SOCS1 Methylation in Patients With Newly
Diagnosed Acute Myeloid Leukemia
Chien-Yuan Chen,1Woei Tsay,1Jih-Luh Tang,1Hwei-Ling Shen,1Shu-Wha Lin,2Sheng-Yi Huang,1Ming Yao,1 Yao-Chang Chen,1,2Ming-Ching Shen,1,2Chiu-Hwa Wang,1,2and Hwei-Fang Tien1*
1Department of Internal Medicine, National Taiwan University Hospital, Taipei, Taiwan 2Department of Laboratory Medicine, National Taiwan University Hospital, Taipei, Taiwan
The proliferation and differentiation of hematopoietic precursor cells depend on various cytokines. The suppressor of cytokine signaling-1 (SOCS1) down-regulates Janus kinases/signal transducers and activators of transcription (JAK/STAT) pathway activity and inhibits the biological effects of cytokines. SOCS1 has been shown to have tumor-suppressor activity, and methylation of this gene, resulting in transcriptional silencing, has been found in 65% of hepatocellular carcinoma and has been suggested to play an important role in the development of the cancer. The methylation status of the SOCS1 gene in acute myeloid leukemia (AML) has not been reported before. In this study, we analyzed SOCS1 methylation in 89 patients with newly diagnosed AML and correlated the result with immunophenotypes, cytogenetics, clinical features, and treatment outcome. SOCS1 methylation was found in the leukemic cells from 53 patients (60%). Thirteen (76%) of the 17 patients with t(15;17) had SOCS1 methylation, whereas this gene was methylated in only one (11%) of the nine patients with t(8;21). The frequencies of SOCS1 methylation among various cytogenetic subgroups differed significantly (P⫽ 0.014). Other clinical and laboratory parameters and the disease-free survival and overall survival were similar between patients with and without SOCS1 methylation. In conclusion, SOCS1 methylation occurs in more than half of AML cases, correlates with cytogenetic abnor-malities, and may play an important role in the development of subsets of AML. ©2003 Wiley-Liss, Inc.
INTRODUCTION
The proliferation and differentiation of hemato-poietic precursor cells are regulated by various cy-tokines (Lotem and Sachs, 2002). These cycy-tokines act in part through activation of the Janus kinase/ signal transducers and activators of transcription (JAK/STAT) pathway (Coffer et al., 2000; Ravandi et al., 2002). Inappropriate activation of the STAT signaling pathway may play an important role in the pathogenesis of leukemias (Coffer et al., 2000; Lin et al., 2000; Spiekermann et al., 2001, 2002). Constitutive activation of STAT transcription fac-tors in acute myeloid leukemia (AML) is associated with short disease-free survival (Benekli et al., 2002). The suppressor of the cytokine signaling (SOCS) family of proteins negatively regulates cy-tokine signaling (Krebs and Hilton, 2001).
The members of the SOCS family (SOCS1 to SOCS7 and CIS) are composed of a poorly con-served amino-terminal region, a central SH2 do-main, and a SOCS box (Hilton et al., 1998). SOCS1 is a negative regulator of the JAK/STAT pathway (Yoshikawa et al., 2001). It inhibits the biological effects of various cytokines, including IL-2, IL-3, IL-4, IL-6, interferon (INF)-␥, and INF-␣/ (Endo et al., 1997; Krebs and Hilton, 2001; O’shea et al., 2002). SOCS1-deficient mice die within the first 3 weeks of life from a myeloproliferative disorder,
which is driven by excessive IFN signaling (Naka et al., 1997; Starr et al., 1998). SOCS1 expression results in suppression of IL-6 and leukemia inhib-itory factor (LIF)– dependent STAT3 activation in M1 leukemia cells (Suzuki et al., 1998). Cytokines such as IL-4, IL-13, INF-␥, LIF, and GM-CSF as well as IL-6 induce SOCS1 gene expression in hematological cells (Naka et al., 1997; Starr et al., 1997). In vitro, the interactions between SOCS1 and various cytokines in hematopoietic cells are complex. In vivo, the role for SOCS1 in leukemia has not yet been investigated.
