Obesity is a well-known risk factor for insulin resis-tance, which in turn increases the risk of developing type 2 diabetes [1]; however, the pathological mecha-nism that links obesity and insulin resistance is not fully understood. Recent studies suggested that adi-pocytokines, products of adipocytes, play a role in this Kaohsiung J Med Sci March 2009 • Vol 25 • No 3 116
Received: Sep 15, 2008 Accepted: Feb 3, 2009 Address correspondence and reprint requests to: Dr Chai-Jan Chang, Department of Family Medicine, Kaohsiung Municipal Hsiao-Kang Hospital, 482 San-Ming Road, Hsiao-Kang, Kaohsiung, Taiwan.
E-mail: [email protected]
A
SSOCIATION
B
ETWEEN
S
ERUM
L
EPTIN AND
A
DIPONECTIN
L
EVELS WITH
R
ISK OF
I
NSULIN
R
ESISTANCE AND
I
MPAIRED
G
LUCOSE
T
OLERANCE
IN
N
ON
-
DIABETIC
W
OMEN
Chun-Ying Lee,1,2Chien-Hung Lee,2Sharon Tsai,5Chia-Tsuan Huang,1Ming-Tsang Wu,1,3
Shu-Yu Tai,1,2Fang-Fei Lin,6Nien-Chan Chao,1and Chai-Jan Chang4
1Department of Family Medicine, Kaohsiung Medical University Hospital, Graduate Institutes of 2Public Health and 3Occupational Safety and Health,
Kaohsiung Medical University, and Departments of 4Family Medicine, 5Laboratory Medicine and 6Nursing, Kaohsiung Municipal Hsiao-Kang Hospital,
Kaohsiung, Taiwan.
Obesity is a well known risk factor for insulin resistance and type 2 diabetes. Recently discovered adipocyte-derived proteins (leptin and adiponectin) might contribute to the pathologic mechanism linking obesity and insulin resistance. A total of 190 non-diabetic women were recruited from the Obesity Clinic of Kaohsiung Municipal Hsiao-Kang Hospital, Taiwan, between February 2003 and February 2004. All participants completed a simple questionnaire. Blood pressure and body mass index were measured; blood samples for fasting glucose, total cholesterol, high-density lipoprotein cholesterol, triglyceride, leptin, adiponectin, and fasting insulin level were collected after an overnight fast. Two-hour glucose level after a 75-g glucose tolerance test was deter-mined. Homeostasis model assessment of insulin resistance (HOMA-IR) was calculated as the index of insulin resistance. Multivariate linear regression analyses were used to analyze the rela-tionship between adipocytokines and insulin resistance after adjusting for possible confounding factors. Leptin and adiponectin were found to be independently associated with HOMA-IR and fasting insulin concentration, but in divergent directions, after adjusting for potential confound-ing factors. Adiponectin, but not leptin, was associated with impaired glucose tolerance after adjusting for potential confounding factors. The results suggest that leptin and adiponectin may be involved in the pathophysiologic link between obesity and insulin resistance independently. Low levels of adiponectin may increase the risks of developing impaired glucose metabolism and type 2 diabetes.
Key Words:adipocytokines, adiponectin, insulin resistance, leptin, obesity (Kaohsiung J Med Sci 2009;25:116–25)
association. Leptin and adiponectin, two of the most abundant adipocyte products, are thought to link obesity, insulin resistance, and related disorders [2,3]. Leptin, an obese gene product, is a 16-kDa protein and is predominantly expressed in adipose tissue [4]. Leptin regulates body weight by modulating appetite and energy expenditure by acting on the hypothala-mus and inhibiting the release of neuropeptide Y in mice and humans [5]. Rising levels of leptin will cause a signal to be sent to the brain, which then attempts to protect the subject from obesity by decreasing appetite and increasing energy expenditure. Nevertheless, most obese humans have elevated concentrations of leptin [6], suggesting that these people develop leptin resis-tance. Meanwhile, studies have also shown that insulin resistance is particularly prevalent in obese humans and have reported an independent association between insulin resistance and elevated plasma leptin levels [7,8]. The coinciding plasma leptin levels, plasma insulin levels and body fat suggest that leptin might be involved in the link between obesity and β-cell hypersecretion.
