CMU96-101
糖尿病誘發心臟細胞凋亡機轉與胰島素抗凋亡療效之探討
Effects of insulin replacement on cardiac apoptotic and survival pathways in streptozotocin-induced diabetic rats
Shin-Da Lee
Department of Physical Therapy, Graduate Institute of Rehabilitation Science, China Medical University, Taichung, Taiwan;
Supported by the China Medical University (CMU96-101)
Short Title: Insulin reverse the cardiac apoptosis in STZ-induced rats
*
Share equal contribution
Correspondence Author: Shin-Da Lee, PhD Department of Physical Therapy,
Graduate Institute of Rehabilitation Science College of Health Care
China Medical University 91 Hsueh-Shih Road,Taichung, Taichung, 40202, Taiwan TEL: 886-4-22053366 ext 7300 E-mail:[email protected]
Abstract
Background. Very limited information regarding cardiac apoptotic and survival pathways in diabetes mellitus (DM) was available. The purpose of this study was to evaluate the effects of insulin on cardiac apoptotic and survival pathways in streptozotocin (STZ)-induced diabetic rats. Methods. Forty-eight male Wistar rats at 8 weeks of age were randomly divided into control group with citrate buffer (Control), streptozotocin-induced (65 mg/kg i.v.) Type 1-like diabetic rats with citrate buffer (DM), and streptozotocin-induced diabetic rats with insulin replacement (DI) for 4 weeks and 8 weeks. The excised hearts were measured by Western blotting. Results. Cardiac mitochondrial-dependent apoptotic pathways, such as Bad, cytosolic cytochrome c, activated caspase 9 and activated caspase 3 and calcinerin-NFAT3 hypertrophic pathway in DM were significantly increased compared with the control group and attenuated back toward basal level in DI group after 8 weeks whereas those were not found after 4 weeks. Cardiac anti-apoptotic Bcl2 was significantly decreased in DM group not in DI group. Insulin-like growth factor-I receptor (IGF-IR), phosphatidylinositol 3’-kinase (PI3K) and the protein kinase B (Akt) were significantly decreased in DM relative to Control and DI after 8 weeks whereas those were not found after 4 weeks. Conclusions. Insulin replacement not only prevents STZ-induced cardiac mitochondrial-dependent apoptotic pathway activity and calcineurin-related hypertrophic pathway activity but also enhances IGF-IR-PI3K-Akt cardiac survival pathway activity, all of which are with insulin therapeutic duration-dependent manners. The findings may provide possible diabetes-enhanced apoptotic pathways for potentially preventing cardiac abnormality in diabetes.
Introduction
Patients with diabetes mellitus have an increased risk for cardiovascular diseases such as coronary heart disease and heart failure that contribute to decreased life expectancy (1). Diabetes, a common co-morbidity in patients with heart failure, is associated with worse long-term outcomes in patients with heart failure (2). The cellular apoptosis in terminally differentiated cardiomyocyte is a very critical pathological mechanism which causes heart failure and, on the other hand, the process of apoptotic interruptus may allow developing a novel strategy to reverse or attenuate heart failure (3). Cardiac cell death or cell apoptosis after myocardial infarction in experimental diabetes were roughly observed in diabetes (4) but cardiac apoptotic or survival pathways have not yet been investigated.
Apoptosis, a physiological program of cellular death, in heart may contribute to many cardiac disorders (5, 6). Cardiac apoptosis was found in many chronic cardiometablic diseases such as obesity (4, 7), diabetes, and hypertension (6, 8), and was also found under various stressors, such as long-term hypoxia (7, 9), or smoke (10). The occurrence of apoptosis has been reported to contribute to the loss of cardiomyocytes in cardiomyopathies, and is recognized as a predictor of adverse outcomes in subjects with cardiac diseases or heart failure (3). The mitochondrial-dependent apoptotic pathway is activated within the cell resulting in the release of a number of pro-apoptotic factors from the intermembrane space of mitochondria (11, 12). The mitochondria is the main site of action for members of the apoptosis-regulating protein family exemplified by Bcl-2 family, such as Bcl-2 and Bad (Bcl-2 antagonist of cell death) (11). Bcl-2, an anti-apoptotic protein, prevents cytochrome c release whereas Bad, a pro-apoptotic protein, enhances cytochrome c release from mitochondria (11). When cytochrome c is released from mitochondria
into cytosol, it is responsible for activating caspase-9, which further activates caspase-3 and executes the apoptotic program (13). However, it is unclear whether mitochondrial-dependent apoptotic pathway mediates diabetes-related cardiac apoptosis.
Calcineurin has been reported as a critical mediator for cardiac hypertrophy and cardiac myocyte apoptosis (14-16). Transgenic mice that express the activated forms of calcineurin in the hearts developed cardiac hypertrophy and heart failure that mimic human heart diseases (15). It has been reported that the activation of Ca2+ induces the calcineurin-nuclear factor activation transcription (NFAT) pathway and in turn enhances hypertrophy (15, 17). In addition, calcineurin has been reported to activate pro-apoptotic protein Bad and cause apoptosis of cardiomyocytes (18).