The expression of inducible SOCS1 is associated with tumor-suppressor activity (Rottapel et al., 2002). Aberrant methylation of the SOCS1 gene, which results in transcriptional silencing, was re-cently demonstrated in 17 of 26 human hepatocel-lular carcinomas (Yoshikawa et al., 2001). The
res-Supported by: The National Science Council of the Republic of China; Grant numbers: NSC 90-2314-B002-267 and 91-2314-B002-133.
*Correspondence to: Dr. Hwei-Fang Tien, Department of Inter-nal Medicine, NatioInter-nal Taiwan University Hospital, No. 7, Chung-Shan South Road, Taipei 100, Taiwan.
E-mail: [email protected]
Received 29 October 2002; Accepted 25 February 2003 DOI 10.1002/gcc.10222
toration of SOCS1 suppressed growth of tumor cells in which SOCS1 was methylation-silenced (Yo-shikawa et al., 2001). Aberrant DNA methylation in promoter regions of suppressor genes, including HIC1 (Issa et al., 1997b), WT1 (Plass et al., 1999), CDKN2B (Wong et al., 2000; Tien et al., 2001), and CDKN2A (Faderl et al., 2000), can be detected in AML and is usually associated with a poor progno-sis and increased relapse rates. The incidence and the clinical and biological implications of SOCS1 methylation in human AML are unknown. In this study, we analyzed the methylation status of the SOCS1 gene in leukemic cells and correlated the result with the clinical and laboratory characteris-tics of 89 patients with newly diagnosed AML.
MATERIALS AND METHODS Patients
The methylation status of the SOCS1 CpG island was studied in bone marrow cells from 89 patients (54 men, 35 women) with newly diagnosed AML at the National Taiwan University from 1995 to 2000. Pretreatment characteristics are shown in Table 1. Eighty-two patients were adults and seven were children, and the median age was 48 years (range 1– 85 years). The French–American–British (FAB) subtypes of AML included M1 (23 patients), M2 (30), M3 (17), M4 (14), M5 (4), and M7 (1).
Of the patients with AML other than the M3 subtype, most received conventional induction chemotherapy with cytarabine (AraC) for 7 days and one anthracycline (doxorubicin, idarubicin, or mitoxantrone) for 3 days. Some patients with old age and/or poor performance status received no treatment or only low dose AraC (10 mg/m2) for 14 to 21 days. The acute promyelocytic leukemia (APL) patients received all-trans retinoic acid with or without concurrent induction chemotherapy. Af-ter complete remission was achieved, the patients received consolidation chemotherapy with a con-ventional dose of AraC and one anthracycline or with high dose AraC (2 to 3 g/m2) twice a day for 3– 4 days.
Immunophenotype
A panel of monoclonal antibodies to myeloid-associated antigens including CD13, CD33, CD11b, CD15, CD14, and CD41a, as well as lym-phoid-associated antigens including CD2, CD5, CD7, CD19, CD10, and CD20, and lineage non-specific antigens HLA-DR, CD34, and CD56 was used to characterize the phenotypes of the leuke-mic cells. Expression of surface antigens on the leukemic cells was shown by an indirect immuno-alkaline phosphatase method as described before (Tien et al., 1993).
