國立交通大學
機械工程學系
碩士論文
細胞伸展和突出行為之數值模擬
Numerical Simulation of Cell Spreading and
Protrusion
研 究 生:廖乾廷
指導教授:陳大潘 博士
細胞伸展和突出行為之數值模擬
Numerical Simulation of Cell Spreading and Protrusion
研 究 生:廖乾廷 Student:Chien-Ting Liao
指導教授:陳大潘 Advisor:Dr.Da-Pan Chan
國 立 交 通 大 學 機 械 工 程 學 系 碩 士 論 文 A ThesisSubmitted to Department of Mechanical Engineering College of Engineering
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master of Science
in
Mechanical Engineering July 2008
Hsinchu, Taiwan, Republic of China
細胞伸展和突出行為之數值模擬
研究生:廖乾廷 指導教授:陳大潘 博士 國立交通大學機械工程學系(研究所)碩士班摘 要
本論文利用有限元素方法對生物細胞之細胞膜張力做模擬分 析。模擬方向主要分為兩大部分,一為探討紅血球細胞的細胞膜張力 和楊氏系數之間關係。另一方面探討,白血球細胞的細胞膜張力和偽 足突出之間關係,而後整理兩部分之結果資料並與相關的生物研究報 告之數據做比較,成功的利用機械工程科學模擬了生物細胞之行為表 現,亦代表了工程科學理論於生物醫學方面可佔有相當的發揮空間。
Numerical Simulation of Cell Spreading and Protrusion
Student:Chien-Ting Liao Advisor:Dr.Da-Pan Chan
Department(Institute)of Mechanical Engineering National Chiao Tung University
Abstract
This research employs finite elements for the simulation of the spreading and protrusion of biocell. The simulation has two concentrations. One is to study theerythrocyte cell membrane tension and elastic modulus relations, and the other is to study neutrophil cell membrane tension and pseudopod protrusion relations. Our finite element predications are in consistence with experimental results. These works bring out the opportunity to simulate biocell phenomena with computational mechanics.
誌 謝 二年的碩士班生活,在陳大潘教授的細心的指導和教誨之下,讓 我在專業知識上有所進步,能以正確的方法及嚴謹的態度來從事研究 工作,在此致上最誠摰的敬意與感謝。 此外還要感謝實驗室的博士班學長,在我有疑惑時,能提供我新 的思考方向順利解決問題。還要感謝一起努力了二年的同學及相處一 年的學弟,能夠在論文上提供我意見,謝謝幫助過我的師長和同學。 最後感謝我的父母親與家人,謝謝他們在這二年來不斷地的給我 加油和鼓勵,在他們的支持下,我才能夠無後顧之憂的完成學業。
目錄 中文摘要... i 英文摘要...ii 誌謝...iii 目錄...iv 圖目錄...vi 表目錄...viii 第一章 緒論...1 第二章 細胞簡介...3 2-1 細胞概論...3 2-2 細胞的基本結構和功能...3 2-2-1 原核細胞與真核細胞...3 2-2-2 細胞骨架...4 2-2-3 細胞質...4 2-2-4 細胞膜...6 2-3 細胞遷移運動(cell migration) ...8 2-4 高分子之聚合(Polymerization)...12 2-5 偽足...13 2-6 紅血球細胞介紹...14 2-7 細胞與細胞間及細胞與基質間黏附關係...17
第三章、研究架構和方法及理論介紹...20 3-1 研究方向...20 3-2 研究工具簡介...20 3-2-1 ANSYS 簡介...21 3-2-2 MATLAB 簡介...26 3-2-3 Amira 軟體簡介...26 3-3 研究方法...27 第四章、紅血球黏附力所引起的細胞膜張力...30 第五章、白血球細胞伸展行為和偽足突出之模擬分析...40 5-1 細胞伸展行為模擬流程...40 5-2 細胞在平板上的偽足突出行為模擬...47 5-2-1 單根偽足之模擬...47 第六章、結論 6-1 結果比較...52 6-2 未來展望...53 參考文獻...54
圖目錄 圖 2-1 細胞組織圖...4 圖 2-2 細胞骨架元素圖,由左而右分別為微纖維、中間絲及微管....5 圖 2-3 細胞骨架在細胞內的排列情形...5 圖 2-4 細胞膜結構之「流體鑲嵌模型」...7 圖 2-5 細胞遷移系列反應示意圖...9 圖 2-6 細胞運動示意圖...11 圖 2-7 細胞遷移簡易示意圖...12 圖 2-8 球狀肌動蛋白(G-Actin)簡圖...13 圖 2-9 紅血球結構圖...15 圖 2-10 紅血球在不同濃度溶液的現象(箭頭為水份進出方向) ...16 圖 2-11 細胞與細胞間的黏附分子(CAMs)示意圖...19 圖 2-12 細胞與細胞外基質(ECM)間的黏著示意圖...19
圖 3-1 全牛頓-拉夫森(Full Newton-Raphson Method)疊代法...23
圖 3-2 ANSYS 分析步驟...25 圖 3-3 細胞伸展行為示意簡圖(L1、L2 為底部接觸長度) ...28 圖 3-4 (a)十二根微絲之肌動蛋白微絲束幾何結構排列圖...29 (b)其排列截面圖 圖 3-5 肌動蛋白微絲束推動細胞膜之簡圖...29 圖 4-1 紅血球模型圖(直徑 6μm,厚度 2nm)...32 圖 4-2 模型拘束邊界示意圖...32 圖 4-3 接觸元素對(圖中的線代表接觸面的法線方向) ...32 圖 4-4 (a)~(h)細胞膜各階段變形程度 (E=15MPa) ...33 圖 4-5 (a)~(h)細胞膜各階段表面張力值 (E=15MPa) ...34
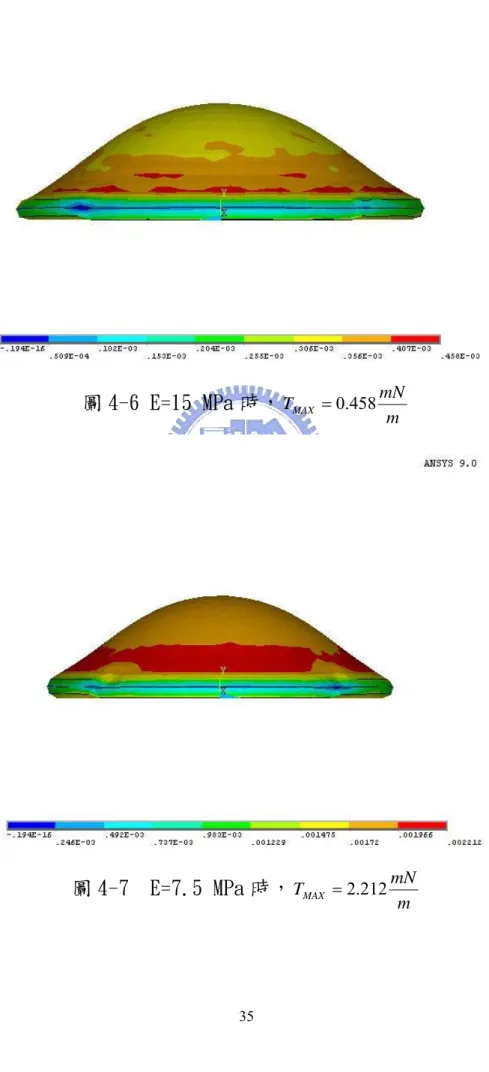
圖 4-6 E=15 MPa 時,TMAX =0.458mN m...35
圖 4-7 E=7.5 MPa 時,TMAX =2.212mN m...35
圖 4-8 E=11.5 MPa 時,TMAX =1.405mN m...36
圖 4-9 E=13.5 MPa 時,TMAX =0.819mN m...36
圖 4-10 E=12.5 MPa 時,TMAX =1.254mN m...37
圖 4-11 E=13 MPa 時,TMAX =0.981mN m...37
圖 4-12 E=12.75 MPa 時,TMAX =1.092mN m...38
