臺北縣三芝鄉淺水灣西側海岸
沙丘地形與植群分布關係之研究
A Study on the Relationship between the
Geomorphology and Vegetation Distribution
of the Coastal Dunes in the Western
Chianshuiwan, Sanjhih of Taipei County
陳彥樺
*李建堂
**Yen-Hua Chen Cheing-Tung Lee
Abstract
Coastal dunes are dynamic landforms which feature complex ecosystem as a result of the interaction among wind process, marine process, geomorphologic evolution and vegetation distribution. Geomorphologic field surveys were conducted twice in the Coastal Dunes on the west side of Chianshuiwan to produce digital elevation models (DEMs), which were then used to investigate the short-term geomorphologic changes within the study area. Meanwhile, vegetation distribution data was attained from surveys of 193 quadrats. After classifying the vegetation with two-way indicator species analysis (TWINSPAN), the relationship between the dune geomorphology and vegetation distribution was analyzed.
The result shows that the erosion and deposition changes of the dunes are due to factors including plant cover, shoreline alignment, beach width, and dune geomorphology. Elevation and the distance from the quadrat to shoreline are factors which affect the differences of the microenvironment within the vegetation zones.
* 國立臺灣大學地理環境資源學系碩士班研究生
Graduate Student, Department of Geography, National Taiwan University.
** 國立臺灣大學地理環境資源學系助理教授,通訊作者 (e-mail: [email protected])
Areas like the blowouts and slacks that are wetter and more sheltered can supply shrubs with more water and nutrients. Whereas in frontal areas like hummocks, wave erosion scarps, and beaches, pioneer herbs are dominant because they can adapt to frequent disturbances and high sand movement. As for the northern slopes, the steepest region in the study area, vines with flexile stems usually climbed upward on the steep and windward slopes. Other differences of vegetation distribution were also found.
As a conclusion, the geomorphologic characteristics led to some differences in coastal dune vegetation construction, composition, functions and species diversity. Therefore, different dominant communities could be found in various landforms.
Keywords: coastal dunes, digital elevation model (DEM), phytogeomorphology, two-way indicator species analysis (TWINPAN)
摘 要
海岸沙丘為動態變化相當快的的風成地形,受到風力作用、海水作用、地形 演育、植群分布等環境因子交互作用的影響,而呈現出複雜的生態系統。本研究 藉由兩次實地的地形量測產出淺水灣西側沙丘的數值高程模型,瞭解研究區沙粒 侵蝕及堆積的變遷情形,同時利用植物樣區法調查研究區內的植物分布狀況,在 調查的 193 個樣區植物資料則利用雙向列表比較法 (TWINSPAN) 進行植群分 類,藉以分析沙丘地形與植群分布之間的關係。 研究結果顯示,沙丘的侵蝕、堆積變遷情形主要受植物覆蓋、海岸線走向、 沙灘寬度、沙丘地形等影響,而高程、與海距離及地形特徵為影響植群型分布的 主要因子。在低凹的地形環境,包括風蝕窪地及低地,因為地表較接近地下水面, 環境較潮濕且背風有遮蔽,以灌木型的植物為主;在最前端近海的地形,包括圓 頂丘、侵蝕坡、沙灘等,因為受風、波浪、鹽霧的擾動較大,沙粒移動性較高, 以先驅性草本植物為優勢;在北側的沙丘斜坡地形,由於坡度較陡且迎風,攀緣 藤本較能適應地形較陡峭且迎風的斜坡脊部。 整體而言,海岸沙丘植群在不同地形因子影響下,植群結構、組成、功能、 多樣性等產生些許差異,且在不同地形環境中亦可發現有不盡相同的優勢族群。 關鍵字:海岸沙丘、數值高程模型 (DEM)、植物地形學、雙向列表比較法 (TWINPAN)前 言
海岸沙丘由於位在海陸交界之處,受到風力作用、海水作用、地形演育和植群分布等交互作用形 成的影響,可視為地形系統單元 (Goldsmith, 1989) 及生物系統單元 (Willis, 1989),或為生態複合系統 (Stallins and Parker, 2003),屬於動態變化快速且複雜的系統,極易受到擾動而發生改變。物質、能量 在系統內循環與流動,經由回饋機制與接鄰的環境進行交換 (Carter et al., 1990)。無論沙粒的移動、地形的演變及其上植群的生長等,都呈現出動態變化現象且其彼此間也有交互影響的關係,進而造成整 個沙丘系統的改變,因此長期以來即吸引許多地形學者、生態學者等從事相關議題的研究。 臺灣四周環海,全島海岸線長約1,200 公里,有海岸沙丘發育分布的海岸線長約 260 公里,約佔 全臺海岸線長的24%,由於受到東北季風的影響,海岸地區有廣大的沙丘分布,面積共約 50,000 多公 頃 (張政亮,1995)。過去臺灣在海岸沙丘方面的研究較偏重於自然營力、分布、規模、形態、沙粒粒 徑分析等地形學方面的研究 (如:連偵欽,1991;鄭瑞壬與鄧國雄,1991;石再添等,1992;1993; 1994;張政亮,1992;1995;李建堂,1998;張瑞津等,1998;張智原,2000;楊美萍,2004;林雅 谷,2005),以及海岸沙丘植物方面的研究,主要在防風定沙物種的研究,想要以人工方式來促進沙丘 的安定,以減少風沙對內陸造成危害 (如:甘偉航,1988;陳財輝等,2004),有關沙丘植群生態方面 的研究則較少 (如:章樂民,1971;黃守先,1974;杜興東,1975;柳榗,1975;郭長生,1977;陳 玉峰,1985;李國輝,1989),而針對沙丘地形與植物分布之間關係探討的研究則更是闕如。Nordstrom and Lotstein (1989) 曾提及需意識到沙丘的動態本質,不應只是聚焦於如何管理及穩定沙丘,而應研究 自然地景的改變過程。 國外研究顯示,沙丘植物會影響到沙丘地形的發育,主要因沙丘植物具有定沙、攔沙及堆沙的功 能,而且植物的生長型會影響到微氣候的改變,進而影響到沙粒的移動變化,因而形成不同的沙丘形 態及其演育 (柳榗,1975; Davies, 1980; Pethick, 1984; Carter et al., 1990; Hesp, 1989;2002)。另一方面, 在不同環境因子及干擾類型的影響下,海岸沙丘的環境特性會有所差異,進而形成不同的植群分布及 物種組成 (Doing, 1985; Hayden et al., 1995; Avis and Lubke, 1996; Costa et al., 1996; Musila et al., 2001; Abuodha et al., 2003; Stallins and Parker, 2003)。
臺灣地區的海岸沙丘由於長期受到人類活動的干擾,幾乎已無完整的天然海岸沙丘地。臺北縣淺 水灣西側的海岸沙丘,由於人為活動干擾程度較小,尚保有較接近天然的沙丘地形及植物分布,因此 本研究以此海岸沙丘為研究區,分別調查短時距的地形變遷、植群分布狀況,進而分析海岸沙丘地形 及植群分布之間的關係。
研究區概述
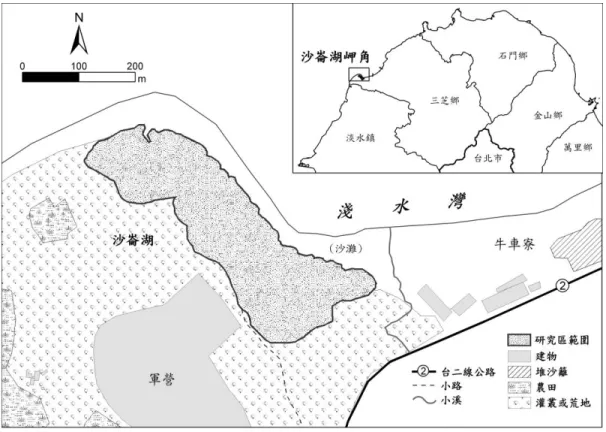
淺水灣位在臺北縣三芝鄉後厝村,沙崙湖岬角東側的海灣 (圖 1),為三芝鄉沙灘面積最大的海岸 (三芝鄉公所,1994)。研究區即位在淺水灣西南岸,靠近沙崙湖岬角的海岸沙丘,靠內陸範圍的界定 主要以黃槿、林投等植物的分布為邊界,面積共約 6.7 公頃,研究區靠海的灘岸主要為藻礁所構成, 並散布著大、小不等的火山岩塊。 淡水河口北側海岸主要的構成物質為大屯火山群的火山岩屑堆積物,沙崙湖附近的沙源部份來自 於淡水河的沉積物,可能為離岸流以滾動的方式帶到此處堆積,其沙粒呈現雙峰的粒徑分布,代表有 兩種沉積物來源,可能為沿岸流及離岸流同時搬運沉積的結果 (吳偉健,2002)。由於淺水灣的海灣迎 向東北季風的吹襲,又有八連溪、海尾溪、北勢坑溪、淡水河等溪流提供沙粒來源,海流將沉積物搬 運至附近堆積。強烈海風將海底的沉積物搬運至岸上堆積,因而於牛車寮、沙崙湖一帶形成海岸沙丘。 三芝鄉的降雨豐沛,近於冬雨型,冬季多為陰沉有雨的天氣,濕度大,日照率低 (三芝鄉公所,圖 1 研究區位置圖 Fig. 1. Location of study area.
