Running Title: anti-inflammatory effects of methanolic extract of Antrodia cinnamomea
Anti-inflammatory effects of methanol extract of Antrodia cinnamomea
mycelia both in vitro and in vivo
Chi-Luan Wen1, 2, #, Chia-Chuan Chang3, #, Shyh-Shyun Huang2, Chao-Lin Kuo2, Shih-Lan Hsu4, Jeng-Shyan Deng5, Guan-Jhong Huang 2, *
1
Taiwan Seed Improvement and propagation Station, Council of Agriculture, Propagation Technology Section, Taichung, Taiwan
2
School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, College of Pharmacy, China Medical University, Taichung 404, Taiwan
3
Division of Medicinal Chemistry, National Research Institute of Chinese Medicine, Taipei 112, Taiwan
4
Department of Education and Research, Taichung Veterans General Hospital, Taichung, Taiwan
5
Department of Health and Nutrition Biotechnology, Asia University, Taichung 413, Taiwan
*
Corresponding author Dr. Guan-Jhong Huang
School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, College of Pharmacy, China Medical University, Taichung 404, Taiwan
Tel.: +886 4 2205 3366 ext 5508 Fax: +886 4 2208 3362.
E-mail address: [email protected]
#
1. Introduction
Inflammation has long been recognized as a localized protective reaction of tissue to irritation, injury, or infection that is characterized by pain, redness and swelling. Usually, inflammation is initiated through the production of specific cytokines or chemokines characterized by recruitment of leukocytes to the damage site. During inflammation, high levels of reactive oxygen species (ROS) were also produced to exert a defense against pathogens (kang et al., 2008). Among them, abnormal excess NO produced by iNOS is believed to act as a toxic radical that can damage cellular macromolecules such as proteins, DNA, and lipids, triggering several unfavorable cellular responses (Cui et al., 2006). However, sustained or excessive inflammation can lead to numerous diseases. Because inflammation involves the participation of various cell types expressing and reacting to diverse mediators along a very precise sequence of events. Therefore, inhibition of the cellular reactions is one of the strategies for treatment of inflammatory diseases. For in vivo tests, inflammation can be induced in animals by many substances. Mice paw edema is the most commonly used model for acute inflammation while subcutaneous implantation of biomaterial is usually used for inflammatory model.
The fruiting body of Antrodia cinnamomea (AC; Polyporaceae, Aphyllophorales) is well known in Taiwan as a traditional medicine. It has been used for the treatment of food and drug intoxication, diarrhea, abdominal pain, hypertension, cancer, and inflammatory disorders (Peng, et al., 2007). After the success in mass production of AC by artificial cultivation, a series of health supplements formulated from AC has been launched with high market value, and are increasingly popular in Taiwan and other Asian countries. Most AC research has been focused on the crude isolated fractions, which are
subjected to pharmacological screening or therapeutical evaluation (Wang, et al., 2003). A recent report demonstrates that the cultured mycelium has the antioxidative, anti-inflammatory, vasorelaxation, anti-cancer, and anti-hepatitis B virus activities (Geethangili, et al., 2009). However, there has been limited investigation supporting the anti-inflammatory property of this medicinal mushroom. This study was therefore aimed to evaluate the anti-inflammatory potential of the methanol extract of liquid cultured mycelia of A. cinnamomea (MEMAC) in both in vitro and in vivo models. Our observations showed that MEMAC significantly suppressed lipopolysaccharide (LPS)-stimulated proinflammatory cytokines and mediators production in RAW264.7 macrophages and human peripheral blood mononuclear cells (PBMCs). Moreover, MEMAC also inhibited the expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX2) in vitro. Besides, MEMAC effectively alleviated λ-carrageenan-induced paw edema and increased the activity of catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx) in mouse liver, as well as reduced the levels of malondialdehyde (MDA) and nitrite oxide (NO) in the edema paw, and decreased the levels of NO and TNF-α in serum of mice.
2. Materials and methods 2.1. Materials
LPS (endotoxin from Escherichia coli, serotype 0127:B8), Pam3CSK4
(Palmitoyl-3-Cysteine-Serine-Lysine-4) were purchased from Invitrogen (San Diego,
purchased from Sigma Chemical Co. (St. Louis, USA). TNF- was purchased from Biosource International Inc. (Camarillo, CA, USA). Anti-iNOS, anti-COX2, and anti--actin antibody (Santa Cruz, USA) and a protein assay kit (Bio-Rad Laboratories Ltd., Watford, Herts, U.K.) were obtained as indicated. Poly-(vinylidene fluoride) membrane (Immobilon-P) was obtained from Millipore Corp. (Bedford, MA, USA).
2.2. Strain and Culture conditions
AC, CCRC35396, was purchased from the Culture Collection and Research Center (Hsinchu, Taiwan). The culture was maintained on malt extract agar of Blakeslee’s formula and transferred into a fresh agar plate grown at 25-28oC for 25 days. Cultured mycelium were cut into disk colony (approximately 1 cm in diameter/disk) and transferred into 600 mL vessels supplemented with 200 mL of malt extract broth, incubated at 28 oC under 130 rpm shaking in the dark for 12 days, after homogenization, the cultured mycelial suspension was used as an inoculum. Ten mL of mycelia culture was inoculated into 600 mL vessels supplemented with 200 mL of modified malt extract broth, and then cultured at 25-28 oC for 60 days by shaking at 130 rpm in a dark room.
2.3. Preparation for the methanolic extracts from MEMAC
After 60 days of the incubation, the cultured mycelia were collected and extracted by methanol, filtered by Whatman No. 1 filter paper, and concentrated to dryness with a yield of 345 mg per liter medium. In RAW264.7 cell experiment, MEMAC was dissolved in dimethyl sulfoxide (DMSO) and stored at -20 oC before use.