Insulin-like growth factor-I (IGF-I) signaling is reported to contribute to the modulation of survival and apoptosis responses in cardiac tissues (6, 19). IGF-I is a survival factor in cardiomyocytes that activates the phosphatidylinositol 3-kinase (PI 3-kinase) and protein kinase B (Akt) pathway via IGF-I receptor (20, 21). Impaired IGF-IR signaling may contribute, at least partially, to the development of hypertension and pathological cardiac apoptosis in hypertensive rats.
The current study was to understand whether mitochondrial-dependent apoptotic pathway, calineurin-Bad apoptotic pathway, calcinerin NFAT3 hypertrophic pathway, and impaired IGF-IR-PI3K-Akt in streptozotocin-induced diabetic rats are more activated and whether these pathways can be improved by insulin replacement. We hypothesized that first, diabetes may predispose to more activated cardiac mitochondrial-dependent apoptotic pathway, more activated calineurin-Bad apoptotic pathway, more activated calcinerin NFAT3 hypertrophic pathway, and impaired IGF-IR-PI3K-Akt survival pathway as well as second, insulin replacement may
prevent changes in these pathways in diabetes.
Materials and Methods
Animals and induction of diabetes
Forty-eight male Wistar rats at 8 weeks of age were obtained from National Laboratory Animal Center, Taiwan. Ambient temperature was maintained at 25°C and the animals were kept on an artificial 12-h light-dark cycle. The light period began at 7:00 A.M. Rats were provided with standard laboratory chow (Lab Diet 5001; PMI Nutrition International Inc., Brentwood, MO, USA) and water ad libitum. All experimental procedures were performed according to the NIH Guide for the Care and Use of Laboratory Animals and all protocols were approved by the Institutional Animal Care and Use Committee of China Medical University, Taichung, Taiwan.
All animals were allowed to adapt to the environment for 1 week after their arrival before the experiment started. All rats were divided into two groups, i.e. control group (n=16) and streptozotocin-injected group (n=32). The rats in streptozotocin-injected group were injected with streptozotocin (65 mg/kg body weight in citrate buffer, pH 4.5) and the rats in control group were injected with an equal volume of vehicle via the lateral tail vein. The rats were considered to be diabetic and used for the study if they had hyperglycemia (>15 mM) at 48 h after injection of streptozotocin as detected by Accu Soft (Hoffmann-La Roche) test strips. Three days later, the 16 of 32 streptozotocin-induced diabetic rats were treated subcutaneously with insulin 4U daily for 4 and 8 weeks. There are six groups in current study, i.e. control group with citrate buffer (Control-4W), streptozotocin-induced diabetic rats with citrate buffer (DM-4W), and streptozotocin-induced diabetic rats with insulin replacement (DI-4W) for 4 weeks as well as control group with citrate buffer (Control-8W), streptozotocin-induced
diabetic rats with citrate buffer (DM-8W), and streptozotocin-induced diabetic rats with insulin replacement (DI-8W) for 8 weeks.
Cardiac characteristics and heart weighting.
The hearts of 6 rats in six groups (i.e. Control-4W, DM-4W, DI-4W, Control-8W, DM-8W, and DI-8W) were analyzed by heart weight index and Western blotting. The hearts of animals were excised and cleaned with PBS. The left ventricles were separated and weighed. The ratios of the whole heart weight (WHW) to body weight (BW), the ratios of the left ventricular weight (LVW) to body weight (BW), the ratios of the left ventricle weight (LVW) to the whole heart weight (WHW) were calculated. Tissue Extraction
Cardiac tissue extracts were obtained by homogenizing the left ventricle samples in a lysis buffer (20mM Tris, 2mM EDTA, 50mM 2-mercaptoethanol, 10% glycerol, PH 7.4, proteinase inhibitor (Roche), phosphatase inhibitor cocktail (sigma)) at a ratio of 100 mg tissue/1ml buffer for 1 min. The homogenates were placed on ice for 10 min and then centrifuged at 12,000 g for 40 min twice. The supernatant was collected and stored at -70°C for further experiments.
Separation of cytosolic and mitochondrial fractions
To detect cytosolic cytochrome c, cardiac tissue extracts were suspended in a buffer (Tris, EDTA and proteinase inhibitor cocktail tablet (Roche)) for 1 min on ice, homogenized by Polytron, and centrifuged at 1,200 g for 10 min. The supernatant was re-centrifuged at 10,000 g for 15 min to collect the mitochondrion-enriched pellet and the supernatant as the cytosolic fraction. The pellet was resuspended in lysis buffer as the mitochondrial fraction.