Adiponectin is a relatively abundant plasma pro-tein of approximately 30 kDa in size specifically secreted from adipose tissue [9]. Although the ex-pression of adiponectin mRNA occurs exclusively in adipose tissue, interestingly, the adiponectin concen-tration is decreased in obese subjects [10]. Previous reports have demonstrated that diabetic subjects have lower levels of plasma adiponectin than non-diabetic subjects, independent of body mass index (BMI) [11], indicating an association between lower adiponectin levels and type 2 diabetes.
Although it appears that leptin and adiponectin may contribute to the development of insulin resistance and type 2 diabetes in obese subjects, the association between adiponectin and insulin resistance and type 2 diabetes may be modulated by leptin, or vice versa. Insulin resistance is an important risk factor for type 2 diabetes. Therefore, we examined the associations between leptin and adiponectin with insulin resis-tance in non-diabetic women.
M
ETHODS ANDS
UBJECTSStudy population
A total of 190 women were recruited from the Obesity Clinic at Kaohsiung Municipal Hsiao-Kang Hospital,
Taiwan, between February 2005 and February 2006. Eligible subjects were those who did not have diabetes or were not taking medications that affect glucose, blood pressure, or lipid metabolism. Subjects whose body weight increased or decreased by more than 5 kg in 6 months before entering the study were also excluded. All participants completed a simple ques-tionnaire on their medication history and lifestyle characteristics, including cigarette smoking accord-ing to the categories “never”, “former” (quit smokaccord-ing at least 1 year ago) and “current smoker” (smoke more than one cigarette/day for at least 1 year) and exercise according to the categories “not regular” (less than 150 minutes per week at less than a moderate inten-sity of exercise) and “regular exerciser” (at least 150 minutes per week at a moderate or greater intensity of exercise). Written informed consent was obtained from all study subjects, and the study was approved by the Human Experiment and Ethics Committee of Kaohsiung Municipal Hsiao-Kang Hospital.
Anthropometry and blood pressure
assessment
Height and body weight were measured without shoes and with the study subjects wearing light clothes. Height was measured to the nearest 0.1 cm, and weight was measured to the nearest 0.1 kg. BMI was calculated as weight/height2 (kg/m2). Subjects were classified into: (1) normal/overweight; (2) obese; and (3) severely obese, based on a definition of obesity for the population in Taiwan, with BMI cut-off values of 27 and 30 kg/m2, respectively. Waist circumference was measured to the nearest 0.1 cm at the level be-tween the midpoint of the lowest rib and iliac crest parallel to the floor in a standing position, while the hip circumference was measured to the nearest 0.1 cm at maximum extension of the buttocks. The waist-to-hip ratio was calculated as an index of central obesity. Percent body fat was measured by bioelectrical imped-ance analysis (BIA) using the Tanita TBF-410 Body Fat Analyzer (Tanita Corp., Tokyo, Japan). The instrument has been previously validated against dual-energy X-ray absorptiometry [12]. Subjects were requested not to eat or drink anything for at least 2 hours and to refrain from strenuous exercise or consuming alcohol for 12 hours before BIA measurement to avoid per-turbation of hydration status. Blood pressure was measured using a sphygmomanometer; two readings were taken at 10-minute intervals after subjects had
been seated for at least 10 minutes. The two readings were averaged.
Adipocytokines, insulin resistance and
metabolic profiles
Fasting glucose, total cholesterol, high-density lipopro-tein (HDL) cholesterol, triglyceride, insulin, leptin, and adiponectin were measured using blood samples obtained after the participants had fasted overnight. Cholesterol, HDL cholesterol, and triglyceride con-centrations were analyzed by enzymatic colorimetric methods and the fasting glucose level was measured by the glucose oxidase method. At 2 hours after a 75-g oral glucose tolerance test, blood was drawn and the glucose level was measured by a glucose dehydroge-nase method. The 2-hour glucose level was used as an index of abnormal glucose metabolism. Using cri-teria established by the World Health Organization in 1999 [13], we classified our subjects into those with normal glucose levels, impaired fasting glucose, impaired glucose tolerance (IGT), or type 2 diabetes. Subjects with type 2 diabetes were excluded from the study. Plasma leptin, adiponectin and insulin levels were measured using a radioimmunoassay from Linco Research, Inc. (St Charles, MO, USA); intra- and in-terassay coefficients of variation were 3.9% and 4.6% for leptin, 6.2% and 6.9% for adiponectin, and 2.2% and 3.8% for insulin, respectively. Insulin resistance was estimated from the homeostasis model assessment of insulin resistance (HOMA-IR) originally described by Matthews et al [14] as follows:
HOMA-IR= [fasting insulin concentration (μU/ mL)× fasting glucose concentration (mmol/L)]/22.5.