1994)。統計鄰近三芝鄉的淡水氣象測站過去十年來 (1997 年 4 月到 2007 年 3 月) 的每日平均風速及 風向資料結果顯示,此區域冬天主要受強烈的北風影響,夏季風向則較為不固定,以東南風為主、西 風為輔。在地形調查期間內 (2006 年 9 月到 2007 年的 3 月) 的風力以北風最為強盛,東北風及東南風 為其次。 研究區後方的最高處為軍營,在研究區內還殘留著以前的軍事碉堡。在研究區東側的沙丘則設有 定沙的木樁,其後方沙丘並有木麻黃防風林,而且該地區目前飆沙、衝沙活動相當盛行,因此研究區 可能多少受到近期人為活動的干擾所波及。為此再進一步比對研究區在第一、二和三版相片基本圖中 的狀況,發現研究區確實是受到一些人為的干擾,主要是多了一些小路徑,然而整體地形並沒有產生 明顯或大範圍的變動,因此研究區大體仍保有較天然的沙丘地形。
研究方法
(一) 地形調查
參考Andrews et al. (2002) 於以色列進行海岸沙丘小尺度地形變遷調查及分析方法,本研究地形 野外測量是利用全測儀 (Total Station) 測得研究區的高程樣點資料,再以 Kriging 進行內插生產出數值高程模型 (digital elevation model, DEM)。本研究分別於 2006 年 9-10 月及 2007 年 4 月各量測一次全區 的沙丘地形高程資料,再利用兩次不同時期所產出的DEMs 得到不同時期的高程變遷資料,以分析短 時距沙丘地形的變化情形,並藉以取得樣區的地形參數資料。
(二) 植群調查方法
植群調查主要採用樣區調查法,雖然研究區的植被主要以草本及灌木為主,但是物種組成的異質 性相當高,若以系統取樣方法調查,常會漏掉一些重要的植群型 (蘇鴻傑與劉靜榆,2004),因此本研 究採用歐陸學派的取樣方法 (劉棠瑞與蘇鴻傑,1983),主要視研究區內植群的分布狀況而選擇樣區, 先判釋研究區內不同的植群分布狀況,然後在各植群內再決定設置樣區的位置及數量。本研究總共設 置193 個植物調查樣區,在草本植被區的樣區大小為 1×1 平方公尺,計有 180 個樣區,灌木植被區的 樣區大小則為4×4 平方公尺,計有 13 樣區。在每個調查樣區內分別記錄物種及每個物種的覆蓋度等級, 共分成十個等級 (0:無;1:<0.1%一株或少數;2:>0.1~<1%偶而出現;3:1~4%;4:5 %;5:6 ~ 15 %;6:16 ~ 25 %;7:26 ~ 50%;8:51 ~ 75%;9:76 ~ 100%)(修改自蘇鴻傑和劉靜榆,2004),同時 利用GPS 將樣區的座標記錄下來,以便在 ArcGIS9.0 中呈現出樣區的分布狀況,並取得各樣區的地形 參數資料。(三) 植群分類
植群分類是將植物組成相似的樣區合併,組合成植群型,本研究採用雙向列表比較法 (two-way indicator species analysis, TWINPAN),來進行植群分類,這是仿照歐陸學派的列表比較法,主要依據分化種的出現與否來合併樣區,並選擇植群型的特徵種,本研究利用 PC-ORD Version 4.14 軟體中的 TWINSPAN 程式進行分析,以得到研究區內不同的植群型。
(四) 地形與植群關係的分析
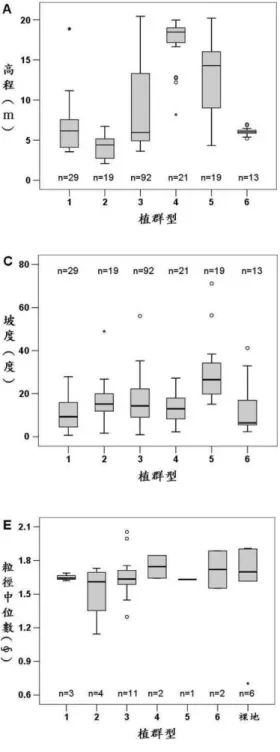
環境因子主要有高程、坡向、坡度、與海距離、高程變遷、粒徑大小等地形參數,利用盒型圖進 行各植群型樣區的地形參數比較,並統計迎風∕背風坡及地形變遷堆積∕侵蝕出現的次數或比例。結果與討論
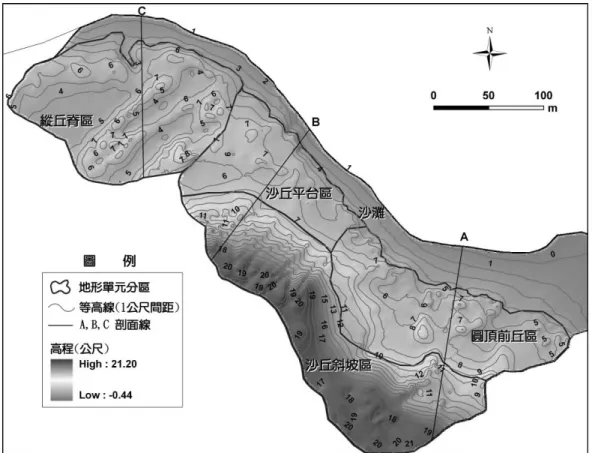
(一) 沙丘地形分區及其特徵
第一次地形調查所產出的DEM 等高線地形圖如圖 2 所示,研究區內的高度多介於 0~20 公尺間, 地勢南高、北低,往東北方傾斜,坡向大多朝向北方及東北方,沙丘脊部及低地的走向多為東北-西 南向,與盛行的東北季風平行,呈縱沙丘的形態,而縱沙丘的形成需要時間,更需要單一風向的強風 (Davies, 1980),此可能與淺水灣的沙丘形成時間較久,植被豐富、較為穩定,又長期受東北季風的影 響所致。研究區內的沙丘地形極具多樣性,有圓頂丘 (hummocks)、侵蝕坡 (wave erosion scarps)、風蝕窪 地 (blowouts)、沙丘平臺 (platforms)、沙丘脊 (dune ridges)、沙丘低地 (dune slacks)、拋物線形沙丘
(parabolic dunes) 等。依主要的地形特徵,研究區可區分為四類不同的沙丘地形單元,包括圓頂前丘區、 沙丘斜坡區、縱丘脊區和沙丘平臺區 (圖 2)。
圖 2 研究區第一次調查的等高線地形圖及地形單元分區
Fig. 2. Contour map and geomorphologic units of study area after the first survey.