2.4. Chemical fingerprint analysis of MEMAC by HPLC.
The HPLC fingerprint of MEMAC fraction (stock solution 2.5 mg/mL) was performed on a Shimadzu instrument equipped with a Shimadzu 10A controller and a
PDA UV-visible absorbance detector. Column: C-18 reverse phase 250 x 4.6 mm (i.d.); mobile phase: A = H2O and B = acetonitrile (ACN); gradient: 0–25 min 90–10% A;
25–45 min 10% A; detection: 280 nm, flow rate: 1 ml/min; run time 45 min, MEMAC sample 20 L.
2.5. Extraction, purification and identification of pure compounds.
The dry MEMAC (6.74 g) were suspended in H2O (300 mL) and liquid-liquid
partitioned in a separatory funnel by n-hexane (Hex, 300 mL x2), ethyl acetate (EtOAc, 300 mL x2) and n-butanol (BuOH, 300 mL x2) recursively, and then each supernatant was concentrated to give the corresponding fractions (Hex 0.11 g, EtOAc 2.31 g, BuOH 0.85 g and H2O 1.91 g). The pure compounds were purified from the EtOAc fraction and
identified by 1H and 13C NMR spectroscopy (Varian VNMRS 600 MHz NMR spectrometer) (Hattori and Sheu, 2006; Nakamura et al., 2004).
2.6. Isolation of the human peripheral blood mononuclear cells (PBMCs) from healthy volunteer
Peripheral bloods isolated from the healthy volunteer were diluted with phosphate buffered saline (PBS) by 1:1 ratio. Volume of Ficoll (Sigma, St. Louis, MO) equal to that of the diluted blood was added to a sterilized centrifuge tube, and then the blood was careful layered onto the top of Ficoll. The blood Ficoll samples were then centrifuged at 400 xg for 30 min at room temperature. After centrifugation, the opaque interface (mononuclear cells) were transferred into a clean centrifuge tube and washed for three times with PBS. The cells were then cultured for test.
2.7. Cell culture and assessment of cell viability
supplemented with 10% FBS, penicillin (100 units/mL) and streptomycin (100 g/mL), in a 95% air, 5% CO2 humidified atmosphere at 37 °C. To examine the cell viability,
RAW264.7 cells and PBMCs were incubated with various concentrations of MEMAC (0, 25, 50, 75, 100, 150, and 200 g/mL) for 24 h, respectively. Viable cells were determined by trypan blue dye exclusion method and direct counting with a hemocytometer.
2.8. Nitrite measurement
The nitrite concentration in the medium and serum levels were measured according to the Griess reaction (Dirsch et al., 1998), and the calculated concentration were taken as an indicator of NO production. The RAW264.7 cells and PBMCs were seeded into a
12-well plate and incubated with the MEMAC (0, 25, 50, and 75 g/mL) in the presence
or absence of LPS (100 ng/mL), or stimulated of RAW264.7 cells with polyIC (20 μg/mL)
and Pam3 (Pam3CSK4, 200 ng/mL) for 24 h, respectively. One hundred microliters of
each supernatant was mixed with the same volume of Griess reagent (0.75% sulfanilamidein 0.5 M HCl and 0.075% naphthylethylenediamine dihydrochloride in water), absorbance of the mixture at 550 nm was determined using an ELISA plate reader (Dynatech MR-7000; Dynatech Laboratories). The optical density at 550 nm (A550) was measured and calculated against a sodium nitrite standard curve.
2.9. Cytokine measurement
Cytokine concentrations in the supernatants and serum levels were determined by ELISA kits. The RAW264.7 macrophages and human PBMCs were seeded into a 12-well
plate at a density of 2×105 cells/well and incubated with MEMAC (0, 25, 50, and 75 g
/mL) in the presence or absence of LPS (100 ng/mL), or stimulated of RAW264.7 cells
levels of TNF- and IL-6 were analyzed by ELISA kits (OptEIA Set from BD Biosciences) according to the manufacturer's instructions.
2.10. Measurement of PGE2 production
RAW264.7 cells and PBMCs were subcultured into 12-well plates and incubated
with MEMAC (0, 25, 50, and 75 g /mL) in the presence or absence of LPS (100 ng/mL)
or stimulated RAW264.7 cells with polyIC (20 μg/mL), Pam3 (Pam3CSK4, 200 ng/mL)
for 24 h, respectively. The levels of PGE2 were assessed by EIA kit (Cayman Chemical,
Ann Arbor, MI, USA) according to the manufacturer's instructions.
2.11. Western blotting analysis
Cells were cultured without or with MEMAC (0, 10, 25, 50, and 75 g/mL) for 30 min, and then treated with LPS (100 ng/mL) for indicated time points. Cells were washed with PBS, and then scraped into microcentrifuge tubes and pelleted. The cell pellets were resuspended with lysis buffer (50 mM Tris-HCL (pH 7.4), 150 mM NaCl, 1% Triton X-100, 5 mM EDTA, 1 mM EGTA, 0.1% NP-40, 0.5 mM phenylmethylsulfonylfluoride, 1 mM dithiothreitol, 1 mM NaF, 5 μg/mL each of leupeptin and aprotinin) and incubated for 30 min at 4 °C. The cell debris was removed by microcentrifugation, followed by quick freezing of the supernatants. The protein concentration was determined by using the Bio-Rad protein assay reagent according to the manufacturer's instruction. The lysates (20 μg/lane) were separated by SDS-PAGE on polyacrylamide gels and transferred onto polyvinylidene difluoride membranes (PerkinElmer Life Sciences, Inc. Boston, MA, USA). The membranes were blocked in 5% milk in TBS-T solution for 1 h. Then membranes were subsequently probed with monoclonal anti-iNOS antibody (upstate, Charlottesville, VA, USA), anti-COX2 antibody (Santa Cruz Biochemicals, Santa Cruz,
CA, USA), and overnight at 4 °C. The blots were washed with TBS-T for three times and incubated with horseradish peroxidase-conjugated rabbit anti-rabbit IgG (1:20,000; PerkinElmer Life Sciences, Inc. Boston, MA, USA) for 1 h at room temperature. Following three further washings in TBS-T, immunoreactive bands were visualized using the ECL-Plus detection system (PerkinElmer Life Sciences, Inc. Boston, MA, USA).
