PPAR-δ促效劑GW501516對疫苗反應之篩選生發中心B細胞和抗體製造之影響
121
0
0
全文
(2) i.
(3) /. B. / 4 8. : N. 1. . :. 2:. 5. 5 9. 9. 8. N. 、 4. 1. :. 8. .. .. 9. N 。. 8. :. 8. 9: 8. :. ii. 9.
(4) :. FcγRIIB B. IgG. 9. 。. : Fc. 。. FcγRIIB. SHIP. FcγRIIB. B. 。. c-Abl FcγRIIB. : :. ,. FcγRIIB. 。 FcγRIIB. N 。 B. FcγRIIB. 。. FcγRIIB. 。. c-Abl. .. 。 。. BCR FcγRIIB/c-Abl. B. N. 。. 1. C57BL/6. NP(4-hydroxy-3-nitrophenylacetyl)- CGG. (chicken gamma globulin) nilotinib . 2. /. P. (). 0P. 。 NP. IgG. IgG. 。 iii. 。. N B. FcγRIIB. .. 。. 。. 。 GW501516. B. B. 。. 。. PPAR-δ. B. 8. c-Abl.
(5) B GW501516. 。. 。. FcγRIIB GW501516 3. B. 。. N 6. P. N. FcγRIIB. B. 。 NP FcγRIIB/c-Abl. A. FcγRIIB. PPAR-δ. GW501516. iv. .. nilotinib. nilotinib.
(6) ABSTRACT FcγRIIB is an inhibitory Fc receptor of IgG and an essential negative regulator of B cells. In response to cognate and non-cognate immune complexes (ICs), FcγRIIB can transduce SHIP-dependent and c-Abl-dependent signaling pathways, respectively, in a mutually exclusive manner. Mice deficient in FcγRIIB display an enhanced humoral immune. response. after. immunization.. Moreover,. FcγRIIB. knockout. mice. spontaneously develop a lupus-like disease as a result of uncontrolled expansion of plasma cells (PCs) and thereby excess generation of ICs that accumulate in tissues, leading to glomerulonephritis. Thus, FcγRIIB is considered to play a checkpoint role in regulating the homeostasis of PC numbers. Because PCs are terminally differentiated B cells and they originate from germinal center (GC) B cells, we investigate the functional role of FcγRIIB in the selection of GC B cells for further differentiation into PCs after secondary immunization, and pharmacological modulation with GW501516, a PPAR-δ agonist, on this election process, known as affinity maturation to generate antibody (Ab) of high affinity to antigen (Ag). Because FcγRIIB/c-Abl signals for apoptosis of B cells and GC B cells of which B-cell receptor (BCR) binds to the Ag with low affinity will be. v.
(7) eliminated by apoptosis, we hypothesized that inhibition of FcγRIIB/c-Abl signaling during affinity maturation could alter the selection outcome and Ab affinity. We immunized C57BL/6 mice with a model Ag NP-CGG (4-hydroxy-3-nitrophenylacetyl hapten-chicken gamma globulin) and treated mice with nilotinib (2 mg/kg/day), a clinically used c-Abl inhibitor, during the seventh to ninth days when GC B cells undergo apoptosis for selection. Indeed, we found an increase of serum NP+ IgGs accompanied by an increased number of low-affinity NP+IgG+ PCs, indicating reduced affinity maturation. Previously, GW501516 was shown to down-regulate the expression level of FcγRIIB. When GW501516 (3,6 mg/kg/day) was administered intraperitoneally to mice during the peak of selection of GC B cells, we found a similar phenotype of affinity maturation. Taken together, we propose that FcγRIIB plays a checkpoint role in the negative selection of low-affinity GC B cells to ensure the production of high-affinity Abs for humoral host protection. Moreover, our preliminary data showed cross-reactivity of low-affinity Abs to cognate Ags. Thus, it will be of interest to examine whether modulation of inhibitory activity of FcγRIIB could enhance cross-protection against distinct influenza A virus by seasonal influenza vaccination. Key words: FcγRIIB germinal center PPRA-δ GW501516 nilotinib cross-protection vi.
(8) CONTENTS .................................................................................................................... i ............................................................................................................................................ ii : .................................................................................................................................. iii ABSTRACT .............................................................................................................................. v LIST of FIGURES ................................................................................................................... x LIST of ABBREVIATION ................................................................................................... xi Chapter 1 Introduction .......................................................................................................... 1 1.1 Germinal center (GC) reaction ............................................................................................... 2 1.1.1 Anatomy and function of germinal center .................................................................................. 2 1.1.2 Somatic Hypermutation .................................................................................................................... 3 1.1.3 Affinity maturation ............................................................................................................................ 4 1.1.4 Class switch recombination ............................................................................................................. 6 1.2 Hapten carrier system as a model for vaccination response ............................................ 6 1.3 Fcγ Receptors .............................................................................................................................. 8 1.4 Fcγ receptor IIB (FcγRIIB, a.k.a. CD32B) ........................................................................ 10 1.4.1 General Properties ............................................................................................................................ 10 1.4.2 Physiological function and signal transduction of FcγRIIB ............................................... 11 1.4.3 FcγRIIB-I232T polymorphism ..................................................................................................... 12 1.5 Peroxisome proliferator activated receptors (PPARs) ................................................... 14 1.6 PPARδ ....................................................................................................................................... 15 1.7 GW501516 ................................................................................................................................ 16 1.8 Nilotinib .................................................................................................................................... 17 1.9 Motivation ................................................................................................................................ 19. Chapter 2 Materials and Methods .................................................................................... 20 2.1 Mice ............................................................................................................................................ 21. 2.2 Enzyme Linked Immunosorbent spot (ELISPOT) assay ................................. 21 2.3 Enzyme Linked Immunosorbent assay (ELISA) ............................................................. 22 2.4 Confocal microscopy .............................................................................................................. 23 vii.
(9) Chapter 3 Results ................................................................................................................. 25 3.1 FcγRIIB232T/T mice show persistent impaired affinity maturation over time ............ 26 3.2 Association of FcγRIIB and BCR with the lipid raft after coligation was impaired in FcγRIIB 232T/T mutant mice ......................................................................................................... 27 3.3 Nilotinib administration during GC reaction has a negative impact on affinity maturation ...................................................................................................................................... 29 3.4 GW501516 increases respective total and low-affinity Ag-specific IgG PCs at the dose of 3 and 6 mg/kg/day during GC reaction ...................................................................... 30 3.5 GW501516 increases the serum levels of total and low-affinity Ag-specific IgG at the dose of 3 and 6 mg/kg/day in wild-type mice ........................................................................... 32 3.6 GW501516 increases total Ag-specific IgM secreting PCs at the dose of 6 mg/kg/day during GC reaction ....................................................................................................................... 33 3.7 GW501516 increases the serum levels of high-affinity Ag-specific IgM at the dose of 3 and 6 mg/kg/day and total Ag-specific IgM at 6 mg/kg/day ............................................. 35. Chapter 4 Discussion ........................................................................................................... 36 4.1 FcγRIIB232T/T mutant mice as a model for investigating the functional role of FcγRIIB in GC reaction ............................................................................................................... 37 4.2 Difference between FcγRIIB 232T/T mutant mice and wild type mice treated with nilotinib or GW501516 ................................................................................................................. 38 4.3 Summary of the overall effect of GW501516 on GC reaction ...................................... 39 4.4 Could GW501516 affect GC reaction other than FcγRIIB down-regulation induced by PPAR-δ activation? ................................................................................................................. 40 4.5 Pharmacological modulation of FcγRIIB activity is an applicable method for novel manipulation of immune response ............................................................................................. 43 4.6 Potential role of macrophages in the GC reaction to influence the generation of Ag-specific Abs ............................................................................................................................... 44. Figures .................................................................................................................................... 47 References .............................................................................................................................. 65 Appendix ................................................................................................................................ 71 Publication List .............................................................................................................................. 72 Publications .................................................................................................................................... 73 The lupus-associated FcγRIIB-I232T polymorphism results in impairment in the negative viii.
(10) selection of low-affinity germinal center B cells via c-Abl in mice ........................................... 74 B-cell ELISpot assay to quantify antigen-specific antibody-secreting cells in human peripheral blood mononuclear cells ....................................................................................................... 87 Dual immuno-renal targeting of 7-benzylidenenaltrexone alleviates lupus nephritis via FcγRIIB and HO-1 ...................................................................................................................................... 96. ix.
(11) LIST of FIGURES Figure 1 Mouse immunization schedules. .................................................................................... 48 Figure 2 Analysis of the differences of affinity maturation status between wild-type and FcγRIIB 232T/T mice over time. .......................................................................................................... 50 Figure 3 Analysis of the differences of affinity maturation status between wild-type and FcγRIIB 232T/T mice over time. .......................................................................................................... 51 Figure 4 Analysis of the effect of nilotinib on the affinity maturation of female wild-type mice. ....................................................................................................................................... 53 Figure 5 Analyzing IgG-secreting PCs in the spleen of female C57BL/6 mice immunized with two different GW501516 doses by ELISPOT assay. ................................ 55 Figure 6 Measurement of serum IgG levels of female C57BL/6 mice immunized with two different GW501516 doses by ELISA assay. ...................................................................... 58 Figure 7 Analyzing IgM-secreting PCs in the spleen of female C57BL/6 mice immunized with two different GW501516 doses by ELISPOT assay ................................. 60 Figure 8 Measurement of the serum IgM levels of female C57BL/6 mice immunized with two different GW501516 doses by ELISA assay .............................................................. 63. x.
(12) LIST of ABBREVIATION. 15-HETE. 15-hydroxyeicosatetraenoic acid. 4-HNE. 4-hydroxynonenal. Ab. Antibody. AEC. 3-amino-9-ethylcarbazole. AF-1. Activation function-1. Ag. Antigen. AID. Activation-induced cytidine deaminase. AMPK. AMP-activated protein kinase. Bcl-6. B-cell lymphoma 6. BCR. B-cell receptor. BCR-Abl. Breakpoint cluster region-Abelson tyrosine kinase . Blimp-1. B lymphocyte-induced maturation protein-1. CML. Chronic myelogenous leukemia. CSR. Class switch recombination . CXCL12. C-X-C motif chemokine 12. CXCL13. C-X-C motif chemokine 13. CXCR4. C-X-C chemokine receptor 4. CXCR5. C-X-C chemokine receptor 5. DMSO. Dimethyl sulfoxide . DZ. Dark zone. ELISA. Enzyme Linked Immunosorbent assay . ELISPOT. Enzyme Linked Immunosorbent spot assay. FACS. Fluorescence-assisted cell sorting. FDA. Food and Drug Administration. GC. Germinal center. GPI. Glycosylphosphatidyl inositol. HBV. Hepatitis B virus. xi.