1. 圓頂前丘區:位於研究區的最東側,沙灘面積較廣,沙源供應較多,海風攜帶沙粒被植物攔截 而堆積成許多不連續的圓頂沙丘,較靠近海岸的圓頂丘的高度小於1 公尺,散布在此區前方;距海較 遠處的圓頂丘則高於2 公尺,會順著東北-西南方向羅列。圓頂丘的頂端有植物覆蓋,其餘則為裸沙 地,幾無植被生長。 2. 沙丘平臺區:位在研究區的中段,沙灘呈狹窄形,寬僅約 10 公尺,沙丘從沙灘順著侵蝕坡上 升,侵蝕坡有明顯的切割面,主要受到波浪侵蝕所致,沙丘的頂端與海岸線平行。此區為地勢起伏最 緩的一區,高程約5~7 公尺,坡度多在 10%以下。 3. 縱丘脊區:位在研究區的最西側,此區主要特徵為 2 道呈直線形並列的沙丘脊,外型為拋物線 沙丘的拖曳臂,沙丘長軸走向與東北季風平行,呈東北-西南走向。沙丘脊部高約2~3 公尺,一道拖 曳臂部長約100 公尺。此地形區內約有 4 道沙丘脊,3 道沙丘低地位在沙丘脊間,呈現脊部-低地並 排,脊部兩翼的坡向多為東南∕南方或西北∕北方。低地的靠海前端多為風蝕窪地。
4. 沙丘斜坡區:此區為陡升的地勢,位於研究區靠內陸的最後方,海岸風沙向岸吹移過程中,遇 到了山體 (沙崙湖岬角) 阻擋風沙順坡上爬堆積所形成的沙丘。坡向主要朝北方至東方,正迎冬天的 盛行風向,在60 公尺的水平距離內,高度差約 11 公尺,坡度約 24%,有十幾條裸露的沙丘低地與富 有植生的沙丘脊參差並列,呈東北-西南走向。長條狀的沙丘脊長約50~60 公尺,寬從 2~20 公尺不等; 沙丘低地長度亦約50~60 公尺,寬約 2 公尺,深約 1~2 公尺。北側斜坡區相對南側而言,沙丘脊與低 地分布較密集且較陡峭,為近期的人為活動開闢出來的路徑。而南側斜坡區的沙丘脊部與低地並列, 呈現2~3 道拋物線形沙丘並連,其拖曳臂區 (脊部) 則相接在一起。
圓頂丘和拋物線沙丘是潮濕地區最為常見有植生分布的沙丘地形 (Pye and Tsoar, 1990)。圓頂丘的
高度通常小於10 公尺,其形態的變異與生長植物的特性 (如:形狀、密度、生長特徵) 與風力侵蝕狀
況有關,常會形成一系列平行濱線而不連續的分布,但有時圓頂丘也會呈不規則性分布 (Pye and Tsoar, 1990)。研究區內靠海前方的圓頂丘規模較小,呈不規則分布於沙丘地上,其上所生長的植物種類主要 為草本的蔓荊和濱刺草,其中,濱刺草分布的沙丘較蔓荊所在沙丘還高,這與此兩種植物的生長形態 不同有關。後方圓頂丘的規模則較前方大,由數個較大型的圓頂丘接連排列而成,呈東北-西南走向, 主要的植生分布為林投灌木叢。
Pye and Tsoar (1990) 認為拋物線沙丘的特徵為 U 或 V 型,擁有 2 道拖曳臂區,而其尖端是朝向 上風處,在拋物線沙丘的下風處大部分會有較陡的滑動面。研究區內的拋物線沙丘主要分布在縱丘脊 區和沙丘斜坡區兩地形單元區內,分別呈現出兩種不同的類型 (圖 3)。在縱丘脊區的拋物線沙丘為單 一形態呈髮夾狀的拋物線沙丘 (圖 3A),此為研究區內縱丘脊區的主要地形特徵,其中拋物線沙丘吹 蝕廊道 (deflation corridor) 的長軸長約 75 公尺,寬約 25 公尺,朝 45 度的方向。沙丘斜坡區內的拋物 線沙丘主要分布在其南側,有2 個比較明顯且相連的拋物線形沙丘,其吹蝕廊道的長軸都約 60 公尺, 主要有3 道沙丘拖曳臂 (圖 3B)。相較於縱丘脊區,沙丘斜坡區內拋物線沙丘的地勢較高,且較為陡斜, 圖 3 研究區內兩種不同類型的拋物線沙丘 Fig. 3. Two different types of parabolic dune in the study area.