Statistical analysis
Descriptive data are expressed as means and standard deviations or frequencies and percentages. Pearson correlation coefficients were used to determine the associations between leptin, adiponectin and contin-uous variables. Linear trends of leptin and adipo-nectin levels and metabolic parameters was tested across the three obesity subgroups using the Mantel-Haenszel test with adjustment for age, exercise and smoking status. Multiple regression analysis was used to examine the association between parameters of in-sulin resistance (HOMA-IR, fasting inin-sulin levels or 2-hour glucose levels) and adipocytokines (leptin and adiponectin) after adjusting for age, smoking status and exercise. Multiple logistic regression analysis was
used to explore the association between IGT (2-hour glucose level > 140 mg/dL) and adipocytokines after controlling for potential confounding factors such as age, BMI, fasting glucose level, fasting insulin level, triglyceride, HDL cholesterol and blood pressure. All statistical operations were performed using Stata SE software version 9.0 (StataCorp LP, College Station, TX, USA). A p value of less than 0.05 was considered significant.
R
ESULTSDescriptive data are shown in Table 1. The subjects with BMI ≥ 30 kg/m2 were, on average, slightly younger than those in the other two groups (Table 1). Regular exercisers were more prevalent in the rela-tively lean group (BMI< 27 kg/m2). The prevalence of current smokers was less than 10% in all three groups. Mean values for fasting glucose, 2-hour glucose, tri-glycerides, insulin, HOMA-IR, leptin, and systolic and diastolic blood pressure increased with increasing BMI, after adjusting for age, exercise and smoking status. In contrast, levels of HDL cholesterol and adipo-nectin increased with decreasing BMI. Table 2 shows that plasma leptin was positively correlated with measures of obesity, fasting insulin and HOMA-IR, but was not correlated with fasting glucose, 2-hour glucose, triglycerides, HDL cholesterol or blood pres-sure. Adiponectin was negatively correlated with measures of obesity, fasting insulin, HOMA-IR, 2-hour glucose levels and triglycerides, and positively corre-lated with HDL cholesterol. Leptin and adiponectin levels were inversely and significantly correlated.
The associations between leptin, adiponectin, insulin resistance and hyperinsulinemia are shown in Table 3. The multiple linear regression analyses re-vealed that serum leptin levels were positively and significantly associated with HOMA-IR (R2= 31.6%) and insulin level (R2= 39.2%), but not 2-hour glucose after controlling for the effects of adiponectin, age, smoking status, exercise and BMI. Serum adiponectin levels were negatively and significantly associated with HOMA-IR, insulin level and 2-hour glucose, after controlling for the effects of leptin, age, smoking status, exercise and BMI.
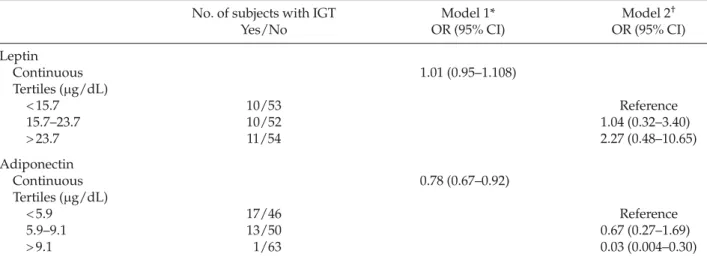
We then dichotomized the subjects for the pres-ence of IGT and found that adiponectin level, but not leptin level, was significantly associated with IGT after
controlling for covariates including age, BMI, exer-cise, smoking status, fasting glucose, HDL cholesterol, triglycerides, fasting insulin, systolic blood pressure and diastolic blood pressure (Table 4). Every 1 ng/dL increase in adiponectin level was associated with a 0.22-fold decreased risk of having IGT. When we cat-egorized subjects according to adiponectin levels into tertiles, we found that subjects in the highest adipo-nectin tertile (>9.1μg/dL) had significantly decreased risk of having IGT (odds ratio, 0.03; 95% confidence interval, 0.004–0.30; p= 0.002) compared with those in the lowest tertile, after controlling for other covariates including age, BMI, exercise, smoking status, fasting glucose, HDL cholesterol, triglycerides, fasting insulin, systolic blood pressure and diastolic blood pressure (Table 4). In contrast, we found no significant effect of leptin level, by tertiles, on the risk of IGT.