(13) HODE. 9-hydroxyoctadecadienoic acid. HRP. Horseradish peroxidase. ICAM-1. Intercellular adhesion molecule 1 . Ics. Immune complexes. Ig. Immunoglobulin. IRF-4. Interferon regulatory protein-4 . ITAMs. Immunorepector tyrosine-based activating motifs. ITIM. Immunoreceptor tyrosine-based inhibitory motif. LZ. Light zone. MMM. Marginal metallophilic macrophage. MZM. Marginal zone macrophage. NIP. 4-Hydroxy-5-iodo-3-nitrophenylacetic acid. NP. 4-hydroxy-3 nitrophenylacetic acid. NP-CGG. 4-hydroxy-3-nitrophenylacetyl hapten-chicken gamma globulin. PBS-T. Phosphate buffered saline with 0.05% Tween 20. PCs. Plasma cells. Ph+. Philadelphia chromosome positive. PPARs. Peroxisome proliferator activated receptors. PPAR-δ. Peroxisome proliferator activated receptor delta. PPRE. PPAR response element . RBCs. Red blood cells. RXR. Retinoid X receptor. SHIP. SH2-containing inositol 5′-Phosphatase . SHM. Somatic hypermutation. SLAM. Signaling lymphocyte activation molecule. SNP. Single nucleotide polymorphism. TBM. Tingible body macrophage. xii.
(14) . Chapter 1 Introduction. 1.
(15) Chapter 1 Introduction 1.1 Germinal center (GC) reaction 1.1.1 Anatomy and function of germinal center The primary follicle is a B cell-rich structure, which locates in the secondary lymphoid organs, including, the spleen, lymph nodes, Peyer’s patches of the intestine and tonsils. In response to antigen (Ag) stimulation, B cells in the primary follicle proliferate and form the secondary follicle, which comprises of the marginal zone, the mantle zone and the germinal center (GC). The GC localizes in the central portion of the secondary follicle and contacts with the T cell zone (Victoria and Nussenzweig, 2012). In the GC, Ag-stimulated B cells sequentially undergo somatic hypermutation, affinity maturation and class switching. Based on the hematoxylin-eosin staining of nuclear contents, it was well established that the GC comprises the dark zone (DZ) and the light zone (LZ), both of which are filled with GC B cells differentiated in different stages, and also called centroblasts and centrocytes, respectively. Histologically, the DZ is densely packed with proliferating centroblasts (Allen, et al., 2015). In contrast, the LZ is mostly composed of centrocytes and follicular dendritic cells that can trap. 2.
(16) immune complexes (ICs) for presentation to B cells. In the DZ, the Ag-stimulated centroblasts rapidly proliferate and undergo somatic hypermutation in the variable region of immunoglobulin (Ig) gene to increase the repertoire of mutated B-cell receptor (BCR) on GC B cells for selection. The centroblasts later cease proliferation and migrate to the LZ to differentiate into centrocytes for affinity maturation for selection, followed by class switching of Ig isotypes (Zhang, et al., 2016).. 1.1.2 Somatic Hypermutation The immature B cells undergo the V(D)J gene recombination to diversify the surface B cell receptor and further mature into naïve B cell. It is process completed in the bone marrow. Somatic hypermutation (SHM) occurs to the rapid proliferating GC B cells, derived from antigen-stimulated naïve B cells. It is a tightly regulated mechanism that the genes of the B-cell receptor (BCR) acquire a higher-than-usual mutation rate during proliferation to expand its repertoire. The mutation rate is estimated about 103 to 105 per base pair, about 106 folds greater than other somatic cells’ proliferation (Martin, et al., 2015). The mutations of BCR are mostly point mutations and are left unaltered to. 3.
(17) accumulate in the variable region of the Ig genes for increasing the Ab diversity. The key enzymes that contribute to SHM include activation-induced cytidine deaminase (AID), uracil N-glycosylase, and error-prone DNA-polymerase. Since SHM is a random process, the majority of the mutations will lead to decreased Ab affinity (Kanyavuz, et al., 2019). Only a small portion of the centroblasts that acquire higher Ag binding affinity after SHM will survive owing to BCR signals.. 1.1.3 Affinity maturation The affinity maturation is a process to select high-affinity centrocytes as a result of success in competing limited Ag on follicular dendritic cells for survival and proliferation (Zhang, et al., 2016). High-affinity GC B cells induce stronger CD40 and BCR signals that enhance c-Myc, interferon regulatory protein-4 (IRF-4) expression levels. IRF-4 upregulates the expression of B lymphocyte-induced maturation protein-1 (Blimp-1), a transcriptional factor that ceases proliferation, induces plasma cell differentiation, and increases antibody synthesis and secretion. CD40 activation leads to higher expression levels of intercellular adhesion molecule 1 (ICAM-1) and signaling. 4.
(18) lymphocyte activation molecule (SLAM), both of which are adhesion molecules that can enhance interaction and adhesion between the T follicular helper cells (Tfh) and centrocytes during affinity maturation and class switching (Shulman, et al, 2013). The current paradigm is that centrocytes with high affinity BCRs promote more signal help from the Tfh for survival and plasma cell (PC) differentiation. In contrast, low-affinity centrocytes that cannot receive Ag-triggered survival signals are doomed to be eliminated by apoptosis or survive as memory B cells (Wataru and Tomohiro, 2018). The organization and migration between the DZ and the LZ are highly dynamic and require chemokine receptor expression and chemokine gradients to direct the destination of GC B cells in the GC. Namely, the centroblasts express higher C-X-C chemokine receptor type 4 (CXCR4) levels and respond to C-X-C motif chemokine ligand 12 (CXCL12) produced in the DZ, while the centrocytes express abundant CXCR5 and migrate to CXCL13 gradients established in the LZ. Recent studies have shown that some positively selected centrocytes with intermediate Ag binding affinity will reenter the DZ for further proliferation and SHM in order to obtain higher affinity to Ag (De Silva and Klein, 2015).. 5.
(19) 1.1.4 Class switch recombination The variable region of an antibody determines its Ag binding affinity. On the other hand, the constant region defines the class (isotype) of the antibody, and its distinct function and distribution. The naïve B cells that express IgM or IgD on the surface will undergo class switch recombination (CSR) to transform into IgG, IgA or IgE (Stavnezer, et al., 2008). It is a process that determine by the type of helper T cells and cytokines, with which the B cells interact. In the GC, Tfh interact with the GC B cells at the interface of the GC and T cell zone for CSR. Nevertheless, triggered by Th1 and Th2 cells, CSR can also take place in extrafollicular regions. In addition to the involvement in SHM, AID is the major enzyme of initiation of CSR (Chua, et al., 2002).. 1.2 Hapten carrier system as a model for vaccination response Vaccination is a clinical procedure and active immunity aimed to enhance host immunity by artificially exposing antigens of target pathogens. The vaccine antigen (Ag) can be live attenuated, inactivated or subunits of pathogens. It is a cost-effective method. 6.
(20) for individuals to generate effective and protective immune responses prepared for encountering the target pathogens (WHO, 2017). When pathogens invade the host, the primary humoral response is slow, inefficient and less in scale. Nevertheless, after the primary response fades away the pathogen-specific memory B and T cells will be generated and stay long term. Because of the presence of memory cells, a quicker and greater secondary immune response can occur as the individual encounters the original or similar pathogens (Kurosaki, et al., 2015). Taking advantage of the memory response of the adaptive immunity, we can generate specific immune memory for better protection by vaccination. Hapten carrier system is a widely used model for research on T-cell-dependent response of vaccination. T-cell-dependent response occurs in the GC. Haptens are small molecules that do not elicit immune response alone but they become immunogenic when they are conjugated with a carrier protein to increase molecular weight. The valence of the hapten-carrier protein is defined as the average number of the haptens that are able to bind to the carrier protein (Oda, et al., 2004). Commonly used carrier proteins include albumin, ovalbumin, gamma globulin, etc. The haptens modulate the. 7.
(21) amino acid epitopes and trigger T-cell-dependent adaptive immunity against the hapten, hapten-carrier epitopes or the carrier after immunization (Gefen, et al., 2015). The GCs induced by immunization undergo the affinity maturation to select GC B cells that carry B-cell receptors with higher Ag affinity, meaning that cells are able to bind against hapten-carrier protein with lower valence. As a result, by detecting the percentage of antibodies that can bind against the hapten-carrier protein with lower or higher valence than the immunogen, researchers can trace the affinity maturation status of vaccination. The fundamental goal of vaccination is to generate antibodies with higher affinity to target Ag for humoral host protection.. 1.3 Fcγ Receptors Fcγ receptors are a class of Fc receptors that bind the constant region of immunoglobulin G (IgG) to function. Different types of Fcγ receptors show different affinity to IgG isotypes, and express on different immune cells. Among the Fcγ receptors, only the FcγRI, an activating receptor, binds the monomeric form of IgGs. FcγRIIB is the only inhibitory receptor in the Fcγ receptor family and it possesses weak. 8.
(22) affinity to IgG in mice and humans. In humans, the other Fcγ receptors are classified into FcγRIIA, FcγRIIC, FcγRIIIA, and FcγRIIIB. In mice, other activating Fcγ receptors are FcγRIII and FcγRIV. With the exception of FcγRI, which have three Ig-like domains in the extracellular part, all the Fcγ receptors have only two Ig-like domains, resulting in weak to medium affinity for IgG and only binding to IgG immune complex (IC). Activating Fcγ receptors of humans contain immunorepector tyrosine-based activating motifs (ITAMs) on either the intracellular domain itself, like FcγRIIA, FcγRIIC or on the adaptor protein Fcγ subunit, such as FcγRIIIA. Human FcγRIIIB is a glycosylphosphatidyl inositol (GPI)-anchored protein without ITAMs that activate intracellular signaling by lipid raft-mediated activation (Yang, et al., 2018). All the activating Fcγ receptors of mice, including FcγRI, FcγRIII and FcγRIV, have their ITAMs on the adaptor protein Fcγ subunit. Aside from the Fcγ receptor subtypes, the IgG isotypes also show difference between mice and human. For example, the human IgG1 with high effector function and affinity toward Fcγ receptors is more similar to the characteristics of mouse IgG2a, instead of the mouse IgG1 (Steward, et al., 2014).. 9.