2.12. Animals
Imprinting control region (ICR) mice, 6-8 weeks male, were obtained from the BioLASCO Taiwan Co., Ltd. The animals were kept in plexiglass cages at a constant temperature of 22 ± 1 °C, relative humidity 55 ± 5 % with 12 h dark-light cycle for at least 2 weeks before the experiment. Mice were given food and water ad libitum. All experimental procedures were performed according to the NIH Guide for the Care and Use of Laboratory Animals. The placebo groups were given saline intraperitoneally (i.p.) using a bent blunted 27-gauge needle connected to a 1 mL syringe. All tests were conducted under the guidelines of the International Association for the Study of Pain (Zimmermann 1983).
After a 2-week adaptation period, male ICR mice (18-25 g) were randomly assigned to five groups (n=6) in the carrageenan-induced edema experiment, there were 6 groups (n=6) of the animals in the study. The control group receives normal saline. The other five groups include a carrageenan-treated, a positive control (carrageenan + indomethecin) and MEMAC administered groups (carrageenan + MEMAC).
2.13. Carrageenan-induced edema
The carrageenan-induced hind paw edema model was used for determination of anti-inflammatory activity (Chang et al., 2009). Animals were i.p. treated with MEMAC
(100, 200 and 400 mg/kg), indomethecin or normal saline, 30 min prior to injection of 1% -carrageenan (50 L) in the plantar side of right hind paws of the mice. The paw volume was measured immediately after -carrageenan injection and at 1, 2, 3, 4, and 5 h intervals after the administration of the edematogenic agent using a plethysmometer (model 7159, Ugo Basile, Varese, Italy). The degree of swelling induced was evaluated by the ratio a/b, where a was the volume of the right hind paw after -carrageenan treatment, and b was the volume of the right hind paw before -carrageenan treatment. Indomethecin was used as a positive control. After 5 h, the animals were sacrificed and the mouse right hind feet were dissected and stored at -80oC. Also, blood were withdrawn and kept at -80oC.
Therefore, the right hind paw tissue and liver tissue were removed at the 5 h. The right hind paw tissue was rinsed in ice-cold normal saline, and homogenized at 4oC, and then the homogenate was centrifuged at 12,000 xg for 5 min. The supernatant was obtained and stored at -20 oC refrigerator for MDA assays. The whole liver tissue was rinsed in ice-cold normal saline, and homogenized at 4 oC. Then the homogenate was centrifuged at 12,000 xg for 5 min. The supernatant was obtained and stored in the refrigerator at -20 oC for the antioxidant enzymes (CAT, SOD, and GPx) activity assays. The protein concentration was determined by the Bradford method (Bio-Rad, Hercules, CA).
2.14. MDA assay
MDA from -carrageenan-induced edema foot was evaluated by the thiobarbituric acid reacting substances (TRARS) method (Chang et al., 2009). Briefly, MDA reacted with thiobarbituric acid in the acidic high temperature and formed a red-complex TBARS.
The absorbance of TBARS was determined at 532 nm.
2.15. Antioxidant enzyme activity measurement
The following biochemical parameters were analyzed to check the hepatoprotective activity of MEMAC by the methods given below. Total superoxide dismutase (SOD) activity was determined by the inhibition of cytochromec reduction (Flohe and Otting 1984). The reductionof cytochrome c was mediated by superoxide anions generatedby the xanthine/xanthine oxidase system and monitored at 550 nm.One unit of SOD was defined as the amount of enzyme requiredto inhibit the rate of cytochrome c reduction by 50%. Total catalase (CAT) activity was estimated as described elsewhere (Aebi 1984). In brief, the reduction of 10 mM H2O2 in 20 mM of phosphate buffer (pH 7) was monitored
by measuring the absorbance at 240 nm. The activity was calculated by using a molar absorption coefficient, and the enzyme activity was defined as nanomoles of dissipating hydrogen peroxide per milligram protein per minute. Total GPx activity was determined as previously reported (Paglia and Valentine, 1967). Briefly, the enzyme solution was added to a mixture containing hydrogen peroxide and glutathione in 0.1 mM Tris buffer (pH 7.2) and the absorbance at 340 nm was measured. Activity was evaluated from a calibration curve, and the enzyme activity was defined as nanomoles of NADPH oxidized per milligram protein per minute.
2.16. Histological Examination
For histological examination, biopsies of paws were taken 5 h following the interplanetary injection of -carrageenan. The tissue slices were fixed in (1.85%
formaldehyde, 1% acetic acid) for 1 week at room temperature, dehydrated by graded ethanol and embedded in Paraffin (Sherwood Medical). Sections (thickness 5 m) were deparaffinized with xylene and stained with hematoxylin and eosin (H&E) stain. All samples were observed and photographed with light microscopy. Every 3~5 tissue slices were randomly chosen from -carrageenan, indomethacin and MEMAC-treated (50 mg/kg) groups. Histological examination of these tissue slices revealed an excessive inflammatory response with massive infiltration of neutrophils [ploymorphonuclear leukocytes (PMNs)] by microscopy. The numbers of neutrophils were counted in each scope (400 x) and thereafter obtain their average count from 5 scopes of every tissue slice.