D
ISCUSSIONThe aim of our study was to examine the associations between leptin, adiponectin and insulin resistance after controlling for obesity and other covariates, be-cause leptin, adiponectin and insulin resistance are all highly correlated with obesity. We found that higher
BMI levels were associated with higher concentra-tions of leptin, fasting insulin, HOMA and lower con-centrations of adiponectin. The physiologic function of leptin is to provide a signal to the brain to decrease appetite, increase fuel consumption and control stores of body fat in rodents and humans [6,15]. However, instead of leptin deficiency, the leptin concentration was positively associated with adiposity in our study. Moreover, other studies have revealed that the elevated circulating plasma leptin does not suppress appetite or prevent fat deposition in obese humans [16], which suggests that hyperleptinemia rather than leptin deficiency is a common feature of human obe-sity, because the concentrations of leptin in plasma may reflect the degree of body adiposity in most human beings. In contrast to leptin, adiponectin was decreased in the obese subjects in our study and in other studies [10,17], even though adiponectin is mainly synthesized in adipose tissue. Taken together, obese subjects tend to have high levels of leptin, low levels of adiponectin and hyperinsulinemia.
Obesity, particularly visceral obesity, is an impor-tant risk factor for insulin resistance. The significant positive association between BMI and insulin resis-tance (HOMA-IR and fasting insulin level) was also demonstrated in our study. Whether the association
Table 1.Participant characteristics and metabolic measures according to degree of obesity
BMI< 27 27 ≤ BMI < 30 BMI ≥ 30
(n= 70) (n= 58) (n= 62) Age (yr) 42.9± 12.0 41.6± 9.4 36.5± 12.0 Current smoker (%) 5 (7.1) 2 (3.5) 5 (8.1) Exercise (%) 32 (45.7) 28 (48.3) 25 (40.3) BMI (kg/m2) 24.4± 1.8 28.4± 0.8 34.6± 4.2 Waist-to-hip ratio 0.82± 0.06 0.85± 0.06 0.85± 0.05 Waist circumference 82.0± 6.4 90.1± 6.4 99.4± 9.7 % body fat 34.0± 4.5 40.0± 4.3 47.6± 7.5 p for trend* Fasting glucose (mg/dL) 91.7± 8.3 94.6± 10.7 93.8± 12.0 0.04 2-hr glucose (mg/dL) 105.3± 27.3 110.1± 29.4 116.6± 31.1 0.004 TG (mg/dL) 105.1± 55.5 134.0± 64.3 138.0± 62.3 0.001 HDL cholesterol (mg/dL) 59.5± 16.2 53.4± 13.4 50.2± 9.0 < 0.0001
Fasting insulin (μIU/mL) 6.2± 5.0 9.2± 6.6 15.7± 9.0 < 0.0001
HOMA-IR (μU⭈mol−1⭈L−3) 1.5± 1.4 2.2± 1.7 3.6± 2.1 < 0.0001
Leptin (μg/dL) 15.5± 8.0 19.6± 6.6 30.6± 12.6 < 0.0001
Adiponectin (μg/dL) 10.5± 5.3 7.5± 3.8 7.0± 3.3 < 0.0001
SBP (mmHg) 123.1± 13.1 131.1± 15.4 131.8± 14.0 < 0.0001
DBP (mmHg) 79.8± 9.5 84.5± 9.8 85.0± 9.4 0.0001
*Trend analysis was adjusted for age, exercise and smoking status. BMI = body mass index; TG = triglycerides; HDL = high-density lipoprotein; HOMA-IR = homeostasis model assessment of insulin resistance; SBP = systolic blood pressure; DBP = diastolic blood pressure.