Electrophoresis and Western Blot
method (Bio-Rad Protein Assay,Hercules,CA).Protein samples(50 μg/lane)were separated on a 10% SDS polyacrylamide gel electrophoresis (SDS-PAGE) with a constant voltage of 75 V. Electrophoresed proteins were transferred to polyvinylidene difluoride(PVDF)membrane(Millipore,Bedford,MA,0.45 μm poresize)with a transferring apparatus (Bio-rad). PVDF membranes were incubated in 5% milk in TBS buffer. Primary antibodies including Fas ligand, Fas receptor, FADD, Bad, Bax, caspase 8, caspase-9, caspase-3and α-tubulin (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and Bcl-2 (BD) were diluted to 1:500 in antibody binding buffer overnight at 4°C. The immunoblots were washed three times in TBS buffer for 10 min and then immersed in the second antibody solution containing goat anti-mouse IgG-HRP, goat anti-rabbit IgG-HRP, or donkey anti goat IgG-HRP (Santa Cruz) for 1 hour and diluted 500-fold in TBS buffer. The immunoblots were then washed in TBS buffer for 10 min three times. The immunoblotted proteins were visualized by using an enhanced chemiluminescence ECL Western blotting luminal Reagent (Santa Cruz, CA, USA) and quantified using a Fujifilm LAS-3000 chemiluminescence detection system (Tokyo, Japan). Densitometric analysis of immunoblots was performed by AlphaImager 2200 digital imaging system (Digital Imaging System, San Leandro, CA, USA).
Statistical Analysis
The data of whole heart weight index and protein levels were compared among groups of animals in six groups (Control-4W, DM-4W, DI-4W, Control-8W, DM-8W, and DI-8W) using one-way analysis of variance (ANOVA) with pre-planned contrast comparison in 4 weeks and 8 weeks. Control group serves as negative control group for DM groups and DM serves as non-therapeutic control group for therapeutic groups, DI. In all cases, a difference at P<0.05 was considered statistically significant.
Results:
Body weight and cardiac characteristics.
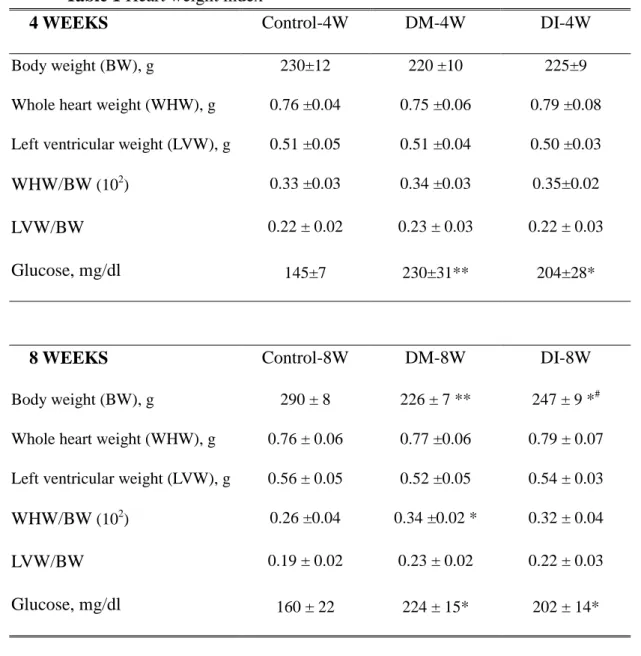
Body weight (BW), whole heart weight (WHW), left ventricular weight (LVW), the whole heart weight corrected by body weight (WHW/BW) and the left ventricle weight corrected by body weight (LVW/BW) were similar among Control-4W, DM-4W, and DI-4W groups. BW was decreased but WHW/BW was increased in DM-8W, compared with Control-8W. WHW, LVW, and LVW/BW were similar among Control-8W, DM-8W, and DI-8W groups (Table 1). However, blood glucose in DM-4W and DI-4W was significantly higher than in Control-4W and blood glucose in DM-8W and DI-8W was significantly higher than in Control-8W (Table 1).
Upstream components of cardiac mitochondrial-dependent apoptotic pathways To further understand the changes of the upstream components of mitochondrial-dependent apoptotic pathways in diabetes and diabetes treated with insulin, the protein levels of the Bcl-2 family (Bcl-2 and Bad) and cytosolic cytochrome c were measured in the excised hearts of six groups (Control-4W, DM-4W, DI-4W, Control-8W, DM-8W, and DI-8W) by Western blotting. Mitochondrial-related pro-apoptotic protein levels of Bad and cytosolic cytochrome c were significantly higher in DM-8W than Control-8W or DI-8W, but were similar among Control-4W, DM-4W, and DI-4W. Mitochondrial-related anti-apoptotic protein levels of Bcl2 in DM-8W were significantly lower than those in DI-8W or Control-8W, but were similar among Control-4W, DM-4W, and DI-4W (Fig 1).
Downstream components of cardiac mitochondrial-dependent apoptotic pathways To identify the changes of the downstream components of cardiac mitochondrial-dependent apoptotic pathways in diabetes and diabetes treated with insulin, the protein levels of activated caspase 9 and 3 were measured in the excised
hearts of six groups (Control-4W, DM-4W, DI-4W, Control-8W, DM-8W, and DI-8W) by western blotting. The activated forms of caspase 9 and 3 protein levels were significantly higher in DM-8W than those in Control-8W or DI-8W, but were similar among Control-4W, DM-4W, and DI-4W (Fig 2).