2.17. Statistical analysis
In the RAW264.7 cell experiment, data was analysed using Statsview® Statistical programme (SAS) and significance of difference between means determined by ANOVA and Least Significant Difference (LSD) test. A P value of <0.05 was considered to be statistically significant. And the animal data are expressed as mean ± S.D. Statistical evaluation was carried out by one-way analysis of variance (ANOVA followed by Scheffe's multiple range tests). Statistical significance is expressed as *p < 0.05, **p < 0.01, and ***p < 0.001.
3. Results
The HPLC fingerprint chromatogram was established for the quality control of MEMAC (Fig. 1A) which showed one major (peak 11) and sixteen minor peaks. The major peak (peak 11) and one of the minor peak (peak 8) were identified as camphorataanhydride A (retention time = 27.13 min, peak 11) and camphorataimide B (retention time = 25.21 min, peak 8) (Fig. 1B). The estimated camphorataanhydride A content was 56.87 mg and camphorataimide B content was 5.95 mg in MEMAC (6.74g).
3.2. Effect of MEMAC on Cell Viability of RAW264.7 macrophage cell line and Human PBMCs
To examine the effect of MEMAC on cell viability, RAW264.7 cells were pretreated with various concentrations (0, 25, 50, 75, 100, 150, and 200 g/mL) of MEMAC for 24 h. As shown in Fig. 2, the survival of RAW264.7 cells were not affected by exposure to lower concentrations (25 to 100 g/mL) of MEMAC compared to that of the controls. However, MEMAC-mediated cytotoxicity occurred at the higher concentrations (up to 150 g/mL). Similar dose-response was observed in human PBMCs (Fig. 2B).
3.2.1. MEMAC suppressed immunogens induced pro-inflammatory cytokine and mediator production
It is well known that immunogens such as LPS, polyIC, Pam3CSK4 can induce the
production of pro-inflammatory cytokines, including TNF-α, IL-6, and IL-1β, as well as stimulates iNOS expression. Next, the effects of MEMAC on LPS, polyIC, Pam3CSK4 induced pro-inflammatory cytokine and mediator production in RAW264.7 macrophages
were investigated, respectively. The levels of TNF-α (Fig. 3A) and IL-6 (Fig. 3B) were dose-dependently decreased by MEMAC treatment at 0-75 g/mL. MEMAC (0-75 g/mL) also blocked NO release (Fig. 3C) and PGE2 production (Fig. 3D) in RAW264.7
cells.
3.2.2. MEMAC reduced LPS-induced pro-inflammatory cytokine and mediator production in human PBMCs
Next, we tested the MEMAC might have direct effects on the expression of proinflammatory cytokines in human PBMCs. As shown in Fig. 4, MEMAC dose-dependently blocked the production of cytokines TNF-α (Fig. 4A), IL-6 (Fig. 4B)
and blocked NO release (Fig. 4C) and PGE2 production (Fig. 4D). More inhibition of
these mediators was achieved at 75 g/mL of MEMAC. MEMAC was not cytotoxicity to
PBMCs at this concentration (Fig. 2B).
3.2.3. Inhibition of LPS-induced iNOS and COX-2 protein expression by MEMAC
In order to investigate whether the inhibition of NO production was due to a decreased iNOS and COX-2 protein levels, the effect of MEMAC on iNOS and COX-2 protein expression was determined by immunoblot, the intensity of protein bands were analyzed using densitometer in three independent experiments. As depicted in Fig. 4, treatment with LPS increased the iNOS and COX-2 protein levels in RAW264.7 cells. However, MEMAC administration caused a time- (Fig. 5A) and dose-dependent (Fig. 5B) reduction of LPS-stimulated iNOS and COX-2 protein expression.
3.3.1. MEMAC alleviated carrageenan-induced mouse paw edema
To determine whether anti-inflammatory effects of MEMAC occurred in vivo, carrageenan-induced mouse hind paw edema test was conducted. As expected, there was a gradual increase in edema paw volume of mice in the carrageenan-treated group. However, in the test groups, the MEMAC significantly inhibited carrageenan-induced mouse paw edemas in a dose-dependent manner with a maximum attend at 50 mg/kg (Fig.
6). The dose-related inhibition of hind paws edema between 3 to 5 h was observed. Indomethacin as positive control (10 mg/kg) produced a significant inhibitory effect comparable to carrageenan-treated group.
3.3.2. MEMAC reduced MDA, NO and TNF- production in vivo
MDA level increased significantly in the edema paw at the 5 h after carrageenan injection (p < 0.001). As expected, administration of 10 mg/kg indomethacin significantly reduced the MDA level in the edema paw. In this time, MDA level was also decreased dose-dependently by treatment with MEMAC (Fig. 7A).
In Fig. 7B, the NO level increased significantly in the edema serum after 5 h carrageenan injection (p < 0.001). MEMAC (25 and 50 mg/kg) markedly decreased the serum NO level (p < 0.001) in carrageenan-treated mice. The inhibitory potency was similar to that of indomethacin (10 mg/kg) at 5th hour after induction.
Data from ELISA assay showed that TNF-α level increased significantly in serum after 5 h carrageenan injection (p < 0.001). However, MEMAC (25 and 50 mg/kg) and indomethacin (10 mg/kg) decreased the TNF-α level in serum at the 5th hour after carrageenan injection (p < 0.001) (Fig. 7C).