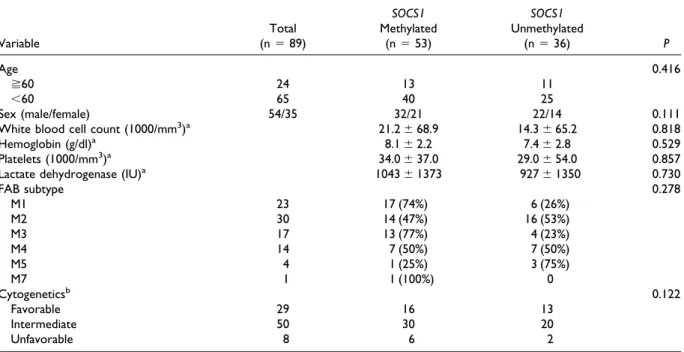
TABLE 1. Clinical Characteristics of the 89 AML Patients Before Treatment
Variable Total (n⫽ 89) SOCS1 Methylated (n⫽ 53) SOCS1 Unmethylated (n⫽ 36) P Age 0.416 ⭌60 24 13 11 ⬍60 65 40 25 Sex (male/female) 54/35 32/21 22/14 0.111
White blood cell count (1000/mm3)a 21.2⫾ 68.9 14.3⫾ 65.2 0.818
Hemoglobin (g/dl)a 8.1⫾ 2.2 7.4⫾ 2.8 0.529
Platelets (1000/mm3)a 34.0⫾ 37.0 29.0⫾ 54.0 0.857
Lactate dehydrogenase (IU)a 1043⫾ 1373 927⫾ 1350 0.730
FAB subtype 0.278 M1 23 17 (74%) 6 (26%) M2 30 14 (47%) 16 (53%) M3 17 13 (77%) 4 (23%) M4 14 7 (50%) 7 (50%) M5 4 1 (25%) 3 (75%) M7 1 1 (100%) 0 Cytogeneticsb 0.122 Favorable 29 16 13 Intermediate 50 30 20 Unfavorable 8 6 2 aMedian⫾ SD.
bChromosomal study in two patients showed no metaphase cells for analysis. Favorable: including t(8;21), t(15;17), and inv(16). Unfavorable: t(7;11)
Cytogenetics
Chromosome analyses were carried out as de-scribed previously (Tien et al., 1995). Bone marrow (BM) cells were harvested directly or after 1–3 days of non-stimulated culture. Metaphase chromo-somes were banded by trypsin-Giemsa and karyo-typed according to the ISCN (Mitelman, 1995). Methylation-Specific Polymerase Chain Reaction (PCR)
The methylation status of the promoter region of the SOCS1 gene was analyzed by methylation-spe-cific PCR as described (Herman et al., 1996; Tien et al., 2001). Mononuclear cells were isolated from BM aspirates by Ficoll-Hypaque gradient centri-fugation. High-molecular-weight DNA was ex-tracted. DNA (4 g) in a volume of 40 l was denatured by the addition of 10 l of 1 mol/L NaOH (final concentration 0.2 mol/L) for 10 min at 37°C. Hydroquinone (30l of 10 mmol/L) (Sigma, St. Louis, MO) and 520 l of 1.5 mol/L sodium bisulfite (Sigma) at pH 5 were added and mixed, and samples were incubated under mineral oil at 50°C for 16 hr. Modified DNA was purified by use of the Wizard DNA purification resin and Vacuum Manifold, according to the manufacturer’s instruc-tion (Promega, Madison, WI), and then eluted into 100l of water. Final desulfonation was achieved by treatment with 50 l of 1 mol/L NaOH (final concentration 0.3 mol/L) at room temperature for 5 min, followed by ethanol precipitation. DNA was resuspended in 45 l of water and used immedi-ately or stored at⫺20°C before use.
The bisulfite-modified DNA was amplified by PCR with either a methylation-specific or un-methylation-specific primer set, designed by Yo-shikawa et al. (2001). The methylation-specific primer sequences were 5⬘-TTC GCG TGT ATT TTT AGG TCG GTC-3⬘ (sense) and 5⬘-CGA CAC AAC TCC TAC AAC GAC CG-3⬘ (anti-sense). The unmethylation-specific primer se-quences were 5⬘-TTA TGA GTA TTT GTG TGT ATT TTT AGG TTG GTT-3⬘ (sense) and 5⬘-CAC TAA CAA CAC AAC TCC TAC AAC AAC CA-3⬘ (antisense). Negative controls (normal DNA and distilled water) were used in each exper-iment. The hepatoma cell lines Hep3B and SNU423 were used as positive controls; the former had an amplified band in PCR with methylation-specific primers, but not in PCR with unmethyla-tion-specific primers, and the latter had positive bands in both conditions. To avoid contamination,
each DNA sample was aliquoted and analyzed by use of at least two different PCRs.