(23) 1.4 Fcγ receptor IIB (FcγRIIB, a.k.a. CD32B) 1.4.1 General Properties. Both FcγRIIB (a.k.a. CD32B) of mice and humans are low-affinity Fcγ receptors and the only inhibitory Fcγ receptors, which contains an immunoreceptor tyrosine-based inhibitory motif (ITIM) (Moldta and Hessellb, 2014). Except for T cells and NK cells, immune cells express certain isoforms of FcγRIIB. FcγRIIB demonstrate highest binding affinity toward IgG1 or IgG3 in humans, whereas IgG2a and IgG2b in mice. FcγRIIB can be further classified into FcγRIIB1, FcγRIIB2 and FcγRIIB3 isoforms, based on alternative splicing of the mRNA. FcγRIIB1 is expressed in B cells and is usually prevented from endocytosis function. FcγRIIB2 is generated by alternative splicing of mRNA, giving rise to a 47-amino acid deletion at the upstream of the ITIM. FcγRIIB2 is mainly expressed on the surface of myeloid cells, including macrophages, monocytes and dendritic cells, and function as an endocytosis mediator. FcγRIIB3 is a soluble isoform without the transmembrane region and signal transduction domain, which can inhibit forming of ICs (Li and Kimberley, 2015). FcγRIIB deficiency is found be correlated with autoimmune diseases, such as 10.
(24) systemic lupus erythematosus, multiple sclerosis, etc. Clinically, FcγRIIB is expressed on malignant B cells enhance the internalization rate of monoclonal antibody and thus, contribute to the resistance of B-cell immunotherapy (Roghanian, et al., 2015). Mice deficient in FcγRIIB spontaneously develop splenomegaly due to uncontrolled expansion of B cells. These mice gradually develop a lupus-like disease resulting from massive IC deposition in the kidney after 6 months of age (Bolland, et al., 2002). Consistent with these results, the surface expression level of wild-type FcγRIIB in memory B cells and PCs is down-regulated, thereby reducing inhibitory activity in patients with SLE (Mackay, et al., 2006). Likewise, lupus-prone mice fail to up-regulate FcγRIIB expression on GC B cells, regardless of their genetic backgrounds, suggesting a role of FcγRIIB in the GC (Rahman, et al., 2005).. 1.4.2 Physiological function and signal transduction of FcγRIIB The main physiological function of FcγRIIB is to serve as a negative regulator by feedback inhibition of humoral response via cognate ICs that co-ligate BCR (via Ag) and FcγRIIB (via IgG Ab), resulting in inhibition on proliferation and differentiation of. 11.
(25) B cells. Coengagement of BCR and FcγRIIB by the IgG ICs triggers the phosphorylation of ITIM and recruitment of SH2-containing inositol 5′-Phosphatase (SHIP) to transduce inhibitory signals (Ambrosso, et al., 1996). On the other hand, non-cognate IgG ICs can mediate FcγRIIB aggregation when BCR is not recognized or has low affinity to the Ag and induce apoptosis of B cells through c-Abl tyrosine kinase in mice (Zheng, et al., 2005). We have recently shown that this pathway mediated by FcγRIIB alone is crucial for the deletion of low-affinity GC B cells in that a human polymorphic allele of FcγRIIB-I232T defective in inhibition leads to retention of low-affinity GC B cells, a major source of autoreactive B cells (Jhou, et al., 2018).. 1.4.3 FcγRIIB-I232T polymorphism Compared to wild-type mice, deficiency in FcγRIIB results in higher antibody titers after immunization (Ono, et al, 1996). This result suggests FcγRIIB as a potential target of immunomodulation for vaccine-elicited antibody response. On the other hand, FcγRIIB-deficient mice are prone to produce self-reactive B cells that secrete anti-nuclear antibodies and ultimately lead to a lupus-like disease (Li and Kimberley,. 12.
(26) 2014). Recently, we have shown that FcγRIIB-I232T variant mice, of which the 232nd amino acid of FcγRIIB protein was mutated from isoleucine to threonine, resulting in reduced inhibition as previously reported (Kono, et al., 2005). We demonstrated that these mutant mice display impairment in the elimination of low-affinity GC B that should have been deleted during affinity maturation. As a result, immunized FcγRIIB232T/T mice exhibit higher levels of both high- and low-affinity IgGs after immunization and also expansion of LZ B cells in the GC due to a failure in the elimination of low-affinity GC B cells (Jhou, et al., 2018). In addition, our preliminary data showed cross-reactivity of 4-hydroxy-3nitrophenylacetyl (NP) Ag-specific IgGs to 4-hydroxy-5-iodo-3-nitrophenylacetyl (NIP), a cognate Ag. This finding suggests the repertoire of low-affinity IgGs is diverse and potentially cross-reactive. Based on these findings, we reasoned that pharmacological down-regulation of FcγRIIB levels during the affinity maturation in the GC might be able to safely generate higher levels of both Ag-specific and cross-reactive Abs without long-term side effects for ongoing affinity maturation can clear low-affinity B cells after a period of time. Such modulation via FcγRIIB might be able to alter vaccine responses. 13.
(27) for better efficacy and cross-protection, e.g. influenza, HBV, etc.. 1.5 Peroxisome proliferator activated receptors (PPARs) PPARs are nuclear receptors that are activated with ligand binding and they function as transcription factors. Various PPARs share similar structures that contain the N-terminal domain, the DNA-binding domain, the hinge and the ligand-binding domain. The activation function-1 (AF-1) domain that can control the activation of PPARs independently of the ligand binding lies in the N-terminal region. The DNA-binding domains are highly conservative with zinc fingers among PPARs (Higashiyama, et al., 2007). During activation, the PPARs heterodimerize with the retinoid X receptor (RXR) and bind to the PPAR response element (PPRE) with DNA sequences of “AGGTCAnAGGTCA” (Zhang and Young, 2008). PPARs are classified into PPAR-α, PPAR-β/δ, PPAR-γ. PPAR-α regulates fatty acid oxygenation and ketogenesis, and is mainly expressed on cardiac muscle, hepatocytes and brown adipose tissue. PPAR-γ is mainly expressed on brown and white adipose tissues and controls adipogenesis and lipid storage (Poulsen, et al., 2012).. 14.
(28) PPARs affect cell metabolism and energy usage, making them potential targets for regulating immune response, cell differentiation and development.. 1.6 PPARδ PPAR-δ is localized in the nucleus in most tissues with exception of cytoplasmic localization in the islet of Langerhans, nervous system and macrophages. PPAR-δ regulates gene expression and mediates cell differentiation, metabolism, tumorigenesis and immune reaction. Thiazolidinediones (TZD) is a PPARγ, another subtype of PPAR receptor, agonist and it has therapeutic effects on type 2 diabetes mellitus (Quinn, et al, 2008). However, no drugs of PPAR-δ have been approved for clinical use so far. The natural ligands of PPAR-δ are intracellular fatty acids and metabolites, including 9-hydroxyoctadecadienoic. acid. (HODE),. 4-hydroxynonenal. (4-HNE),. and. 15-hydroxyeicosatetraenoic acid (15-HETE). Synthetic compounds of PPAR-δ have been developed, based on the ligand binding site structural design (Wu, 2017). With the co-repressor B-cell lymphoma 6 (Bcl-6) binding to the PPAR-δ and RXR, conformational change blocks access of the ligand binding sites for PPAR-δ and RXR. 15.
(29) and thereby inactivates gene transcription. Whereas through ligand and coactivator binding, activated PPAR-δ forms a stable complex with RXR and activates transcription (Cox, 2017). Whether PPAR-δ is involved in the regulation of humoral response by influence the gene expression of FcγRIIB has not been determined. Interestingly, PPAR-δ agonists have been shown to enhance macrophage survival under hypoxic stress and down-regulate gene transcription of FcγRIIB in macrophages (Adhikary, et al., 2015).. 1.7 GW501516 GW501516, also known as cardarine, is a synthetic and specific PPAR-δ agonist, which was designed from the structure of the ligand binding site, and therefore it is highly specific. GW501516 has been shown to confer striking effects on improving serum lipid profile in obese rhesus monkeys at the dose of 3 mg/kg/day (Luo, 2012). However, carcinogenesis was observed in mice by daily oral administration of greater than 10 mg/kg/day (Newsholme, et al, 2012). The tumor promoting effect is considered through PPAR-δ activation rather than genotoxicity. On the other hand, GW501516 is a. 16.
(30) selective androgen receptor modulator that enhances athletic performance (Pokrywka, 2014). The EC50 of GW501516 to PPAR-δ is less than a thousandth of the EC50 of PPAR-α and PPAR-γ, respectively. In vitro study of GW501516 further reveals peak activity at the concentration above 10 nM (Oliver, et al., 2001).. 1.8 Nilotinib Nilotinib is a Food and Drug Administration (FDA) approved medication for treating chronic myelogenous leukemia (CML). It is a small molecule that acts as a tyrosine kinase inhibitor (TKI) that blocks the activity of breakpoint cluster region-Abelson tyrosine kinase (BCR-Abl), discoidin domain receptors, platelet-derived growth factor receptors, etc. Nilotinib can bind to either wild-type or imatinib-resistant BCR-Abl due to its rational design from the protein binding pocket. Nilotinib is effective for most identified imatinib-resistant related mutations of BCR-Abl, with an IC50 below 2000 nM (Manley, et al., 2010). The elimination half-life of nilotinib is 17 hours. The recommended dose for Philadelphia chromosome positive (Ph+) CML pediatric patients is 230 mg/m2 twice daily orally based on two clinical trials:. 17.