between obesity and insulin resistance is causal or due to an underlying association with confounding variables remains to be clarified. Therefore, our analy-ses were adjusted for measures of obesity and poten-tial confounding factors to examine the relationship between leptin, adiponectin and insulin resistance. The results revealed that high leptin levels and low adiponectin levels were independently associated with hyperinsulinemia and insulin resistance, suggesting that high leptin levels and low adiponectin levels are associated with hyperinsulinemia and insulin resis-tance independently of obesity, and may play a role in the development of insulin resistance. Boden et al [18] observed that prolonged infusion of insulin depen-dently increases serum leptin concentration during euglycemic-hyperinsulinemic clamps in non-obese young men. Some studies also showed that insulin can stimulate leptin secretion due to increased triglyceride storage in fat cells [19,20], while leptin may directly suppress insulin secretion from pancreatic islets iso-lated from ob/ob mice and humans [21,22]. However, insulin secretion did not affect leptin synthesis in hyperleptinemic insulin-resistant individuals inde-pendently of the level of obesity [7]. Taken together, these findings suggest that there is a feedback path-way between leptin and insulin secretion, but the development of “insulin insensitivity” or “leptin insensitivity” could adversely affect the association between leptin and insulin, and stimulate the pro-gression to hyperinsulinemia and impaired glucose metabolism.
The relationship between adiponectin and insulin resistance/hyperinsulinemia in our study can be explained by some experimental studies. Expression of the adiponectin receptor was detected in rat and human β-cells [23] and the expression of adiponectin can be stimulated by insulin [17], which suggests that adiponectin may modulate β-cell function. The admin-istration of adiponectin to normal, obese and diabetic rodents decreased gluconeogenesis in the liver and increased fatty acid oxidation in muscle [24] and thus regulates insulin sensitivity and energy homeostasis. However, the effect of adiponectin on insulin secre-tion is controversial. Staiger et al [25] demonstrated that adiponectin does not affect insulin secretory function of healthy human islets in vitro. Winzell et al [26] reported a dual effect of adiponectin to modify insulin secretion in insulin-resistant mice because adiponectin inhibited insulin secretion at low glucose
T
able 2.
Pearson corr
elation coef
ficients between leptin, adiponectin, obesity (including BMI, waist-to-hip ratio and per
cent body fat),
and metabolic pr ofiles BMI WHR % Fat Fasting 2-hr TG HDL-C SBP DBP Insulin HOMA-IR Leptin Adiponectin glucose glucose Leptin 0.72* 0.12 0.69* −0.09 0.06 0.13 −0.08 0.12 0.1 0.51* 0.45* – −0.16 ‡ Adiponectin −0.28* −0.19 † −0.30* −0.1 1 −0.29* −0.28* 0.43* −0.08 −0.14 −0.32* −0.30* −0.16 ‡ – *p < 0.001; †p < 0.01; ‡p < 0.05. BMI =
body mass index; WHR
= waist-to-hip ratio; TG = triglycerides; HDL-C = high-density lipopr otein cholester ol; SBP = systolic blood pr essur e; DBP = diastolic blood pr essur e; HOMA-IR =
homeostasis model assessment of insulin r
(2.8 mM) but augmented insulin secretion at high glucose (16.7 mM) concentration. In contrast, adipo-nectin did not have an acute effect on insulin secre-tion in islets from normal mice. Therefore, we believe that adiponectin modulates insulin secretion but involves a complicated mechanism. Recent studies have found that the adiponectin level is decreased in patients with type 2 diabetes or coronary heart dis-ease [28], and is associated with anti-inflammation [27,28]. One randomized interventional trial of weight reduction revealed that body weight reductions were associated with increased adiponectin levels and decreased levels of inflammatory factors such as C-reactive protein, tumor necrosis factor (TNF)-α, and interleukin (IL)-6 [29], indicating that adiponectin might protect against inflammation and atherosclerosis. However, it has also been reported that inflammatory
mediators, such as TNF-α and IL-6, which are in-creased in obese and insulin resistant individuals, can suppress the transcription of adiponectin in adipo-cyte cell lines [30,31], which explains, at least in part, the decreased level of adiponectin in obese subjects.
