代謝
(Metabolism)
• Metabolism, the sum of all of the enzyme-catalyzed reactions in a living organisms, is
highly coordinated and
purposeful cell activity.
• 註 : 代謝途徑中的關鍵酵素,都受到協調
式 (coordinated manner) 的活化 ( activation )或抑制
代謝的功能
(Four
functions)
• (1)to obtain chemical energy by capturing solar energy or by
degrading energy-rich nutrients from the environment. 取得太陽能或 從營養物取得化學能。
• (2) to convert nutrients
molecules into the cell’s own characteristic molecules,
macromolecular precursor. 把營養 物分子轉成自已的分子。
• (3) to polymerize monomeric
precursors into proteins, nucleic acids, lipids, polysaccharides
and other cell components. 聚合單體 成細胞組成的巨分子。
• (4) to synthesize and degrade biomolecules required in
specialized cellular functions. 合成分解生物分子因應細胞之功能與需要。
• Metabolic pathway are promoted by
sequential enzyme systems. Enzymes
are the simplest units of metabolic activity, each catalyzing a
specific chemical reaction.
• 代謝途徑是由一個接連一個的酵素作用所組成的 ,酵素是代謝活性中最簡單的單位,每個酵素催 化一個特異的化學反應。大部分的代謝過程是循 途徑 (pathway) 方式進行。
Catabolism (degradation)
• Catabolism: Organic nutrient molecules are degraded by stepwise reactions
into smaller, simpler end products. • Catabolism 過程常伴隨自由能釋出,但大部分的
自由能是以 ATP ( energy-carrying
molecule )或以高能的電子即 NADH or FADH2
(energy-rich hydrogen atom) 形式儲
存。 NADH 與 FADH2 進入呼吸鏈 (respiratory
Anabolism (biosynthesis)
• Anabolism (biosynthesis) :
Small precursors or building block molecules are built up into large macromolecular
components of cells.
• Anobolism 過程需輸入自由能包括 ATP 分解成 phosphate 與 ADP 或 AMP 並需供 給高能的電子 (NADPH) 。
Amphibolism
• Amphibolism : It serves in both catabolic and anabolic processes.
• The citric acid cycle is an amphibolic pathway. It not only functions in the
oxidative catabolism of carbohydrate, fatty acids, and amino acids, but also provides precursors for many biosynthetic
Catabolic pathways
converge to few end
products
• 例如 : 主要產能的營養物其 Catabolism 可分 為三個時期 : • 第一時期 : 多糖體 (polysaccharides) 、脂 質 (fats) 與各種蛋白質 (proteins) 分解成種 類不多的建材分子 (building-block molecule) 如單糖、脂肪酸、甘油與各種胺基酸 。 • 第二時期 : 建材分子進一步分解成共同產物 Acetyl-CoA. • 第三時期 : 經由轉胺作用 (transamination) 與檸檬酸循環 (citric acid cycle) 形成 NH3 、 H2O 與 CO2 三個主要產物。Biosynthetic(Anabolic)
pathways diverge to yield
many product
• 例如許多脂類分子是由 acetyl-CoA 衍生 而來。
Bioenergetics
• ATP (adenosine triphosphate) is the universal currency of free energy in biological systems.
• 起初 ATP 被認為與肌肉的活力 (activity) 有關 (1929) ,之後陸陸續續的研究發現,它也存在其 他形式的細胞中,而且動、植物與微生物皆存在。 • In 1941, Fritz Lipmann saw the broad
significance of these observations and postulated the unifying concept that ATP is the primary and universal carrier of chemical energy in cells.
4.3 ATP 扮演的角色
• ATP provides energy by group
transfers not by simple
hydrolysis 。 ATP 的水解是經由原子團 的移轉並非簡單的水解。
• 以熱力學的觀點此水解反應必會自然發生 ( 自由能為負值 ) ,但以動力學的觀點而言 此反應並不一定會進行。
• 只有在特異的酵素存在下反應才能進行。
代謝中的不可逆反應雖然是伴隨著 large
and negative free-energy change 之自發反應 (spontaneous reaction) ,但反應幾乎不會進行,必須在特異的酵 素存在下,反應始能進行。可見在代謝過 程中,酵素是控制反應進行的關鍵。
• 細胞無法以一次反應就將萄葡糖燃燒成二 氧化碳,需經由一系列的代謝反應才能達 成。而其中的氧化反應步驟幾乎佔了總反 應大部分的自由能,在這些氧化反應中所 消失的電子被轉移到一些能攜帶電子的輔 助酵素 (coenzyme) 上、如
NAD+ 、 NADP+ 與 FAD ,進而形成高能的電
子儲存形式即 NADH 、 NADPH or FADH2 (energy-rich hydrogen atom) 。
醱酵的產物 人們使用某些微生物在無氧 下代謝糖而產生乾酪、葡萄酒和麵包。
醣類代謝
• 重要的醣類代謝如下 : • 1 糖解途徑 (glycolysis):catabolic pathway • 2 糖質新生 (gluconeogensis):anabolism • 3 戊糖磷酸途徑 (pentose phosphate pathway) • 4 肝醣合成 (glycogenesis):anabolism • 5 肝醣分解 (glycogenlysis):catabolismGlycolysis( 糖解途徑 )
• Glycolysis( 糖解作用 ):Inglycolysis a molecule of
glucose is degraded in a series of enzyme-catalyzed reactions
to yield two molecules of
pyruvate. 一分子的葡萄糖經一系列酵素 催化的反應分解成兩分子的丙酮酸。
• The breakdown of the six-carbon glucose into two molecules of the three-carbon
pyruvate occurs ten steps.