Statistics
Comparisons were made with the t-test. Survival curves were plotted by use of the Kaplan–Meier method; differences between curves were analyzed by the log-rank test. All statistical analyses were performed by use of SPSS 8.0 for Windows (SPSS, Chicago, IL). Values of P⬍ 0.05 were considered significant.
RESULTS
SOCS1 Methylation in AML and Its Correlation With Clinical Features
Methylation of the promoter region of the SOCS1 gene was detected in 53 (60%) of the 89 patients with newly diagnosed AML (Fig. 1). Blood cells from eight normal donors of hematopoietic stem cells showed no methylation. The FAB M1 and M3 subtypes had higher incidences of SOCS1 methylation (74 and 77%, respectively) than the M2, M4, and M5 subtypes (47, 50, and 25%, re-spectively; Table 1). However, the difference did not reach statistical significance (M1 and M3 vs. M2, M4, and M5: 75 vs. 46%, P ⫽ 0.626). Other clinical and laboratory features, including age, sex, white blood cell count, hemoglobin, platelets, and lactate dehydrogenase, were similar between pa-tients with and without SOCS1 methylation (Table 1). In addition to the amplified bands shown by PCR that used methylation-specific primers, sam-ples from 49 of the 53 patients with SOCS1 meth-ylation could also be amplified with unmethyla-tion-specific primers (Fig. 1). This may be explained by contamination of normal cells or the
Figure 1. Methylation-specific polymerase chain reaction analysis of the SOCS1 promoter region in 12 patients with newly diagnosed AML (left panel: lanes 1 to 5 in experiment I; right panel: lanes 1 to 7 in experiment II). Five patients show SOCS1 methylation (upper row, lanes 2 and 4 in experiment I; lanes 3, 5, and 6 in experiment II). Amplified bands of unmethylated DNA can also be seen in three of them (lower row: lanes 2 and 4 in experiment I; lane 6 in experiment II), but not in the other two. In experiment I, lane 6: normal control; lanes 7 and 8: hepatoma cell lines SNU-423 and Hep3B, respectively; lane 9: water.
presence of unmethylated alleles in the AML cells (Cameron et al., 1999; Tien et al., 2001).
Correlation of SOCS1 Methylation With Cytogenetics and Immunophenotypes
The cytogenetic studies were performed before treatment. Two patients showed no metaphase cells for analysis. The cytogenetic result for the remaining 87 patients is shown in Table 2. The incidence of SOCS1 methylation was low (11%) in AML with t(8;21), but high (76%) in APL with t(15;17); the difference among various cytogenetic subgroups was statistically significant (P ⫽ 0.014; Table 2).
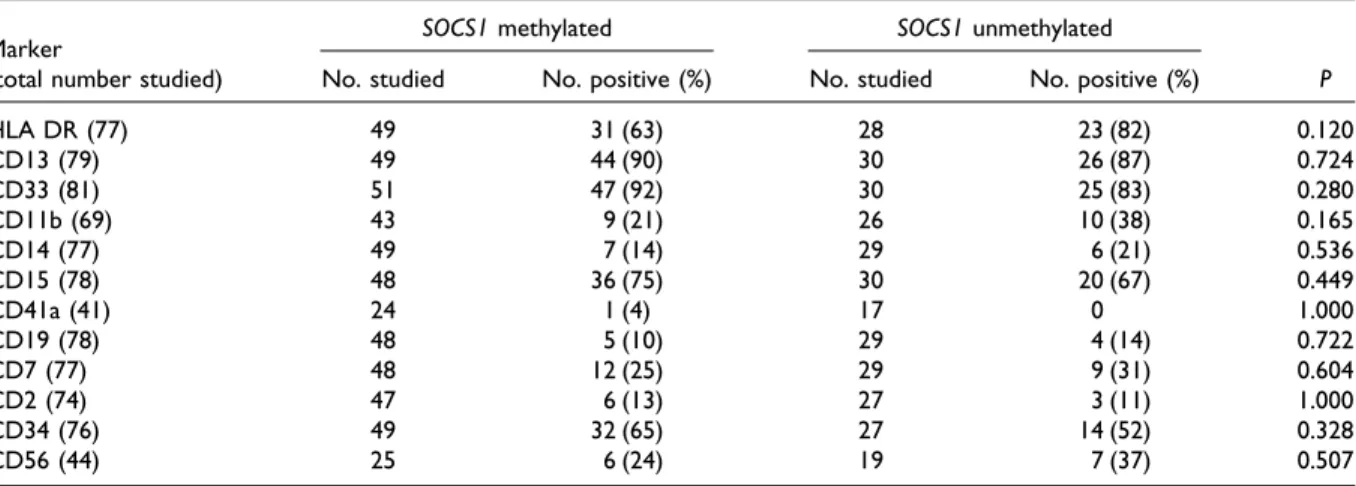
The patients with SOCS1 methylation had a somewhat lower incidence of HLA DR and CD11b expression on the leukemia cells than did those without methylation (63 vs. 82%, P⫽ 0.12 and 21 vs. 38%, P⫽ 0.165, respectively; Table 3). There was no difference in the expression of other anti-gens between the two groups of patients.
