行政院國家科學委員會補助專題研究計劃
期中進度報告
利用基因體方法研究人類病原白色念珠菌致病化因素
之功能
計畫類別:個別型計畫
計畫編號:NSC 96-3112-B-009-004
執行期間:
96 年
05 月
01 日至
97 年
04 月 30 日
計畫主持人:楊昀良
共同主持人:藍忠昱
計畫參與人員:許淑貞、李淑萍、陳亦達
成果報告類型(依經費核定清單規定繳交):完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立交通大學生物科技學系
中
華
民
國
97
年
03
月
18
日
一、中文摘要:
關鍵詞:致病機制調控、訊號傳遞途徑、基因同源置換、去氧核糖核酸微陣列、 功能性基因組 近年來,人類真菌感染 (fungal infections) 的情形有日益增加的趨勢。這 其中以白色念珠菌(Candida albicans) 為最大宗-在美國相關的醫療費用估計 為每年 10 億美金。雖然臨床上有抗真菌藥物,然而這些藥物普遍有副作用、對 新出現及一些特定的菌株無效、及造成抗藥性流行等問題。因此,發展及引進新 的觀念及策略,由不同的角度及方法來瞭解真菌感染的問題是有必要的。 目前已知與調控白色念珠菌致病機制有關的因素有環境刺激、營養之取得、抗 藥性、及毒性因子(virulence factors)等。但是對整個致病機制的調控及各別 因素間彼此的關連仍有很多不清楚的地方。因此,我們以功能性基因組的角度來 分析及探索致病化 (pathogenesis) 過程中,這些因素的訊號傳遞途徑及其與致 病化整體調控迴路(regulatory network)的關係。 目前已知的白色念珠菌主要的致病化調控因之中,Efg1 同時調控幾個致病化 因素,是一個主要的調控樞紐。因此,在這個計劃的第一年中,進行的第一個目 標是由受其調節的 CaEno1 切入,對 Efg1 途徑(pathway)展開功能性研究。CaEno1 本身是多功能性的蛋白,最早是以醣解酵素的功能被發現。它不但是白色念珠菌 細胞壁的主要成分,也是白色念珠菌感染寄主時的主要抗原,同時也影響白色念 珠菌的致病化過程,因此它在 Efg1 途徑中所扮演的角色就很耐人尋味了。由於 目前白色念珠菌有性世代(sexual cycle)尚未被發現,也缺乏適當的質體 (plasmid),因此基因突變的方法將以同源置換的方式直接對白色念珠菌基因體 進行 gene targeting 而達成。目前已成功構築重組 DNA,將 Marker 序列兩側加上CaENO1 的序列,並成功利用此重組 DNA 進行同源置換,以 gene targeting
的方式將CaENO1 的兩套序列皆以 Marker 序列取代,並在特定碳源環境中培養,
迴避醣解酵素的功能,正在進行致病力相關的功能性分析。同時也在啤酒酵母菌 中建立 EGFP reporter,將 CaENO1 與 EGFP 作 fusion,成功的證實此重組基因除 可留置於細胞質中,尚可被分泌至細胞外,與在白色念珠菌中一致。目前已針對 CaENO1-EGFP 的 ENO1 細進行 deletion analysis,以找出相對應於不同 cellular location 的序列,進而瞭解其調控。
第二個進行的目標是以去氧核糖核酸微陣列為工具,刻劃由前述之特定致病機 制及環境因素對白色念珠菌造成的整體基因組的表現。目前已初步在 UltraGAPS 玻片上完成一次 microarray spotting 的流程,正進行 microarray 的品質驗證 及實驗操作的最佳化。主要測試因子有 RNA 分離、樣品標記、及雜合反應的條件。
二、英文摘要
Keywords:Pathogenesis, Regulatory network, DNA microarray, Functional
genomics.
Currently, the known factors associated with C. albicans pathogenesis include environmental cues, nutrition availability, drug resistance, and virulence. Although extensive researches have been devoted to individual factors, the overall picture of the global networking and the coordination among those pathways contributing to the pathogenesis is still missing. Therefore, we employ the tools of functional genomics, particularly homologous replacement and DNA microarray, to analyze and explore the following: (1) the function of genes whose products involved in the
signaling/regulatory pathways contributing to pathogenesis and (2) their roles in the network of pathogenesis.
In first year, our objective is to focus on the functional study of CaEno1, which is under the pathways regulated by Efg1, the key regulatory factor of several
pathogenesis pathways. CaEno1 is a known glycolytic proteins specifically regulated by Efg1 and involved in the pathogenesis. We are interested in knowing what is its function and role in the Efg1 pathway and Candida pathogenesis?
Therefore, our first goal is to perform genetic study on CaENO1 by constructing null mutants. Due to the lack of known sexual cycle and plasmids, mutagenesis to study the function of this gene will rely on the homologous replacement technique.