圖 4-13 E=12.88 MPa 時,TMAX =1.004mN m...38
圖 5-1 細胞模型(直徑 10μm,厚度 3nm)...41
圖 5-2 內壓力 2 0 12pN m P = μ 時之表面張力...41
圖 5-3 利用 Amira 軟體計算體積V0和面積A0...42 圖 5-4 細胞和平板模型示意圖...44 圖 5-5 模型拘束邊界示意圖...44 圖 5-6 「接觸元素對」示意圖(圖中的線代表接觸面的法線方向) ...44 圖 5-7 (a)~(f) 細胞膜各階段變形程度...45 圖 5-7 (a)~(f) 細胞膜各階段表面張力值...46 圖 5-8 細胞模型圖(偽足突出部位之網格加密) ...48 圖 5-9 (a)十二根微絲之肌動蛋白微絲束幾何結構排列圖...48 (b)其排列截面圖偽足 圖 5-10 偽足和細胞膜接觸圓直徑...49 圖 5-11 邊界條件設定:固定細胞膜和平板接觸部份,並在直徑 49nm 圓上施加負載...49 圖 5-12 細胞偽足突出變形圖...50 圖 5-13 細胞膜不同程度的突出示意圖...50 圖 5-14 施加負載 F 與偽足突出長度之關系圖...51 圖 6-1 偽足突出長度和負載關係...53
表目錄:
表 4-1 細胞各階段變形資料 (E=15 MPa) ...39
表 4-2 不同 E 值所對應的 Tmax 值...39
表 5-1 細胞各階段變形資料...46
第一章、 緒論 生醫工程是一個跨學門的領域,將工程科學之理論及技術應用於醫學 的研究及發展,其中生物力學更是將這兩大領域結合的關鍵。麻省理工學 者蘇爾什曾提出:力學在疾病的控制及醫療上有極大可發揮的空間。以目 前所知,機械外力對於細胞有著甚為顯著之影響,例如機械壓力能大幅提 昇軟骨細胞的成長,血管內血流所產生的切力能促進血管內皮細胞之正常 分佈及排列。近來由於電子掃描顯微技術及雷射光鉗技術之快速演進,成 為生物力學及細胞研究之兩大工具,細胞力學因而有長足之進展。在細胞 力學領域中,細胞之遷移運動(Cell Migration)尤其在最近成為醫學界 針對癌症之預防及治療研究之重點。因為癌症之發展在初期有擴散轉移之 階段,因此了解細胞遷移運動之機制,可在癌症之預防及治療上提供有效 的控制。 在細胞生物學裡,細胞之遷移運動為細胞運動(Cell Motility)大綱 內之一分支,其主要之運動機構為細胞內部之肌動蛋白(Actin)以高分子 合成(Polymerization)之機制合成一較具剛度(Stiffnes)之纖維束 ( Filament Bundle ), 向 前 延 伸 一 適 當 之 距 離 而 形 成 細 胞 突 出 (Protrusion),此即為細胞運動之重要器官—偽足(Filopodium)。細胞 以此往前延伸之偽足吸附於細胞外之介質,再以肌動蛋白纖維束之收縮力
將細胞之其餘部分往前拖移而完成往前之ㄧ小步,此爬行(Cell Crawling) 之連續重複動作將細胞遷移至目的地。 在生物學領域裡細胞運動已具相當成熟之理論。本研究擬以機械力學 之有限元素來模擬「紅血球」和「白血球」細胞,以適當之有限元素模擬 細胞之皮膜,計算細胞伸展行為細胞內壓力施加在皮膜上的力量和位移, 從而得到整個運動過程的能量變化。將此比較抽象之生物現象作機械力學 方式之具體表現。研究重點有 2 點: 1. 紅血球細胞在平板上做伸展行為黏附到平板時,楊氏係數和細胞膜表面 張力之間關係。 2. 模擬白血球細胞在平板上的伸展行為和偽足突出行為。 以下為本研究論文之內容:第二章為生物細胞類別概論,並簡介紅血 球細胞和真核細胞之各種組織,且對於細胞能動性、偽足的形成、細胞內 部發生的高份子之聚合現象及細胞黏附關係做介紹,第三章為研究架構和 方法作描述,並介紹研究工具,第四章以有限元素軟體—ANSYS 建立細胞膜 模型,以之進行模擬紅血球細胞因黏附行為所引起的細胞膜張力,第五章 為白血球細胞伸展行為和偽足突出之模擬。第六章便針對第四、五章所得 之結果作討論,並與相關的生物細胞研究報告做比較。
第二章、 細胞簡介 2-1 細胞概論 細胞是生物活動的基本單位,一般而言生物可分為單細胞生物及多細 胞生物兩大類,前者如細菌等絕大部分微生物及原生動物,而高等植物與 高等動物則為多細胞生物。細胞又可分為兩類:原核細胞與真核細胞,原 核細胞內之細胞核之界限並不分明,與其他細胞質混合為一體。真核細胞 之細胞核有一層核膜,藉以和其他細胞質作明顯之分隔,可參考圖 2-1。 2-2 細胞的基本結構和功能 2-2-1 原核細胞與真核細胞 原核細胞是比較低級和原始的一類細胞。其主要特點是沒有成形的細 胞核,遺傳物質散在於細胞質中。除核糖體外,沒有成形的細胞器。 真核微生物細胞,通常具有細胞膜(Membrane)、細胞核(Nucleus)、 細胞骨架(Cytoskeleton)、細胞質(Cyotplasm)…等多種組織,可參考 圖 2-1。 細胞核中含有染色質,而其於細胞分裂期時將會濃縮形成染色體,並 儲存細胞的遺傳訊息、控制細胞代謝、指導蛋白質合成,故細胞核為真核 細胞中相當重要的細胞器。
2-2-2 細胞骨架 細胞骨架主要由微管(Microtubules)、微纖維(Microfilament)、中間 絲(Intermediate Filament)等三種纖維組成。微管呈中空管狀,為細胞骨 架的架構主幹。微纖維是實心狀的纖維,能確定細胞表面特徵並使細胞運 動和收縮。中間絲為中空的骨狀結構,其化學組成比較複雜,在不同細胞 中成分變化較大,且中間纖維使細胞具有張力和抗剪切力,可參考圖 2-2、 2-3。而細胞骨架提供細胞機械性支撐用以維持細胞型態、承受外力,保持 細胞內部結構的有序性等方面起了重要作用,且可隨細胞的生命活動而呈 現高度的動態性,於細胞運動中亦扮演相當重要角色。 2-2-3 細胞質 細胞質包含了多種元素,視乎環境情況及當時細胞的活動,可以是凝膠 狀亦可為可流動的液體,在細胞內有傳送或聚集的功能。
圖 2-2 細胞骨架元素圖,由左而右分別為微纖維、中間絲及微管
2-2-4 細胞膜 生物細胞膜的發展對早期生命的發展是一個相當重要的關鍵,細胞膜 包圍著每個細胞而界定了細胞的範圍,並維持細胞內含物與其環境有相當 的差異,但它並非僅是一層被動的障礙物,更為一層具有高度選擇性的過 濾門戶,維持著膜內外兩邊不同的離子濃度,且可使養分進入細胞而排出 代謝廢物。 儘管目前還沒有一種能夠直接觀察膜的分子結構的較為方便的技術和 方法,但從 1930 年代以來提出了各種假說有數十種,其中得到較多實驗事 實 支 持 而 目 前 仍 為 大 多 數 人 所 接 受 的 是 美 國 的 S.J.Singer 和 G.L.Nicholsom 於 1972 年提出的流體鑲嵌模型(fluid mosaic model),可 參考圖 2-4。 「流體鑲嵌模型」是目前被最廣泛接受和認可的觀點。這種觀點主張, 構成膜的蛋白質和脂類分子具有鑲嵌關係,而且膜的結構處於流體變化之 中。許多實驗結果支持流體鑲嵌模型的模式。關於細胞的流體鑲嵌模型, 有個著名的實驗。科學家取了兩顆細胞,在兩顆細胞的蛋白質上分別染色, 再將他們倆個擠在一起,結果竟然能融合,而且兩細胞的蛋白質分子也混 和在一起(不是一半一半),這證明了細胞膜與其上的蛋白質都可以流動在 流體鑲嵌模型學說中,細胞膜具有流動性,也就是說,他並不是像我們的保 鮮膜一樣是固體,而是液體膜中的磷脂質分子以雙層排列,構成了膜的網