Leptin, adiponectin and IGT
We found that the level of adiponectin, but not leptin, was associated with 2-hour glucose level after con-trolling for BMI, age, cigarette smoking and exercise status. We then classified the study subjects into those with or without IGT, and found that high adiponectin levels were independently associated with decreased risk of developing IGT (odds ratio, 0.78) after control-ling for possible confounding variables including BMI, age, fasting glucose level, HDL cholesterol, tri-glycerides, fasting insulin concentration, systolic and
Table 3.Multiple regression analyses of adipocytokines and insulin resistance*
HOMA-IR Insulin 2-hr glucose
B SE p B SE p B SE p
BMI 0.10 0.04 0.004 0.44 0.14 0.002 0.31 0.60 NS
Leptin 0.03 0.02 0.036 0.16 0.06 0.009 0.16 0.26 NS
Adiponectin −0.09 0.03 0.001 −0.36 0.11 0.001 −1.87 0.46 < 0.0001
Model R2 31.6 39.2 15.3
*Each model was adjusted for age, cigarette smoking and exercise. HOMA-IR= homeostasis model assessment of insulin resistance; BMI= body mass index; SE = standard error.
Table 4.Multiple regression analysis for leptin, adiponectin and the presence of impaired glucose tolerance
No. of subjects with IGT Model 1* Model 2†
Yes/No OR (95% CI) OR (95% CI)
Leptin Continuous 1.01 (0.95–1.108) Tertiles (μg/dL) < 15.7 10/53 Reference 15.7–23.7 10/52 1.04 (0.32–3.40) > 23.7 11/54 2.27 (0.48–10.65) Adiponectin Continuous 0.78 (0.67–0.92) Tertiles (μg/dL) < 5.9 17/46 Reference 5.9–9.1 13/50 0.67 (0.27–1.69) > 9.1 1/63 0.03 (0.004–0.30)
*Adjusted for age, body mass index, exercise, smoking status, fasting glucose, high-density lipoprotein cholesterol, triglycerides, fasting insulin, systolic and diastolic blood pressure, and leptin and adiponectin were analyzed as continuous variables in the model;
†adjusted for age, body mass index, exercise, smoking status, fasting glucose, high-density lipoprotein cholesterol, triglycerides,
fasting insulin, systolic and diastolic blood pressure, and leptin and adiponectin were analyzed as categorical variables in the model. IGT = impaired glucose tolerance; OR = odds ratio; CI = confidence interval.
diastolic blood pressure, cigarette smoking and exer-cise. Meanwhile, subjects in the highest adiponectin tertile had the lowest risk of having IGT. IGT has been used as a marker for insulin resistance [32], denoting a pre-diabetic state in which subjects are at increased risk of progressing to diabetes [33], premature mortality and cardiovascular disease [34]. These findings sug-gest that low levels of adiponectin may be involved in the pathogenesis of abnormal glucose metabolism. A prospective cohort study by Snijder et al [35] has reported that low levels of adiponectin are indepen-dently associated with a higher risk of IGT and type 2 diabetes in elderly Caucasian women. Another study by Tso et al [36] revealed that polymorphisms of
ADIPOQ, the gene encoding adiponectin, was
associ-ated with glycemic status in southern Chinese people, suggesting a biomolecular mechanism for adiponectin and abnormal glucose metabolism. However, we did not find an association between leptin and 2-hour glucose level and IGT after controlling for BMI and other possible confounding factors, suggesting that leptin is less important in the development of IGT. Wannamethee et al [37] reported that the association between leptin and type 2 diabetes disappeared in older men after adjusting for insulin resistance in a prospective cohort study. Similarly, Kanaya et al [38] reported that the association between leptin and the incidence of type 2 diabetes disappeared after adjust-ing for metabolic syndrome-related factors. However, Panarotto et al [39] reported that leptin is indepen-dently associated with IGT and type 2 diabetes in Caucasian females, although that study did not deter-mine adiponectin. Therefore, the associations between leptin and impaired glucose metabolism may be mediated by insulin resistance and other factors.
The limitation of our study is that the cross-sectional design prohibits us from inferring a causal relationship. Second, most of our study subjects were obese because they were recruited from obesity clinics. Therefore, the study subjects may be more insulin-resistant and thus skew the data; however, after log-transforming our data (which was carried out before data analysis and which we have not shown in this paper) and adjusting for potential confounding factors, the results remained valid and the significance should not be ignored. Third, we did not collect nutritional information from the study subjects; therefore, possible associations between nutritional effects, adipocytokines and insulin resistance are not clear in our study.