糖解途徑的反應
1. 合成葡萄糖 -6- 磷酸 (glucose-6-phosphate;G-6-P) 。
5. 甘油醛 -3- 磷酸 (G-3-P) 和二羥丙酮磷酸 酯 (DHAP) 的互相轉換。
8. 3- 磷酸甘油酸 (3-phosphoglycerate) 和 2-磷酸甘油酸的交互轉換。
圖
8.3
甘
油
醛
-3
-
磷
酸
的
去
氫
反
應
• stage 1.
• Glucose is phosphorylated and cleaved to form two molecules of glyceraldehyde-3-phosphate (G-3-P). The two ATP
molecules consumed during this stage are like an investment.
• The energetic gain comes in the stage 2 of glycolysis.
• Glyceraldehyde-3-phosphate is converted to pyruvate. Four ATP and two NADH are produced.
• The stage 1 of glycolysis requires 2ATP • The stage 2 of glycolysis produces 4ATP
and 2NADH
Fate of Pyruvate
• Pyruvate can be converted into ethanol, lactate, or acetyl-coenzyme A.
• pyruvate
• Anaerobic aerobic condition • condition
丙酮酸的命運
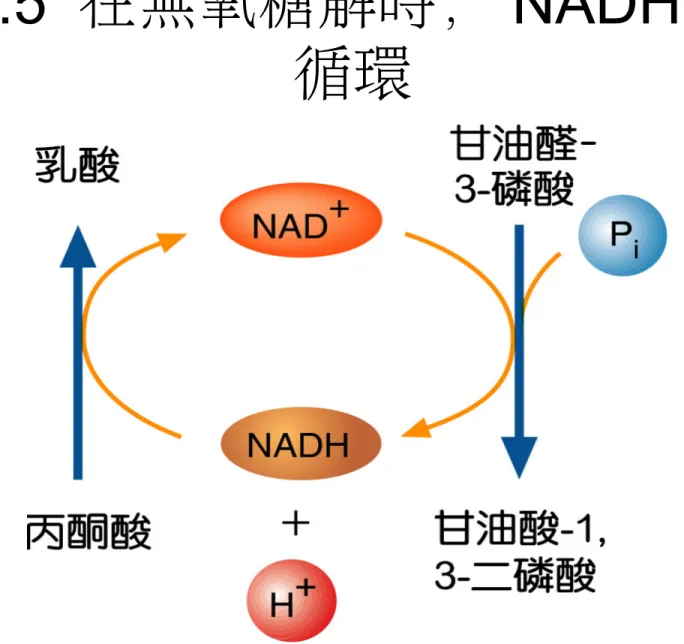
• 肌肉細胞和某細菌的品系 [ 例如,乳酸桿菌 (Lactobacillus)] ,是藉由轉換丙酮酸成為 乳酸來生產 NAD +:
圖 8.5 在無氧糖解時, NADH 的再
循環
Fermentation
• Lactic acid fermentation:
• 1. Homolactic fermenters produce only lactate.
• 2. Heterolactic or mixed acid fermenters produce several organic acids (e.g., lactic, acetic, propionic, and butyric acid.)
• Alcoholic fermentation:
• Alcoholic fermentation occurs in yeast several bacterial species.