架,是膜的基質。磷脂質分子為雙性分子,分為親水頭端和疏水尾端,雙 層磷脂質分子之頭端皆朝向水相,疏水尾端則兩兩相接埋於膜內。而使脂 雙層分子之親水頭端的內層(面對細胞質之面)與外層(面對外界之面) 之結構不對稱原因,主要在於脂雙層分子兩親水頭端的化學組成不同。 圖 2-4 細胞膜結構之「流體鑲嵌模型」 膜的另一種主要成分是蛋白質,細胞膜上面具有許多蛋白質,而他們就 像是嵌在大教堂牆壁上小小的瓦片一樣,嵌在流動的細胞膜上。根據在膜 上存在位置的不同,膜蛋白可分為兩類,一是通過強疏水或親水作用同膜 脂牢固結合不易分開的,稱為整合蛋白(integral protein);二是附著在 膜的表層,與膜結合比較疏鬆容易分離的,稱為膜周邊蛋白(peripheral protein)。 細胞膜之主要功能有以下五點: 1. 維持細胞之結構完整。
2. 保護細胞之內部成分。
3. 為細胞內外選擇性物質輸送之通道。 4. 細胞表面絨毛、纖毛、鞭毛的著生位點。
5. 細胞抗原-抗體特異性識別的物質基礎和位置。
2-3 細胞遷移運動(cell migration)
於 1675 年科學家安東尼.凡 列文虎克(Antonie Van Leeuwenhoek) 發表了一份描述細菌運動的相關研究,而此發表亦開啟了科學家對於觀察 細胞運動的研究大門,至今由於新科技不斷創新、突破,對於細胞的運動 研究亦有極大的推動、幫助,我們對生物細胞的運動也有了相當的研究成 果。生物細胞的運動包括了細胞的整體運動和部份運動,可稱之為細胞能 動性(Cell Motility),其中整體運動代表細胞遷移(Cell Migration),
而部份運動則是指細胞的突出行為(Protrusion),亦是本研究之探討重點。 細胞的遷移運動主要為細胞在接受到遷移信號或感受到某物質的濃度 梯度後而產生的移動,參與細胞遷移的細胞外信號分子有相當多的種類, 如肽、代謝產物、細胞膜的殘片,但其作用皆是先與細胞膜表面的受體結 合而啟動細胞內訊號,接而完成一系列的反應,可參考圖 2-5,而所謂的濃 度梯度則是依離源(如細菌)的遠近,所產生不同的濃度定義,靠近源的 區域濃度梯度高,反之遠離源的區域濃度梯度低。
遷移過程中,細胞將不斷的重複伸出偽足來牽引胞體的循環,故細胞 突出行為可說是遷移運動的先驅。兩種行為雖然不全相同,但都有相同的 生化反應過程,包括極化現象(Polarisation)、細胞骨架的重新組織及 Cell-ECM 間黏著點的改變,ECM(Extracellular Matrix)為細胞外基質,是 由細胞分泌出去的多種蛋白質構成,包括膠原蛋白(Collagens)、纖維黏 蛋白(Fibronectin)、層粘連蛋白(Laminins)和醣蛋白(Proteoglycans), 架構成的複雜纖維網狀組織,可加強組織的強度及韌性。
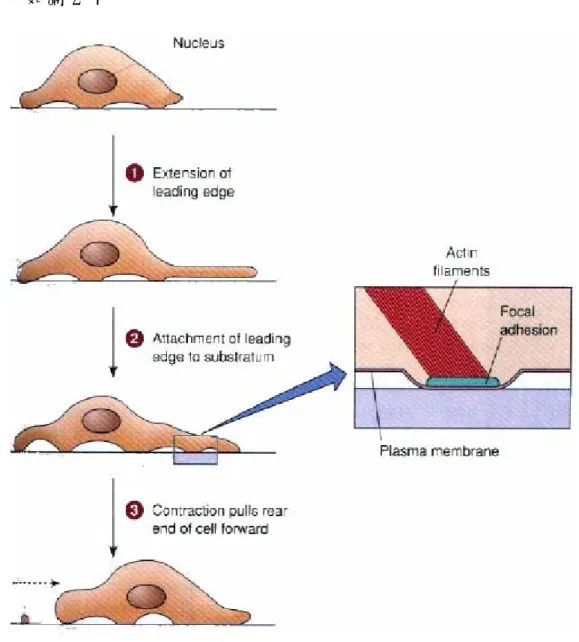
細胞運動過程是由一連串的突出(Protrusion)、黏附(Attachment) 及收縮(Retraction)等動作反覆循環,可將之分為五個主要步驟,可參 考圖 2-6: 1. Membrane Extension: 細胞的遷移是由極化現象開始,極化現象即為定位出細胞的前後以及 移動方向。一旦細胞極化後,憑藉著肌凝蛋白(Myosin)肌動蛋白(Actin) 可在細胞內移動,並於細胞前進方向之前端發生肌動蛋白聚合(Actin Polymerzation),產生肌動蛋白纖維(Actin Filament)推動細胞膜向前 突出,形成偽足(Filopodia)或瓣狀偽足(Lamellipodium),引導細胞開始 前進運動。
2. Attachment to the Substrate
細胞向前伸展後,為避免細胞縮回,故偽足必須找到新的 Cell-ECM 接 觸點並且附著上去。細胞內的受器蛋白,如黏合素(Integrins),會被送到 細胞前緣並附著於細胞外基質上,附著後因偽足之接觸面積大於尾端,因 此可提供足夠的抓力拉動整個細胞。 3. Cell Contraction 經前面的偽足固定後,細胞內的胞器將會向前流動,而造成流動之原 因主要為於細胞後面已回收的肌動蛋白纖維和細胞膜受器被送往前面,故 產生一股向前的流動。
4. Release of the Attachment at the Trailing Edge 尾端部分和 ECM 的接觸點會逐漸分離並縮回。 5. Recycling of the Receptors
最後尾端的受器會被回收到細胞內並往前推送,將繼續進行下一次的 循環。
若將整個過程濃縮,主要可分為 1.細胞膜向遷徙方向伸展 2.細胞本體 的移動,如圖 2-7。
圖 2-7 細胞遷移簡易示意圖 2-4 高分子之聚合現象 從上述章節可知生物細胞運動其實是由數種不同過程反覆循環,而聚 合現象(Polymerization)便是其中相當有趣的一環,生物細胞內的聚合 現象類似於高分子材料的聚合,由大量的簡單小分子合併成具有大分子量 的物質,而能聚合成高聚物的小分子物質可稱之為單體。 以本研究所選取的白血球細胞為例,當白血球受到細菌所分泌的化學 物質吸引影響,將啟動細胞內的遷移信號並產生聚合現象,此時在細胞內 扮演小分子的單體便為肌動蛋白單體(G-Actin),全稱為球狀肌動蛋白
(Globular Actin),可參考圖 2-8,一端為有刺端(Barbed End),另一端
為尖頭端(Pointed End)。當大量的球狀肌動蛋白一個接一個組合,可連 成一串肌動蛋白鏈(F-Actin),這種肌動蛋白多聚體可稱作纖維形肌動蛋 白(Fibrous Actin),再經由兩串肌動蛋白鏈互相纏繞則可形成一條肌動 蛋白纖維(Actin Filament),這一連串的反應便稱為高分子之聚合現象。
從相關研究已得知肌動蛋白微絲的成長速度約為每秒鐘可生長十分之一微 米(0.1μm/s)。 圖 2-8 球狀肌動蛋白(G-Actin)簡圖 2-5 偽足 偽足是生物細胞運動過程中主要的運動器官,聚合現象產生的目的即 是形成偽足。當發生肌動蛋白聚合,慢慢形成條狀的肌動蛋白纖維後,經 由 Arp 複合體(Actin Related-Protein)將數根肌動蛋白纖維結合成一束, 捆在一起猶如一圓柱,Arp 複合體是一種能與肌動蛋白結合的蛋白,由 Arp2、Arp3 和其它數種蛋白構成。而肌動蛋白纖維成長到一定長度後,封 閉蛋白(End-Blocking Protein)會像帽子般覆蓋在纖維頂端,肌動蛋白 纖維將停止組合,此時整束肌動蛋白纖維束即可朝運動方向推動細胞膜, 在細胞運動方向前端形成突出,此突出物便為偽足,故偽足內主要是由佈 滿的肌動蛋白纖維所支撐,而由鄰近數個偽足突出後將可形成瓣狀偽足, 為葉子形狀般瓣狀的突出物。
2-6 紅血球細胞介紹 紅血球,是血液中數量最多的一種血細胞。紅血球的主要功能分 子是「血紅蛋白」(紅血球的 90%由血紅蛋白組成)。血紅蛋白是一種 含有血紅素的蛋白質分子,它可以在肺部或鰓部與氧氣分子結合,然 後在身體的組織中將結合的氧氣分子釋放。氧氣分子可以很容易地以 「擴散方式」通過紅血球的細胞膜。血紅蛋白也可以運送由器官使用 氧氣后產生的二氧化碳(不到氧氣總量的 2%,更多的二氧化碳由血 漿來運輸)。另一種相關的蛋白質分子「肌紅蛋白」,可以在肌肉細胞 中存儲氧氣。此外,血紅蛋白與一氧化碳的結合活性要遠高於氧氣, 因此當空氣中存在一定量的一氧化碳時,血紅蛋白失去攜氧能力,導 致一氧化碳中毒,嚴重時可致死。 紅血球這種攜氧細胞(即將攜氧蛋白質包含在細胞中而不是直接 包含于體液中)的出現,是脊椎動物進化過程中的重要一步,它使得 血液在低黏度情況下仍具有高攜氧性。在哺乳動物中,成熟的紅血球 是無核(細胞核)的,因為細胞核會佔掉能帶氧的面積,由於缺少細 胞核,因此紅血球不能生產結構蛋白、修復蛋白或酶,使得其只有有 限的壽命。紅血球的主要作用是負責運送氧氣,所以它不會去消耗自 己攜帶的氧氣。因此,紅血球也無粒線體結構存在,它們通過糖酵解 產生能量,行無氧呼吸作用。