Obesity is associated with the development of the metabolic syndrome, a cluster of metabolic abnor-malities, including hypertension, abnormal glucose metabolism, dyslipidemia, and hyperuricemia. Insulin resistance and compensatory hyperinsulinemia may be a potential link between obesity [40]. Therefore, early detection of insulin resistance is important for early intervention and prevention of obesity-related comorbidities.
In conclusion, we found associations between leptin and adiponectin and obesity and related metabolic disorders, although the associations were in opposite directions. Leptin and adiponectin were both associated with insulin resistance and hyperinsulinemia inde-pendently of obesity, suggesting that leptin and adipo-nectin may be involved in the pathophysiologic link between obesity and insulin resistance. However, low levels of adiponectin, but not leptin, were associated with 2-hour glucose levels and IGT, suggesting that adiponectin is a potential determinant for the progres-sion of insulin resistance to type 2 diabetes, and low levels of adiponectin might predict the increased risk of developing impaired glucose metabolism, type 2 dia-betes, premature mortality and cardiovascular disease.
A
CKNOWLEDGMENTSThis study was supported by research grants from Kaohsiung Municipal Hsiao-Kang Hospital, Taiwan (KMHK-94-004).
R
EFERENCES1. Obesity: preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ
Tech Rep Ser 2000;894:1–253.
2. Ukkola O, Santaniemi M. Adiponectin: a link between excess adiposity and associated comorbidities? J Mol
Med 2002;80:696–702.
3. Matsuzawa Y, Shimomura I, Kihara S, et al. Impor-tance of adipocytokines in obesity-related diseases.
Horm Res 2003;60 Suppl 3:56–9.
4. Zhang Y, Proenca R, Maffei M, et al. Positional cloning of the mouse obese gene and its human homologue.
Nature 1994;372:425–32.
5. Jequier E. Leptin signaling, adiposity, and energy bal-ance. Ann N Y Acad Sci 2002;967:379–88.
6. Lonnqvist F, Arner P, Nordfors L, et al. Overexpression of the obese (ob) gene in adipose tissue of human obese subjects. Nat Med 1995;1:950–3.
7. Segal KR, Landt M, Klein S. Relationship between insulin sensitivity and plasma leptin concentration in lean and obese men. Diabetes 1996;45:988–91.
8. Bodkin NL, Nicolson M, Ortmeyer HK, et al. Hyper-leptinemia: relationship to adiposity and insulin resis-tance in the spontaneously obese rhesus monkey. Horm
Metab Res 1996;28:674–8.
9. Maeda K, Okubo K, Shimomura I, et al. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene tran-script 1). Biochem Biophys Res 1996;221:286–9.
10. Arita Y, Kihara S, Ouchi N, et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity.
Biochem Biophys Res Commun 1999;257:79–83.
11. Hotta K, Funahashi T, Arita Y, et al. Plasma concentra-tions of a novel, adipose-specific protein, adiponectin, in type 2 diabetic patients. Arterioscler Thromb Vasc Biol 2000;20:1595–9.
12. Jebb SA, Cole TJ, Doman D, et al. Evaluation of the novel Tanita body-fat analyser to measure body com-position by comparison with a four-compartment model. Br J Nutr 2000;83:115–22.
13. World Health Organization. Definition, Diagnosis, and
Classification of Diabetes Mellitus and its Complications. Report of a WHO Consultation. Part 1: Diagnosis and Classification of Diabetes Mellitus. Geneva: World Health
Organization, 1999.
14. Matthews DR, Hosker JP, Rudenski AS, et al. Homeo-stasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985;28:412–9. 15. Campfield LA, Smith FJ, Burn P, et al. The OB protein
(leptin) pathway—a link between adipose tissue mass and central neural networks. Horm Metab Res 1996;28: 619–32.
16. Hamann A, Matthaei S. Regulation of energy balance by leptin. Exp Clin Endocrinol Diabetes 1996;104:293–300. 17. Scherer PE, Williams S, Fogliano M, et al. A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 1995;270:26746–9.
18. Boden G, Chen X, Kolaczynski JW, et al. Effects of pro-longed hyperinsulinemia on serum leptin in normal human subjects. J Clin Invest 1997;100:1107–13.
19. Meinders AE, Toornvliet AC, Pijl H, et al. Leptin. Neth J
Med 1996;49:247–52.