糖解途徑的調節
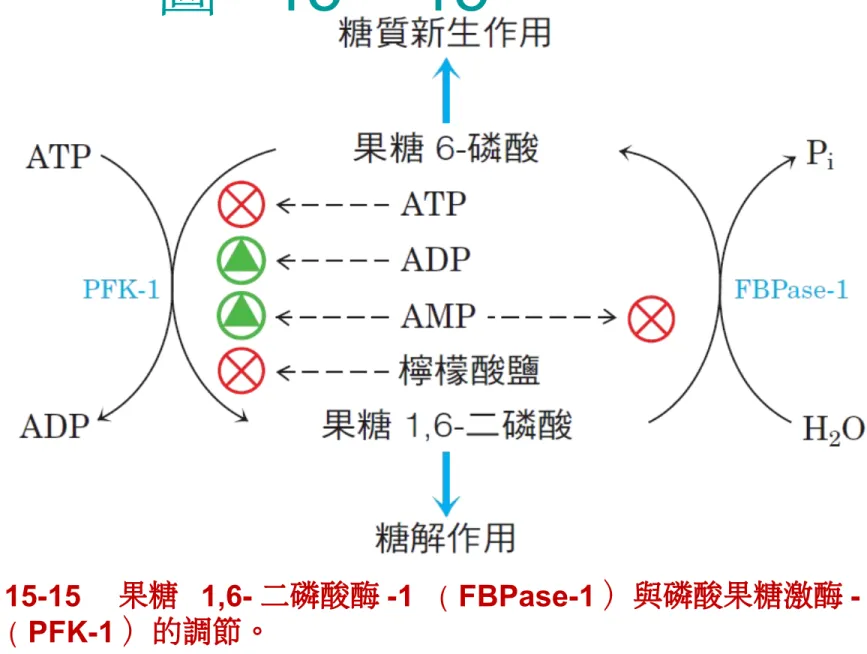
• 糖解途徑的速率調節主要是透過異位作用 (allostery) 的控制。 • Hexokinase 、 PFK-1 與 Pyruvate kinase 是三個會受到異位調節的酵素,他 們是透過 allosteric effectors 來開 啟或關閉反應。圖
15 – 15
p.625 圖 1515 果糖 1,6 二磷酸酶 1 ( FBPase1 )與磷酸果糖激酶 -1 ( PFK-1 )的調節。果糖 2,6- 二磷酸在這個受質循環的調節所扮演 的重要角色會在接下來的圖中詳述。Gluconeogenesis( 糖質新生
)
• Gluconeogenesis, the formation of new glucose molecules from noncarbohydrate precursors (e.g., lactate, glycerol,
pyruvate, and certain α-keto acids ), occurs in primarily in the liver and
糖質新生
(GLUCONEOGENESIS)
• 這些先驅物包括乳酸 (lactate) 、丙
胺酸
(alanine) 、丙酮酸
(pyruvate) 、甘油 (glycerol) 與一
些 α-Keto acid ( 來自胺基酸 ) 。
• Gluconeogenesis 是由非糖的先驅
物合成葡葡糖,主要發生在肝臟。
• 在一些情況 ( 如飢餓與代謝性酸中毒 )
腎臟也能產生
glucose 。
Gluconeogenesis reactions :
• Gluconeogenesis reactions : The
reaction sequence is largely the reverse of glycolysis. Recall that three glycolytic
reactions are irreversible.
• In gluconeogenesis, alternate energetically favorable reactions
catalyzed by different enzymes are used
8.2 糖質新生作用
• 糖質新生的反應
2. 果糖 -1,6- 二磷酸轉換為果糖 -6- 磷酸。
糖質新生的途徑大部分是糖解作用
(glycolysis) 的逆反應。
糖解作用的
hexokinase
、 PFK-1 與
Pyruvate kinase 所催化的反應是不可
逆的,所以
Gluconeogenesis 選擇了
有利能量的迂迴反應途徑避開此障礙
。
圖
8.7
醣
代
謝
:
糖
質
新
生
作
用
和
糖
解
作
用
三種迂迴途徑的反應
• 1.