植物只分布在拋物線沙丘的脊部,吹蝕廊道地區則為裸露地。由於縱丘脊區的拋物線沙丘已為植物所 覆蓋,因此穩定性較高,相對地,在沙丘斜坡區的拋物線沙丘,其吹蝕廊道仍為裸地,屬於較不穩定 的地區,因此未來地形的變化程度應會較大。
Pye and Tsoar (1990) 認為拋物線沙丘幾乎都是從有植生分布的風蝕窪地發育而來,若植被遇到干 擾時,如:植生被破壞、放牧、踩踏、病害或土壤性質改變等,便會加速風蝕窪地的形成。一旦沙地 表面結皮或植物根叢的孔隙擴大時,擾動的漩渦流會促進凹洞的擴大,進而擴大風蝕窪地的範圍,而 從風蝕窪地所侵蝕搬運的沙粒,會因背風坡植群的攔截而堆積下來,而逐漸增加拋物線沙丘的高度。 研究區內的風蝕窪地主要出現在沙丘平臺區和縱丘脊區的侵蝕坡,此處的侵蝕坡是指最前方的沙丘因 波浪切割坡腳所形成的斜坡,當侵蝕坡持續受風、波浪、人為等干擾而破壞其上生長的植被時,便會 加速風蝕窪地的形成。當風蝕窪地逐漸擴大後,反而會促使風、波浪等營力沿著風蝕窪地的缺口入侵 內地,因此風蝕窪地便會持續受到侵蝕而逐漸形成吹蝕廊道,並往內延伸成長軸狀,此作用即為主導 縱丘脊區內單一拋物線沙丘的形成原因,研究區內吹蝕廊道的延伸方向約與冬季東北季風的盛行風向 一致。 在沙丘斜坡區內拋物線沙丘的前方有圓頂丘的散布,由於圓頂丘間有較大範圍的裸地,氣流順著 此裸地向內陸吹襲,因此在裸地上並無植被生長,而原本的斜坡處由於有部分植生的固定,因而形成 較穩定的脊部。在圓頂前丘區與沙丘斜坡區交界處的裸地,由於地形的改變,坡度陡、高度驟升,氣 流在此遇到驟升的迎風坡,朝頂端行進時受地形壓縮而加快風速,特別在迎風面凹坡的風速會最快, 過了頂端到達背風面時,則因氣流分散,所攜帶的沙粒會堆積於背風面 (Pye and Tsoar, 1990),因而逐 漸形成研究區內的拋物線沙丘。
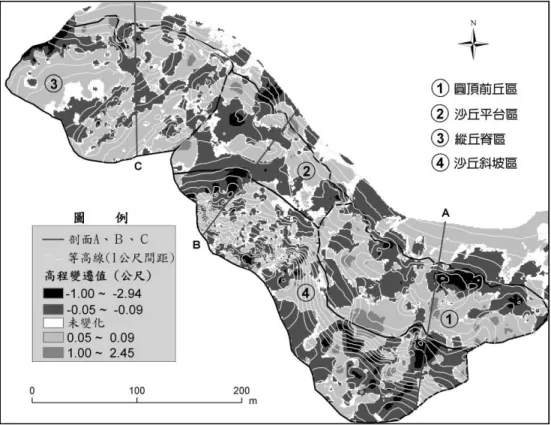
(二) 沙丘地形的短時距變遷
研究區沙丘地形短時距的高程變遷及高程剖面變遷分別如圖4、圖 5 所示,沙丘侵蝕與堆積的方 向大致朝南至西南方,主要應是受研究期間內盛行的東北季風影響所致。高程及侵蝕∕堆積的體積變 化最明顯的地區是在圓頂前丘區和沙丘斜坡區,尤其是圓頂前丘區的東側和其後方所連接上去斜坡區 的南側,這是研究區內侵蝕與堆積變遷最顯著的地區。 由於圓頂前丘區的前方沙灘位於海灣最凹處,擁有平坦且較寬的沙灘,波浪、沿岸流攜帶的沙粒 先在此堆積,再藉由盛行季風將沙粒往內陸攜帶。途中沙粒在此區受到植群的攔截而堆積下來,成為 沙源最充沛的地區。裸沙地為沙粒輸送的廊道,為堆積和侵蝕的主要所在,是沙粒移動性最高的一區, 圓頂前丘區的沙丘裸地比例相當高,特別是在最前端的圓頂丘,因冬季蔓荊的莖葉枯萎且地勢高起, 在迎風面受風侵蝕的沙粒易被搬往後方堆積,其後退可達20 幾公尺 (圖 5)。 沙丘斜坡區因為距海較遠,沙源補充不若前方的沙丘多,且因為坡度陡、坡向朝北及東北方,受 東北季風影響更為明顯,風力侵蝕大於堆積,因此沙丘斜坡區主要為侵蝕的地區,尤其南側的侵蝕與 堆積變動情形最明顯。侵蝕主要發生在裸露的凹坡,而在有植被的脊部則大多呈堆積狀況。由於凹坡 為拋物線形沙丘的吹蝕廊道,且前方的裸地比例較高,風速較不易受植被影響而減小,氣流會順著凹 坡向上升,也由於地勢驟升的緣故,氣流受壓縮而加速向頂端侵蝕,導致沙丘斜坡區內裸露凹坡的侵 蝕情形最為明顯。斜坡區北側由於植被覆蓋的比例較高,因此侵蝕與堆積變遷不若南側明顯。說明:負值為侵蝕,正值為堆積,∣Value∣≦0.05公尺代表沒變化。
圖 4 兩次地形調查的高程變遷圖 Fig. 4. Elevation changes between two field surveys.
在沙丘平臺區及縱丘脊區最前端的侵蝕坡,由於距海較近,能接受到沙源供給,大多呈堆積的結 果,但在風蝕窪地則出現侵蝕情形,主要因沒有植群攔截沙粒,且坡度較陡導致風速加快而侵蝕,因 而呈現出侵蝕的結果。沙丘平臺及縱丘脊區前方的外灘有約30~50 公尺寬的藻礁分布,其上有火山岩 塊散布,沙源的補充受到阻礙而減少,且因沙灘較為狹窄,較沒有適當的堆積環境,所以不若圓頂前 丘區有較多的沙粒堆積量。在沙丘平臺區後方有較大面積的侵蝕區域,推測因其前方有東西向生長的 灌木叢分布,沙粒會被灌木叢攔截而堆積下來,因此位於其後方的地區,在沙源供給有限、植被稀疏 下,會持續受到風力侵蝕。縱丘脊區的沙丘植物覆蓋最為完整,沙丘較為穩定,除了在其前方的風蝕 窪地有較明顯的變動外,其餘地區均未有明顯的變遷,為地形變化最少、最穩定的一區。 Carter et al. (1990) 認為有些沙丘系統擁有兩類不連續的沙丘地形,分別呈現穩定及活動沙丘鑲嵌 的複雜類型,這是因為波浪和風的作用造成小區域的沙粒堆積、侵蝕或重新分配所致,這樣的地形多 樣性對維持系統的半穩定性很重要。Goldsmith (1989) 認為風沙的輸送情形會隨地域不同而改變,會 受當地的植群、沙灘坡度、沙灘寬度、海岸線走向、當地地形等因子的影響。研究區內的沙丘擁有多 樣的地形組成,不同地形單元的侵蝕、堆積變遷情形,主要應是受到植物覆蓋、海岸線走向、沙灘寬 度、沙丘地形等的影響,而呈現出不同的特性,這應該有助於研究區內沙丘系統的穩定。
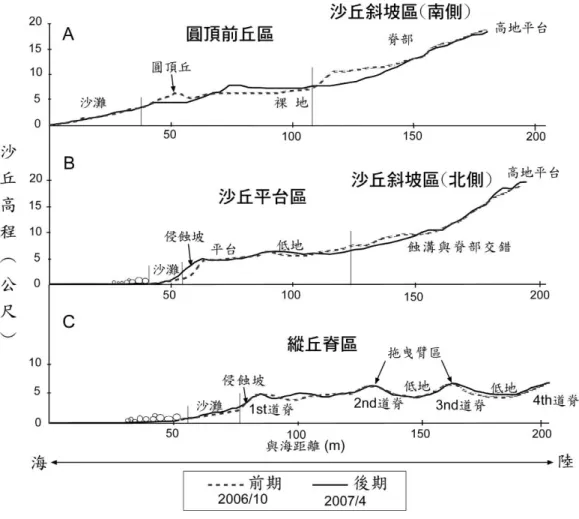
圖 5 兩次地形調查的高程剖面變遷圖 (剖面位置參見圖 4)
Fig. 5. Profiles of elevation change between two field surveys. (Location of profiles see Fig. 4.)