(31) CAMN107A2120, CAMN107A2203. For newly diagnosed Ph+ adult CML patients, 300 mg orally twice daily is recommended (Prod Info Tasigna, 2007).. 18.
(32) 1.9 Motivation A recent study of human monocyte-macrophage has identified a paradoxical phenomenon that both anti-inflammatory and pro-inflammatory genes were activated by GW501516 (Adhikary, et al., 2015). The FCGR2B gene expression is suppressed during hypoxic stress based on fluorescence-assisted cell sorting (FACS) analysis. Interestingly, the LZ of the GCs is a hypoxic environment (Cho, 2016). Taken together, we hypothesized that by activation of PPAR-δ during the stage of affinity maturation in the LZ, the surface expression level of FcγRIIB on macrophages and/or B cells could be down-regulated. If our hypothesis is correct, the number of PCs will increase substantially and so do the titers of the low-affinity Abs owing to a reduced inhibition by FcγRIIB. In this study, we use the commonly used hapten 4-hydroxy-3-nitrophenyl acetic acid conjugated with chicken gamma globulin (NP-CGG) as the immunogen to investigate the Ab titers and distinguish their affinity to the NP Ag. We performed ELISA and ELISPOT assays for detecting serum Ag-specific Abs and splenic PC numbers as a functional readout of the stringency of selection of GC B cells in the LZ.. 19.
(33) Chapter 2 Materials and Methods. 20.
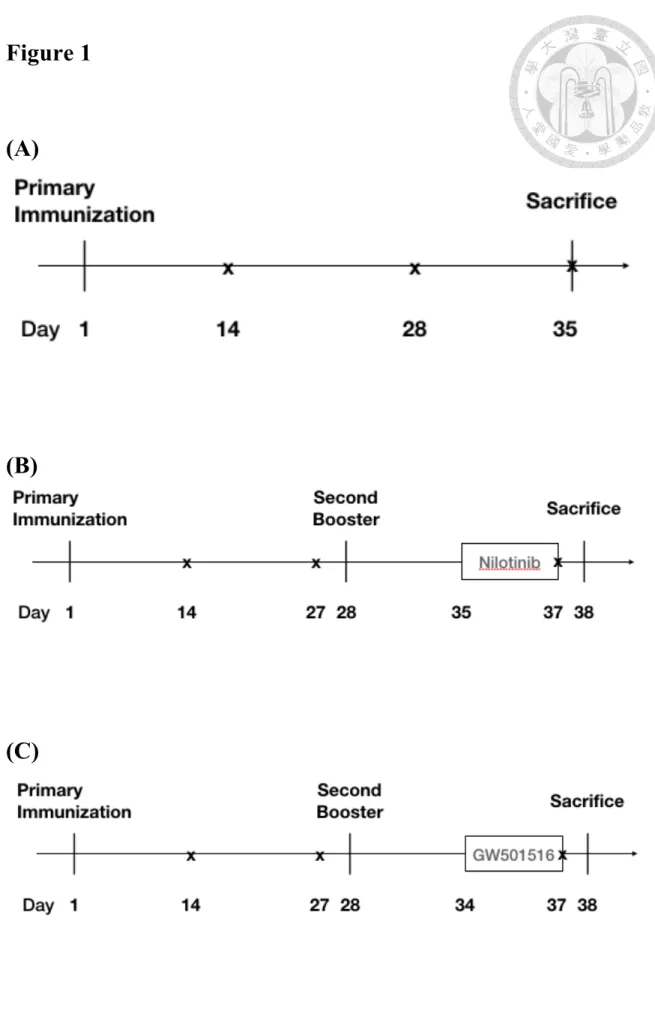
(34) Chapter 2 Materials and Methods 2.1 Mice. Female C57BL/6 mice were obtained from the Laboratory Animal Center of National Taiwan University College of Medicine. We immunized 7 to 10-week-old mice intraperitoneally with 50 µg nitrophenyl-chicken gamma globulin (NP-CGG, ratio 20, 1 mg/ml, Bioresearch Technology) admixed with equal volume alum precipitate (Alhydrogel® adjuvant 2%). The same antigen was immunized a second time 28 days later. GW501516 was administered intraperitoneally on the 6th day to 9th day after the second booster of immunization. Nilotinib was administered intraperitoneally on the 7th day to 9th day after the second booster of immunization. Mice were then sacrificed on the next day of the last injection. The animal protocol and schedule are illustrated in Figure 1.. 2.2 Enzyme Linked Immunosorbent spot (ELISPOT) assay PVDF-based ELISPOT plates were pretreated with 30% ethanol to increase hydrophobicity. After PBS wash twice, NP7-BSA and NP30-BSA were coated with 50 µl/well at the concentration of 10 µg/ml in PBS at 4°C overnight. Splenocytes were 21.
(35) harvested by crushing the spleen with two pieces of microslides and then filtered into a 15 ml centrifuge tube containing sterile PBS. To eliminate the red blood cells (RBCs), RBC lysis buffer was added and incubated for 5 min. Cells were centrifuged and resuspended in RPMI 1640 containing 10% fetal bovine serum. Splenocytes were seeded at about 3 x 105/well for detection of Ag-specific IgG and IgM secreting PCs and incubated at 37°C, 5% CO2 overnight. Horseradish peroxidase (HRP) conjugated rabbit anti-mouse IgG and goat anti-mouse IgM Abs were incubated at room temperature for detection of PCs for 2 hours. The 3-amino-9-ethylcarbazole (AEC substrate, BD Bioscience) of 50 µl was added as the substrate of HRP to develop spots resulting from PCs. To quench the reaction, plates were washed with ddH2O. The spots were counted after completely air dry by C.T.L. S6 universal analyzer (Cellular Technology Limited, Cleveland, OH, USA).. 2.3 Enzyme Linked Immunosorbent assay (ELISA). High binding 96-well microplates (Corning® 96 Well EIA/RIA Assay Microplate) were coated with anti-mouse IgG + IgM Abs, NP2-BSA, NP7-BSA, and NP30-BSA 22.
(36) with 10 µg/ml in 50 µl at 4°C overnight, respectively. Plates were blocked with BlockPROTM blocking buffer in room temperature for 2 hours. Serum samples collected from submandibular vein puncture of the mice were diluted in blocking buffer with 1: 40000 or 20000 dilutions for measuring IgG and IgM levels, respectively. Mouse reference serum (mouse reference serum, RS10-101, Bethyl Laboratories) in 2-fold serial dilution was used for generation of standard curves for quantification. Samples and diluted reference serum were incubated at room temperature for 2 hours. HRP conjugated rabbit anti-mouse IgG and goat anti-mouse IgM were incubated at room temperature for detection for 2 hours. After washing with PBS-T (phosphate buffered saline with 0.05% Tween 20), tetramethylbenzidine substrate was added for development. The reaction was terminated with 2N H2SO4.The absorbance OD450 nm and OD570 nm was read by an ELISA plate reader (BioTek, Winooski, VT, USA).. 2.4 Confocal microscopy. Mouse splenic B cells were isolated from wild-type and FcγRIIB232T/T mice of 8 weeks of age using the mouse B cell isolation kit (BD Pharmingen) to more than 95% 23.
(37) purity. Cells were treated with 10 µg of rabbit anti-mouse IgM (Jackson Immunoresearch) for 15 min on ice. After a brief wash, either FITC-labeled cholera toxin B (Sigma) or Cy3-labeled anti-rabbit IgGs (Jackson Immunoresearch) were added for 15 min on ice, followed by additional incubation for 5-60 min at room temperature. Cells were washed by PBS, fixed by 4% paraformaldehyde and then mounted. Metamorph software was applied to quantify colocalization of FcγRIIB with lipid raft.. 24.
(38) Chapter 3 Results. 25.
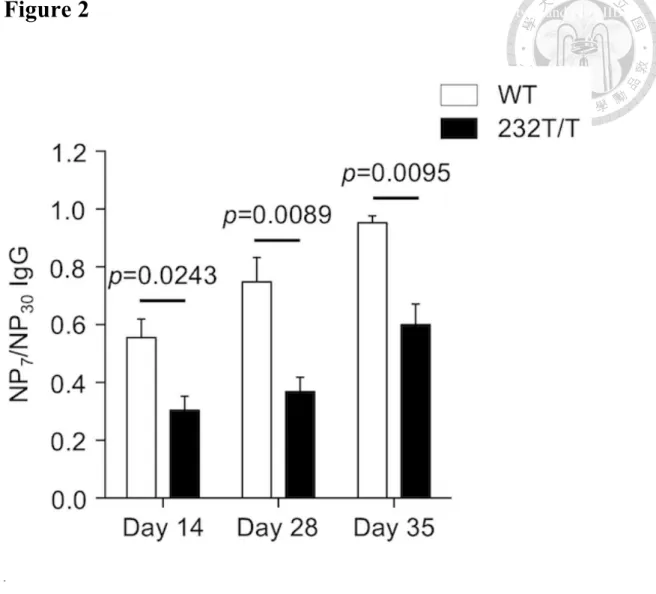
(39) Chapter 3 Results 3.1 FcγRIIB232T/T mice show persistent impaired affinity maturation over time To investigate whether reduced inhibitory function of FcγRIIB could affect the selection of GC B cells, we have generated FcγRIIB232T/T mice to test our concept. We have recently shown that the FcγRIIB-I232T polymorphism, of which the 232th amino acid was substituted from isoleucine to threonine, in mice results in impairment in the negative selection of GC B cells during GC reaction (Jhou, et al., 2018). To evaluate the long-term effect of FcγRIIB232T/T on affinity maturation, we immunized female wild-type and FcγRIIB232T/T mice with 50 µg of NP-CGG admixed with an equal volume of alum adjuvant. Serum samples were collected on days 14, 28 and 35. Serum levels of high-affinity NP-specific IgG and total NP-specific IgG were measured with ELISA plates coated with NP7-BSA and NP30-BSA proteins, respectively. The affinity maturation status was measured by dividing high-affinity NP-specific IgG by total NP-specific IgG from ELISA readings. We found that on day 14, the serum levels of NP-specific IgG of FcγRIIB232T/T mice showed significantly lower affinity maturation. 26.