合 PEP(Phosphoenolpyruvate) :
• 合成 PEP 需要兩個酵素 (Pyruvate
carboxylase
與 PEP carboxykinase)
Pyruvate carboxylase( 位於粒腺体 )
能將
Pyruvate 轉成 oxaloacetate 。
• 3.Fructose 1-6 bisphosphatase 催化 fructose 1-6 bisphosphate 轉換成
fructose-6-phosphate ,此方應的△ G0’=-4 Kcal/mole 。 fructose 1-6
bisphosphatase 是一個 allosteric enzyme, 檸檬酸會促進其活性, AMP 與 fructose 2-6 bisphosphate 會抑制其活性。
• Glucose-6-phosphatase 催化
Glucose-6-phosphate 水解成 Glucose
,此酵素是肝臟特有的酵素。
• 糖質新生是耗能的過程,需要水解 6
個 ATP 分子
圖
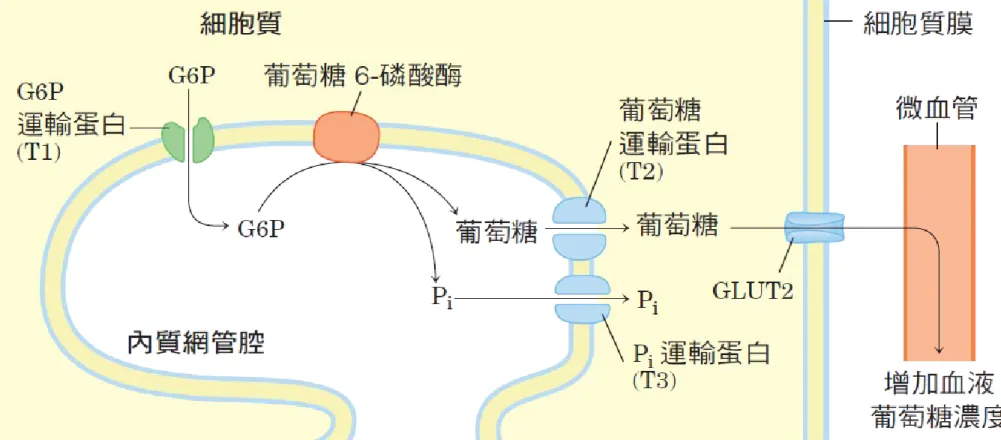
15 – 28
p.635 圖 15-28 藉由內質網的葡萄糖 6- 磷酸酶水解葡萄糖 6- 磷酸。葡萄 糖 6- 磷酸酶的催化部位在內質網管腔內側。葡萄糖 6- 磷酸( G6P ) 運輸蛋白( T1 )會將受質從細胞質運送到管腔內,而產物葡萄糖和 Pi 會由特定的運輸蛋白( T2 和 T3 )送至細胞質。最後葡萄糖再經由細 胞質膜上的運輸蛋白( GLUT2 )離開細胞。• Pyruvate carboxylase 是一個粒線體 酵素, glucose-6-phosphatase 在內質 網。其餘的酵素皆屬於細胞質酵素,因 glycolysis 與 gluconeogenesis 皆發 生在細胞質中,因此 Pyruvate carboxylase 催化的產物 oxaloacetate 須先轉成 malate 穿過粒線體的膜,然後在 細胞質中再轉換成 oxaloacetate ( 因 oxaloacetate 無法穿過粒線體膜 ) 。
肝細胞中糖解與糖質新生作用受
hormone 調節
• 昇糖素 (Glucagon): • 胰島素 (Insulin):
p.625 果糖 2,6- 二磷酸是 PFK-1 和 FBPase-1 強而有力的異位調節物 •當血糖降低時,升糖素( glucagon )會傳 遞訊息給肝臟來產生和釋放更多葡萄糖,並 且停止肝臟本身消耗葡萄糖。 •糖解作用和糖質新生作用中的快速荷爾蒙調 節是藉由果糖 2,6- 二磷酸( fructose 2,6-bisphosphate )達成,其為 PFK-1 和 FBPase-1 的異位作用物:
p.626
• 當果糖 2,6- 二磷酸結合到 PFK-1 的異位部位, 它增加酵素結合到自己受質(果糖 6- 磷酸)的親 和力,並且減少對異位抑制劑 ATP 和檸檬酸鹽的 親和力(圖 15-16 )。
圖
15 – 16(c)
p.626
圖 15-16 果糖 2,6- 二磷酸在調節糖解作用和糖質新生作用所扮演的角 色。 。 (c) F26BP 的調節總整理。
p.626 • 異位作用物果糖 2,6- 二磷酸在細胞內的濃度 取決於其形成與分解的相對速率(圖 15-17a )。 • 果糖 2,6- 二磷酸是由磷酸果糖激酶 -2 ( phosphofructokinase-2 ;簡稱 PFK-2 ) 催化果糖 6- 磷酸的磷酸化而形成的,並由果 糖 2,6- 二磷酸酶( fructose 2,6-bisphosphatase ;催化其分解(注意這些酵素 不同於磷酸果糖激酶 -1 與果糖 1,6- 二磷酸酶 ,分別催化果糖 1,6- 二磷酸的形成與分解) 。
圖
15 – 17(a)
p.627 圖 15-17 果糖 2,6- 二磷酸濃度的調節。 (a) 調節物果糖 2,6- 二磷酸 ( F26BP )在細胞內的濃度取決於磷酸果糖激酶 -2 ( PFK-2 ) 催化其 合成以及果糖 2,6- 二磷酸酶( FBPase-2 )催化其分解的速率。圖
15 – 17(b)
p.627
圖 15-17 果糖 2,6- 二磷酸濃度的調節。 (b) 這兩個酵素皆為相同多肽 鏈的一部分,並且都受到胰島素和升糖素的相反調節。
圖
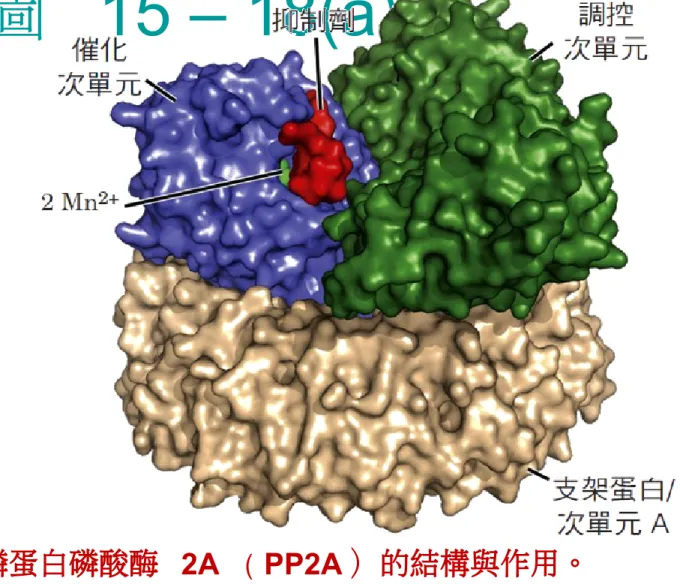
15 – 18(a)
p.627 圖 15-18 磷蛋白磷酸酶 2A ( PP2A )的結構與作用。 (a) 催化次單元 的活性部位帶有兩個 Mn2+ 離子,位在靠近由催化次單元和調控次單元之 間介面( PDB ID 2NPP )所形成的受質辨識表面。 Microcystin-LR ,以 紅色表示,是 PP2A 的專一性抑制劑。催化和調控次單元由支架蛋白 (次單元 A )支撐,使它們的位子相互靠近並使受質辨識位置成形。圖