Calcineurin-NFAT3 hypertrophic pathway
To identify the activities of calcineurin-NFAT3 hypertrophic pathway in diabetes and diabetes treated with insulin, the protein levels of calcineurin and NFAT3 were measured in the excised hearts of six groups (Control-4W, DM-4W, DI-4W, Control-8W, DM-8W, and DI-8W) by Western blotting. The protein levels of calcineurin were significantly higher in DM-4W than those in Control-4W or DI-4W and were also significantly higher in DM-8W than those in Control-8W or DI-8W. The protein levels of NFAT3 were significantly higher in DM-8W than those in Control-8W or DI-8W, but were similar among Control-4W, DM-4W, and DI-4W (Fig 3).
IGFIR, PI3K, and p-Akt survival pathway
To identify insulin-like growth factor-I receptor (IGFIR), phosphatidylinositol 3’-kinase (PI3K) and phosphorylated Akt (p-Akt) survival pathway in diabetes and diabetes treated with insulin, the protein levels of IGFIR, PI3K, and p-Akt were measured in the excised hearts of six groups (Control-4W, DM-4W, DI-4W, Control-8W, DM-8W, and DI-8W) by Western blotting. The protein levels of IGFIR, PI3K, and p-Akt were similar among Control-4W, DM-4W, and DI-4W. The protein levels of IGFIR, PI3K, and p-Akt were significantly decreased in DM relative to control and these lower levels of IGFIR, PI3K, and p-Akt were significantly increased relative to DM (Fig 4).
Discussion:
Our main findings based on current results can be summarized as follows: (1) Body weight was decreased but the ratio of whole heart weight to body weight was increased after, compared with Control-8W. Changes in heart weight index were not found in STZ-induced diabetes and diabetes with insulin replacement for 4 weeks (2) The key coponents of cardiac mitochondrial-dependent apoptotic pathways, such as Bad, cytosolic cytochrome c, activated caspase 9 and activated caspase 3 in STZ-induced diabetes were significantly increased compared with control group and these STZ-induced mitochondrial-dependent apoptotic pathways were significantly attenuated after insulin replacement for 8 weeks not for 4 weeks. (3) Calcinerin-NFAT3 hypertrophic pathways in STZ-induced diabetes were significantly increased compared with Control group and these STZ-induced calcinerin-NFAT3 hypertrophic pathways were significantly attenuated after insulin replacement for 8 weeks. (4) Insulin-like growth factor-I receptor (IGF-IR), phosphatidylinositol 3’-kinase (PI3K) and the protein kinase B (Akt) were significantly decreased compared with control group and these STZ-induced impaired IGF-IR-PI3K-Akt pathways were improved after insulin replacement for 8 weeks not for 4 weeks. Our major conclusion showed that insulin replacement not only prevented STZ-induced cardiac mitochondrial-dependent apoptotic pathway and calcineurin-related hypertrophic pathway but also enhanced IGF-IR-PI3K-Akt cardiac survival pathway. The findings may provide possible diabetes-enhanced apoptotic pathways for potentially preventing cardiac abnormalities in diabetes.
Human type 1 and 2 diabetes mellitus, a multi-system disorder, results from failure of insulin production and insulin resistance, respectively. High single doses of streptozotocin, a genotoxic methylating agent that is targeted to the beta cells, cause
extensive beta-cell necrosis (22). The streptozotocin (STZ)-induced diabetic rats were often used as an animal model of type 1 diabetes mellitus (23). The STZ-induced diabetes (hyperglycemic) rats present many of the same pathophysiologic deficits as noted in diabetic humans, such as hypoinsulinemia, hyperglycemia, atherosclerosis, cardiac hypertrophy, cardiomyopathy, cardiovascular dysfunction, and finally resulting in heart failure (23-26). Diabetes predisposes to develop a multi-system disorder and had shown to be impacted by multiple factors, such as endocrine dysfunctions, metabolic disorders, oxidative stresses, inflammatory processes or immune systems (27, 28). Therefore, in the current experimental design, we have to add a cautious note that any deleterious effect of STZ-induced diabetes (hyperglycemic) or any preventive effect of insulin replacement on cardiac changes cannot be isolated to one specific factor or any specific system, but may be affected directly or indirectly by various factors, such as insulin, glucose, endocrine oxidative stress, inflammatory status, autoimmune or unclear interacting factors.