(三) 植群分布與地形的關係
研究區內共記錄到42 科 73 種植物 (附錄),在 193 個植物樣區內則記錄到 29 科 53 種植物,物種 多具有匍伏地下莖、節節生根的特性,以適應沙埋的環境,其中以白茅、濱艾為研究區內最優勢的植 物。依TWINSPAN 分類,研究區內的植物社會可分為六種植群型,分別為狗牙根-裂葉月見草型、濱 刺草-天蓬草舅型、白茅-濱艾型、大花咸豐草-濱豇豆型、槭葉牽牛-雞屎藤型、林投-海桐型。 各植群型的分布位置如圖6 所示,而樣區不同植群型的位置在不同的高程及與海距離的影響下大致呈 現出帶狀分布。 盒型圖所呈現出各植群型地形參數的集中及離散程度如圖7 所示,質性統計資料則如表 1 所示。 大花咸豐草-濱豇豆型的高程及與海距離幾乎都大於其他類型,槭葉牽牛-雞屎藤型次之,此兩植群 型都分布在距海較遠的沙丘斜坡區;而濱刺草-天蓬草舅型的高程及與海距離則小於其他類型,屬於 最靠近海岸的植群,另外其他三型則是介於中間。雖然高程與距海遠近決定了整體植群型分布的變異圖 6 不同植群型樣區的分布位置
Fig. 6. Distribution map of sampled quadrats classified by vegetation types.
結果,但其他的地形參數對植物分布多少也有些影響,以下分別就植群型偏好出現的沙丘環境及植物 生長特性進行討論。 1. 狗牙根-裂葉月見草型 為人為干擾頻繁的植群型,主要分布在人車干擾較頻繁的平坦路徑兩側,多為適應踩踏的物種生 存,如狗牙根、鋪地黍、牛筋草及裂葉月見草等。狗牙根、鋪地黍為常出現在平野地的野草,而裂葉 月見草為北部海岸逸散的外來種,適合生活在海邊沙地。而這些物種易受人為干擾而影響其分布,過 去已有一些海岸沙丘研究證實外來種常出現在人為干擾多的地區 (Greipsson, 2002; Kim, 2005),這現象 在研究區內亦可發現,一些人為干擾較多的區塊,常有一些歸化種 (naturalized species) 的分布,如裂 葉月見草、大花咸豐草、銀合歡、槭葉牽牛等。 2. 濱刺草-天蓬草舅型 為沙地前端先驅植群型,主要有濱剪刀股、濱箬草、天蓬草舅、濱刺草等沙丘先驅草本植物,受 到高度和與海距離的影響顯著,其分布較其他植群型來的低且近海,主要分布在最前端的圓頂丘、侵 蝕坡和沙灘上,其所在的沙粒粒徑較大,顯示其生長之處的水分留存最差。因為其近海位置,易受強 風、沙埋、侵蝕及波浪等干擾,植物卻能在此擾動相當大的地方生存,進而穩定沙丘,為最能適應海 岸最前端擾動大、高鹽霧的環境,因而此型的物種數量最少,最具有沙地前端的指標性質,表示沙灘 和沙丘的交界處 (Carter et al., 1990)。
說明:植群型代號:1:狗牙根-裂葉月見草型;2:濱刺草-天蓬草舅型;3:白茅-濱艾型;4:大花咸豐草-濱 豇豆型;5:槭葉牽牛-雞屎藤型;6:林投-海桐型。 圖 7 不同植群型地形參數的盒型圖
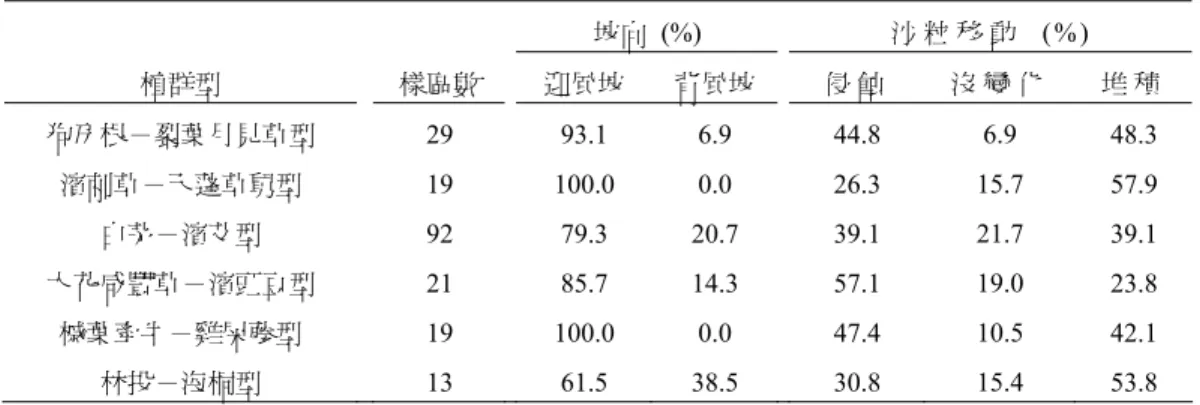
表 1 植群型所佔迎風∕背風及侵蝕∕堆積的比例
Table 1. Percentages of windward/leeward and erosion/deposition in the six vegetation types.
坡向 (%) 沙 粒 移 動 (% ) 植群型 樣區數 迎風坡 背風坡 侵 蝕 沒 變 化 堆 積 狗牙根-裂葉月見草型 29 93.1 6.9 44.8 6.9 48.3 濱刺草-天蓬草舅型 19 100.0 0.0 26.3 15.7 57.9 白茅-濱艾型 92 79.3 20.7 39.1 21.7 39.1 大花咸豐草-濱豇豆型 21 85.7 14.3 57.1 19.0 23.8 槭葉牽牛-雞屎藤型 19 100.0 0.0 47.4 10.5 42.1 林投-海桐型 13 61.5 38.5 30.8 15.4 53.8 3. 白茅-濱艾型 為研究區內分布最廣泛的一型,地形參數的離散程度也最大,其優勢的白茅、濱艾、蔓荊、馬鞍 藤為研究區典型的沙地植物,在較為穩定的沙丘平臺、拋物線沙丘脊部及低地均可見其分布。 4. 大花咸豐草-濱豇豆型 為沙丘高地平臺型,主要由濱豇豆、雙花蟛蜞菊、大花咸豐草等所組成。樣區與海距離的資料較 為集中,分布較侷限在後方斜坡區中段朝東的斜坡和其後方平臺上,為植群型分布位置最高且離海最 遠的一型,主要出現在斜坡後方坡度較緩處或有遮蔽處的凹坡。濱豇豆為豆科植物,根部與根瘤菌共 生,可行固氮作用,能定沙且改善沙土的貧脊,提供後方內陸物種較多生存所需的無機氮。 5. 槭葉牽牛-雞屎藤型 為陡坡攀緣型,組成以雙花蟛蜞菊、雞屎藤、槭葉牽牛、串鼻龍等攀緣性藤本為主,此型主要分 布在斜坡區北側沙丘脊上,坡度顯著大於其他植群型,而高程僅次於大花咸豐草-濱豇豆型。由於斜 坡陡峭,較不利於直立的草本生長,而此型優勢的蔓藤植物莖部柔軟,極易延長生長且擁有蔓莖或卷 鬚攀爬而上,但需依賴支持物供其向上生長,因此能適應地形非常陡峭上升且迎風的斜坡脊部,較陡 的地勢也是區別此型與沙丘斜坡區另外主要兩型 (白茅-濱艾型和大花咸豐草-濱豇豆型) 在空間分 布差異的主因。 6. 林投-海桐型 為凹地灌木型,主要為林投、海桐、銀合歡等灌木所組成,分布於圓頂前丘區的沙丘背風處、低 凹地、沙丘平臺區的背風低凹地及縱丘脊區的風蝕窪地等,在研究區分布的高程非常集中於6 公尺附 近,由於其生長的環境地勢低凹,較接近地下水面潮濕的環境,因而能支持木本植物生長。在圓頂前 丘區的低凹地常為林投等灌木型植物生長,這可能與Pye and Tsoar (1990) 所提到的作用有關,由於圓 頂丘大都形成於植物覆蓋不連續和有強風之處,使得相鄰圓頂丘間的凹地會逐漸擴大成更深的低槽, 因而較接近地下水面,反而有利於林投等木本植物的生長,而沙粒在林投等木本植物附近開始進行堆 積,進而形成較大型的圓頂丘。在縱丘脊區最前方的風蝕窪地,由於其凹狀地形會促使氣流漩渦加速 侵蝕,使窪地加大、加深,直到接近地下水面為止 (Pye, 1982),因此會有利於支持此類型的植物生長。
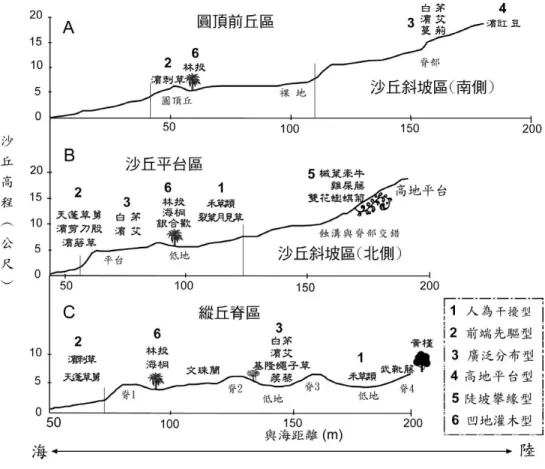
Stallins and Parker (2003) 認為海浪沖刷及淹沒頻率較小的沙洲島,因為地形起伏變化較多樣化, 棲地異質度較高,可以共存更多物種,因此物種結構較為豐富,物種多樣性會遠較於干擾較多的沙丘 高,而Hayden et al. (1995) 也認為地表高程的異質性是海岸植群呈現區塊分布的主要原因。研究區內 不同地形單元與植群型分布的關係如圖8 所示,縱丘脊區的物種豐富度最高,這可能與沙丘地形的多 樣性較高、環境較為穩定有關,而物種次多的沙丘斜坡區同樣也擁有較多樣的地形,但因沙粒移動性 較高且受到人為的干擾較大,因而減少其物種數。沙丘平臺區雖然屬於較穩定的環境,但其地形變化 較少,因此其物種數並不如前兩區多,但是單位面積的物種數卻是最多的。圓頂前丘區的沙粒移動性 最高、環境相對屬於最不穩定的地區,因此其物種數也最少。 圖 8 不同地形分區剖面的植群分布圖 (剖面位置參見圖 4)