3.4. Effects of MEMAC on activities of antioxidant enzymes
At the 5th hour following carrageenan injection, liver tissues were analyzed for the biochemical parameters such as CAT, SOD, and GPx activities (Table 1). CAT, SOD, and GPx activities in liver tissue were decreased significantly by Carr administration. CAT, SOD and GPx activity were increased significantly after treated with MEMAC and 10 mg/kg indomethacin (P<0.01or P<0.001) (Table 1). High dose (50 mg/kg) of MEMAC treatment was more potent than indomethacin.
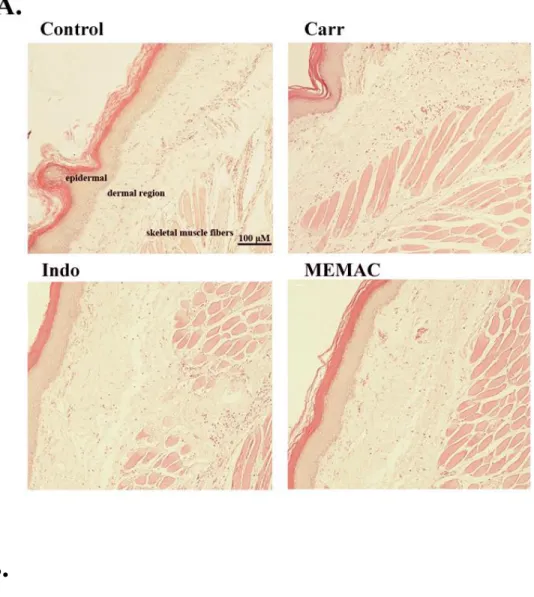
3.5. Histological examination.
Paw biopsies of control animals showed marked cellular infiltration in the connective
tissue. The infiltrates accumulated between collagen fibers and into intercellular spaces. Paw biopsies of animals treated with MEMAC (50 mg/kg) showed a reduction in carrageenan-induced inflammatory response. Actually inflammatory cells were reduced in numbers and were confined to near the vascular areas. Intercellular spaces did not show any cellular infiltrations. Collagen fibers were regular in shape and showed a reduction of intercellular spaces. Moreover, the hypodermal connective tissue was not damaged (Fig. 8A). The number of neutrophil was significantly increased upon carrageenan treatment (P < 0.001). However, indomethacin and MEMAC (50 mg/kg) could effectively decrease the neutrophil numbers as compared to the carrageenan-treated group (P < 0.001) (Fig. 8B).
In the present study, we demonstrated anti-inflammatory activities of MEMAC in both in vitro and in vivo experimental systems, using LPS-stimulated RAW264.7 macrophages and a carrageenan-induced topical inflammation model, respectively. Dual inhibitory activities against iNOS as shown in in vitro assay appear to confer on MEMAC a potent in vivo efficacy in carrageenan-induced mouse paw edema, comparable with a potent and well known COX inhibitor, indomethacin, suggesting its potential therapeutic usage as a novel anti-inflammatory source.
It is well known that LPS can induce the production of pro-inflammatory cytokines, including TNF-α, IL-6, IL-1β, and NO, as well as stimulates iNOS and COX2 expression (Sheeba, and Asha,2009). Excessive production of TNF-α, IL-6, and NO plays a critical
role in chronic inflammatory diseases (Franzotti, et al., 2000). Inhibition of inflammatory
cytokine and mediator production serves as a key mechanism in the control of inflammation that suppress the expression of these inflammation-associated cytokines and mediators have therapeutic potential for treatment of inflammatory diseases. A previous study demonstrated that the mechanisms involved anti-inflammatory activity of
methanol extract (50 g/mL) from wild fruiting bodies of A. cinnamomea was through
inhibiting iNOS, COX-2, and TNF- expression and inhibiting the phosphorylation of
extracellular signal-regulated protein kinases (ERK), c-Jun NH2-terminal protein kinases (JNK) and signal transducer and activator of transcription-1 (STAT-1) in LPS-treated
mouse microglia cell line EOC13.31 (Liu et al., 2007). It has been reported that the
aqueous extract from submerged cultivation mycelium of A. cinnamomea dose-dependently (25-100 g/mL) inhibited the LPS-induced NO, TNF-, IL-1 and PGE2 production, iNOS and COX2 protein expression via NF-B pathway in
macrophages (Hseu et al., 2005). Consistently, this study shows that suppression of iNOS and COX2 expression by MEMAC is in parallel with the comparable inhibition of NO and PGE2 production in LPS-activated RAW264.7 macrophage cell line. Moreover,
LPS-induced TNF- and L production was also markedly attenuated by MEMAC. These results reveal that A. cinnamomea has the anti-inflammatory activity in vitro.
Carrageenan-induced paw edema is a model of acute inflammation. Acute inflammation is a short-term process characterized by swelling, redness, pain, heat generation and infiltration of leukocytes at the inflammatory site. The carrageenan-induced model is highly sensitive to non-steroidal anti-inflammatory drugs, and has long been accepted as a useful phlogistic tool for investigating new drug therapies (Salvemini et al., 1996).
In the studies of the mechanism on the inflammation, TNF- and L-arginine-NO pathways have been proposed to play important roles in the carrageenan-induced inflammatory response (Yun, et al., 2008). TNF- is a mediator of carrageenan-induced inflammatory incapacitation, and is able to induce the further release of kinins and leukotrienes, which is suggested to have an important role in the maintenance of long-lasting nociceptive response (Rao et al., 2007). In this study, we found that MEMAC decreased the TNF-α level in immunogen (LPS, polyIC, and Pam3CSK4) stimulated RAW264.7 macrophages as well as in serum of carrageenan-injected mice.