15 – 19
p.628
圖
15 – 20
p.629 圖 15-20 丙酮酸的兩種代 謝命運。丙酮酸可利用糖質 新生作用轉化成葡萄糖和肝 醣,或氧化成乙醯輔酶 A 以提供能量產生。兩種代謝 路徑的第一個酵素均被異位 調節;由脂肪酸氧化或丙酮 酸去氫酶複合物所產生的乙 醯輔酶 A 能刺激丙酮酸羧 化酶且抑制丙酮酸去氫酶。糖質新生的受質
• 數種代謝物是糖質新生的先驅物, 3 種
重要的受質描述如下:
• 1. 乳酸:乳酸由紅血球 ( 缺粒腺体的細
胞 ) 或肌肉運動後所產生的乳酸,在柯
立循環
(Cori cycle) 到達肝臟,在肝中
有乳酸去氫脢
(lactate dehydrogenase)
將乳酸轉成丙酮酸
(Pyruvate) 然後再重
新合成萄葡糖。
Cori cycle
• Cori cycle: The sequence of glucose oxidation to lactate in peripheral tissue,
delivery of lactate to the liver, formation of glucose from lactate in the liver, and
delivery of glucose back to peripheral tissues.
• 2. 丙胺酸 (alanine) :
• 生糖的胺基酸中以丙胺酸 (alanine) 最
重要;運動的肌肉也會產生大量的丙酮
酸,丙酮酸經轉胺作用形成丙胺酸後再
釋出於血液中,再流至肝臟,在肝中
alanine 再轉成 Pyruvate 合成 glucose 。
• 3. 甘油:是脂肪在脂肪細胞代謝物,甘油 隨著血液至肝臟轉成 glycerol-3-phosphate ( 由 glycerol kinase) ,然後氧化成 DHAP 。
– 甘油,是脂肪組織的脂類新陳代謝的ㄧ種產物 ,會被血液傳送到肝臟,然後藉甘油激酶轉換 成甘油 -3- 磷酸 (glycerol-3-phosphate)[ 甘油激酶 (glycerol kinase) 只有在肝中被發現 ] 。甘油 -3- 磷 酸的氧化作用,當細胞質 NAD +濃度相對地高 時,會形成二羥丙酮磷酸 (DHAP) 。
糖質新生途徑的調控
• 糖質新生的速率主要受到受質的來源
, allosteric effectors 與荷爾蒙的影
響。
• 糖質新生在高乳酸、甘油與胺基酸時
被促進,高脂食物、飢餓與長時間絕
食會使這些三個碳的受質分子大量產
生。
• 在 Gluconeogenesis 的途徑中的四個關鍵 酵素,皆是 allosteric enzyme 。例如
fructose 1-6 bisphosphatase 受 ATP 活化 ,但受 AMP 與 fructose 2-6 bisphosphate 的抑制 Pyruvate carboxylase 受
acetyl-coA 的活化。 (acetyl-acetyl-coA 是脂肪酸分解的
• 另外可經由荷爾蒙改變 allosteric effector 與 關鍵酵素的合成來影響 Gluconeogenesis 。 • 昇糖素 (glucagon) 會減少 fructose 2-6 bisphosphate 能抑制 PFK-1 與活化 fructose 1-6 bisphosphatase 經由促進及其基因的表 現。 • 昇糖素結合肝細胞能抑制醣解酵素 Pyruvate kinase( 經由磷酸化反應 ) 。
• 可松体 (cortisol): 一種在腎上腺皮質合
成的類固醇激素,能促進糖質新生酵
素的合成。
( 可松体使身体有利於受壓
力狀態
) 。
• 胰島素會抑制糖質新生關鍵酵素的合
成。
• 2.PEP carboxykinase( 在人類中位於粒腺 体與胞質 ) 能水解 GTP 將 OAA 轉成 PEP 。
由於粒体膜無法讓 OAA 通過,因此需將它
轉成 Malate( 蘋果酸 )( 因為粒線体膜具有 Malate shuttle) , Malate dehydrogenase 催化此反應,且位於粒線体膜內與胞質中 。
• malate shuttle: It includes malate dehydrogenase
• in both the cytosol and the mitochondrial matrix, in addition to transporters in the inner mitochondrial membrane.
• As stated, each of the above reactions is matched by an opposing irreversible