Fig. 8. Vegetation types of different geomorphologic profiles. (Location of profiles see Fig. 4.)
結 論
本研究利用全測儀兩次測量研究區的地形,藉由GIS 產出 DEMs,以了解沙丘地形地形特徵及短
時距的變遷情形,同時也藉以取得植物樣區所在的地形參數,並利用樣區調查研究區的植物分布情形, 以分析研究區內地形與植物分布之間的關係。
淺水灣的西側海岸沙丘地形高程介於 0~20 公尺,在風、植物及其他外在營力作用下產生多樣的 沙丘地形,包括:圓頂丘、侵蝕坡、風蝕窪地、沙丘平臺、沙丘脊、沙丘低地、拋物線沙丘等。研究 區依地形特徵主要可區分成圓頂前丘區、沙丘平臺區、縱丘脊區和沙丘斜坡區四類不同的地形單元區。 圓頂前丘區主要為圓頂丘所組成,分布於沙源供應充足的海岸前端,海風攜帶沙粒被植物攔截而堆積 成許多不連續的圓頂沙丘。若植物生長較旺盛或分布較密時,有助於沙粒的攔截與堆積,因而會形成 較廣大的沙丘平臺,研究區的沙丘平臺區高程介於5~7 公尺,地勢起伏很小。縱丘脊區最主要的地形 特徵為單一髮夾狀拋物線沙丘,可能是發育自侵蝕坡的風蝕窪地,因受東北季風長期的影響所致,其 吹蝕廊道的長軸多朝向東北方。而沙丘斜坡區的地勢較陡升,主要為沙粒沿著岬角山體向上爬升堆積 形成,甚至到20 公尺高的平臺處仍有沙丘分布。沙丘斜坡區南側主要為耙子狀的拋物線沙丘,由於地 形陡升,氣流受地形壓縮加速侵蝕所形成的沙丘地形。 由兩次地形調查的變遷分析顯示,圓頂丘、侵蝕坡等前端沙丘及沙丘斜坡的坡腳處變化較為明 顯,此與東北季風、沙丘地形坡度∕坡向、與海遠近、植被覆蓋等因子有關,特別在一些裸地面積較 大、地勢較陡、迎風坡的地區,變遷最為明顯,如圓頂前丘區前端的小型沙丘,被風侵蝕的沙粒往內 陸搬運堆積,移動距離近20 公尺。沙丘斜坡區的坡腳則因為坡度驟升且植被稀疏,氣流受壓縮而加速 向頂端侵蝕,導致此區的侵蝕量是最多的。侵蝕坡和圓頂丘皆因距海較近,較易遭受風力和波浪侵蝕 而消失,因此變動也較為明顯。縱丘脊區則因植被覆蓋較完整、干擾較少,相對環境較為穩定,因而 地形變遷最不明顯。 地形參數盒型圖及植群分布圖顯示,高程與距海遠近為影響植群呈帶狀分布的主要因子,坡度、 坡向、高程變遷、粒徑等地形參數則影響到少數植群類型的分布。研究區內的植群大致可區分為六類: (1) 狗牙根-裂葉月見草型:為人為干擾頻繁的植群型,主要出現在路徑兩旁;(2) 濱刺草-天蓬草舅 型:為沙地前端先驅型,生長於圓頂丘、侵蝕坡、沙灘等前端環境,因為沙源豐富且有植被生長,能 堆積沙粒、適應沙埋環境,擾動的環境使其物種較稀少卻居於優勢;(3) 白茅-濱艾型:為最廣泛分 布的一型,是研究區最典型的植群型;(4) 大花咸豐草-濱豇豆型:為沙丘高地平臺型,分布於距海 最遠之處;(5) 槭葉牽牛-雞屎藤型:為陡坡攀緣藤本型,攀爬而上的特性使其能適應坡度非常陡峭 且迎風之斜坡;(6) 林投-海桐型為凹地灌木型:長期受風吹蝕所形成的低凹地較接近地下水面,凹 地的環境可累積水分、有機質,可提供較大型的灌木生存所需的水分及養分。 海岸沙丘為一複雜的生態系,其內生物與非生物的組成相互影響,本研究結果顯示在這小尺度的 時空範圍內,地形梯度及特徵與植物分布具有相互影響的結果。本研究為一短時距、小範圍的植物地 形學研究,未來會持續進行長時期的調查與監測,期望對於臺灣地區海岸沙丘環境以及沙丘地形和植 物的交互作用能有進一步的瞭解,以提供從事沙丘環境復育之參考。
引用文獻
三芝鄉公所 (1994) 三芝鄉志,臺北:三芝鄉公所。 石再添、張瑞津、林雪美、張政亮、連偵欽 (1992) 臺灣北部海岸沙丘之研究,國立臺灣師範大學地 理研究報告,17: 147-167。石再添、張瑞津、林雪美、張政亮、連偵欽 (1993) 臺灣西部海岸沙丘之地形學研究,國立臺灣師範 大學地理研究報告,19: 104-139。 石再添、張瑞津、林雪美、張政亮、連偵欽 (1994) 臺灣南端與東部海岸沙丘之地形學研究,國立臺 灣師範大學地理研究報告,21: 2-34。 甘偉航 (1988) 海岸竹枝籬堆砂功效評估及植物是砂功能調查,林業試驗所研究報告季刊,3: 225-240。 吳偉健 (2002) 淡水河口海岸帶地形地貌及沉積物特性,國立臺灣大學海洋研究所碩士論文。 李建堂 (1998) 對數-雙曲線分佈於粒度分析的應用:老梅前濱沙丘個案研究,中國地理學會會刊, 26: 129-142。 李國輝 (1989) 彰化縣沙丘植群之研究,國立中興大學植物研究所碩士論文。 杜興東 (1975) 桃園海岸砂地植物之生態研究,國立臺灣大學森林學研究所碩士論文。 林雅谷 (2005) 風沙與地形變動之試驗研究,國立成功大學水利及海洋工程研究所碩士論文。 柳榗 (1975) 臺灣西海岸沙丘生態之研究,臺灣省林業試驗所報告,266: 1-16。 連偵欽 (1991) 臺灣北部海岸福隆和富貴角地區沙丘之研究,國立臺灣師範大學地理學研究所碩士論 文。 張政亮 (1992) 蘭陽平原海岸沙丘之地形學研究,國立臺灣師範大學地理學研究所碩士論文。 張政亮 (1995) 臺灣海岸沙丘之地形學研究,私立中國文化大學地學研究所博士論文。 張智原 (2000) 臺灣西北部海岸變遷之研究-淡水河口至新竹頭前溪口段,私立中國文化大學地學研 究所地理組碩士論文。 張瑞津、石再添、陳翰霖 (1998) 臺灣西南部嘉南平原的海岸變遷研究,國立臺灣師範大學地理研究 報告,28: 83-106。 郭長生 (1977) 風吹沙地區沙地植物生態調查,科學教育,7: 48-52。 陳玉峰 (1985) 墾丁國家公園海岸植被,屏東:墾丁國家公園管理處。 陳財輝、游漢明、洪富文 (2004) 桃園許厝港海岸飛砂之移動及定砂植物之生長,中華林學季刊,36: 367-377。 章樂民 (1971) 臺灣西海岸鹽溼地之植物地理與生態之研究,臺灣省林業試驗所報告第 213 號。 黃守先 (1974) 臺灣之沙丘及其植物社會,郭宗德、周昌弘、張崑雄、林飛棧主編,生物與環境專題 研討會講稿集,臺北:中央研究院生物研究中心,109-117。 楊美萍 (2004) 桃園縣海岸地形變遷之研究,國立臺灣大學地理環境資源研究所碩士論文。 鄭瑞壬、鄧國雄 (1991) 桃園臺地海岸沙丘型態與沙粒度分析,中國地理學會會刊,19: 21-31。 劉棠瑞、蘇鴻傑 (1983) 森林植物生態學,臺北:臺灣商務印書館。 蘇鴻傑、劉靜榆 (2004) 論植相社會學之植群分類法,臺大實驗林研究報告,18 (3): 129-152。 Abuodha, J. O. Z., Musila, W. M. and van der Hagen, H. (2003) Floristic composition and vegetation ecology