(40) (P < 0.05, Figure 2) than those of wild-type mice. By day 35, the affinity maturation status of serum NP-specific IgGs of wild-type mice was close to 1, indicative of nearly total elimination of the low-affinity NP-specific GC B cells to produce mostly high-affinity NP-specific IgGs. In contrast, FcγRIIB232T/T mice revealed continued retention of low-affinity NP-specific IgGs and a slower affinity maturation kinetic toward maturation to high-affinity NP-specific IgGs over time. These data from FcγRIIB232T/T mice suggest that pharmacological down-regulation of FcγRIIB expression might induce a similar Ab phenotype resulting from reduced inhibitory activity of FcγRIIB.. 3.2 Association of FcγRIIB and BCR with the lipid raft after coligation was impaired in FcγRIIB 232T/T mutant mice The I232T polymorphism is a single nucleotide polymorphism (SNP) in the transmembrane domain of FcγRIIB proteins. Coligation of FcγRIIB with immune complex results in apoptosis of the centrocytes. We hypothesized that the mouse FcγRIIB-I232T would alter its stability in the association with lipid raft as observed in. 27.
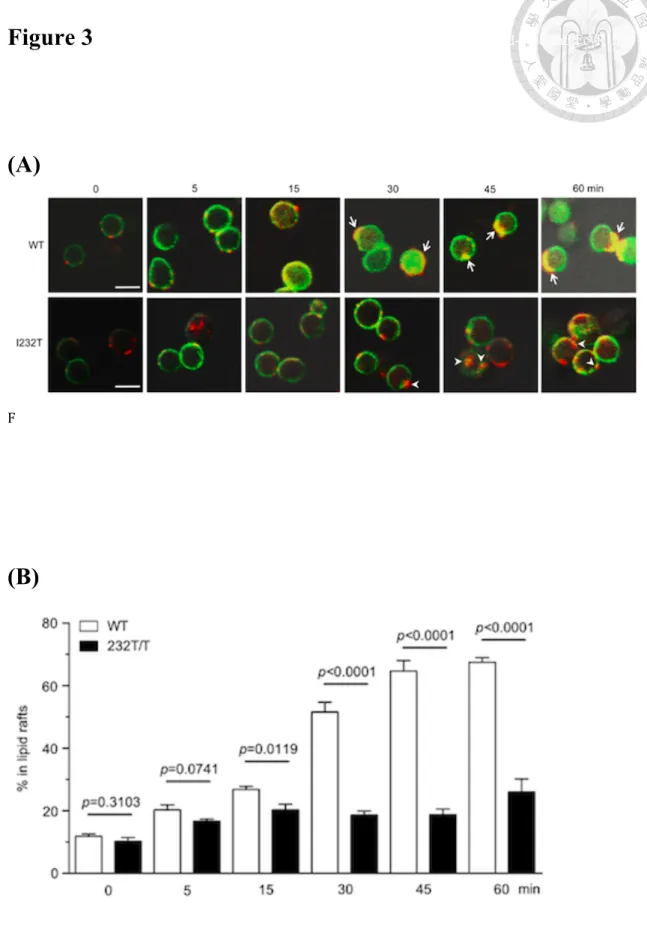
(41) human counterpart. To investigate this, confocal microscopy was applied to visualize the effect of FcγRIIB-I232T on BCR in the lipid microdomain at the cell surface. The ganglioside GM1, a lipid raft marker that binds to the subunit B of cholera toxin, and FcγRIIB were respectively labeled as green with FITC and red with Cy3-conjugated Abs. As shown in Figure 3A, the wild-type B lymphocytes showed remarkable yellow cap structure at 30 to 60 min, indicating that the FcγRIIB were stably coligated within the lipid raft with BCR in response to whole anti-Ig crosslinking. In contrast, B lymphocytes from FcγRIIB232T/T mice showed discrete FcγRIIB and lipid raft co-localization. Metamorph analysis tool was applied to quantify the percentage of FcγRIIB localized in the lipid raft. From 15 to 60 min, we found that a significantly lower co-localization in isolated FcγRIIB-232T B cells in comparison with the wild-type B cells in a time-dependent fashion (Figure 3B). Our findings FcγRIIB-232T B cells are consistent with those observed in human peripheral mononuclear cells isolated from FcγRIIB-232T carriers (Kono, et al., 2005).. 28.
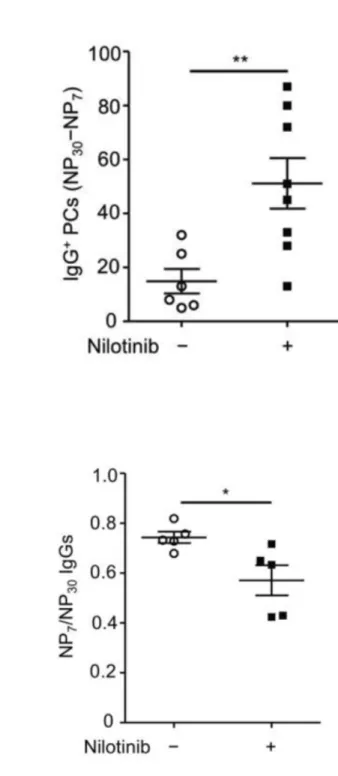
(42) 3.3 Nilotinib administration during GC reaction has a negative impact on affinity maturation To assess whether it is applicable to manipulate affinity maturation with FcγRIIB targeting agent, we examine the effects of nilotinib, a specific c-Abl inhibitor. As reviewed in the introduction, c-Abl is the key downstream signal protein involved in the apoptosis pathway induced by aggregation of FcγRIIB via non-cognate ICs. Mice were immunized twice with NP-CGG 50 ug admix with equal volume of alum on day 1 and day 28 (Figure 1). After the second booster of NP-CGG, we administered the wild-type female mice with nilotinib with the dose of 2 mg/kg/day daily from the sixth day (day 35) to the ninth day (day 37), which was the most active time of GC reaction, clonal selection and apoptosis of low-affinity GC B cells. Immunized mice were sacrificed on day 38, the following day after last treatment of nilotinib. Sera and splenocytes were collected. NP-specific IgG secreting PCs and IgG levels were detected and quantified with ELISPOT and ELISA assays, respectively. Low-affinity NP-specific IgG secreting PCs were measured with subtracting the NP7-specific IgG-secreting PC count from. 29.
(43) NP30-specific IgG-secreting PC count. In nilotinib-treated group, the low-affinity NP-specific IgG secreting cell count was significantly higher (P < 0.05, Figure 4A). Correspondingly, the affinity maturation status of the circulating NP-specific IgGs was significantly reduced (P < 0.05, Figure 4B).. 3.4 GW501516 increases respective total and low-affinity Ag-specific IgG PCs at the dose of 3 and 6 mg/kg/day during GC reaction To observe the effects of GW501516 on the selection process of GC reaction, we gave each wild-type female mouse with either 3 or 6 mg/kg/day on the sixth to the ninth day after the secondary immunization when IgG ICs were abundantly present. The choice of doses was based on a study of obese rhesus monkey, of which lipid-lowering effect by GW501516 was dramatically observed at 3 mg/kg/day given for 1 month. More importantly, the dosage of 3 mg/kg/day was safe and not reported to induce carcinogenesis in mice in previous studies. Due to the short-term treatment of GW501516 for 4 days, a higher dose of 6 mg/kg/day of GW501516 was also included in our study to observe a potential dose-dependent effect. The control group was given. 30.
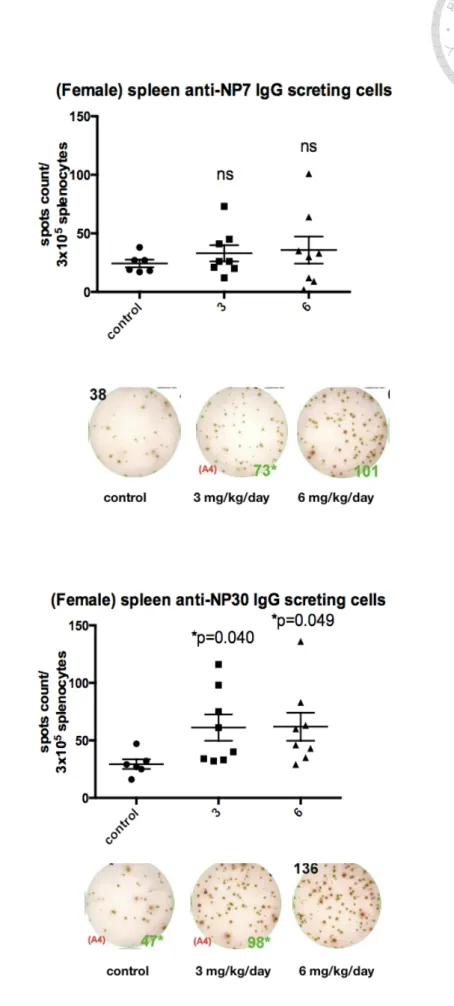
(44) an equal volume of diluted dimethyl sulfoxide (DMSO) in PBS, which is the vehicle of GW501516. We performed ELISPOT assay to assess Ag-specific IgG secreting PCs in spleen. We found that mice respectively given 3 and 6 mg/kg/day of GW501516 could generate more NP30-specific IgG secreting PCs than those of control mice. Moreover, the increase in the both groups of 3 and 6 mg/kg/day was statistically significant (P < 0.05, Figure 5B). The total Ag-specific IgG secreting cells show increment in each dose but no dose dependent effect was observed. The number of NP7-specific IgG secreting cells, considered as high affinity NP-specific IgG secreting cells, did not show a significant change (Figure 5A). The low-affinity Ag-specific IgG secreting PC count, defined as the numbers of NP30-specific IgG secreting PC numbers minus NP7-specific IgG secreting PC numbers, increased at the dose of 3 and 6 mg/kg/day in a dose-dependent fashion (P < 0.05, Figure 5C). The effect was apparent on 6 mg/kg/day probably because of no significant increase and also mild decrease in some of the mice in the numbers of high-affinity NP+ PCs.. 31.