Diabetes is associated with a high incidence of cardiovascular disease, which is the major cause of morbidity and mortality (28). Left ventricular function was significantly impaired in diabetic animals (29). The presence of diabetes is a powerful risk factor for the development of heart failure (30). Individuals with diabetes mellitus have an increased risk of developing heart failure, usually as a consequence of coronary artery disease, although a specific diabetic cardiomyopathy, secondary to a microangiopathy, may also exist (31). Diabetes causes systolic heart failure in coronary artery disease prone patients but not patients with a higher myocardial infarction rate, because the coronary artery disease is often a microvascular impairment and it leads to heart failure rather than myocardial necrosis (32). Additively, diabetes is an independent risk factor for chronic heart failure, probably in
part due to disturbances in myocardial metabolism (33).
The mitochondrial-dependent apoptotic pathway is mediated by Bcl2 family, such as Bad and Bcl2 (34). Shifting the balance of Bcl2 family members toward pro-apoptotic effects will enhance cytochrome c release and will activate caspase-9, which further activates caspase-3 and executes the apoptotic program (13). Previous study shows increased apoptosis via increases in caspase-9 and caspase-3 activities in streptozotocin (STZ)-induced diabetic rat heart (35). After infarction, the numbers of apoptotic cells and cardiac caspase 3 were higher in the diabetic rats as compared to non-diabetic rats in the border zone of infarction and in non-infarcted area (36). In the current study, after 8 weeks but not 4 weeks, the mitochondrial-dependent apoptotic pathways were significantly activated in cardiac tissues in STZ-induced diabetic rats and the STZ-induced mitochondrial-dependent apoptotic pathways were suppressed with the dosage of 4 units insulin from a series of evidence, such as cardiac Bad, Bcl2, cytosolic cytochrome c, activated caspase-9, and activated caspase-3 levels. Therefore, our findings suggest that STZ-induced diabetes did cause more mitochondrial-dependent apoptosis as well as insulin replacement significantly attenuated diabetes-induced cardiac mitochondrial-dependent apoptosis and it is therapeutic duration dependent.
Calcineurin and its downstream transcriptional effector NFAT3 have been reported as a critical mediator for cardiac hypertrophy and cardiac myocyte apoptosis (14, 15). In addition, calcineurin will activate pro-apoptotic protein Bad and induce cytochrome c release from mitochondria to cause apoptosis of cardiomyocytes (18). Previous study had reported that cardiac hypertrophy was found in STZ-induced diabetic rats (37). This study represents the first to report that calcineurin-NFAT3 hypertrophic pathway and calcineurin-Bad related apoptotic pathway were activated
in STZ-induced diabetes and insulin replacement significantly reversed the deleterious effects of diabetes.
IGF-I/IGF-I receptor and its downstream PI3K and Akt signaling pathway are reported to contribute to the modulation of survival responses in cardiac tissue (20, 21). One previous study suggests that exogenous IGF-I treatment may ameliorate contractile disturbances in cardiomyocytes from diabetic animals and could provide therapeutic potential in the treatment of diabetic cardiomyopathy (38). This study represents the first to report that impaired IGF-IR-PI3K-Akt survival pathways were found in diabetic hearts and insulin replacement therapy significantly improved this impairment.
Hypothesized clinical application
Since cardiac tissues are difficult to be obtaied from diabetic patients, the findings of the current STZ-induced animal experiment should provide an important mechanism for explaining the apoptosis-related cardiac diseases in diabetes. If diabetes-related cardiac apoptosis and diabetes-related cardiac hypertrophy also occur in human, most of diabetic patients should be highly aware of the progressive development in cardiac abnormality and should actively promote heart health. The apoptotic and hypertrophic pathways might provide one possible mechanism to interrupt the development of heart failure and pathological cardiac hypertrophy. Of course, further clinical and experimental studies are required to clarify the apoptotic or hypertrophic mechanisms in human diabetic hearts and possible mechanisms between insulin and cardiac apoptosis and hypertrophy.
Reference
1. Sobel BE. Optimizing cardiovascular outcomes in diabetes mellitus. Am J Med. 2007;120:S3-11.
2. Masoudi FA, Inzucchi SE. Diabetes mellitus and heart failure: epidemiology, mechanisms, and pharmacotherapy. Am J Cardiol. 2007;99:113B-32B.
3. Narula J, Pandey P, Arbustini E, et al. Apoptosis in heart failure: release of cytochrome c from mitochondria and activation of caspase-3 in human cardiomyopathy. Proc Natl Acad Sci USA. 1999;96:8144-9.
4. Backlund T, Lakkisto P, Palojoki E, et al. Activation of protective and damaging components of the cardiac renin-angiotensin system after myocardial infarction in experimental diabetes. J Renin Angiotensin Aldosterone Syst. 2007;8:66-73.
5. Haunstetter A, Izumo S. Apoptosis: basic mechanisms and implications for cardiovascular disease. Circ Res. 1998;82:1111-29.
6. Lee SD, Chu CH, Huang EJ, et al. Roles of insulin-like growth factor II in cardiomyoblast apoptosis and in hypertensive rat heart with abdominal aorta ligation. Am J Physiol Endocrinol Metab. 2006;291:E306-14.