This pharmacological property may be attributed to a possible molecular mechanism by effectively decreasing the production of the pro-inflammatory cytokines (TNF-and IL-6) and the expression of COX2 and iNOS in the carrageenan-injected mouse paw tissues. Furthermore, the present study also showed that carrageenan-induced paw edema was
accompanied by the production of NO. The expression of the iNOS has been proposed as an important mediator of inflammation (Dawson et al., 1991). In our study, the level of NO was decreased significantly by treatment with 25 and 50 mg/kg MEMAC in carrageenan-injected animals. Moreover, MEMAC also inhibited the NO production and iNOS expression in immunogen- (LPS, polyIC, or Pam3CSK4) stimulated RAW264.7
macrophages. Furthermore, MEMAC inhibition of LPS-stimulated TNF-, IL-6, NO and
PGE2, IL-6, NO, and PGE2 release from PBMCs was significant. We suggest the
anti-inflammatory mechanism of MEMAC may be partly through the L-arginine-NO pathway.
The carrageenan-induced inflammatory response has been also demonstrated to be linked to neutrophils infiltration and the production of neutrophils-derived free radicals, such as hydrogen peroxide, superoxide and hydroxyl radicals, as well as the release of other neutrophils-derived mediators (Hseu, et al., 2005). The MDA production is due to free radical attack plasma membrane. Thus, carrageenan-induced inflammatory effect would result in the accumulation of MDA. Glutathione (GSH) is a known oxyradical scavenger. Enhances the level of GSH conducive toward favor reduces MDA the production. It was suggested that endogenous GSH plays an important role against carrageenan-induced local inflammation (Cheng et al., 2008). In this study, there is significantly increased in CAT, SOD, and GPx activities with MEMAC treatment (Table 1). Furthermore, there are significantly decreases in MDA level with MEMAC treatment (Fig. 5A). We assume the suppression of MDA production is probably due to the increases of CAT, SOD, and GPx activities.
triterpenes, benzenoids, lignans, benzoquinone derivatives, succinic and maleic derivatives, and polysaccharides (Geethangili and Tzeng, 2009). Our preliminary phytochemical screening demonstrated the presence of terpenoids, sterols, triterpenes, maleic and succinic acid derivatives in MEMAC. Two known compounds, camphorataanhydride A (Hattori and Sheu, 2006) and camphorataimide B (Nakamura et al., 2004) (which belong to maleic and succinic acid derivatives) were isolated and identified from MEMAC. However, the pharmacological properties, including anti-inflammatory activity of camphorataanhydride A and camphorataimide B are never evaluated, limiting by difficult to produce these compounds. Zhankuic acid and antrocamphin A, isolated from ethanol extracts of wild fruiting body has shown anti-inflammatory effects (Hsieh et al., 2010). And zhankuic acids effectively inhibited the N-formyl-methionyl-leucyl-phenylalanine (fMLP) - or phorbol myristate acetate (PMA)-induced ROS production and firm adhesion by neutrophils has no significant
cytotoxic effect (Shen et al., 2004). Therefore, the observed anti-inflammatory activity of
MEMAC may be attributed from maleic and succinic acid derivatives.
In conclusion, our observations demonstrated that methanol extract from A. cinnamomea mycelia possessed anti-inflammatory and antioxidative effects. The mechanism actions of this methanol extract may be related to TNF-, COX2, iNOS, and antioxidant enzymes (CAT, SOD, and GPx). These results provide better understanding of A. cinnamomea pharmacology and positive health impact through characterization of the antioxidant and anti-inflammatory potential of this Taiwanese mushroom medicine. However, the phytochemical studies together with pharmacological and toxicological investigations are essential for complete understanding of the medicinal application.
Acknowledgement
The authors want to thank the financial supports from the National Science Council (NSC 97-2313-B-039 -001 -MY3), China Medical University (CMU) (CMU96-171,
CMU99-S-29, CCM-P99-RD-042, and CMU99-COL-10) and Taiwan Department of Heath Clinical Trial and Research Center of Excellence (DOH100-TD-B-111-004). The
LITERATURE CITED
Aebi, H., 1984. Catalase in vitro. Methods in Enzymology. 105, 121-126.
Chang, H.Y., Sheu, M.J., Yang, C.H., Leu, Z.C., Chang, Y.S., Peng, W.H., Huang, S.S., Huang, G.J., 2009. Analgesic effects and the mechanisms of anti-inflammation of hispolon in mice. Evidence-based Complementary and Alternative Medicine doi:10.1093/ecam/nep027.
Cui, X.Y., Kim, J.H., Zhao, X., Chen, B.Q., Lee, B.C., Pyo, H.B., Yun, Y.P., Zhang, Y.H., 2006. Antioxidative and acute anti-inflammatory effects of Campsis grandiflora flower. Journal of Ethnopharmacology, 103, 223-228.
Dawson, J., Sedgwick, A.D., Edwards, J.C., Lees, P., 1991. A comparative study of the cellular, exudative and histological responses to carrageenan, dextran and zymosan in the mouse. International Journal of Tissue Reactions 13, 171–185.
Dirsch, V.M., Stuppner, H., Vollmar, A.M., 1998. The Griess assay: suitable for a bio-guided fractionation of anti-inflammatory plant extracts? Planta Med. 64, 423–426.
Flohe, L., Otting, F., 1984. Superoxide dismutase assays. Methods in Enzymology 105, 93 –104.
Franzotti, E.M., Santos, C.V., Rodrigues, H.M., Mourao, R.H., Andrade, M.R., Antoniolli, A.R., 2000. Anti-inflammatory, analgesic activity and acute toxicity of Sida cordifolia L. (Malva-branca). Journal of Ethnopharmacology 72, 273–727.
Geethangili, M., Tzeng, Y.M., 2009. Review of Pharmacological Effects of Antrodia
camphorata and its Bioactive Compounds. Evidence-based Complementary and
Alternative Medicine doi:10.1093/ecam/nep108.