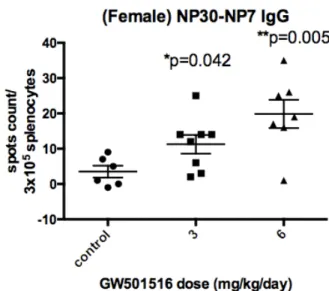
(45) 3.5 GW501516 increases the serum levels of total and low-affinity Ag-specific IgG at the dose of 3 and 6 mg/kg/day in wild-type mice In order to assess whether the serum IgG level was associated with the numbers of IgG secreting PCs, we performed indirect ELISA with either NP2-BSA (1 BSA conjugated with 2 NPs) or NP30-BSA as the Ag coated onto 96-well plates. Serum samples were diluted with PBS in a factor of 4 x 104. The relative NP Ag-specific IgG concentrations expressed in arbitrary units per ml (AU/ml) were calculated from semilog regression of the OD450 levels of standard serum. The average of total antigen-specific (NP30-specific) IgGs increased approximately 2 folds in both 3 and 6 mg/kg/day groups but the results did not show a dose-dependent pattern (P < 0.05, Figure 6B). The level of high-affinity IgGs (NP2-specific) did not show significant differences (Figure 6A). We divided the high-affinity Ag-specific IgG level over total Ag-specific IgGs as the antibody avidity index as a functional readout of affinity maturation. The higher the ratio, the better the affinity maturation in terms of high-affinity IgG production. The results showed a significant decrease in the affinity maturation in 3 mg/kg/day but not in 6 mg/kg/day. As a result, the circulating IgG. 32.
(46) levels were highly relevant to the IgG secreting cell count in the spleen. Notably, the interpretation of IgG levels is slightly distinct from antigen secreting cell count since the affinity maturation terminates and the germinal centers diminished in the spleen after the primary immunization. But the circulating IgGs continue to increase affinity till day 35 as shown in Figure 2. The average ratio was about 0.8 in 3 mg/kg/day (Figure 6C) compared with 1 in control group, which indicating complete affinity maturation on day10 after the second booster. The results were consistent with the prediction that GW501516 can increase low-affinity antibody production and impair negative selection during the GC reaction. On average, high-affinity antibody levels were not affected, but the GW501516-treated mice presented a greater variation than the control group. Namely, the GW501516 treatment has an ambiguous effect on high-affinity Abs.. 3.6 GW501516 increases total Ag-specific IgM secreting PCs at the dose of 6 mg/kg/day during GC reaction Our results had shown effects of GW501516 on the GC reaction and humoral response of repeated immunization. Traditionally, IgM is not considered as the main. 33.
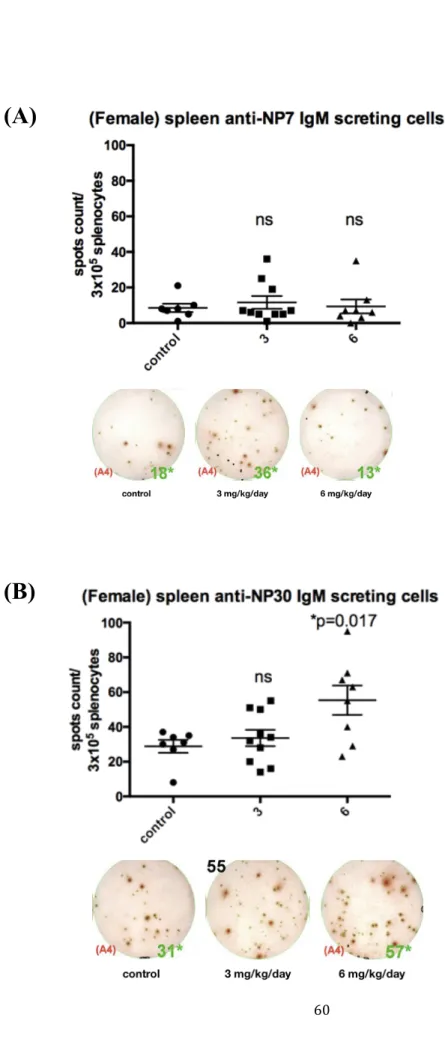
(47) component of affinity maturation since the centrocytes undergo class switch to IgG secreting PCs. Especially after the secondary immunization, IgM has little role in the humoral immunity. Circulating IgMs are produced from extrafollicular PCs that do not participated in the GC reaction. To evaluate whether GW501516 had effects on extrafollicular IgM secreting PCs, we performed ELISPOT assay to determine the numbers of IgM secreting PCs in the spleen. GW501516 administration increased the levels of NP30-specific (total NP-specific) IgM at the dose of 6 mg/kg/day (P < 0.05, Figure 7B) but not 3 mg/kg/day. In the group of mice treated with 3 mg/kg/day, the effect of GW501516 varies that some of the mice decreased but some increased in total numbers of NP-specific PCs (Figure 7A). When the numbers of high-affinity IgM PCs were analyzed, no significant difference was observed in each dose. However, the numbers of low-affinity IgM NP+ PCs (total IgM PCs minus high-affinity IgM PCs) increased significantly in mice treated with 6 mg/kg/day for 4 days (P < 0.05, Figure 7C). The avidity index was 0.5 in control group but the GW501516-treated group display an average 0.6 and up to 0.8 in some mice.. 34.
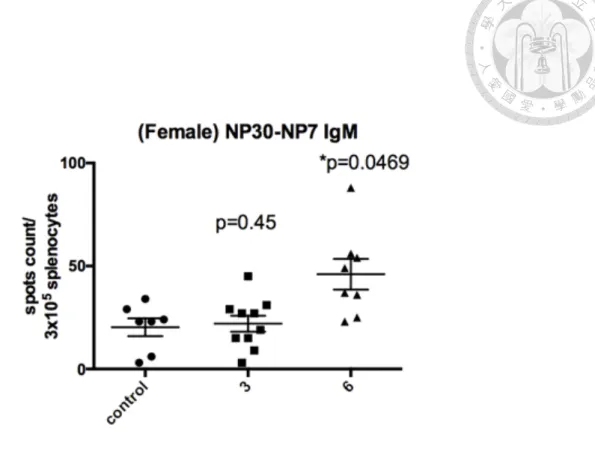
(48) 3.7 GW501516 increases the serum levels of high-affinity Ag-specific IgM at the dose of 3 and 6 mg/kg/day and total Ag-specific IgM at 6 mg/kg/day To assess whether the result of Ag-specific IgM PCs is associated with serum levels of Ag-specific IgMs, we performed ELISAs. The serum samples were diluted in a factor of 2 x 104. The serum Ab levels were displayed in arbitrary units per ml. Consistent with the findings in NP30-specific (total NP-specific) IgM secreting PCs, the serum total Ag-specific IgM levels increased in mice treated with 6 mg/kg/day (P <0.05, Figure 8B) but not in 3 mg/kg/day. On the other hand, the NP2-specific (high affinity NP-specific) IgMs increased in both 3 and 6 mg/kg/day in a dose-dependent pattern, which was not concordant with the results in IgM-secreting PCs (Figure 8B). The antibody avidity index of the high-affinity Ag-specific IgMs over the total NP-specific IgMs showed increased ratios in both groups of 3 and 6 mg/kg/day treatments owing to the higher levels of high-affinity Ag-specific IgMs (Figure 8C). The result indicates that the effect of GW501516 on FcγRIIB might not be the only factor to influence the humoral immunity and affinity maturation. GW501516 might also affect the memory B cell response, Ab secretion, and class switch of centrocytes and Tfh activity.. 35.
(49) . Chapter 4 Discussion. 36.
(50) Chapter 4 Discussion 4.1 FcγRIIB232T/T mutant mice as a model for investigating the functional role of FcγRIIB in GC reaction FcγRIIB act as the only inhibitory Fcγ receptor of immune response. Impairment of FcγRIIB leads to development of persistent GC reaction and autoimmune diseases. I232T is a single nucleotide polymorphism of Fcgr2B in human that is highly associated with SLE. As previously mentioned, the I232T SNP locates in the transmembrane domain, impairing signal transduction and stability during coengagement with BCR in the lipid raft. Despite that FcγRIIB232T/T mutant mice are functionally impaired of the receptor on all the immune cells, FcγRIIB232T/T mice did not present SLE symptoms even after immunization. Because FcγRIIB-deficient mice spontaneously develop lupus symptoms after 6 months of age, advanced age of FcγRIIB232T/T is likely required to trigger lupus pathology. However, we can still observe impaired negative selection, larger GCs and more PCs in FcγRIIB232T/T mice. On the other hand, FcγRIIB-deficient mice present with hyper-immunity and are used as a model for SLE. The properties of FcγRIIB232T/T mice make them an ideal human disease-relevant model for research on. 37.
(51) the role of Fcγ receptors in the GC reaction and also pharmacological manipulation of Fcγ receptors for vaccination, e.g. phenotype rescue, or the treatment of autoimmune disorders, as opposed to FcγRIIB-deficient mice.. 4.2 Difference between FcγRIIB. 232T/T. mutant mice and wild type mice. treated with nilotinib or GW501516 We tested nilotinib and GW501516 for the potential of manipulating the selection outcome of GC reaction. We hypothesized that nilotinib inhibits the downstream c-Abl signal pathway triggered by coengagement of FcγRIIB, negative selection and apoptosis of low-affinity centrocytes in the LZ were impaired, leading to accumulation of low-affinity GC B cells. For GW501516, it can reduce FcγRIIB expression levels, mimicking the condition of reduced FcγRIIB inhibitory activity. The timing of administering nilotinib and GW501516 was designed to meet the maximum negative selection and apoptosis of GC B cells, but to avoid affecting early phase GC reaction. As shown in Figure 2, FcγRIIB232T/T mice remained impaired on affinity maturation till 35 days after primary immunization. For wild-type mice on day 28 after primary. 38.
(52) immunization, the day for second booster, the circulating IgG affinity maturation was 0.8. Therefore, the baseline conditions for FcγRIIB232T/T mice and wild type mice treated with nilotinib or GW501516 are different. We could reason that memory cells generated from the primary immunization are not exposed to nilotinib or GW501516, which might affect the secondary immune response. It is reasonable that by administering nilotinib or GW501516 cannot fully replicate the functional impairment in FcγRIIB-I232T polymorphism. After drug elimination, we expected that the affinity maturation would go on and FcγRIIB activity or expression would return to normal conditions. Thus, the risk for inducing autoimmune disease is expected to be low. The long-term effect requires further investigation.. 4.3 Summary of the overall effect of GW501516 on GC reaction In this study, we showed that GW501516 at the dose of 3 and 6 mg/kg/day can influence the GC reaction, resulting in an increase of the numbers of total Ag-specific IgG PCs and serum levels of Ag-specific IgGs. The high-affinity Ag-specific PCs and Ag-specific IgG levels were not significantly different when compared with the control. 39.