7. Lee SD, Kuo WW, Lin JA, et al. Effects of long-term intermittent hypoxia on mitochondrial and Fas death receptor dependent apoptotic pathways in rat hearts. Int J Cardiol. 2007.
8. Kuo WW, Liu CJ, Chen LM, et al. Cardiomyoblast apoptosis induced by insulin-like growth factor (IGF)-I resistance is IGF-II dependent and synergistically enhanced by angiotensin II. Apoptosis. 2006;11:1075-89. 9. Lee SD, Kuo WW, Wu CH, et al. Effects of short- and long-term hypobaric
hypoxia on Bcl2 family in rat heart. Int J Cardiol. 2006;108:376-84.
10. Kuo WW, Wu CH, Lee SD, et al. Second-hand smoke-induced cardiac fibrosis is related to the Fas death receptor apoptotic pathway without mitochondria-dependent pathway involvement in rats. Environ Health Perspect. 2005;113:1349-53.
11. Bishopric NH, Andreka P, Slepak T, Webster KA. Molecular mechanisms of apoptosis in the cardiac myocyte. Curr Opin Pharmacol. 2001;1:141-50. 12. Barnhart BC, Alappat EC, Peter ME. The CD95 type I/type II model. Semin
Immunol. 2003;15:185-93.
13. Brown GC, Borutaite V. Nitric oxide, cytochrome c and mitochondria. Biochem Soc Symp. 1999;66:17-25.
14. Wilkins BJ, Molkentin JD. Calcineurin and cardiac hypertrophy: where have we been? Where are we going? J Physiol. 2002;541:1-8.
pathway for cardiac hypertrophy. Cell. 1998;93:215-28.
16. Lakshmikuttyamma A, Selvakumar P, Kakkar R, et al. Activation of calcineurin expression in ischemia-reperfused rat heart and in human ischemic myocardium. J Cell Biochem. 2003;90:987-97.
17. Yue TL, Ohlstein EH, Ruffolo RR, Jr. Apoptosis: a potential target for discovering novel therapies for cardiovascular diseases. Curr Opin Chem Biol. 1999;3:474-80.
18. Molkentin JD. Calcineurin, mitochondrial membrane potential, and cardiomyocyte apoptosis. Circ Res. 2001;88:1220-2.
19. Kuo WW, Chu CY, Wu CH, et al. Impaired IGF-I signalling of hypertrophic hearts in the developmental phase of hypertension in genetically hypertensive rats. Cell Biochem Funct. 2005;23:325-31.
20. Sun HY, Zhao RR, Zhi JM. Insulin-like growth factor I inhibits cardiomyocyte apoptosis and the underlying signal transduction pathways. Methods Find Exp Clin Pharmacol. 2000;22:601-7.
21. Parrizas M, Saltiel AR, LeRoith D. Insulin-like growth factor 1 inhibits apoptosis using the phosphatidylinositol 3'-kinase and mitogen-activated protein kinase pathways. J Biol Chem. 1997;272:154-61.
22. Cardinal JW, Margison GP, Mynett KJ, et al. Increased susceptibility to streptozotocin-induced beta-cell apoptosis and delayed autoimmune diabetes in alkylpurine-DNA-N-glycosylase-deficient mice. Mol Cell Biol. 2001;21:5605-13.
23. Seemayer TA, Oligny LL, Tannenbaum GS, Goldman H, Colle E. Animal model of human disease. Diabetes mellitus. Am J Pathol. 1980;101:485-8. 24. De Angelis K, Schaan BD, Maeda CY, et al. Cardiovascular control in
experimental diabetes. Braz J Med Biol Res. 2002;35:1091-100.
25. Howarth FC, Al-Shamsi N, Al-Qaydi M, et al. Effects of brain natriuretic peptide on contraction and intracellular Ca2+ in ventricular myocytes from the streptozotocin-induced diabetic rat. Ann N Y Acad Sci. 2006;1084:155-65. 26. Shiomi T, Tsutsui H, Ikeuchi M, et al. Streptozotocin-induced hyperglycemia
exacerbates left ventricular remodeling and failure after experimental myocardial infarction. J Am Coll Cardiol. 2003;42:165-72.
27. Chbinou N, Frenette J. Insulin-dependent diabetes impairs the inflammatory response and delays angiogenesis following Achilles tendon injury. Am J Physiol Regul Integr Comp Physiol. 2004;286:R952-7.
28. Lopes-Virella MF, Virella G. The role of immune and inflammatory processes in the development of macrovascular disease in diabetes. Front Biosci. 2003;8:s750-68.
29. Westermann D, Van Linthout S, Dhayat S, et al. Cardioprotective and anti-inflammatory effects of interleukin converting enzyme inhibition in experimental diabetic cardiomyopathy. Diabetes. 2007;56:1834-41.
30. Estep JD, Aguilar D. Diabetes and heart failure in the post-myocardial infarction patient. Curr Heart Fail Rep. 2006;3:164-9.
31. Macfarlane DP, Fisher M. Thiazolidinediones in patients with diabetes mellitus and heart failure : implications of emerging data. Am J Cardiovasc Drugs. 2006;6:297-304.