紅血球的形狀像是雙凹盤狀,也就是說,邊緣較厚,而中間較薄。 由於這種特別的形狀而且體積比較小(直徑約 7μm),所以表面積對體 積的比值較大,使氧氣以及二氧化碳能夠快速地滲透細胞內、外。同 時它還具有柔韌性,這使得它可以通過毛細血管,並釋放氧分子。如 圖 2-9 圖 2-9 紅血球結構圖 紅血球細胞在不同濃度溶液時,會有不同的現象產生(參考圖 2-10),以下介紹三種濃度的溶液,:
(A). 等張溶液(isotonic solution):水分子進出細胞膜的速率相同, 所以水分子淨移動為零。
(濃度為 0.9%NaCl 的靜脈注射為等張溶液)
(B). 低張溶液(hypotonic solution):細胞內溶質的濃度大,所以水 分子不斷的進入細胞內,會使細胞膨脹,甚至脹破。
(C). 高張溶液(hypertonic solution):細胞內溶質的濃度小,所以水 分子不斷的離開細胞內,水分子的流失,導致細胞的皺縮。 (濃度>0.9%NaCl 的靜脈注射為高張溶液)
2-7 細胞與細胞間及細胞與基質間黏附關係 (Cell-Cell and Cell-Matrix Adhesion)
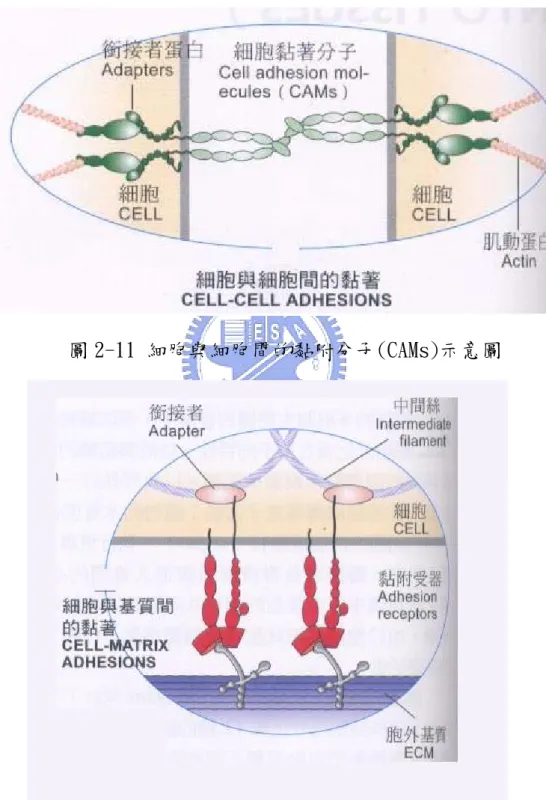
在複雜的多細胞生物體的發展中,例如動物,前驅細胞分化成各 自不同特性、功能與結構型態的細胞。像類似的細胞時常聚集成組 織,以共同執行一個功能,類如肌肉收縮、神經組織傳導電子脈動... 等。不同組織又能組成器官(organ),執行更專一的功能,例如肌肉、 瓣膜與負責擠壓血液進入身體的心臟工作的血管。在組織中許多類型 的細胞共同協調功能組成多樣性之特化組織,由於他們的彼此配合使 個體移動、代謝、生殖與其他的基礎活動。 在細胞的觀點上,不同組織聚集而成的器官,可以用分子間的交 互作用來觀看。組織中的細胞可以直接經由特化的貫穿性膜蛋白,即 被稱為「細胞黏附分子」(cell adhesion molecules;CAMs)來彼此相 互黏附(如圖 2-11)。細胞黏附分子常成叢的特化成細胞連接(cell junction)。在動物組織中的細胞也可經由與環繞細胞表面膜的「細 胞外基質」(extracellular matrix;ECM)上的黏著受器鍵結,以達到 間接黏附的作用(如圖 2-12)。細胞外基質是由細胞分泌多糖類與蛋 白質進細胞間,交叉組合成的網狀複合物。兩種基本的交互作用型態 不只讓細胞聚集成不同的組織;而且也提供了細胞內外雙向傳的管 道。
許多的黏附分子是由多個不同的單位拼湊而成,而且可從其中發 現,有很多都有超過一種以上的細胞黏附分子。一個細胞的細胞黏附 分子可以直接與鄰近的同種細胞的細胞黏附分子相鍵結(嗜同種鍵 結,homophilic binding),或與不同種細胞黏附分子相鍵結(嗜異種 鍵結,heterphilic binding)。細胞與細胞間的黏著可以是緊密而長 時間持久的;也可以是弱而短暫的。例如,脊髓神經細胞間的聯繫或 肝臟的新陳代謝細胞表現緊密連接,相反地,血液中的免疫細胞可以 呈現弱而短暫的交互作用,使其可以沿著血管滾動或在組織內與感染 的病菌相打鬥時,穿過細胞壁。 一定的細胞表面受器包含一些整合蛋白,他們可以鍵結到細胞外 基質(extracellular matrix;ECM)的組成物上,因此經由與基質的交 互作用來達到彼此之間的黏附。雖然細胞外的基質通常提供組織機械 性的支撐,也可提供其他幾種功能。為了不同的目得,細胞外基質的 成分會形成不同的組合方式。例如,軟骨墊與大部分組織的黏附;骨 骼、牙齒與筋的加強。除此之外,基質組成會根據組織在不同的生理 狀態而有差異,他也能讓一個細胞知道他在何處與他需要做什麼。細 胞外基質成份的改變,包括不斷地重新改形、分解與局部性的重新合 成,可以緩和細胞與環境之間的互動關係。基質也可當作許多細胞外 用來控制細胞生長與分化的訊息傳遞分子的儲存庫。除此之外,特別
在細胞聚集的早期,基質提供細胞移動的路徑。型態發生在胚胎發展 的晚期,組織、器官與身體部分的形成是靠細胞的移動與重組,也就 是依賴細胞與基質間的黏附和細胞與細胞間的黏附。
圖 2-11 細胞與細胞間的黏附分子(CAMs)示意圖
第三章 研究架構和方法及工具介紹 3-1 研究方向 隨著新科技的不斷進步,生物細胞內的秘密不斷被挖掘出來,而 在工程科學領域中,對於生物細胞的運動及其所受之應力影響亦期待 有更全面的研究及發展,於本研究中將以機械工程科學的觀點及理 論,藉由工程軟體的應用,針對(1) 紅血球黏附力所引起的細胞膜張 力(2)白血球的伸展行為及偽足突出,做相關的力學模型建立和細胞 膜表面張力模擬作研究。期待經由力學模型,讓我們對生物細胞的幾 何形狀和結構之行為變化與其所產生的應力和位移之間關係有更深 入的了解。 3-2 研究工具簡介 現今電腦科技急速的發展,不但對人類生活帶來許多幫助及貢 獻,對科學的研究更是提供了莫大的幫助,許多工程問題皆可藉由電 腦的輔助,獲得快速的解答和大量的資料,故電腦於工程科學所扮演 的角色日趨重要且不可或缺。 本研究藉由程式語言軟體—MATLAB 建立模型所需的網格節點及 元素資料,且利用 CAE(Computer Aided Engineering)軟體—ANSYS 建立模型並進行力學分析,並且利用 amira 軟體計算模型的體積和面 積,在下章節將對這三個工程軟體做簡單介紹。
3-2-1 ANSYS 簡介
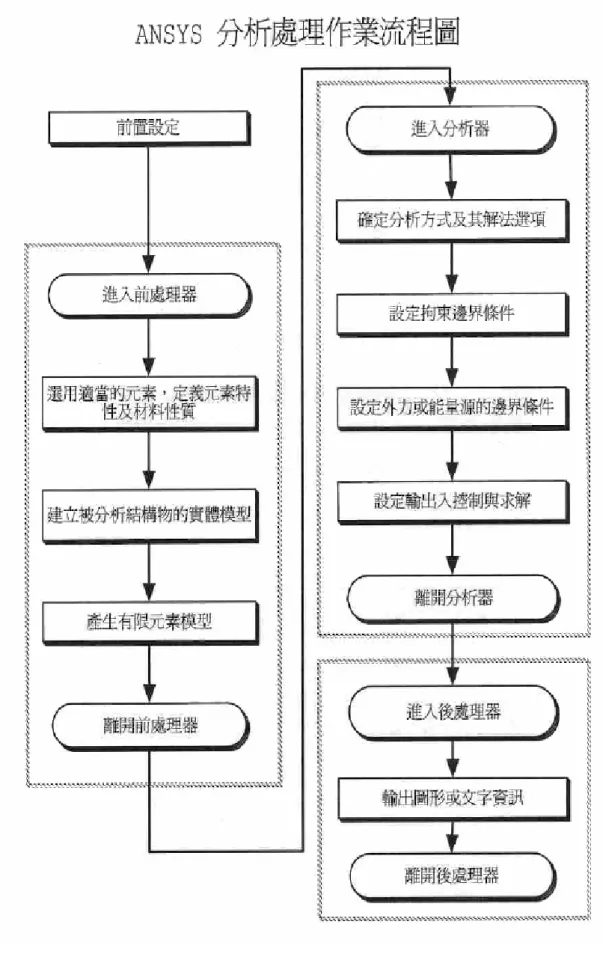
本研究所採用的 ANSYS 是一套功能強大的 CAE 軟體,利用有限元 素法(Finite Element Method)的觀念,是目前在工業上最常使用 的電腦輔助分析套裝軟體之一。它功能相當強大,應用的範圍亦相當 廣泛,能同時分析結構物受到靜力、動力、熱傳、電磁、流力等。當 結構物的模型在軟體中被建立並被網格化後,只要附加上所需的條 件,就可直接利用有限元素法來計算,進而了解到所建立的模型或的 力學特性。 ANSYS 的分析步驟主要可分為前處理(Pre-Processing)、求解 (Solver)及後處理(Post-Processing)這三大部分,操作流程可 參考圖 3-2,將其步驟簡述如下: (1) 前處理(Pre-Processing): 進 行 分 析 的 首 要 步 驟 , 主 要 為 設 定 模 型 所 使 用 的 元 素 類 型 (Element Type)。元素的選用是屬於分析開始的第一項作業,它是 最簡單也是最難的動作,當元素選用完畢,就已經決定模型要如何簡 化與建構方式,同時也決定了求解與邊界條件的可能方式,因此在元 素的選用上要非常的謹慎小心。當元素選擇完畢後,元素的特性會因 為元素的種類不一樣而有不同的意義,例如薄殼元素的特性代表元素 的厚度資料,而梁元素的特性代表元素的截面積和慣性矩等資料。當