(53) group (Figures 2 and 3). However, GW501516 administration at 6 mg/kg/day showed paradoxical results in that some mice displayed lower levels of total Ag-specific IgG PCs, whereas some mice showed the opposite findings (Figures 6 and7). Our results showed a concomitant increase of the low-affinity PCs and serum IgGs (Figures 6 and 7) in treated groups. These results support the involvement of GW501516 in GCs with negative influence on the affinity maturation reminiscent of the findings observed in FcγRIIB-232T mice. Interestingly, GW501516 appears to increase the numbers of high-affinity Ag-specific IgG ASCs and serum IgG. On the other hand, the levels of high-affinity Ag-specific IgM increased more markedly than the increased number of ASCs. As the extrafollicular ASCs mostly carry IgM on their surface and they do not undergo affinity maturation in the germinal center, we reasoned that GW501516 might have effects on the secretion of IgM but this needs to be further investigated.. 4.4 Could GW501516 affect GC reaction other than FcγRIIB down-regulation induced by PPAR-δ activation? A previous study demonstrates that PPAR-δ activation down-regulates the. 40.
(54) expression of FcγRIIB in a macrophage cell line (Adhikary, et al., 2015). In addition, our previous study on FcγRIIB-I232T mice revealed enhanced antibody production and reduced affinity maturation (Jhou, et al., 2018). The down-regulation of FcγRIIB in macrophages could be a result of direct regulation by PPAR-δ but whether the effect of PPAR-δ activation is the same in B cells remained unknown. On the other hand, PPAR-δ is a transcription factor with different functions on T cells, B cells and macrophages.iii How activation of PPAR-δ might lead to alternation of the interaction between B cells and Tfh or between Tfh and B cells remains unclear. However, our preliminary data from the analysis of flow cytometric results did not show a significant change in surface FcγRIIB levels in CD11b+ myeloid cells in the spleen. A more detailed analysis of the phenotypes of GC myeloid cells, especially at the marginal zone of the GC is warranted. Unlike splenic macrophages, splenic B cells showed a trend of decreased expression of FcγRIIB at the dose of 6 mg/kg/day, suggesting greater drug sensitivity. However, the relative numbers of DZ and LZ GC B cells did not show significant differences in vehicle- and GW501516-treated mice, suggesting that GW501516 did not affect the differentiation of DZ GC B cells (centroblasts) into LZ. 41.
(55) GC B cells (centrocytes) and not significantly influenced by FcγRIIB. A definitive answer can be obtained from inducible deletion of FcγRIIB gene at a defined time during GC reaction. FcγRIIB gene deleted at the same period of GW501516 administration after secondary immunization is expected to show more profound effects in the change of IgG PCs and their Abs secreted. PPAR-δ activates signaling pathways that affect immune responses and inflammation. The phenomenon in humoral response observed in our study might be an integrated result from multiple factors. For instance, Bcl-6, an important transcription factor in selection of B cells entering the GC, is also a transcriptional repressor of PPAR-δ. GW501516 induces activation of PPAR-δ and leads to unbinding and activation of Bcl-6 (Coleman, et al., 2013). In contrast, NF-κB is a transcription factor involved in pro-inflammatory effects and it is suppressed during PPAR-δ activation by AMP kinase (AMPK) and SIRT1 (Barroso, et al., 2011). PPAR-δ also modulates inflammatory responses that involve JAK-STAT pathway. On the other hand, GW501516 had been reported to increase fatty acid oxidation and glucose uptake in skeletal muscles via activation of AMPK, a kinase activated in hypoxic environment,. 42.
(56) and PPAR-δ (Kramer, et al., 2007). Since the GC is a hypoxic environment, it then suggests that energy metabolism might modulate antibody production (Cho, 2016). It is possible that PPAR-δ activation alters the metabolism in the GCs and then affects humoral immunity. It should be noted that our study focused on the response after a second booster of immunogen. The effect on memory B cells should also be considered for investigation. In addition, it is likely that the effect on the primary response will be different because of different natures in the production of IgM and IgG.. 4.5 Pharmacological modulation of FcγRIIB activity is an applicable method for novel manipulation of immune response Although the GW501516-mediated mechanisms of our findings are not clear at present, we showed immunomodulatory effects of GW501516, a PPAR-δ agonist, in vivo. The effects of widely used anti-diabetic drugs, such as metformin, TZDs, have been reported to be associated with PPARs activation. It is an interesting question to answer that if these anti-diabetic medications affect our immune response to pathogens and to vaccination. Importantly, our results reveal that PPAR-δ could be a potential. 43.
(57) target for innovative adjuvants in vaccine development and the development of cross-protective effects of vaccines. On the flip side, PPAR-δ inactivation might be a prospective therapy toward autoimmune diseases. Our ongoing research aims to unravel the molecular mechanism of immunomodulatory effects of GW501516 and PPAR-δ activation both in vitro and in vivo by using conditional and inducible knockout mice of FcγRIIB gene and to modulate GC response using various candidate compounds.. 4.6 Potential role of macrophages in the GC reaction to influence the generation of Ag-specific Abs It has been reported that defective phagocytosis of macrophages can lead to autoimmunity (Fond and Ravichandran, 2016). FcγRIIB can block IC-mediated activation of FcγR and other activating receptors in macrophages. Moreover, overexpression of FcγRIIB in myeloid cells suppresses host immunity against bacterial infection. Conversely, FcγRIIB-deficient macrophages increase their phagocytic property (Brownlie, et al., 2008). The gene transcription of FcγRIIB has been shown to be down-regulated in a macrophage cell line by PPAR-δ agonist (Adhikary et al., 2015).. 44.
(58) It is then crucial to determine whether the absence of FcγRIIB gene can promote macrophage function in vivo. Splenic macrophage subpopulations are diverse and their functions remain elusive. The unique tingible body macrophages (TBMs; Mer+DNaseI+) are predominantly scattered in the GC. TBMs express Mer receptor tyrosine kinase, which mediates phagocytic activity and regulates cytokine production (Rahman, 2011). They phagocytose apoptotic cells and thereby contain condensed chromatin fragments. TBMs are thought to play a role in down-regulation of GC reaction (Rahman, 2011). The marginal zone (MZ) surrounds lymphoid follicles, where MZ macrophages (MZMs; MARCO+CD169+) and metallophilic macrophages (MMMs; MOMA+) are abundantly present (McGaha and Karlsson, 2016). MZMs are specialized macrophages that phagocytosed apoptotic materials entering the spleen from circulation to minimize the immunogenicity of autoantigens (McGaha et al., 2011). MMMs distribute adjacent to the T- and B-cell-rich zones. Interestingly, MZMs and MMMs disappear when B cells were absent before birth or gradually depleted after birth, suggesting a critical role of B cells in the maintenance of splenic MZ structure (Nolte, et al., 2004). Interaction with. 45.
(59) MZ B cells is also crucial for efficient homing of MZMs and MMMs and for efficient removal of blood-borne pathogens coming into the MZ. To the best of our knowledge, whether the fate of GC B cells during affinity maturation is influenced by macrophages, whether FcγRIIB regulates phagocytic activity of GC and MZ macrophages, and how macrophages might interact with B cells in the GC are all incompletely understood. We have recently generated conditional knockout mice of FcγRIIB gene in myeloid cells to investigate the potentially important role of macrophages in the selection of GC B cells. Moreover, this mouse strain will help explain and conclude our findings on the effects of GW501516 of macrophages during affinity maturation of GC reaction in secondary immunization.. 46.
(60) Figures. 47.
(61) Figure 1 Mouse immunization schedules.. (A). (B). (C). 48.
(62) Figure 1. Mouse immunization schedules. (A~C) 7-to-10-week-old female mice were primarily immunized with 50 µg of NP20-CGG admixed with 50µl of alum intraperitoneally on day 1. The second booster was given on day 28 with the same antigen and route. The “x” denotes the day of collecting serum samples. Before the second booster, serum samples were collected on days 14 and 27. (A) Comparison of wild-type and FcγRIIB232T/T mice. Mice were sacrificed on day 35 after primary immunization. Sera were collected on day 14, day 28 and day 35. No second booster was given. (B) C57BL/6 mice were daily given nilotinib at 2 mg/kg/day or vehicle intraperitoneally from the sixth day to the ninth day after the second booster. Mice were sacrificed on the tenth day after the second booster (day 38). Serum was collected on day 37. (C) C57BL/6 mice were daily given vehicle, GW501516 3 or 6 mg/kg/day intraperitoneally from the sixth day to the ninth day after the second booster. Mice were sacrificed on the tenth day after the second booster (day 38). Serum samples were collected on day 37.. 49.
(63) Figure 2 Analysis of the differences of affinity maturation status between wild-type and FcγRIIB 232T/T. mice over time.. .. Figure 2. Analysis of the differences of affinity maturation status between wild-type and FcγRIIB 232T/T mice over time. On days 14, 28 and 35 after the primary immunization with NP-CGG, sera were collected. The serum NP-specific IgG levels were analyzed by ELISA assay. The status of affinity maturation was measured by the OD450 ratio of serum NP7-specific IgG/NP30-specific IgGs between wild-type (n=3) and FcγRIIB232T/T (n=4) mice on day 14 (P=0.0243), day 28 (P=0.0089) and day 35 (P=0.0095). 50.
(64) Figure 3 Analysis of the differences of affinity maturation status between wild-type and FcγRIIB 232T/T. mice over time.. (A). F. (B). 51.