Correlation of SOCS1 Methylation With Treatment Outcome
Among the 68 patients who received standard induction chemotherapy, 34 (83%) of the 41 pa-tients with SOCS1 methylation and 24 (89%) of the 27 without methylation obtained a complete remis-sion (Table 4). The median disease-free survival was 15 months in the former group and 10 months in the latter group (P ⫽ 0.97). Also, the overall survival was not different between the two groups (median, 30 vs. 58 months, P⫽ 0.524).
DISCUSSION
This is the first report concerning SOCS1 meth-ylation in AML. Sixty percent of newly diagnosed AML cases were identified to have aberrant meth-ylation in the SOCS1 CpG island. SOCS1 is a neg-ative regulator of the JAK/STAT signaling pathway, and inappropriate activation of JAK and STAT proteins has been associated with the oncogenic process (Bowman et al., 2000; Coffer et al., 2000). Direct implication of the JAK/STAT signaling path-way in human hematological malignancies has been demonstrated by the identification of trans-locations involving JAK and STAT encoding genes. For example, the STAT5B gene is fused to the retinoic acid receptor alpha (RARA) gene in an acute promyelocytic-like leukemia (Arnould et al., 1999), and a translocation involving the JAK2 and ETV6 genes, which results in constitutive activa-tion of the JAK2 protein tyrosine kinase, has been described in leukemia (Lacronique et al., 1997). SOCS1 interferes with the ETV6/JAK2-induced phosphorylation and activates proteasome-depen-dent degradation (Frantsve et al., 2001). Moreover, the constitutive expression of SOCS1 blocks the proliferation of cells transformed with ETV6/JAK2, BCR/ABL, or v-ABL (Rottapel et al., 2002). Dys-regulation of the SOCS1 gene may hence play a role in leukemogenesis.
The CpG islands of genes are essentially un-methylated in normal tissues (Bird, 1986), but be-come hypermethylated in some tumor-suppressor genes in malignancies, including leukemias (Issa et al., 1997a; John et al., 1999). Aberrant methylation of CpG islands is associated with gene inactivation (Singal and Ginder, 1999) and may contribute to the pathogenesis of neoplasia. SOCS1 has been shown to have tumor-suppressor activity (Rottapel et al., 2002). Yoshikawa et al. (2001) demonstrated that aberrant methylation in the CpG island of the SOCS1 gene resulted in its transcriptional silencing in hepatocellular carcinoma and that restoration of SOCS1-suppressed growth of tumor cells in which SOCS1 was methylation-silenced and JAK2 was constitutively activated. Because SOCS1 methyl-ation was demonstrated in most newly diagnosed AML, we suggest that SOCS1 may collaborate with other genetic abnormalities to facilitate the devel-opment of leukemia.