Hattori, M., Sheu, C.C., 2006. Compounds from Antrodia camphorata having anti-inflammatory and anti-tumor activity. US 7109232-2006-9-19.
Hseu, Y.C., Wu, F.Y., Wu, J.J., Chen, J.Y., Chang, W.H., Lu, F.J., 2005. Antiinflammatory potential of Antrodia camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-kB pathway. International Immunopharmacology 5, 1914–1925.
Hsieh, Y.H., Chu, F.H., Wang, Y.S., Chien, S.C., Chang, S.T,, Shaw, J.F., Chen, C.Y., Hsiao, W.W., Kuo, Y.H., Wang, S.Y., 2010. Antrocamphin A, an anti-inflammatory principal from the fruiting body of Taiwanofungus camphoratus, and its mechanisms. Journal of Agricultural and Food Chemistry. 58, 3153-3158.
Kang, H.S., Lee, J.Y., Kim, C.J., 2008. Anti-inflammatory activity of arctigenin from Forsythiae Fructus. Journal of Ethnopharmacology 116, 305-312.
Liu, D.Z., Liang, H.J., Chen, C.H., Su, C.H., Lee, T.H., Huang, C.T., Hou, W.C., Lin, S.Y., Zhong, W.B., Lin, P.J., Hung, L.F., Liang, Y.C., 2007. Comparative anti-inflammatory characterization of wild fruiting body, liquid-state fermentation, and solid-state culture of Taiwanofungus camphoratus in microglia and the mechanism of its action. Journal of Ethnopharmacology 113, 45–53.
Nakamura, N., Hirakawa, A., Gao, J.J., Kakuda, H., Shiro, M., Komatsu, Y., Sheu, C.C. and Hattori, M., 2004. Five new maleic and succinic acid derivatives from the mycelium of Antrodia camphorata and their cytotoxic effects on LLC tumor cell line. Journal of Natural Products 67, 46−48.
Paglia, E.D., Valentine, W.N., 1967. Studies on the quantitative and qualitative characterization of erythrocytes glutathione peroxidase. Journal of Laboratory Clinical Medicine 70, 158–169.
Peng, C.C., Chen, K.C., Peng, R.Y., Chyau, C.C., Su, C.H., Hsieh-Li, H.M., 2007. Antrodia camphorata extract induces replicative senescence in superficial TCC, and inhibits the absolute migration capability in invasive bladder carcinoma cells. Journal of Ethnopharmacology 109, 93-103.
Rao, Y.K., Fang, S.H., Tzeng, Y.M., 2007. Evaluation of the anti-inflammatory and anti-proliferation tumoral cells activities of Antrodia camphorata, Cordyceps sinensis, and Cinnamomum osmophloeum bark extracts. Journal of Ethnopharmacology 114, 78–85.
Sheeba, M.S., Asha, V.V., 2009. Cardiospermum halicacabum ethanol extract inhibits LPS induced COX-2, TNF-alpha and iNOS expression, which is mediated by NF-kappa B regulation, in RAW264.7 cells. Journal of Ethnopharmacology 124, 39-44.
Shen, Y.C., Wang, Y.H., Chou, Y.C., Chen, C.F., Lin, L.C., Chang, T.T., Tien, J.H., Chou, C.J., 2004. Evaluation of the anti-inflammatory activity of zhankuic acids isolated
from the fruiting bodies of Antrodia camphorata. Planta Med. 70, 310-314.
Wang, G.J., Tseng, H.W., Chou, C.J., Tsai, T.H., Chen, C.T., Lu, M.K.
2003.
The vasorelaxation of Antrodia camphorata mycelia: involvement of endothelial Ca(2+)-NO-cGMP pathway. Life Science 73, 2769-2783.Yun, K.J., Koh, D.J., Kim, S.H., Park, S.J., Ryu, J.H., Kim, D.G., Lee, J.Y., Lee, K.T., 2008. Anti-Inflammatory effects of sinapic acid through the suppression of inducible nitric oxide synthase, cyclooxygase-2, and proinflammatory cytokines expressions via nuclear factor-κB inactivation. Journal of Agricultural and Food Chemistry 56, 10265–10272.
Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 16, 109-110.
Figure Legends
Figue 1. HPLC analysis of MEMAC and compounds identification. (A) HPLC fingerprint of MEMAC. (B) Camphorataanhydride A (retention time = 27.13 min, peak 11) and camphorataimide B (retention time = 25.21 min, peak 8) were isolated and identified described as Materials and Methods section.
Figure. 2. Effect of MEMAC on cell viability of RAW264.7 and Human PBMCs.
RAW264.7 (2×105 cells/mL) cells and Human PBMCs (1×106 cells/mL) were treated with 0, 25, 50, 75, 100, 150 and 200 μg/mL of MEMAC for 24 h. Cell viability was checked after treatment with MEMAC using trypan blue exclusion method and examined using phase contrast microscopy. The results were displayed in percentage of control samples. In RAW264.7 cell experiment, data are presented as the mean ± S.D. for three independent experiments. Each data point represents the meant ± S.D. from one representative donor (n=3) in
human PBMCs experiment; **p<0.01, ***p<0.001 as compared with the
MEMAC treatment.