reaction in glycolysis. Each set of such paired reactions is referred to as a
substrate cycle.
• Futile cycle: The loss of large amounts of energy, a situation of substrate cycle, is
usually prevented by metabolic control mechanisms.
戊糖磷酸途徑
(pentose
phosphate pathway)
• Pentose phosphate pathway• After it has been converted to glucose 6-phosphate, glucose can also enter the
pentose phosphate pathway.
• Otto Warburg 在 1931 年發現這個途徑的 第一個酵素即 glucose 6-phosphate
dehydrogenase; 然後由 Fritz Lipmann 等人 合力發表全部反應過程。
• The pentose phosphate pathway has two primary functions:
• 1 the production of NADPH
• 2 formation of ribose 5-phosphate
• NADPH is used for reductive biosynthesis. Ribose 5-phosphate is required for
biosynthesis of ribonucleotides and their derivatives, which are incorporated into RNA, DNA, and certain coenzymes.
• The pentose phosphate pathway can be divided into an oxidative stage and
nonoxidative stage.
• In the oxidative stage, NADPH is produced as glucose 6-phosphate is converted to
five-carbon compound ribulose 5-phosphate.
• In the nonoxidative stage, ribulose
5-phosphate is converted to the glycolytic intermediates fructose 6-phosphate and glyceraldehyde 3-phosphate.
• Ribulose 5-phosphate has two fates: an epimerase can catalyze the formation of xylulose 5-phosphate, or an isomerase can catalyze the formation of ribose 5-phosphate.
圖 8.11 醣代謝:糖解作用和五碳糖
磷酸途徑
• 其他重要單糖的代謝 • 果糖 (fructose) 的代謝:日常果糖的來源包 括水果、蜂蜜與蔗糖。果糖是排第二重要 的糖,能進到糖解途徑。在肝中,果糖可 直接轉成 fructose 1- phosphate 再轉成 DHAP 與 glyceraldehyde ,再進入糖解途 徑。在肌肉與脂肪組織,果糖轉成 fructose-6-phosphate 。
8.4 其他重要的糖的新陳代謝
• 半乳糖 (galactose) 代謝
• 半乳糖會經由酵素作用轉成 galactose-1-phosphate ,然後再轉為 UDP-galactose. UDP-glucose 可形成 glucose 1phosphate 或進行肝醣合成, glucose-1-phosphate 也 可轉 glucose-6-phosphate 進行糖解途徑。
• 甘露糖 (mannose) 代謝 :
• 甘露糖 (mannose) 是糖蛋白中糖基主要的
成分,雖然在食物中佔少部分, mannose
轉成 mannose-6-phosphate 再經異構脢轉 成 fructose-6-phosphate 。
圖
8.1
2
糖
類
代
謝
:
其
他
重
要
的
糖
類
肝醣代謝
• 肝醣合成 (Glycogenesis):
• Glycogen synthesis occurs after a meal, when blood glucose levels are high.