元素的特性設定完成後,緊接著就要考量材料性質等參數,再來是幾 何模型的建立(Modeling)和網格分割(Meshing)亦屬於前處理。 所謂網格分割便是根據建立的幾何模型來分割網格,並可針對模型幾 何形狀的變化或邊界條件的施予,設定網格的疏密程度以提升計算效 率並降低解的誤差值。 (2) 求解器(Solver): 當進入分析器後首先必須要先確定到底是要使用靜態方程式或 動態方程式,或是以何種方式來求解動態方程式,例如求解振動模態 或暫態分析。在 ANSYS 中預設為進行靜態分析。當決定了解題方式 後,有時還會有一些相關選項也要設定,如此才算完成分析器中第一 階段的任務。 根據分析目標施予模型邊界條件(Boundary Conditions),並選 擇適當的分析方法類別(Analysis Type)來進行計算和求解。因邊 界條件的設定及分析方法類別的選擇,將直接影響解的正確性,故必 須謹慎考慮目標物的真實狀況來給予正確的設定,避免分析結果錯 誤。 而本論文在使用 ANSYS 求解分析時是利用到全牛頓-拉夫森 (Full Newton-Raphson Method)疊代法則,介紹如下:
[Kt ]{Du}= {Fa }− {Fnr
[KT ]:切線剛性矩陣(Tangent Stiffness Matrix) {Du}:增量位移向量(Displacement Increment) {Fa }:外力向量(Applied Load Vector)
{Fnr}:內力向量(Internal Force Vector)
假設在系統中給定一外力{Fa},而系統即可由節點(Node)來求 解出內力{Fnr}的大小,利用力平衡{Fa}− {Fnr }= 0 的規則來判斷是否 為收斂。此一差值稱為Newton-Raphson Residual(以下簡稱NRR), 而在一般的問題中NRR 不一定為剛好為零,系統是判斷NRR 值小於一 個容許誤差值,來作為是否收斂的依據。其簡意圖如圖 3-1 所示。 假設系統在處其切線剛性矩陣u0 處其切線剛性矩陣[Kt ]為已知,則 位移可由及Δu可由 ΔF及[Kt]求出,而後位移就移至u1。如此增加外 力向量之值,逐次計算,直到實際施加外力之值。
(3) 後處理器(Post-processor) 後處理器最主要的任務乃是將分析的結果以圖形或文字的型式 表現出來,以作為判斷分析之用。在文字輸出的資訊方面,包含有節 點及元素的變形量、應力及應變等資料。對構造簡單的結構物而言, 所提供的文字資訊或許就已足夠;但對複雜的結構物而言,龐大的文 字資訊反而造成使用者無所適從的感覺,所以往往需要由圖形資訊 中,利用顏色深淺變化的情形以了解結構物的應力應變分佈情形。 後處理器除了輸出結果外,另一個重要的任務是驗證結果。驗證 的第一項工作是檢核其是否有違反原先在前處理器中設定的假設。首 先從最大應力值來判斷其值是否大於降伏強度,若是大於降伏強度, 則結構已經違反材料線性的假設,因此必須要重新處理。接下來要判 斷結構的位移量是否過大而產生幾何非線性,若是產生此種結果,則 必須要重新計算。另外在後處理器中也要觀察應力梯度的變化,如此 才能判斷結構網格的密度是否足夠。總之,後處理器最重要的角色在 驗證結果,使用者必須要謹慎,才能得到正確的結果。
3-2-2 MATLAB 簡介
MATLAB 此名稱是由矩陣實驗室(Matrix Laboratory)所合成, 因此可知其發展理念是提供一套非常完善的矩陣運算指令,但隨著數 值運算需求的演變,MATLAB 已成為各種系統模擬、數位訊號處理、 科學目視的標準程式語言。 MATLAB 是一個計算核心,圍繞這個計算核心,有許多對不同應 用所開發的應用程式,稱之為工具箱(Toolboxes),因為這些工具箱 的開發,也使本軟體更便於使用、操作。 3-2-3 Amira 軟體簡介 amira 對於三維資料的處理如生物學與工程學等提供了極好的解 決方案。藉由視覺化的方式讓使用者更能洞察資料的細節。而且由硬 體加速繪圖的功能也讓使用者與電腦以互動方式傳輸大量資料更有 效率。強大的自動化與交互式的分割工具支援 3D 影像資料的處理。 創新性、快速性和穩定性的重建模型演算法使得以片段的物體去建立 多邊形的模型更為容易。此外,還可對先進的有限元素法模擬產生真 實容量的四面體網格。在大量的視覺化方法的調查下,模擬結果與其 他許多不同的格點定義的資料在視覺化的表現是一樣的好。
3-3 研究方法
本論文利用 CAE 軟體—ANSYS 並加以程式語言軟體—MATLAB 和 Amira 軟體的輔助,建立各種模型,對紅血球伸展行為和白血球的伸 展行為及偽足突出行為做力學分析,相關的模擬方法參考以下說明: 1. 紅血球伸展行為模擬方法: 假設細胞膜和平板一開始為一點接觸的型式,固定兩者間的 接觸點及底部平板,並在細胞膜上施加壓力ΔP1,使細胞膜微量向 外膨脹,並接觸到底部平板。當細胞膜接觸到平板時,立即將接 觸部份固定在平板上,然後再施加壓力ΔP2,使細胞膜再次微量 向外膨脹接觸到底部平板,並固定新的接觸部份(參考圖 3-3)。 依此類推,直到伸展行為達到相關參考文獻[1][2]的細胞幾何形狀 時,觀察楊氏係數、細胞膜張力之間的關係。(模擬過程需保持 紅血球體積固定)
圖 3-3 細胞伸展行為示意簡圖(L1、L2 為底部接觸長度) 2. 白血球伸展行為和偽足突出模擬方法 (a) 白血球的伸展行為大致和紅血球模擬過程一樣,不同的地方 在於施加壓力的起始值。因為由相關文獻[17]指出白血球細胞在常 態下,表面張力會介於 0.027~0.035 m mN 之間,因此,需要先求 得初始內壓力 ,並計算此時的體積 ,後續的模擬過程皆保持 在V 。然後,再內壓力 的情況之下,再施加一 0 P V0 0 P0 ΔP1,使細胞膜微 量向外膨脹接觸到底部平板。 (b) 白血球的偽足突出模擬,由相關文獻[29]指出,生物細胞運動 時所形成之偽足,其內部需 10 至 30 根肌動蛋白纖維捆在一起以 對抗細胞膜阻力,本論文採用十二根纖維結合的肌動蛋白纖維束
構推動細胞膜,其直徑為七個肌動蛋白纖維直徑距離,即 49 (如圖 3-5)。 nm 圖 3-4 (a)十二根微絲之肌動蛋白微絲束幾何結構排列圖 (b)其排列截面圖 圖 3-5 肌動蛋白微絲束推動細胞膜之簡圖
第四章 紅血球黏附力所引起的細胞膜張力 紅血球是人體內重要的細胞之一,負責輸送氧氣和養份到人體各 器官。紅血球能夠攜帶的氧氣量和體積有關系,在考量最多的氧氣攜 帶及自身體積膨脹但又不造成細胞膜破裂的前提之下,達到最佳的生 物行為。由相關文獻[1][2]得知,紅血球的細胞膜厚度為 1.5~2.5 , 楊氏係數介於E=0.73 ~ 30 MPa之間,本論文在這個範圍內,探討紅 血球的楊氏系數及細胞膜表面張力之間的關係。 nm m 本論文以直徑 6μ ,厚度 2 的圓球薄殼來模擬紅血球在平板上 的伸展行為,假設兩者一開始是以一點接觸的型式。 nm
ANSYS 的一般靜力分析(Static Analysis)標準流程為(1)建
立模型,(2)設定分析類別及方法,(3)輸入邊界條件,(4)求解並 觀看結果,詳細步驟紀錄如下: (1) 建立模型:選擇 shell 63 彈性薄殼元素模擬紅血球細胞 膜並建立直徑 6μm的圓球薄殼,設定紅血球細胞膜厚度為 2nm, E= 15 MPa。再以 plane 42 元素來模擬簡化成 2-D 平面的平板。(參考圖 4-1)。 (2) 設定分析類別及方法:此為簡單的靜力分析問題,故分析 類別選擇為 Static。 (3) 輸入邊界條件:固定底部平板和兩者間的接觸點(參考圖