Figure 3. MEMAC inhibited immunogen induced pro-inflammatory cytokine and
mediator production. RAW264.7 (2×105 cells/mL) cells were pretreated with
MEMAC (0, 25, 50, and 75 μg/mL) for 30 min, and then stimulated with LPS (100 ng/mL), or other stimuli polyIC (20 μg/mL), Pam3 (Pam3CSK4, 200
ng/mL). Culture media were collected, the levels of (A) TNF-α and (B) IL-6
were measured at 18 h, the levels of (C) NO and (D) PGE2 were determined at
24 h. Data are presented as the mean ± S.D. for three independent experiments;
the same color column compare,*p<0.05, **p<0.01, ***p<0.001 as compared
with the LPS treatment.
production by human PBMCs. PBMCs (1×106 cells/mL) cells were pretreated with MEMAC (0, 25, 50, and 75 μg/mL) for 30 min, and then stimulated with LPS (100 ng/mL). Culture media were collected, the levels of (A) TNF-α and
(B) IL-6 were measured at 18 h, the levels of (C) NO and (D) PGE2 were
determined at 24 h. Each data point represents the meant ± S.D. from one
representative donor (n=3); *p<0.05, **p<0.01, ***p<0.001 as compared with
the MEMAC treatment.
Figure 5. MEMAC suppressed LPS-induced COX-2 and iNOS expression. (A) Time-course effect. RAW264.7 cells were pretreated with MEMAC (75 μg/mL) for 30 min, and then stimulated with LPS (100 ng/mL) for 8 and 14 h. (B) Dose-dependent effect. RAW264.7 cells were treated with MEMAC (0, 10, 25, 50, and 75 μg/mL) for 30 min, and then stimulated with LPS (100 ng/mL) for another 24 h. Protein extract was analyzed by Western blotting using specific antibodies against COX-2 and iNOS. β-actin was used as an internal loading control.
Figure 6. Effects of MEMAC and indomethacin on hind paw edema induced by carrageenan in mice. Mice were i.p. treated with MEMAC (100, 200, and 400 mg/kg), indomethacin or normal saline, 30 min prior to subcutaneous injection of 1% -carrageenan (50 L) in the plantar side of right hind paws of the mice. The paw volume was measured at 1, 2, 3, 4, and 5 h intervals after challenge using a plethysmometer. Each value was represented as mean ± S.D. *p < 0.05,
**
p < 0.01 and ***p < 0.001 as compared with the carrageenan group (one-way ANOVA followed by Scheffe’s multiple range test). Carr, carrageenan; Indo, indomethacin.
Figure 7. Effects of MEMAC and indomethacin on the levels of MDA, NO, and TNF-
production. Mice were i.p. treated with MEMAC (100, 200, and 400 mg/kg), indomethacin or normal saline, 30 min prior to injection of 1% -carrageenan (50 L) in the plantar side of right hind paws of the mice. After 5 h carrageenan challenge, the levels of (A) tissue MDA of hind paw in mice and the levels of (B) NO and (C) TNF-of serum in mice were measured. Each value represents as mean ± S.D. ###p < 0.001 as compared with the control group. *p < 0.05 and ***p < 0.001 as compared with the carrageenan group (one-way ANOVA followed by Scheffe’s multiple range test). Carr, carrageenan; Indo, indomethacin.
Figure 8. MEMAC alleviated carrageenan-induced hemorrhage, edema and leucocyte infiltration in mouse paw. (A) Mice were i.p. treated with MEMAC (100, 200, and 400 mg/kg), indomethacin or normal saline, 30 min prior to injection of 1% -carrageenan (50 L) in the plantar side of right hind paws of the mice. After 5 h carrageenan challenge, the animals were sacrificed and the mouse right hind paw tissues were removed, the tissue slices were fixed and stained with hematoxylin and eosin (H&E) stain. All samples were observed and photographed with light microscopy. Carrageenan-treated mice showed
hemorrhage with moderately extravascular red blood cell and large amount of inflammatory leukocyte mainly neutrophils infiltration in the subdermis interstitial tissue. However, indomethacin (10 mg/kg) and MEMAC (50 mg/kg) significantly ameliorated carrageenan-induced pathological events (magnification, 100x). (B) Numbers of neutrophil infiltration in paw tissue was estimated (magnification, 400x). Each value was represented as mean ± S.D. ###p < 0.001 as compared with the control group. ***p < 0.001 as compared with carrageenan group. Scale bar, 100 μm. Carr, carrageenan; Indo, indomethacin.
Table 1: Effects of MEMAC, indomethacin, and carrageenan on the liver catalase, superoxide dismutase, and glutathione peroxidase activities in mice.
Groups Catalase (U/mg protein) Superoxide dismutase (U/mg protein) Glutathione peroxidase (U/mg protein) Control 4.87 ± 0.06 26.21 ± 0.26 2.56 ± 0.32 Carr 3.13 ± 0.42### 16.45 ± 0.22### 1.42 ± 0.08### Carr + Indo 3.95 ± 0.37** 20.25 ± 0.34** 2.15 ± 0.19* Carr + MEMAC (5 mg/Kg) 3.51 ± 0.23* 19.56 ± 0.17* 1.54 ± 0.07 Carr + MEMAC (25 mg/Kg) 3.98 ± 0.21** 23.46 ± 0.27** 1.83 ± 0.16* Carr + MEMAC (50 mg/Kg) 4.28 ± 0.32*** 24.89 ± 0.12*** 2.34 ± 0.12**
Each value was represented as mean ± S.D. ###p < 0.001 as compared with the control group. *p < 0.05, **p < 0.01, ***p < 0.001 as compared with the carrageenan group (one-way ANOVA followed by Scheffe’s multiple range test). Carr, carrageenan; Indo, indomethacin.
Figure
6
Time (h)
0 1 2 3 4 5 Change s of e de ma volum e (m L) 0.00 0.02 0.04 0.06 0.08 CarrCarr and Indo
Carr and MEMAC 5 mg/kg Carr and MEMAC 25 mg/kg Carr and MEMAC 50 mg/kg
***
***
** ** **