It is noteworthy that the incidence of SOCS1 methylation is different among the various cytoge-netic subgroups, being higher in APL with t(15;17), and lower in AML with t(8;21). Because the sig-naling cascades in AML with different cytogenetic
TABLE 2. Correlation of SOCS1 Methylation With Cytogenetic Results
Chromosome changes
Number of patients
P Total With SOCS1 methylation (%)
t(8;21) 9 1 (11) t(15;17) 17 13 (76) Inv(16) 3 2 (67) t(7;11) 3 2 (67) t(9;11) 2 1 (50) Trisomy 8 sole 5 4 (80) Simplea 21 12 (57) Complexa 5 4 (80) Normal 22 13 (59) Totalb 87 52 (60) 0.014
aExcluding nonrandom abnormalities. Simple indicates three or fewer
abnormalities and complex, four or more.
bChromosomal study in two patients showed no metaphase cells for
abnormalities are not fully understood, it is not clear why SOCS1 methylation occurs frequently in some AMLs but not in others.
The SOCS1 methylation had no impact on dis-ease-free survival or overall survival in this study. Methylation of the CpG islands in HIC1, WT1, CDKN2B, and CDKN2A is associated with a poor outcome and a high relapse rate in patients with AML (Issa et al., 1997b; Plass et al., 1999; Faderl et al., 2000; Wong et al., 2000; Tien et al., 2001). In contrast, methylation of the estrogen receptor gene has been associated with improved prognosis in AML (Li et al., 1995). Thus, the correlation of CpG-island methylation and prognosis is still con-troversial, and the clinical implications of DNA methylation possibly depend on the function of the involved genes.
REFERENCES
Arnould C, Philippe C, Bourdon V, Gre´goire MJ, Berger R, Jon-veaux P. 1999. The signal transducer and activator of transcription STAT5b gene is a new partner of retinoid receptor alpha in acute promyelocytic-like leukemia. Hum Mol Genet 8:1741–1749. Benekli M, Xia Z, Donohue KA, Ford LA, Pixley LA, Baer MR,
Baumann H, Wetzler M. 2002. Constitutive activity of signal transducer and activator of transcription 3 protein in acute
my-eloid leukemia blasts is associated with short disease free survival. Blood 99:252–257.
Bird AP. 1986. CpG-rich islands and the function of DNA methyl-ation. Nature 321:209 –213.
Bowman T, Garcia R, Turkson J, Jove R. 2000. STATs in oncogen-esis. Oncogene 19:2474 –2488.
Cameron EE, Baylin SB, Herman JG. 1999. p15INK4BCpG-island
methylation in primary acute leukemia is heterogeneous and suggests density as a critical factor for transcriptional silencing. Blood 94:2445–2451.
Coffer PJ, Koenderman L, de Groot RP. 2000. The role of STATs in myeloid differentiation and leukemia. Oncogene 19:2511–2522. Endo TA, Masuhara M, Yokouchi M, Suzuki R, Sakamoto H, Mitsui K, Matsumoto A, Tanimura S, Ohtsubo M, Misawa H, Miyazaki T, Leonor N, Taniguchi T, Fujita T, Kanakura Y, Komiya S, Yoshimura A. 1997. A new protein containing an SH2 domain that inhibits JAK kinase. Nature 387:921–924.
Faderl S, Kantarjian HM, Estey E, Manshouri T, Chan CY, Rahman Elsaied A, Kornblau SM, Cortes J, Thomas DA, Pierce S, Keating MJ, Estrov Z, Albitar M. 2000. The prognostic significance of p16(INK4a)/p14(ARF) locus deletion and MDM-2 protein ex-pression in adult acute myelogenous leukemia. Cancer 89:1976 – 1982.
Frantsve J, Schwaller J, Sternberg DW, Kutock J, Gilliland DG. 2001. SOCS-1 inhibits TEL-JAK2 mediated transformation of hematopoietic cells through inhibition of JAK2 kinase activity and induction of proteasome–mediated degradation. Mol Cell Biol 21:3547–3557.
Herman JG, Graff JR, Myohanen S, Nelkin BD, Baylin SB. 1996. Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci USA 93:9821–9826. Hilton DJ, Richardson RT, Alexander WS, Viney EM, Wilson TA,
Spring NS, Starr R, Nicholson SE, Metcalf D, Nicola NA. 1998. Twenty proteins containing a C-terminal SOCS box from five structural classes. Proc Natl Acad Sci USA 95:114 –119.