20. Carantoni M, Abbasi F, Azhar S, et al. Plasma leptin con-centrations do not appear to decrease insulin-mediated glucose disposal or glucose-stimulated insulin secretion in women with normal glucose tolerance. Diabetes 1997; 47:244–7.
21. Kieffer TJ, Heller RS, Leech CA, et al. Leptin suppression of insulin secretion by the activation of ATP-sensitive K+ channels in pancreatic beta-cells. Diabetes 1997;46: 1087–93.
22. Seufert J, Kieffer TJ, Leech CA, et al. Leptin suppres-sion of insulin secretion and gene expressuppres-sion in human
pancreatic islets: implications for the development of adipogenic diabetes mellitus. J Clin Endocrinol Metab 1999;84:670–6.
23. Kharroubi I, Rasschaert J, Eizirik DL, et al. Expression of adiponectin receptors in pancreatic beta cells. Biochem
Biophys Res Commun 2003;312:1118–22.
24. Yamauchi T, Kamon J, Waki H, et al. The fat-derived hormone adiponectin reverses insulin resistance asso-ciated with both lipoatrophy and obesity. Nat Med 2001;7:941–6.
25. Staiger K, Stefan N, Staiger H, et al. Adiponectin is functionally active in human islets but does not affect insulin secretory function or beta-cell lipoapoptosis.
J Clin Endocrinol Metab 2005;90:6707–13.
26. Winzell MS, Nogueiras R, Dieguez C, et al. Dual action of adiponectin on insulin secretion in insulin-resistant mice. Biochem Biophys Res Commun 2004;321:154–60. 27. Ouchi N, Kihara S, Arita Y, et al. Novel modulator
for endothelial adhesion molecules: adipocyte-derived plasma protein adiponectin. Circulation 1999;100: 2473–6.
28. Yokota T, Oritani K, Takahashi I, et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood 2000;96:1723–32.
29. Esposito K, Pontillo A, Di Palo C, et al. Effect of weight loss and lifestyle changes on vascular inflammatory markers in obese women: a randomized trial. JAMA 2003;289:1799–804.
30. Maeda N, Shimomura I, Kishida K, et al. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30.
Nat Med 2002;8:731–7.
31. Fasshauer M, Kralisch S, Klier M, et al. Adiponectin gene expression and secretion is inhibited by interleukin-6 in 3T3-L1 adipocytes. Biochem Biophys Res Commun 2003; 301:1045–50.
32. Unwin N, Shaw J, Zimmet P, et al. Impaired glucose tolerance and impaired fasting glycaemia: the current status on definition and intervention. Diabet Med 2002; 19:708–23.
33. National Diabetes Data Group. Classification and diagnosis of diabetes mellitus and other categories of glucose intolerance. Diabetes 1979;28:1039–57.
34. Santaguida PL, Balion C, Hunt D, et al. Diagnosis, prognosis and treatment of impaired glucose tolerance and impaired fasting glucose. Evid Rep Technol Assess
(Summ) 2005;128:1–11.
35. Snijder MB, Heine RJ, Seidell JC, et al. Associations of adiponectin levels with incident impaired glucose meta-bolism and type 2 diabetes in older men and women: the Hoorn study. Diabetes Care 2006;29:2498–503. 36. Tso AW, Sham PC, Wat NM, et al. Polymorphisms of
the gene encoding adiponectin and glycaemic outcome of Chinese subjects with impaired glucose tolerance: a 5-year follow-up study. Diabetologia 2006;49:1806–15.
37. Wannamethee SG, Lowe GD, Rumley A, et al. Adipo-kines and risk of type 2 diabetes in older men. Diabetes
Care 2007;30:1200–5.
38. Kanaya AM, Wassel Fyr C, Vittinghoff E, et al. Adipo-cytokines and incident diabetes mellitus in older adults: the independent effect of plasminogen activator inhibitor 1. Arch Intern Med 2006;166:350–6.
39. Panarotto D, Ardilouze JL, Tessier D, et al. The degree of hyperinsulinemia and impaired glucose tolerance predicts plasma leptin concentrations in women only: a new exploratory paradigm. Metabolism 2000;49: 1055–62.
40. Reaven GM. Pathophysiology of insulin resistance in human disease. Physiol Rev 1995;75:473–86.