4-2),並且在平板和薄殼下半圓面上建立「接觸元素對」 (參考圖 4-3)。「接觸元素對」的功用為當薄殼和平板發 生接觸時,即將接觸部位固定在平板上,不會發生細胞穿 透平板的非正常現象,以符合實際物理情況。在薄殼面上 施加「動態壓力負載」, 「動態壓力負載」的功用為在第 1 個時間區內系統施加ΔP1對模型造成影響,然後在第 2 個 時間區內系統再施加ΔP2持續對模型造成影響,以此類推。 (4) 求解並觀看結果:啟動求解器獲得解,並可利用圖形顯示 其 變 形 程 度 , 如 圖 4-4(a)~(h) , 表 面 張 力 如 圖 4-5(a)~(h)。 隨著細胞膜接觸到底部平板的面積漸漸增加,而自由面的細胞膜 漸漸的被拉緊造成表面張力的增加。持續模擬直到細胞模型高度 Y 2.5≈ μm高時,觀察細胞膜各階段變形程度和細胞膜的表面張力,並 將其資料整理如表 4-1。 將細胞模型Y≈2.5μm高 (圖 4-5 (h) ) 時的細胞膜表面張力和參 考值作比較。如果表面張力小於或大於參考值( m mN 1 ~ ),則重新調整 E值,在相同的邊界條件下重新分析,直到表面張力趨近於參考值 ( m mN 1 ~ )。經過數次調整E值分析後,將不同E值所得到的Y 2.5≈ μm高 時之表面張力圖圖示,見圖 4-6 ~ 4-13,並將其結果整理如表 4-2。
由表 4-2 中,可看出當紅血球楊氏系數E=12.88 MPa時,細胞膜表面 張力會趨近於參考值( m mN 1 ~ )。 m μ ,厚度 2nm) 圖 4-1 紅血球模型圖(直徑 6 圖 4-2 模型拘束邊界示意圖
(a) Da=2.6 mμ ,Y=5.63 mμ (b) Da=3.4 mμ ,Y=5.42 mμ
(c) Da=4.2 mμ ,Y=5.20 mμ (d) Da=5.6 mμ ,Y=4.62 mμ
(e) Da=6.9 mμ ,Y=4.11 mμ (f) Da=7.8 mμ ,Y=3.53 mμ
(g) Da=8.2 mμ ,Y=3.1 mμ (h) Da=8.8 mμ ,Y=2.52 mμ 圖 4-4 (a)~(h)細胞膜各階段變形程度 (E=15MPa)
(a) Tmax =0.107mN m (b) Tmax =0.144mN m
(c) Tmax =0.153mN m (d) Tmax =0.213mN m
(e) Tmax =0.267mN m (f) Tmax =0.307mN m
(g) Tmax =0.376mN m (h) Tmax =0.458mN m
圖 4-6 E=15 MPa 時, m mN TMAX =0.458 圖 4-7 E=7.5 MPa 時, m mN TMAX =2.212
圖 4-8 E=11.5 MPa 時, m mN TMAX =1.405 圖 4-9 E=13.5 MPa 時, m mN TMAX =0.819
圖 4-10 E=12.5 MPa 時, m mN TMAX =1.254 圖 4-11 E=13 MPa 時, m mN TMAX =0.981
圖 4-12 E=12.75 MPa 時, m mN TMAX =1.092 圖 4-13 E=12.88 MPa 時, m mN TMAX =1.004
表 4-1 細胞各階段變形資料 (E=15 MPa ) E=15 Mpa 編號 總內壓力 (pN μm2) 底部接觸長度 Da (μm) 頂點高度 Y (μm) Tmax (mN m) ( a ) 25 2.6 5.63 0.107 ( b ) 33 3.4 5.42 0.144 ( c ) 40 4.2 5.2 0.153 ( d ) 54 5.6 4.62 0.213 ( e ) 66 6.9 4.11 0.267 ( f ) 75 7.8 3.53 0.307 ( g ) 79 8.2 3.11 0.376 ( h ) 85 8.8 2.52 0.458 表 4-2 不同 E 值所對應的 Tmax 值 case 楊氏係數 E (Mpa) Tmax (mN m)
1 15 0.458 2 7.5 2.212 3 11.5 1.405 4 13.5 0.819 5 12.5 1.254 6 13 0.981 7 12.75 1.092 8 12.88 1.004 (參考值 Tmax= ~1 m mN )
第五章 白血球細胞伸展行為和偽足突出之模擬分析 白血球是血液中相當重要的血細胞,與紅血球及血小板合稱三大 血細胞,其幾何形狀近似球形,直徑約為 10μm。本章模擬重點為細 胞在平板上的伸展運動行為,同時觀察伸展行為對細胞表面的細胞膜 張力之影響,接著在伸展行為到一定程度後,模擬細胞偽足突出行 為。本論文的模擬過程還有另一個重點,模擬伸展過程中,細胞的體 積皆要保持在體積V0,求得V0的方式在後面 5-1(a)中介紹。 5-1 細胞伸展行為模擬流程 (a) 求得體積 的方法 由相關文獻 0 V [17] 指出,細胞在沒有受到外力作用之下,只 憑 本 身 的 內 壓 力 會 使 得 細 胞 膜 表 面 張 力 為 T=0.027~0.035 m mN 。根據以上的資料,本論文取平均值 T=0.03 m mN 。 一開始先建立直徑 10 的圓球薄殼模型,選擇shell 63 彈性薄殼元素,薄膜厚度為 3.5 ,E=1 MPa,並固定模型 底部一點且在圓球內部施加一均勻壓力P(參考圖 5-1)。求 解分析後,當壓力P使得細胞表面的細胞膜達到 0.03 nm m mN 張 力值,此時的壓力P稱為細胞初始內壓力 ,並且利用Amira 軟體計算其體積(V )和面積( )。 0 P 0 A0
經過模擬分析後當 0 12 2 um pN P = 時,細胞膜表面張力達到 0.03 m mN (如圖 5-2 ),此時的體積 和面積 (如圖 5-3 )。後續的細胞伸展模擬過程皆要 保持在細胞體積( )。 3 0 532.437 m V = μ 2 0 317.744 m A = μ 0 V 圖 5-1 細胞模型(直徑 10μm,厚度 3.5nm) 圖 5-2 內壓力 2 0 12pN um P = 時之表面張力
圖 5-3 利用 Amira 軟體計算體積V0和面積A0
(b) 細胞伸展模擬流程:
ANSYS 的一般靜力分析(Static Analysis)標準流程 為(1)建立模型,(2)設定分析類別及方法,(3)輸入邊 界條件,(4)求解並觀看結果,詳細步驟紀錄如下: (1)建立模型:選擇 shell 63 彈性薄殼元素,建立直徑 10μm 的圓球薄殼,設定細胞膜厚度為 3.5nm, E= 1 MPa。再 以 plane 42 元素來模擬簡化成 2-D 平面的平板(如圖 5-4)。 (2)設定分析類別及方法:此為簡單的靜力分析問題,故僅 需將分析類別選擇為 Static。 (3)輸入邊界條件:細胞和載體一開始是點接觸的清況下作 模擬分析的第一步。固定底部平板和兩者之間的接觸點(如
圖 5-5),並且在圓球和平板之間建立「接觸元素對」(參考 圖 5-6)。建立「接觸元素對」的用意為當細胞接觸到平板 時,即固定在平板之上,不會發生細胞穿透平板的非正常現 象,以符合實際物理情況。在薄殼面上施加「動態壓力負 載」,「動態壓力負載」的功用為在第 1 個時間區內 0 12 2 m pN P μ = 對模型造成影響,然後在第 2 個時間區內系統再施加 持 續對模型造成影響,依此類推。直到細胞接觸到平板的面積 足夠大,以提供偽足向外突出之基礎。 (4)求解並觀看結果:啟動求解器獲得解,並可利用圖形顯 示其變形程度。各階段變形程度如圖 5-7(a)~(f),和各階段 表面張力情況如圖 5-8(a)~(f)。 觀察各階段的變形程度、圓球薄殼模型和平板的接觸長 度及表面張力,並將其資料整理如表 5-1。由表中可觀察得 知, 伸展行為會引起細胞膜表面積增加導致細胞膜張力跟 著增加。 1 P Δ
圖 5-4 細胞和平板模型示意圖