of the Malindi Bay coastal dune field, Kenya, Journal of Coastal Conservation, 9: 97-112.
Andrews, B. D., Gares, P. A. and Colby, J. D. (2002) Techniques for GIS modeling of coastal dunes,
Geomorphology, 48: 289-308.
South Africa, Landscape and Urban Planning, 34: 237-254.
Carter, R. W. G., Nordstrom, K. F. and Psuty, N. P. (1990) The study of coastal dunes. In: Nordstrom, K., Psuty, N. and Carter, B. (eds.) Coastal Dunes: Forms and Process, New York: John Wiley & Sons, 1-14. Costa, C. S. B., Cordazzo, C. V. and Seeliger, U. (1996) Shore disturbance and dune plant distribution,
Journal of Coastal Research, 12: 133-140.
Davies, J. L. (1980) Geographical Variation in Coastal Development, 2nd ed., New York: Longman.
Doing, H. (1985) Coastal fore-dune zonation and succession in various parts of the world, Vegetatio, 64: 65-75.
Goldsmith, V. (1989) Coastal sand dunes as geomorphological systems, Proceedings of the Royal Society of
Edinburgh, 96: 3-15.
Greipsson, S. (2002) Coastal dunes. In: Perrow, M. R. and Davy, A. J. (eds.) Handbook of Ecological
Restoration, Cambridge: Cambridge University Press, 214-237.
Hayden, B. P., Santos, M. C. F. V., Shao, G. and Kochel, R. C. (1995) Geomorphological controls on coastal vegetation at the Virginia Coastal Reserve, Geomorphology, 13: 283-300.
Hesp, P. A. (1989) A review of biological and geomorphological processes involved in the initiation and development of incipient foredunes, Proceedings of the Royal Society of Edinburgh, 96 (B): 181-201. Hesp, P. A. (2002) Foredunes and blowouts: initiation, geomorphology and dynamics, Geomorphology, 48 (1):
245-268.
Kim, K. D. (2005) Invasive plants on disturbed Korean sand dunes, Estuarine, Coastal and Shelf Science, 62: 353-364.
Musila, W. M., Kinyamario, J. I. and Jungerius, P. D. (2001) Vegetation dynamics of coastal sand dunes near Malindi, Kenya, African Journal of Ecology, 39: 170-177.
Nordstrom, K. F. and Lotstein, E. L. (1989) Perspectives on resource use of dynamic coastal dunes,
Geographical Review, 79 (1): 1-12.
Pethick, J. (1984) An Introduction to Coastal Geomorphology, London: Edward Arnold.
Pye, K. (1982) Morphological development of coastal dunes in a humid tropical environment, Cape Bedford and Cape Flattery, North Queensland, Geografiska Annaler, 64 (A): 213-227.
Pye, K. and Tsoar, H. (1990) Aeolian Sand and Sand Dunes, London: Unwin Hyman.
Stallins J. A. and Parker, A. J. (2003) The influence of complex systems interactions on barrier island dune vegetation pattern and process, Annals of the Association of American Geographers, 93: 13-29. Willis, A. J. (1989) Coastal sand dunes as biological systems, Proceedings of the Royal Society of Edinburgh,
96: 17-36.