• Glycogen is synthesized and degraded by different pathways
• 1957 年, Luis Leloir 和他的共事同仁發 現,肝糖的合成是經由一個與
glycogenolysis 不同的途徑。萄葡糖基 (glycosyl) 的提供者是 UDP-glucose 並非 glucose 1-phosphate ,即 glycogenesis 並 不是 glycogenolysis 的逆反應。在生物系 統中, catabolism 和分 anabolism 的
• Glucose 合成 glycogen 的啟始步驟是 glucose 經 muscle 中的 Hexokinase 或 liver 中的 Glucokinase 催化下,形成 Glucose-6-phosphate 。
• The synthetic pathway of glycogen:
• 形成肝醣直鏈 : glucose 以 1 – 4 連結 (1 – 4 linkage) 的方式,使肝醣分子鏈增長。
肝醣生成作用
• Glycogen synthase catalyzes the transfer of glucose from UDP-glucose to a growing chain.
• 是肝醣合成的速率決定步驟 (rate-determining step) 。
• Amylo-α(1,4 – 1,6)-glucosyl
transferase(branching enzyme), which creates the α(1,6) linkages for branches in the molecule. • 形成肝醣支鏈 : 當聚葡萄糖鏈 (polyglucose chain) 增加到差不多是 6 至 11 個 glucose 時,則 有另一種酵素稱 branching enzyme ,會將約 6 個 1 – 4 連結 glucose 組成,其中一個 1 – 4 連 結的 glucose 連結轉成 1— 6 連結,形成枝狀分 子結構。
• Glycogenolysis: Glycogen degradation requires
• glycogen phosphorylase, which removes glucose residues from the nonreducing ends on the outer branches of glycogen, and
• amylo-α(1,6)-glucosidase (debranching enzyme), which hydrolyzes the α(1,6)
圖
8.1
5
肝
糖
降
解
圖
8.1
6
肝
醣
降
解
p.639 肝醣素提供肝醣合成時最初的糖基引子 • 令人疑惑的蛋白肝醣素( glycogenin )(圖 15-32 )是新肝醣鏈合成所需的引子,在此引 子上新鏈可以聚集,也是催化它們聚集的酵素 。 • 合成新肝醣分子的第一步是將 UDP- 葡萄糖上 的葡萄糖殘基轉移至肝醣素上的 Tyr194 羥基 ,此作用是由肝醣素本身的葡萄糖基轉移酶 ( glucosyltransferase )活性所催化(圖 15-33 )。
圖
15 – 32
p.639
圖
15 – 33
p.640
p.636
BOX 15-4 Carl 及 Gerty Cori :肝醣代謝與疾病的開 拓者
肝醣代謝的調節
• 肝醣磷酸脢 (Glycogen phosphorylase) 為 肝醣分解的啟始步驟,也是肝醣分解的速 率決定步驟 (rate-determining step) 。
• Regulation of Glycogen Metabolism: Glycogen metabolism is carefully
regulated to avoid wasting energy. Both synthesis and degradation are controlled through a complex mechanism involving insulin, glucagon, and epinephrine.
• 肝醣的分解及合成受到 glycogen
phosphorylase 與 Glycogen synthase 的控 制。控制機轉是經由 allosteric mechanism 或 phosphorylation 及 dephosphorylation 所造成的共價修飾等複雜的反應來完成。
• Glycogen phosphorylase and glycogen synthase have both active and inactive conformation 。 These forms are
interconverted by covalent modification.
The active form of glycogen synthase,
known as nonphosphorylation form, is converted to the inactive form, known as phosphorylation form, by phosphorylation.
• The active form of glycogen
phosphorylase, known as
phosphorylation form, is also converted
to the inactive form, known as nonphosphorylation form, by phosphorylation.