32. Palmiero P, Macello M, De Pascalis S. Heart failure and diabetes: left ventricular systolic function. Minerva Cardioangiol. 2006;54:241-8.
33. Thrainsdottir I, Malmberg K, Olsson A, Gutniak M, Ryden L. Initial experience with GLP-1 treatment on metabolic control and myocardial function in patients with type 2 diabetes mellitus and heart failure. Diab Vasc Dis Res. 2004;1:40-3.
34. Kubasiak LA, Hernandez OM, Bishopric NH, Webster KA. Hypoxia and acidosis activate cardiac myocyte death through the Bcl-2 family protein BNIP3. Proc Natl Acad Sci USA. 2002;99:12825-30.
35. Ghosh S, Pulinilkunnil T, Yuen G, et al. Cardiomyocyte apoptosis induced by short-term diabetes requires mitochondrial GSH depletion. Am J Physiol Heart Circ Physiol. 2005;289:H768-76.
36. Backlund T, Palojoki E, Saraste A, et al. Sustained cardiomyocyte apoptosis and left ventricular remodelling after myocardial infarction in experimental diabetes. Diabetologia. 2004;47:325-30.
37. Chang KC, Hsu KL, Tseng CD, et al. Aminoguanidine prevents arterial stiffening and cardiac hypertrophy in streptozotocin-induced diabetes in rats. Br J Pharmacol. 2006;147:944-50.
38. Norby FL, Wold LE, Duan J, Hintz KK, Ren J. IGF-I attenuates diabetes-induced cardiac contractile dysfunction in ventricular myocytes. Am J Physiol Endocrinol Metab. 2002;283:E658-66.
Table 1 Heart weight index
4 WEEKS Control-4W DM-4W DI-4W
Body weight (BW), g 230±12 220 ±10 225±9
Whole heart weight (WHW), g 0.76 ±0.04 0.75 ±0.06 0.79 ±0.08
Left ventricular weight (LVW), g 0.51 ±0.05 0.51 ±0.04 0.50 ±0.03
WHW/BW(102) 0.33 ±0.03 0.34 ±0.03 0.35±0.02
LVW/BW 0.22 ± 0.02 0.23 ± 0.03 0.22 ± 0.03
Glucose, mg/dl 145±7 230±31** 204±28*
8 WEEKS Control-8W DM-8W DI-8W
Body weight (BW), g 290 ± 8 226 ± 7 ** 247 ± 9 *#
Whole heart weight (WHW), g 0.76 ± 0.06 0.77 ±0.06 0.79 ± 0.07
Left ventricular weight (LVW), g 0.56 ± 0.05 0.52 ±0.05 0.54 ± 0.03
WHW/BW(102) 0.26 ±0.04 0.34 ±0.02 * 0.32 ± 0.04
LVW/BW 0.19 ± 0.02 0.23 ± 0.02 0.22 ± 0.03
Glucose, mg/dl 160 ± 22 224 ± 15* 202 ± 14*
Values are means± SEM (n=6 in each group). Body weight and Heart weight index in Wistar rats with citrate buffer (Control), streptozotocin-induced diabetic rats with citrate buffer (DM), and streptozotocin-induced diabetic rats with insulin replacement (DI) for 4 weeks and 8 weeks. * P<0.05, ** P<0.01, significant differences vs. control group.#P<0.05, significant differences vs. DM group.
Figure legend Fig 1.
(A) the representative protein levels of Bcl2, Bad, and cytosolic cytochrome c extracted from the left ventricles of excised hearts in Wistar rats with citrate buffer (Control, C), streptozotocin-induced diabetic rats with citrate buffer (DM), and streptozotocin-induced diabetic rats with insulin replacement (DI) for 4 weeks and 8 weeks were measured by Western blotting analysis. (B)(C)(D) Bars represent the relative protein quantification of Bcl2, Bad, and cytosolic cytochrome c on the basis ofα-tubulin, and indicate mean values±SD (n=6 in each group). *P<0.05, **P<0.01, significant differences from control group. #P<0.05, ##P<0.01, significant differences from STZ group.
Fig 2.
(A) the representative protein levels of activated caspase-3, activated caspase-8 and activated caspase-9 extracted from the left ventricles of excised hearts in Wistar rats with citrate buffer (Control, C), streptozotocin-induced diabetic rats with citrate buffer (DM), and streptozotocin-induced diabetic rats with insulin replacement (DI) for 4 weeks and 8 weeks were measured by Western blotting analysis. (B)(C)(D) Bars represent the relative protein quantification of activated caspase-8, activated caspase-9 and activated caspase-3on thebasisofα-tubulin, and indicate mean values±SD (n=6 in each group). *P<0.05, **P<0.01, significant differences from control group. #P<0.05, ##P<0.01, significant differences from STZ group.