(65) Figure 3. Compare FcγRIIB232T/T mutant and wild type mice of FcγRIIB cluster in the lipid raft with confocal microscopy. (A) Confocal image (400x magnification) of B-lymphocyte retrieved from the spleens 8-week-old FcγRIIB232T/T mutant and wild type mice. FITC-labeled cholera toxin B binds to the GM1 ganglioside of the lipid raft and display green fluorescence. Cy3-labeled anti-rabbit IgG binds to the anti-mouse IgM and display red fluorescence. FcγRIIB stabilized in the lipid raft would display yellow fluorescence. In wild type mice, cap structures were formed (arrows) at 30 to 60 min but FcγRIIB232T/T mutant mice did not show much merged yellow fluorescence (arrow head). (B) Quantitative representation of FcγRIIB localized in the lipid raft with Metamorph analysis tool. FcγRIIB232T/T mutant mice showed significant impaired lipid raft colocalization over 15 min (P < 0.05).. 52.
(66) Figure 4 Analysis of the effect of nilotinib on the affinity maturation of female wild-type mice.. (A). (B). 53.
(67) Figure 4. Analysis of the effect of nilotinib on the affinity maturation of female wild-type mice. (A, B) Wild type female mice were given nilotinib 2 mg/kg/day or vehicle given from the seventh day to the ninth day after the second booster. (A) Analysis of IgG secreting PC numbers with ELISPOT assay. Number of low affinity NP-specific IgG secreting PCs per 2.4 x 104 splenocytes by subtracting the NP7 specific IgG-secreting PCs from NP30 specific IgG-secreting PCs. Comparisons between vehicle (n=6) and nilotinib-treated group (n=8) (P=0.0087). (B) Analysis of serum NP-specific IgG concentration by ELISA assay. The affinity maturation was measured by the OD450 ratio of serum NP7-specific IgG/NP30-specific IgG between vehicle (n=5) and nilotinib-treated group (n=5) (P=0.0283).. 54.
(68) Figure 5 Analyzing IgG-secreting PCs in the spleen of female C57BL/6 mice immunized with two different GW501516 doses by ELISPOT assay.. (A). . (B). . 55.
(69) . (C). Figure 5. Analyzing IgG-secreting PCs in the spleen of female C57BL/6 mice immunized with two different GW501516 doses by ELISPOT assay. (A, B) ELISPOT quantification (upper panel) and demonstrative graph (lower panel) of the high-affinity and total NP-specific IgG PCs per 3 x 105 splenocytes. Female C57BL/6 mice, 10 days after the second booster (day 39). Vehicle (n=6, as control group), GW501516 3 mg/kg/day (n=9), GW501516 6 mg/kg/day (n=7) were administered, respectively, during the sixth to the ninth day after the second booster. (A) High-affinity NP7-specific IgG-secreting PC count: GW501516 3 mg/kg/day (ns), 6. 56.
(70) mg/kg/day (ns). (B) Total NP30-specific IgG-secreting PC count: GW501516 3 mg/kg/day (P=0.040), 6 mg/kg/day (P=0.049), compared with the control group. (C) Low-affinity NP-specific IgG secreting PCs measure by subtracting the NP7 specific IgG-secreting PCs from NP30 specific IgG-secreting cells. Low-affinity NP-specific IgG secreting PC count: GW501516 3 mg/kg/day (P=0.042), 6 mg/kg/day (P=0.005). Results were analyzed with unpaired, two-tailed student t test and shown as mean. SEM. *P < 0.05, **P < 0.01, ***P < 0.001 and ns, no significance. 57.
(71) Figure 6 Measurement of serum IgG levels of female C57BL/6 mice immunized with two different GW501516 doses by ELISA assay.. (A). (B). (C). . 58.
(72) F. Figure 6. Measurement of serum IgG levels of female C57BL/6 mice immunized with two different GW501516 doses by ELISA assay. (A, B) Serum high-affinity and total NP-specific IgGs in the serum of female C57BL/6 mice, 10 days after the second booster (day 39). Vehicle (n=5, as control group), GW501516 3 mg/kg/day (n=6), GW501516 6 mg/kg/day (n=6) were administered, respectively, during the sixth to the ninth day after the second booster. Serum was diluted in 4 x104 folds. (A) Serum high-affinity NP2-specific IgG: GW501516 3mg/kg/day (ns), 6mg/kg/day (ns). (B) Serum total NP30-specific IgG of treatment with GW501516 3 mg/kg/day (P=0.037) or 6 mg/kg/day (P=0.025), compared with the control group. (C) The affinity maturation measurement by dividing the NP2 specific IgG serum level by the NP30 specific IgG serum levels. NP2/NP30: GW501516 3 mg/kg/day (P=0.035), 6 mg/kg/day (ns). Results were analyzed with unpaired, two-tailed student t test and shown as mean. SEM. *P < 0.05, **P < 0.01, ***P <. 0.001 and ns, no significance. AU denotes arbitrary units. 59.
(73) Figure 7. Analyzing IgM-secreting PCs in the spleen of female C57BL/6 mice. immunized with two different GW501516 doses by ELISPOT assay. (A). (B). 60.
(74) (C). Figure 7. Analyzing IgM-secreting PCs in the spleen of female C57BL/6 mice immunized with two different GW501516 doses by ELISPOT assay (A, B) ELISPOT quantification (upper panel) and demonstrative graph (lower panel) of high-affinity and total NP-specific IgM-secreting cells per 3x105 splenocytes of female C57BL/6 mice, 10 days after the second booster (day 39). Vehicle (n=7, as control group), GW501516 3 mg/kg/day (n=10), GW501516 6 mg/kg/day (n=8) were administered, respectively, during the sixth to the ninth day after the second booster. (A) High-affinity NP7-specific IgM-secreting PC count: GW501516 3 mg/kg/day (ns),. 61.
(75) 6mg/kg/day (ns). (B) Total NP30-specific IgM-secreting PC count: GW501516 3 mg/kg/day (ns), 6 mg/kg/day (P=0.017), compared with the control group. (C) Low-affinity NP-specific IgM secreting PCs measure by subtracting the NP7 specific IgM-secreting PCs from NP30 specific IgM-secreting PCs. Low-affinity NP-specific IgM secreting PC count: GW501516 3 mg/kg/day (P=0.45), 6 mg/kg/day (P=0.0469) Results were analyzed with unpaired, two-tailed student t test and shown as mean. SEM,*P < 0.05, **P < 0.01, ***P < 0.001 and ns, no significance.. 62.
(76) Figure 8 Measurement of the serum IgM levels of female C57BL/6 mice immunized with two different GW501516 doses by ELISA assay. (A). (B). (C). . y. 63.
(77) Figure 8. Measurement of the serum IgM levels of female C57BL/6 mice immunized with two different GW501516 doses by ELISA assay. (A, B) Serum high-affinity and total NP-specific IgMs in the serum of female C57BL/6 mice, 10 days after the second booster (day 39). Vehicle (n=5, as control group), GW501516 3 mg/kg/day (n=5), GW501516 6 mg/kg/day (n=5) were administered, respectively, during the sixth to the ninth day after the second booster. Serum was diluted 2 x 104 times. (A) Serum high-affinity NP2-specific IgGs: GW501516 3 mg/kg/day (P=0.046), 6 mg/kg/day (P=0.013). (B) Serum total NP30-specific IgMs. GW501516 3 mg/kg/day (P=0.14), 6 mg/kg/day (P=0.045), compared with the control group. (C) Affinity maturation measured by dividing the NP2 specific IgM serum levels by the NP30 specific IgM serum levels. NP2/NP30: GW501516 3 mg/kg/day (P=0.03), 6 mg/kg/day (P=0.019). Results were analyzed with unpaired, two-tailed student t test and shown as mean. SEM. *P < 0.05, **P < 0.01, ***P < 0.001 and ns, no. significance. AU denotes arbitrary units.. 64.
(78) References Ademokun, Alexander A, and Dunn-Walters, Deborah (Sep 2010) , Immune responses: primary and secondary. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0000947.pub2] Adhikary T, et al. (2015), The Transcriptional PPARβ/δ Network in human macrophages defines a unique agonist-induced activation state. Nucleic Acids Research, 43(10): 5033–5051 Allen C, Okada T, Cyster JG (2015), Germinal center organization and cellular dynamics. Immunity, 27(2): 190–202 Barroso E, Eyre E, Palomer X, Vázquez-Carrera M (2011), The peroxisome proliferator-activated. receptor. β/δ. (PPARβ/δ). agonist. GW501516. prevents. TNF-α-induced NF-κB activation in human HaCaT cells by reducing p65 acetylation through AMPK and SIRT1. Biochemical Pharmacology, 15, 81(4):534-543 Baxter D (2007), Active and passive immunity, vaccine types, excipients and licensing. Occupational Medicine, 57(8): 552–556 Cho SH, et al.(2016), Germinal centre hypoxia and regulation of antibody qualities by a hypoxia response system. Nature, 537: 234–238 Coleman JD, Thompson JT, Smith RW III, Prokopczyk B and Vanden Heuvel JP (2013), Role of peroxisome proliferator-activated receptor and B-Cell Lymphoma-6 in regulation of genes involved in metastasis and migration in pancreatic cancer cells. PPAR Research 2013:121956. https:// doi: 10.1155/2013/121956 Cox RL (2017), Rationally designed PPARδ-specific agonists and their therapeutic potential for metabolic syndrome. Proceedings of the National Academy of Sciences of 65.
數據
+7

相關文件
To investigate the characteristics of Tsongkhapa’s meditation thought, the study is divided into five parts: (1) introduction, (2) Tsongkhapa’s exposition of meditation practice,
Then, we tested the influence of θ for the rate of convergence of Algorithm 4.1, by using this algorithm with α = 15 and four different θ to solve a test ex- ample generated as
Numerical results are reported for some convex second-order cone programs (SOCPs) by solving the unconstrained minimization reformulation of the KKT optimality conditions,
Particularly, combining the numerical results of the two papers, we may obtain such a conclusion that the merit function method based on ϕ p has a better a global convergence and
Then, it is easy to see that there are 9 problems for which the iterative numbers of the algorithm using ψ α,θ,p in the case of θ = 1 and p = 3 are less than the one of the
By exploiting the Cartesian P -properties for a nonlinear transformation, we show that the class of regularized merit functions provides a global error bound for the solution of
2019 冠狀病毒病的疫情仍在發展中。世界各地的 科學家不斷監測有關情況,包括病毒變種、接種疫
2. How would you say the following sentence? Write the stress level 1, 2, or 3 over each word. Draw a slash to show the thought groups. Circle the true statements about