圖 5-5 模型拘束邊界示意圖
m m m m μ , Y=9.87μ (b) Da=6.6μ , Y=9.36μ (a) Da=3.1 m m m m μ , Y=8.55μ (d) Da=9.51μ , Y=8.13μ (c) Da=8.8 m m m m
μ , Y=7.67μ (f) Da=12.2μ , Y=7.04μ (e) Da=10.52
圖 5-7 (a)~(f) 細胞膜各階段變形程度
(a) TMAX =0.0482mN m (b) TMAX =0.0528mN m
(c) TMAX =0.0618mN m (d) TMAX =0.0696mN m
(e) TMAX =0.102mN m (f) TMAX =0.121mN m 圖 5-7 (a)~(f) 細胞膜各階段表面張力值 表 5-1 細胞各階段變形資料 編號 總內壓力 (pN/μm2) 底部接觸長度 Da ( mμ ) 頂點高度 Y ( mμ ) Tmax (mN/m) ( a ) 15.5 3.1 9.87 0.0482 ( b ) 19.7 6.6 9.36 0.0528 ( c ) 24.8 8.8 8.55 0.0618 ( d ) 28.7 9.58 8.13 0.0696 ( e ) 34.6 10.52 7.67 0.102
5-2 細胞在平板上的偽足突出行為模擬 細胞在平板上經過伸展過程後,細胞和平板之間的接觸面積達到 一定程度,足夠提供細胞偽足向外突出的基礎。本節將利用 5-1 節中 步驟五的細胞模型,將偽足突出的地方劃分更密的網格,接續做細胞 偽 足 突 出 行 為 模 擬 ( 如 圖 5-8) 。 此 時 細 胞 的 表 面 已 經 有 張 力 (TMAX =0.102mN m nm nm )的存在,偽足突出需要更多的力量來抵抗表面張力 以便達到向外伸出的目的。 5-2-1 單根偽足之模擬 由相關文獻[29]指出,偽足主要由近似圓柱狀的肌動蛋白纖維束向 前推動細胞膜所形成的突出,生物細胞運動時所形成之偽足,其內部 需 10 至 30 根肌動蛋白纖維捆在一起以對抗細胞膜阻力。本論文採用 12 根肌動蛋白纖維,其幾何結構排列(參考圖 5-9),將之視為一圓柱 結構推動細胞膜,其直徑為七個肌動蛋白纖維直徑距離,即 49 。 欲探討偽足突出行為,需要更密的網格才能得到精確的解。因 此,將模型網格加密以提供足夠的節點數密度來施加偽足向外突出的 機械力。網格中直徑為 49 的近似圓,正為肌動蛋白纖維束推動細 胞膜時,與之接觸的位置,可(參考圖 5-10),故可將圓上分佈之節 點設為負載施予之位置,固定細胞和平板接觸部份(參考圖 5-11)。 啟動求解器獲得解,並可利用圖形顯示其變形程度,如圖 5-12,亦
可列出詳細變形位移資料。 隨施加負載的變化,細胞膜亦產生不同程度變形,如圖 5-13,將 其資料整理可得表 5-2 及圖 5-14,觀察圖 5-14 可得知偽足突出長度 會 隨 施 加 負 載 變 大 而 增 長 , 且 其 關 係 近 似 正 比 的 線 性 關 係 。 圖 5-8 細胞模型圖(偽足突出部位之網格加密) 圖 5-9 (a)十二根微絲之肌動蛋白微絲束幾何結構排列圖
圖 5-10 偽足和細胞膜接觸圓直徑
nm
圖 5-11 邊界條件設定:固定細胞膜和平板接觸部份,並在直徑 49 圓上施加負載
圖 5-12 細胞偽足突出變形圖
0 0.5 1 1.5 2 2.5 3 8 10 12 14 16 18 20 F (pN) 偽足突出長度 (u m ) 圖 5-14 施加負載 F 與偽足突出長度之關系圖 表 5-2 施加負載對細胞膜突出長度之影響 單節點所施加之負載 (pN) 負載總和 ( pN) 細胞膜突出之長度 (μm) 1.0000 8 0.98 1.2500 10 1.23 1.5000 12 1.48 1.7500 14 1.72 2.0000 16 1.97 2.2500 18 2.21 2.5000 20 2.46
第六章 結論 6-1 結果比較 (1) 在第四章中,本論文的細胞模型高度從 6μm降到 2.5μm,且底 部接觸長度~8.8μm,與相關文獻[1][2]的細胞模型相近。相關文獻[1][2]之 細胞在如此形狀之下的細胞膜張力~1mN m,但量測所得之紅血球楊 氏係數為 0.73~30MPa,此屬相當大的範圍。本論文利用數值模擬的 方式,在 0.73~30MPa之內反複調整E值,經過數次模擬後,E=12.88 MPa 時,細胞膜張力~1mN m(參考圖 4-13)。將紅血球楊氏係數以更精準 的結果呈現。 (2) 在第五章中,本論文模擬之單根偽足突出長度和力量間的關係, 和相關文獻[29]作比較。由圖 6-1 中可看出在相同力量負載之下,偽足 突出長度會比相關文獻[29]短。此結果印證了,細胞膜張力的存在會使 得偽足更難突出,要達到相同長度,則需要更多根肌動蛋白纖維束提 供力量,同時防止肌動蛋白纖維束發生挫曲現象。
0 1 2 3 4 8 10 12 14 16 18 20 F (pN) 偽足突 出長 度 ( um ) 圖 6-1 偽足突出長度和負載關係 :本論文資料,細胞膜表面拉緊時,負載和偽足間的關係。 :相關文獻[29]資料,細胞膜表面未拉緊時,負載和偽足間的關係。 6-2 未來展望 受限於分析軟體功能,未建立細胞質,且細胞質模擬牽扯到流場 分析、細胞質的黏滯性、細胞質的流動方式…等,待未來分析軟體具 備更齊全的功能,可同時考慮多種清況下的模型分析,便可以建立更 接近真實細胞的模型。本論文之些許成果亦表示了工程科學理論於生 物醫學方面還有許多發展空間,本論文之成果亦提供了細胞模擬研究 之雛型,期待兩大領域的緊密結合研究發展。
參考文獻
1. R. Skalak, A. Tozeren, R. P. Zarda and S. Chien.「Strain Energy Function of Red Blood Cell Membranes」.Biophysical Journal 13: 245-264 (1973)
2. Alina Hategan, Richard Law, Samuel Kahn and Dennis E. Discher.「Adhesively-Tensed Cell Membranes: Lysis Kinetics and Atomic Force Microscopy Probing」.Biophysical Journal 85:2746-2759 (2003)
3. Tsai, M. A., R. S. Frank, and R. E. Waugh. 「 Passive mechanical behavior of human neutrophils: effect of cytochalasin B」. Biophys. J. 65:2078 –2088. 1994.
4. D. A. Simson, F. Ziemann, M. Strigl, and R. Merkel. 「 Micropipet-Based Pico Force Transducer: In Depth Analysis and Experimental Verification 」 . Biophysical Journal Volume 74 April 1998 2080–2088
5. Jianwu Dai and Michael P. Sheetz. 「 Membrane Tether Formation from Blebbing Cells」. Biophysical Journal Volume 77 December 1999 3363–3370
