu, O., and Gilles, A. M. (1999). The highly similar TMP kinases of Yersinia pestis and Escherichia coli differ markedly in their AZTMP phosphorylating activity. Eur. J. Biochem. 265, 112–119. Cheng, Y. C. (1976). Deoxythymidine kinase induced in HeLa TK⫺cells
by herpes simplex virus type I and type II. Biochim. Biophys. Acta 452, 370–381.
Cheng, Y. C., and Ostrander, M. (1976). Deoxythymidine kinase induced in HeLa TK⫺ cells by herpes simplex virus type I and type II. II.
Purification and characterization. J. Biol. Chem. 251, 2605–2610. Chou, H. Y., Huang, C. Y., Wang, C. H., Chiang, H. C., and Lo, C. F. (1995).
Pathogenicity of a baculovirus infection causing white spot syn-drome in cultured penaeid shrimp in Taiwan. Dis. Aquat. Org. 23, 165–173.
De Clercq, E., Descamps, J., De Somer, P., Barr, P. J., Jones, A. S., and Walker, R. T. (1979). (E)-5-(2-Bromovinyl)-2⬘-deoxyuridine: A potent and selective anti-herpes agent. Proc. Natl. Acad. Sci. USA 76, 2947–2951.
De Clercq, E. (1983). Antiviral activity of 5-substituted pyrimidine nucle-oside analogs. Pure Appl. Chem. 55, 623–626.
Elion, G. B., Furman, P. A., Fyfe, J. A., de Miranda, P., Beauchamp, L., and Schaeffer, H. J. (1977). Selectivity of action of an antiherpetic agent, 9-(2-hydroxyethoxymethyl) guanine. Proc. Natl. Acad. Sci. USA 74, 5716–5720.
Eriksson, S., Kierdaszuk, B., Munch-Petersen, B., Oberg, B., and Johan-sson, N. G. (1991). Comparison of the substrate specificities of human thymidine kinase 1 and 2 and deoxycytidine kinase toward antiviral and cytostatic nucleoside analogs. Biochem. Biophys. Res.
Commun. 176, 586–592.
Flegel, T. W. (1997). Special topic review: Mayor viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J. Microbiol.
Biotechnol. 13, 433–442.
Gehring, H., and Schroder, A. (1991). Thymidine secretion by cultured chicken embryo fibroblasts and NIH/3T3 cells: Quantification and time course. Biochem. Biophys. Res. Commun. 177, 259–264. Gentry, G. A. (1992). Viral thymidine kinases and their relatives.
Phar-macol. Ther. 54, 319–355.
Hoglund, L., Pontis, E., and Reichard, P. (1988). Effects of deoxycytidine and thymidine kinase deficiency on substrate cycles between de-oxyribonucleosides and their 5-phosphates. Cancer Res. 48, 3681– 3687.
Inouye, K., Miwa, S., Oseko, N., Nakano, H., and Kimura, T. (1994). Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, in Japan in 1993: Electron microscopic evidence of the causative virus. Fish
Pathol. 29, 149–158 (in Japanese).
Kit, S., and Dubbs, D. R. (1965). Properties of deoxythymidine kinase partially purified from noninfected and virus-infected mouse fibro-blast cells. Virology 26, 16–27.
Kit, S. (1985). Thymidine kinase. Microbiol. Sci. 2, 369–375.
Kit, S., Leung, W.-C., Jorgensen, G. N., Trkula, D., and Dubbs, D. R. (1974). Thymidine kinase isozymes of normal and virus-infected cells.
Cold Spring Harbor Symp. Quant. Biol. 39, 703–705.
Liu, W. J., Yu, H. T., Peng, S. E., Chang, Y. S., Pien, H. W., Lin, C. J., Huang, C. J., Tsai, M. F., Huang, C. J., Wang, C. H., Lin, J. Y., Lo, C. F., and Kou, G. H (2001). Cloning, characterization and phylogenetic analysis of a shrimp white spot syndrome virus (WSSV) gene that encodes a protein kinase. Virology 289, 262–77.
Lo, C. F, Ho, C. H., Peng, S. E., Chen, C. H., Hsu, H. C., Chiu, Y. L., Chang, C. F., Liu, K. F., Su, M. S., Wang, C. H., and Kou, G. H. (1996). White spot syndrome baculovirus (WSBV) detected in cultured and cap-tured shrimp, crabs and other arthropods. Dis. Aquat. Org. 27, 215– 225.
Lo, C. F., Ho, C. H., Chen, C. H., Liu, K. F., Chiu, Y. L., Yeh, P. Y., Peng, S. E., Hsu, H. C., Liu, H. C., Chang, C. F., Su, M. S., Wang, C. H., and Kou, G. H. (1997). Dectection and tissue tropism of white spot syn-drome baculovirus (WSBV) in captured brooders of Penaeus
mon-odon with a special emphasis on reproductive organs. Dis. Aquat. Org. 30, 53–72.