TABLE 3. Surface Antigen Expression in Patients With and Without SOCS1 Methylation Marker
(total number studied)
SOCS1 methylated SOCS1 unmethylated
P No. studied No. positive (%) No. studied No. positive (%)
HLA DR (77) 49 31 (63) 28 23 (82) 0.120 CD13 (79) 49 44 (90) 30 26 (87) 0.724 CD33 (81) 51 47 (92) 30 25 (83) 0.280 CD11b (69) 43 9 (21) 26 10 (38) 0.165 CD14 (77) 49 7 (14) 29 6 (21) 0.536 CD15 (78) 48 36 (75) 30 20 (67) 0.449 CD41a (41) 24 1 (4) 17 0 1.000 CD19 (78) 48 5 (10) 29 4 (14) 0.722 CD7 (77) 48 12 (25) 29 9 (31) 0.604 CD2 (74) 47 6 (13) 27 3 (11) 1.000 CD34 (76) 49 32 (65) 27 14 (52) 0.328 CD56 (44) 25 6 (24) 19 7 (37) 0.507
TABLE 4. SOCS1 Methylation and Treatment Outcome
Outcome SOCS1 methylated (n⫽ 41) SOCS1 unmethylated (n⫽ 27) P Complete remission (%) 34/41 (83%) 24/27 (89%) 0.242 Consolidation chemotherapy 0.579 Conventional regimen 29/41 13/27
High dose regimen 12/41 14/27
Stem cell transplantation 9 5 0.117
Disease-free survival
(median, months) 15 10 0.245
Overall survival (median,
Huang SY, Tang JL, Liang YJ, Wang CH, Chen YC, Tien HF. 1997. Clinical, hematological and molecular studies in patients with chromosome translocation t(7;11): a study of four Chinese patients in Taiwan. Br J Haematol 96:682– 687.
Issa JP, Baylin SB, Herman JG. 1997a. DNA methylation changes in hematologic malignancies: biological and clinical implications. Leukemia 11:s7–s11.
Issa JP, Zehnbauer BA, Kaufmann SH, Biel MA, Baylin SB. 1997b. HIC1 methylation is a late event in hematopoietic neoplasms. Cancer Res 57:1678 –1681.
John RM, Paul CV, Susan JC. 1999. Concurrent DNA methylation of multiple genes in acute myeloid leukemia. Cancer Res 59: 3730 –3740.
Krebs DL, Hilton DJ. 2001. SOCS proteins: negative regulator of cytokine signaling. Stem Cells 19:378 –387.
Lacronique V, Boureux A, Valle VD, Poirel H, Quang CT, Mauchauffe M, Berthou C, Lessard M, Berger R, Ghysdael J, Bernard OA. 1997. A TEL-Jak2 fusion protein with constitutive kinase activity in human leukemia. Science 278:1309 –1312. Li Q, Kopecky KJ, Mohan A, Willman CL, Appelbaum FR, Weick
JK, Issa JP. 1995. Estrogen receptor methylation is associated with improved survival in adult myeloid leukemia. Clin Cancer Res 5:1077–1084.
Lin TS, Mahajan S, Frank DA. 2000. STAT signaling in the patho-genesis and treatment of leukemias. Oncogene 19:2496 –2504. Lotem J, Sachs L. 2002. Cytokine control of developmental
pro-grams in normal hematopoiesis and leukemia. Oncogene 21:3284 –3294.
Mitelman F, editor. 1995. ISCN: an international system for human cytogenetic nomenclature. Basel: S. Karger.
Naka T, Narazaki M, Hirata M, Matsumoto T, Minamoto S, Aono A, Nishimoto N, Kajita T, Taga T, Yoshizaki K, Akira S, Kishimoto T. 1997. Structure and function of a new STAT induced STAT inhibitor. Nature 387:924 –929.