Red Cell Membrane 」Biophysical Journal 4: 115-135 (1964) 7. 「 Changes in the mechanical properties of fibroblasts during spreading:a micromanipulation study」Biophys J (1999) 28: 222–234
8. Tomomi Tani, Richard D. Allen and Yutaka Naitoh.「Cellular membranes that undergo cyclic changes in tension: Direct measurement of force generation by an in vitro contractile vacuole of Paramecium multimicronucleatum」.Journal of Cell Science 114, 785-795
9. R. M. Hochmuth, N. Mohandas, and P. L. Blackshear, Jr. 「Measurement of the Elastic Modulus for Red Cell Membrane Using a Fluid Mechanical Technique」Biophys J. 1973 August; 13(8): 747–762.
10. Pierre Sens and Matthew S. Turner. 「 Budded membrane microdomains as regulators for cellular tension」
11. WIELAND FRICKE , MICHAELC. JARVIS AND CHRISTOPHER T. BRETT 「 Turgor pressure, membrane tension and the control of exocytosis in higher plants」.
cell motility by membrane tension」. Trends in cell Biology 6,85-89.
13. Tatsuru Togo,Tatiana B. Krasieva, and Richard A. Steinhardt 「 A Decrease in Membrane Tension Precedes Successful Cell-Membrane Repair」. Molecular Biology of the Cell Vol. 11, 4339–4346, December 2000.
14. R.A., Bi, G.-Q., and Alderton, J.M. (1994). 「Cell membrane resealing by a vesicular mechanism similar to neurotransmitter release」. Science 263, 390–393
15. D., and Sheetz, M.P. (2000). 「 Cell spreading and lamellipodial extension rate is regulated by membrane tension」. J. Cell Biol. 148, 127–136.
16. Dai, J., and Sheetz, M.P. (1998). 「Cell membrane mechanics Methods」 Cell Biol. 55, 157–171.
17. 「Role of the Membrane Cortex in Neutrophil Deformation in Small Pipets」, Biophysical journal Volume 67.
18. 「Quantification and localization of phosphorylated myosin I isoforms in Acanthamoeba castellanii.」 J. Cell Biol. 130, 591-603.
19. 「Mechanics of neutrophil phagocytosis: behavior of the cortical tension」, journal of cell science.
20. A. Mogilner, B. Rubinstein, 「 The physics of filopodial protrusion」, Biophysical Journal, Volume 89, pp.782-795, August 2005.
21. Alexander Mogilner, George Oster,「Cell motility driven by actin polymerization」, Biophysical Journal,1996.
22. J.G. McGarry, P.J. Prendergast, 「 A three-dimensional finite element model of an adherent eukaryotic cell 」 , European Cells and Materials, Vol.7, pp.27-34, 2004. 23. J.C.Shillcock, R.Lipowsky, 「 Shape fluctuations and
elastic properties of two-component bilayer membranes", Europhy.」 Lett.,69(4), pp.650-656, 2005.
24. Joseph E. Shigley,Charles R. Mischke, 「 Mechanical Engineering Design」, six edition, McGraw-Hill, New York, 2001.
25. Thomas D. Pollard, William C. Earnshaw, Cell biology, Philadelphia, 2002.
27. 龔皇光,黃柏文,陳鴻雄, ANSYS 與電腦輔助工程分析, 初版, 全 華, 台北市, 2002。 28. 林欣暐, 「狄勞尼三角化之有限單元網格自動建立」, 國立交通 大學, 碩士論文, 民國八十七年。 29. 顏志涵,「生物細胞突出行為之有限單元分析」,國立交通大學, 碩士論文,民國九十六年。





![圖 3-3 細胞伸展行為示意簡圖(L1、L2 為底部接觸長度) 2. 白血球伸展行為和偽足突出模擬方法 (a) 白血球的伸展行為大致和紅血球模擬過程一樣,不同的地方 在於施加壓力的起始值。因為由相關文獻 [17] 指出白血球細胞在常 態下,表面張力會介於 0.027~0.035 m mN 之間,因此,需要先求 得初始內壓力 ,並計算此時的體積 ,後續的模擬過程皆保持 在 V 。然後,再內壓力 的情況之下,再施加一P0V0 0 P 0 Δ P 1 ,使細胞膜微 量向外膨脹接觸到底部平板。 (b) 白](https://thumb-ap.123doks.com/thumbv2/9libinfo/8636520.192877/38.892.133.746.116.420/白血球白血球白血球細得初始內壓力並計算此時的體積後續皆保持.webp)