Lo, C. F., and Kou, G. H. (1998). Virus-associated white spot syndrome of shrimp in Taiwan: A review. Fish Pathol. 33, 365–371.
Lo, C. F., Hsu, H. C., Tsai, M. F., Ho, C. H., Peng, S. E., Kou, G. H., and Lightner, D. V. (1999). Specific genomic fragment analysis of different geographical clinical samples of shrimp white spot syndrome virus.
Dis. Aquat. Org. 35, 175–185.
Munch-Petersen, B., Cloos, L., Tyrsted, G., and Eriksson, S. (1991). Diverging substrate specificity of pure human thymidine kinases 1 and 2 against antiviral dideoxynucleosides. J. Biol. Chem. 266, 9032– 9038.
Peng, S. E., Lo C. F., Lin S. C., Chen L. L., Chang Y. S., Su M. S., and Kou G. H. (2001). Performance of WSSV-infected and WSSV-negative
Penaeus monodon postlarvae in culture ponds. Dis. Aquat. Org. 46,
165–172.
Plagemann, P. G. W., and Erebe, J. (1972). Thymidine transport by cultured Novikoff hepatoma cells and uptake by simple diffusion and relationship to incorporation into deoxyribonucleic acid. J. Cell Biol. 55, 161–178.
Reichard, P. (1988). Interactions between deoxyribonucleotide and DNA synthesis. Annu. Rev. Biochem. 57, 349–374.
Schaeffer, H. J., Beauchamp, L., de Miranda, P., Elion, G. B., Bauer, D. J., and Collins, P. (1978). 9-(2-Hydroxyethoxymethyl)guanine activity against viruses of the herpes group. Nature 272, 583–585.
Smith, K. O., Galloway, K. S., Kennell, W. L., Ogilvie, K. K., and Radatus, B. K. (1982). A new nucleoside analog, 9-[[2-hydroxy-1-(hydroxymeth-yl)ethoxyl]methyl]guanine, highly active in vitro against herpes sim-plex virus types 1 and 2. Antimicrob. Agents Chemother. 22, 55–61. Stubbe, J. (1990). Ribonucleotide reductases. Adv. Enzymol. Relat. Areas
Mol. Biol. 63, 349–419.
Tenser, R. B. (1991). Role of herpes simplex virus thymidine kinase expression in viral pathogenesis and latency. Intevirology. 32, 76–92. Thelander, L., and Reichard, P. (1979). Reduction of ribonucleotides.
Annu. Rev. Biochem. 48, 133–158.
Tsai, M. F., Lo, C. F., van Hulten, M. C. W., Tzeng, H. F., Chou, C. M., Huang, C. J., Wang, C. H., Lin, J. Y., Vlak, J. M., and Kou, G. H. (2000a). Transcriptional analysis of the ribonucleotide reductase genes of shrimp white spot syndrome. Virology 277, 92–99, doi:10.1006/ viro.2000.0956.
Tsai, M. F., Yu, H. T., Tzeng, H. F., Leu, J. H., Chou, C. M., Huang, C. J., Wang, C. H., Lin, J. Y., Kou, G. H., and Lo, C. F. (2000b). Identification and characterization of a shrimp white spot syndrome virus (WSSV) gene that encodes a novel chimeric polypeptide of cellular-type thymidine kinase and thymidylate kinase. Virology 277, 100–110, doi:10.1006/viro.2000.0957.
van Hulten, M. C. W., Witteveldt, J., Peters, S., Kloosterboer, N., Tarchini, R., Fiers, M., Sandbrink, H., Lankhorst, R. K., and Vlak, J. M. (2001). The white spot syndrome virus DNA genome sequence. Virology 286, 7–22, doi:10.1006/viro.2001.1002.
Wang, C. H., Lo, C. F., Leu, J. H., Chou, C. M., Yeh, P. Y., Chou, H. Y., Tung, M. C., Chang, C. F., Su, M. S., and Kou, G. H. (1995). Purification and genomic analysis of baculovirus associated with white spot syn-drome (WSBV) of Penaeus monodon. Dis. Aquat. Org. 23, 239–242. Wang, C. H., Yang, H. N., Tang, C. Y., Lu, C. H., Kou, G. H., and Lo. C. F.
(2000). Ultrastructure of white spot syndrome virus development in primary lymphoid organ cell cutlures. Dis. Aquat. Org. 41, 91–104. Wongteerasupaya, C., Vickers, J. E., Sriurairatana, S., Nash,G. L.,
Akara-jamorn, A., Boonsaeng, V., Panyim, S., Tassanakajon, A., Withya-chumnarnkul, B., and Flegel, T. W. (1995). A non-occluded, systemic baculovirus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in the black tiger prawn Penaeus
monodon. Dis. Aquat. Org. 21, 69–77.
Yang, F., He, J., Lin, X., Li, Q., Pan, D., Zhang, X., and Xu, X. (2001). Complete genome sequence of the shrimp white spot bacilliform virus. J. Virol. 75, 11811–11820.