Fig 3.
transcription 3 (NFAT3) extracted from the left ventricles of excised hearts in Wistar rats with citrate buffer (Control, C), streptozotocin-induced diabetic rats with citrate buffer (DM), and streptozotocin-induced diabetic rats with insulin replacement (DI) for 4 weeks and 8 weeks were measured by Western blotting analysis. (B)(C) Bars represent the relative protein quantification of calcinerin and NFAT3 on the basis of α-tubulin, and indicate mean values±SD (n=6 in each group). *P<0.05, **P<0.01, significant differences from control group. #P<0.05, ##P<0.01, significant differences from STZ group.
Fig 4.
(A) the representative protein levels of insulin-like growth factor-I receptor (IGFIR), phosphatidylinositol3’-kinase (PI3K) and phosphorylated Akt (p-Akt) extracted from the left ventricles of excised hearts in Wistar rats with citrate buffer (Control, C), streptozotocin-induced diabetic rats with citrate buffer (DM), and streptozotocin-induced diabetic rats with insulin replacement (DI) for 4 weeks and 8 weeks were measured by Western blotting analysis. (B)(C) Bars represent the relative protein quantification of activated caspase-8, activated caspase-9 and activated caspase-3on thebasisofα-tubulin, and indicate mean values±SD (n=6 in each group). *P<0.05, significant differences from control group. #P<0.05, significant differences from STZ group.
Fig 1 (A) (B) (C) 0 1 2 3 C DM DI C DM DI B cl -2 / α -t ub ul in ** ## 4W 8W 0 2 4 6 8 C DM DI C DM DI B ad / α-tu bu li n ** ## 4W 8W (D) 0 1 2 3 4 C DM DI C DM DI C y t c / α-tu bu li n ** ## 4W 8W Bcl2 Bad Cyt c α-tubulin 4W 8W C DM DI C DM DI
Fig 2 (A) (B) (C) 0 1 2 3 4 C DM DI C DM DI C as pa se 9 / α-tu bu lin ** ## 4W 8W 0 1 2 3 4 C DM DI C DM DI C as pa se 3 / α-tu bu li n * # 4W 8W Caspase 9 Caspase 3 α-tubulin 4W 8W C DM DI C DM DI
Fig 3 (A) (B) (C) 0 1 2 3 4 C DM DI C DM DI C al ci ne ur in / α-tu bu li n * # 4W 8W ** 0 2 4 6 C DM DI C DM DI N F A T 3 / α -t ub ul in ** ## 4W 8W Calcineurin NFAT3 α-tubulin 4W 8W C DM DI C DM DI
Fig 4 (A) (B) 0 1 2 3 C DM DI C DM DI IG F -I R / α-tu bu lin 4W 8W * # 0 1 2 3 C DM DI C DM DI P I3 K / α-tu bu lin 4W 8W * 0 1 2 3 C DM DI C DM DI p-A kt / α-tu bu li n 4W 8W * # 4W 8W C DM DI C DM DI IGF-IR PI3K p-Akt α-tubulin
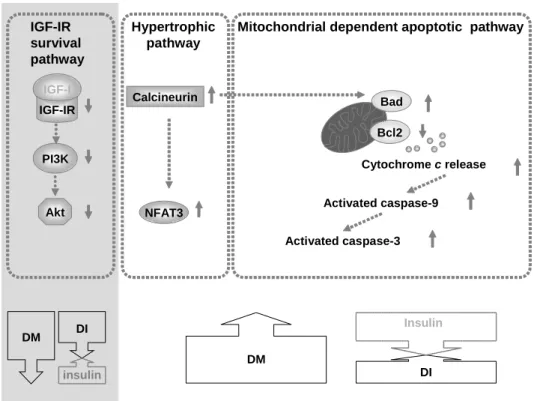
Fig 5 IGF-I IGF-IR PI3K Akt Bcl2 Bad Activated caspase-9 Cytochrome c release Activated caspase-3
Mitochondrial dependent apoptotic pathway IGF-IR survival pathway DM DI Insulin Calcineurin NFAT3 Hypertrophic pathway DM DI insulin
Fig. 5 Our proposed hypothesis that cardiac mitochondrial-dependent apoptotic pathway may be more activated in diabetes due to decreased anti-apoptotic Bcl2, increased pro-apoptotic Bad, increased cytochrome c release, increased activated caspase-9, and increased caspase-3 (right upper column). Up arrows and down arrows on the right side represent increases and decreases, respectively. These STZ-induced mitochondrial-dependent apoptotic pathways were significantly attenuated after insulin replacement (right lower arrow). Calcinerin-NFAT3 hypertrophic pathways in diabetes may be more activated in diabetes and these STZ-induced calcinerin-NFAT3 hypertrophic pathways were significantly attenuated after insulin replacement (middle upper column). Insulin-like growth factor-I receptor (IGF-IR), phosphatidylinositol 3’-kinase (PI3K) and the protein kinase B (Akt) may be more activated in diabetes and these STZ-induced impaired IGF-IR-PI3K-Akt pathways were improved after insulin replacement (right column).