2007 年 7 月 30 日 收稿 2007 年 12 月 28 日 修正 2008 年 1 月 8 日 接受
附錄 研究區物種名錄
I. Dicotyledons 雙子葉植物 1. Acanthaceae 爵床科Justicia procumbens Linn. 爵床 (H, V, C)
2. Aizoaceae 番杏科
Tetragonia tetragonoides (Pall.) Kuntze 番杏 (H, V, C) **
3. Araliaceae 五加科
Eleutherococcus trifoliatus (L.) var. trifoliatus 三葉五加 (S, V, C) ** 4. Asteraceae 菊科
Artemisia fukudo Makino 濱艾 (H, V, C)
Bidens pilosa L. var. radiata Sch. Bip. 大花咸豐草 (H, R, C)
Conyza canadensis (L.) Cronq. var. canadensis 加拿大蓬 (H, R,C)
Gynura divaricata (L.) DC. subsp. formosana (Kitam.) F. G. Davies 白鳳菜 (H, E, C)
Ixeris repens (L.) A. Gray 濱剪刀股 (H, V, C)
Pterocypsela indica (L.) C. Shih 鵝仔草 (H, V, C) Taraxacum formosanum Kitam. 臺灣蒲公英 (H,V, M)
Wedelia biflora (L.) DC. var. biflora 雙花蟛蜞菊 (S,V,C)
Wedelia prostrata (Hook. & Arn.) Hemsl. var. prostrata 天蓬草舅 (H, V, C)
5. Asclepiadaceae 蘿藦科
Gymnema sylvestre (Retz.) Schultes 武靴藤 (S, V, C) 6. Brassicaceae 十字花科
Lepidium virginicum L. 獨行菜 (H, R, C)
7. Caryophyllaceae 石竹科
Silene fortunei Vis. var. fortunei 蠅子草 (H, V, C)**
Silene fortunei Vis. var. kiruninsularis (Masamune) Ying 基隆蠅子草 (H, E, M)
8. Casuarinaceae 木麻黃科
Casuarina equisetfolia L. 木麻黃 (T, R, M) **
9. Chenopodiaceae 藜科
Chenopodium acuminatum Willd. subsp. virgatum (Thunb.) Kitamura 變葉藜 (H, V, C) **
10. Convolvulaceae 旋花科
Calystegia soldanella (L.) R. Br. 濱旋花 (H, V, C)
Cuscuta australis R. Br. 菟絲子 (H, V, C)
Ipomoea cairica (L.) Sweet 槭葉牽牛 (C, R, C)
Ipomoea imperati (Vahl) Griseb. 厚葉牽牛 (H, V, C)
Ipomoea pes-caprae (L.) R. Br. subsp. Brasiliensis (L.) Oostst. 馬鞍藤 (C, V, C)
11. Crassulaceae 景天科
Sedum formosanum N. E. Br. 臺灣佛甲草 (H, V, C) **
12. Euphorbiaceae 大戟科
Breynia officinalis Hemsley var. officinalis. 紅仔珠 (S, V, C)
13. Fabaceae 豆科
Leucaena leucocephala (Lam.) de Wit 銀合歡 (T, R, C) Millettia reticulata Benth. 老荊藤 (S, V, C)
Vigna marina (Burm.) Merr. 濱豇豆 (C, V, C)
14. Lamiaceae 唇形花科
Clinopodium chinense (Benth.) Kuntze 風輪菜 (H, V, C)
15. Lauraceae 樟科
Machilus thunbergii Sieb. & Zucc. 豬腳楠 (T, V, C) **
16. Malvaceae 錦葵科
Hibiscus tiliaceus L. 黃槿 (T, V, C) **
17. Menispermaceae 防己科
Cocculus orbiculatus (L.) DC. 土牛入石 (S, V, C)
18. Moraceae 桑科
Broussonetia papyrifera (L.) L'Herit. ex Vent. 構樹 (T, V, C) **
Morus australis Poir. 小桑樹 (T, V, C)
19. Onagraceae 柳葉菜科
Oenothera laciniata J. Hill 裂葉月見草 (H, R, C)
20. Orobanchaceae 列當科
Orobanche coerulescens Stephan 列當 (H, V, C) **
21. Oxalidaceae 酢醬草科
Oxalis corniculata L. 酢醬草 (H, V, C) **
22. Pandanaceae 露兜樹科
Pandanus odoratissimus L. f. 林投 (S, V, C)
23. Pittosporaceae 海桐科
Pittosporum tobira Ait. 海桐 (S, V, C)
24. Polygonaceae 蓼科
Polygonum chinense L. 火炭母草 (H, V, C)
Rumex crispus var. japonicus (Houtt.) Makino 羊蹄 (H, R, C) **
25. Portulacaceae 馬齒莧科
Portulaca oleracea L. 馬齒莧 (H, V, C) **
26. Primulaceae 櫻草科
Anagalis arvensis L. 琉璃繁縷 (H, V, C) **
Lysimachia mauritiana Lam. 茅毛珍珠菜 (H, V, C)
27. Ranunculaceae 毛茛科
Clematis grata Wall. 串鼻龍 (C, V, C)
28. Rhamnaceae 鼠李科
Berchemia lineata (L.) DC. 小葉黃鱔藤 (S, V, C)
29. Rubiaceae 茜草科
Paederia foetida L. 雞屎藤 (C, V, C)
30. Rutaceae 芸香科
31. Sapotaceae 山欖科
Planchonella obovata (R. Br.) Pierre 樹青 (T, V, C) **
32. Thymelaeaceae 瑞香科
Wikstroemia indica (L.) C. A. Mey. 南嶺蕘花 (S, V, C)
33. Ulmaceae 榆科
Celtis sinensis Pers. 朴樹 (T, V, C)
34. Umbelliferae 繖形科
Glehnia littoralis F. 濱防風 (H, V, C) **
Torilis japonica (Houtt.) DC. 竊衣 (H, V, C)**
35. Verbenaceae 馬鞭草科
Lantana camara L. 馬櫻丹 (S, R, C)
Phyla nodiflora (L.) Greene 鴨舌癀 (H, V, C)
Vitex rotundifolia L. F. 蔓荊 (S, V, C)
36. Vitaceae 葡萄科
Ampelopsis brevipedunculata (Maxim.) Traut var. hancei (Planch.) Rehder 漢氏山葡萄 (C, V, C) Cayratia japonica (Thunb.) Gagnep. 虎葛 (C, V, C)
37. Zygophyllaceae 蒺藜科 Tribulus terrestris L. 蒺藜 (H, V, C) II. Monocotyledons 單子葉植物 38. Amaryllidaceae 石蒜科 Crinum asiaticum L. 文珠蘭 (H, V, C) 39. Cyperaceae 莎草科
Carex pumila Thunb. 小海米 (H, V, C)
40. Liliaceae 百合科
Asparagus cochinchinensis (Lour.) Merr. 天門冬 (H, V, C)
Dianella ensifolia (L.) DC. in Red. 山菅蘭 (H, V, C)
Lilium formosanum Wallace 臺灣百合 (H, E, C) **
41. Poaceae 禾本科
Cynodon dactylon (L.) Pers. 狗牙根 (H, V, C)
Eleusine indica (L.) Gaertn. 牛筋草 (H, V, C)
Imperata cylindrica (L.) P. Beauv. var. major (Nees) C. E. Hubb. & Vaughan 白茅 (H, V, C)
Miscanthus floridulus (Labill) Warb. ex Schum. & Laut. 五節芒 (H, V, C)
Panicum repens L. 鋪地黍 (H, V, C)
Spinifex littoreus (Burm. f.) Merr. 濱刺草 (H, V, C)
Thuarea involuta (G. Forst) R. Br. Ex Sm. 濱箬草 (H, V, C)
42. Zingiberaceae 薑科
Alpinia zerumbet (Persoon) B. L. Burtt & R. M. Smith 月桃 (H, V, C)
共紀錄到42 科 73 種
備註:
1. 屬性代碼 (A, B, C) 對照表:欄 A──T:木本,S:灌木,C:藤本,H:草本。欄 B──E:特有,V:原生,R:
歸化。欄C──C:普遍,M:中等。