O’shea JJ, Gadina M, Schreiber RD. 2002. Cytokine signaling in 2002: new surprises in the Jat/Stat pathway. Cell 109:S121–S131. Plass C, Yu F, Yu L, Strout MP, El-Rifai W, Elonen E, Knuutila S, Marcucci G, Young DC, Held WA, Bloomfield CD, Caligiuri MA. 1999. Restriction landmark genome scanning for aberrant mehy-lation in primary refractory and relapsed acute myeloid leukemia: involvement of the WIT-1 gene. Oncogene 18:3159 –3165. Ravandi F, Talpaz M, Kantarjian H, Estrov Z. 2002. Cellular
sig-naling pathways: new targets in leukemia therapy. Br J Haematol 116:57–77.
Rottapel R, Ilangumaran S, Neale C, La Rose J, Ho JM, Nguyen MH, Barber D, Dubrenil P, deSepulveda P. 2002. The tumor suppressor activity of SOCS-1. Oncogene 21:4351– 4362.
Singal R, Ginder GD. 1999. DNA methylation. Blood 93:4059 – 4070.
Spiekermann K, Biethahn S, Wilde S, Hiddemann W, Alves F. 2001. Constitutive activation of STAT transcription factors in acute myelogenous leukemia. Eur J Hematol 67:63–71.
Spiekermann K, Pau M, Schwab R, Schmieja K, Franzrahe S, Hid-demann W. 2002. Constitutive activation of STAT3 and STAT5 is induced by leukemic fusion proteins with protein tyrosine kinase activity and is sufficient for transformation of hematopoi-etic precursor cells. Exp Hematol 30:262–271.
Starr R, Willson TA, Viney EM, Murray LJ, Rayner JR, Jenkins BJ, Gonda TJ, Alexander WS, Metcalf D, Nicola NA, Hilton DJ. 1997. A family of cytokine-inducible inhibitors of signaling. Na-ture 387:917–921.
Starr R, Metcalf D, Elefanty AG, Brysha M, Willson TA, Nicola NA, Hilton DJ, Alexander WS. 1998. Liver degeneration and lym-phoid deficiencies in mice lacking suppressor cytokine signal-ing-1. Proc Natl Acad Sci USA 95:14395–14399.
Suzuki R, Sakamoto H, Yasukawa H, Masuhara M, Wakioka T, Sasaki A, Yuge K, Komiya S, Inoue A, Yoshimura A. 1998. CIS3 and JAB have different regulatory roles in interleukin-6 mediated differentiation and STAT 3 activation in M1 leukemia cells. Oncogene 17:2271–2278.
Tien HF, Wang CH, Chen YC, Shen MC, Lin DT, Lin KH. 1993. Characterization of acute myeloid leukemia (AML) coexpressing lymphoid markers: different biologic features between T-cell antigen positive and B-cell antigen positive AML. Leukemia 7:688 – 695.
Tien HF, Wang CH, Lin MT, Lee FY, Liu MC, Chuang SM, Chen YC, Shen MC, Lin KH, Lin DT. 1995. Correlation of cytogenetic results with immunophenotype, genotype, clinical features and ras mutation in acute myeloid leukemia. A study of 235 Chinese patients in Taiwan. Cancer Genet Cytogenet 84:60 – 68. Tien HF, Tang JL, Tsay W, Liu MC, Lee FY, Wang CH, Chen YC,
Shen MC. 2001. Methylation of the p15 (INK4B) gene in myelo-dysplastic syndrome: it can be detected early at diagnosis or during disease progression and is highly associated with leukemic transformation. Br J Haematol 112:148 –154.
Wong IH, Ng MH, Huang DP, Lee JC. 2000. Aberrant p15 promo-tor methylation in adult and childhood acute leukemia of nearly all morphologic subtypes: potential prognostic implications. Blood 95:1942–1949.
Yoshikawa H, Matsubara K, Qian GS, Jackson P, Groopman JD, Manning JE, Harris CC, Herman JG. 2001. SOCS-1, a negative regulator of the JAK/STAT pathway, is silenced by methylation in human hepatocellular carcinoma and shows growth-suppression activity. Nat Genet 28:29 –35.