• (1) 肝中 Glycogen phosphorylase 的活 化與去活化 :
• Phosphorylase kinase: 此酵素能利用 ATP 將去活化型 phosphorylase b 轉成活化型
phosphorylase a 。
• Protein phosphatase: 可水解活化型
phosphorylase a 形成去活化型 phosphorylase b 。
• (2) 肌肉中 Glycogen phosphorylase 亦 有兩種型式存,其活性與 AMP 濃度有關。 • phosphorylase a: 為磷酸化的脢,不論 AMP 是否存在,均具有活性。 • phosphorylase b: 去磷酸的脢,當 AMP 存在時才有活性,反之 ATP 與 Glucose-6-phosphate 則會抑制 phosphorylase b 的 活性。
• 肝醣分解的控制 : 主要以 glycogen phosphorylase 的活化與去活化來控 制, cAMP 與 Ca2+ 則是間接控制肝醣分解 反應的第二信使 (Second messengers) 。 • 第二信使 (Second messengers): 在胞質中 傳遞水溶性激素或類激素訊息的分子。如 cGMP 、 IP3(Inositol Triphosphate) 、 DAG(Diacylglycerol) 、 cA MP 與 Ca2+
• cAMP
• 可活化 cAMP-dependent protein kinase , cAMP-dependent protein kinase 催化
phosphorylase kinase 的磷酸
化, phosphorylase kinase 再磷酸化
Glycogen phosphorylase ,將非活化型
的 phosphorylase b 轉成活化型 phosphorylase a 。
圖
15 – 34
p.641 圖 15-34 肌肉肝醣磷酸 化酶由共價性修飾所調節。 在較具活性狀態的酵素磷 酸化酶上,其每個次單元 的 Ser14 殘基都會被磷 酸化。磷酸化酶 a 會被 磷酸化酶 a 磷酸酶(也 是已知的磷蛋白磷酸酶 1 , PP1 )催化而移除這 些磷醯基,轉變成較不具 活性的磷酸化酶 b 。磷酸 化酶 b 能被磷酸化酶 b 激酶作用而再轉變回(再 活化成)磷酸化酶 a 。 (同時參閱圖 6-36 的肝 醣磷酸化酶調控。)p.641 • PKA 接著磷酸化並活化磷酸化酶 b 激酶 ( phosphorylase b kinase ),其能催化肝醣 磷酸化酶上的兩個相同次單元的 Ser 殘基磷 酸化,而將之活化,因此促進肝醣分解。 • 當肌肉回復到休息狀態時,第二個酵素磷酸化 酶 a 磷酸酶( phosphorylase a phosphatase ),也稱為磷蛋白磷酸酶 1 ( phosphoprotein phosphatase 1 ;簡稱 PP1 ),此酵素會將磷 酸化酶 a 上的磷醯基移除,而將之轉變成較 不具活性的磷酸化酶 b 。
圖
15 – 36
p.642 圖 15-36 肝臟的肝醣磷酸化酶可以當作葡萄糖感測器。葡萄糖結合到 肝臟磷酸化酶 a 同功酶的異位部位,因此誘導構形改變而暴露出磷酸化 的 Ser 殘基,使得磷酸化酶 a 磷酸酶 1 ( PP1 )可以作用。此磷酸 酶將磷酸化酶 a 轉變成磷酸化酶 b ,明確地降低磷酸化酶的活性,因 此在因應高血糖時會減緩肝醣的分解。胰島素也可以間接刺激 PP1 而 減緩肝醣的分解。p.643 肝醣合成酶也是由磷酸化與去磷酸化所調節 • 如同肝醣磷酸化酶,肝醣合成酶有磷酸化與去磷酸化 兩種構形(圖 15-37 )。當處於活化態時,肝醣合成 酶 a ( glycogen synthase a )是未磷酸化的。 • 最重要的調節激酶為肝醣合成酶激酶 3 ( glycogen synthase kinase 3 ,簡稱 GSK3 ),其會在肝醣合成 酶羧端附近的三個絲胺酸殘基加上磷醯基,造成肝醣 合成酶強烈失活。 • GSK3 的作用具有等級制度之分;它不會磷酸化肝醣 合成酶,直到另一個蛋白激酶酪蛋白激酶 II ( casein kinase II ,簡稱 CKII )先磷酸化肝醣合成酶附近的殘 基後,此反應稱為引導( priming )(圖 15-38a )。
水溶性激素或類激素作用機轉
• 肝醣的合成和降解被小心地調節,以
便足夠的葡萄糖可用來滿足身體的能
量需要。糖質新生作用和肝醣分解作
用,被三個主要的荷爾蒙控制著:胰
島素、胰升糖激素和腎上腺素。
圖
15 – 35
p.642 圖 15-35 腎上腺素與升糖素作用的 串聯機制。藉由與特殊的表面受體結 合,腎上腺素作用在肌原細胞(左 邊 ) ,或升糖素作用在肝細胞(右邊 ) ,皆活化 GTP- 結合蛋白 Gsα (見 圖 12-4 )。活化的 Gsα 促使 [cAMP] 增加,進而活化蛋白激酶 A ( PKA )。如此開始一連串磷酸化 作用; PKA 活化磷酸化酶 b 激酶, 磷酸化酶 b 激酶接著活化肝醣磷酸 化酶。這種串聯反應將最初訊息極度 放大;圖中粉紅色框格內可能低估此 串聯酶一步驟中分子數目實際的增加 量。肝醣分解之後供應葡萄糖,其在 肌原細胞可提供 ATP (經由糖解作 用)支持肌肉收縮,在肝細胞中葡萄 糖會被釋放至血液中以彌補低血糖。圖
15 – 39
p.644