Marker genes flanked with short homologous sequences at both sides will be used to knockout both alleles of CaENO1 via homology recombination. We have completed the construction of recombinant DNA containing selection markers franked by
homologous sequence of CaENO1. These recombinant DNAs were used to
knock-out r both alleles of CaENO1. According to the PCR and Southern analyses, we have successfully obtained the null mutant of CaENO1. In addition, CaEno1p appeared in several cellular locations yet there is no known localization signal. Hence, we have established a localization reporter system in baker yeast to identify the sequences on CaENO1 responsible to various locations, which are tightly connected to the pathogenesis and virulence of C. albicans. We are now in the process of deletion analysis to define those sequences.
Our second objective is the fabrication of DNA microarray for studying the global networking of pathogenesis and environmental responses. We have spotted on the UltraGAPS coated slides and started testing the quality of the products and have complete optimizing the conditions, such as RNA isolation, sample labeling, and hybridization, for the experiments.
三、目錄:
一、中文摘要………1
二、英文摘要………2
三、目錄………3
四、報告內容………4
A. Background and Significance………4
B. Specific Aims………8
C. Research Design and Methods………10
D. Results and Discussions………16
五、參考文獻………26
四、報告內容
A. Background and Significance
Yeast infections in human have increased significantly in recent years. Among the pathogens, Candida albicans is the most dominant one. Currently, the available antifungal drugs have undesirable issues such as side effects, ineffective against new or reemerging fungi, and leading to the rapid development of resistance (White et al., 1998; Yang and Lo, 2001). Candida albicans is an opportunistic fungal pathogen,
commensally colonizes various anatomical sites in humans. In the
immunocompromise individuals, such as the ones with HIV infection, diabetes, organ transplantation and cancer chemotherapy, C. albicans can become virulent and
invasive (Edwards, 1990). Candida albicans not only causes mucosal diseases (such
as oropharyngeal/esophageal and vulvovaginal candidiasis), but can also invade the bloodstream (candidaemia). It has emerged as the fourth most common cause of nosocomial infection (Beck-Sague and Jarvis, 1993). The estimated cost for treating this Candida nosocomial infection approaches 1 billion US Dollars per year in the United States (Miller et al., 2001). Understanding the regulatory mechanisms and gene functions of pathogenesis-related pathways may provide us the knowledge and drug targets for anti-fungal application. Although several factors associated with pathogenesis, such as environmental cues, nutrition availability, drug resistance, and virulence factors (such as morphogenesis, extracellular hydrolytic activities, and phenotype switch) have been identified in C. albicans, the picture of the global
networking and coordination of those regulations and signaling pathways contributing to pathogenesis in Candida albicans is still lacking. Noteworthy, those pathways are intertwined and only handful controlling points are known to modulate the global activities. There are the well-known major controlling point EFG1 and the two lesser candidates, CPH1, and TUP1. In this project, we will focus on the
relationship between environmental cues, stress, and morphogenesis, with emphasis on the connection to one of the major pathogenesis control point, Efg1 pathway.
Complexity of pathogenesis in C. albicans and Efg1 as a major controlling point
Signaling pathways commonly sense and transfer environmental signals to downstream regulators that lead to the expression of subsets of genes potentially related to Candida pathogenesis. Those subsets of genes are known as virulence factors. Several virulence factors of C. albicans have been proposed, including hyphal morphogenesis, extracellular hydrolytic activities (e.g. secreted aspartyl proteinases and lipases) (Calderone and Fonzi, 2001; Gow et al., 2002; Naglik et al., 2004; Stehr et al., 2004; Sundstrom, 2002; Yang, 2003), and phenotypic switching.
In hyphal morphogenesis, cells transit from an ovoid yeast shape to filament forms (pseudohyphae and hyphae); both filamentous forms are able to again produce yeast forms (Lo et al., 1997). Hyphal morphogenesis can be induced by a number of environmental cues, including the presence of serum, N-acetylglucosamine (GlcNAc), proline, neutral pH and elevated temperature (Ernst, 2000). The secreted aspartyl proteinases are encoded by a large SAP gene family of ten members, each
differentially regulated under a variety of conditions (e.g. pH, temperature and cell morphology) (Naglik et al., 2004). Finally, C. albicans reversibly switches phenotypes with a high frequency (Soll, 2002) known as the phenotypic switching. It occurs spontaneously and is also induced by temperature and low doses of UV irradiation (Soll, 1997). These cells that undergo the switch of phenotypes vary in morphology, physiology, metabolism and pathogenicity (Lan et al., 2002; Soll, 1997).
But then how do these "factors" connect to the pathogenesis? In C. albicans yeast-hyphae morphogenesis, the presence of serum activates adenylate cyclase (Cdc35), and thus promotes cyclic adenosine monophosphate (cAMP) production. Cyclic AMP acts as an intracellular regulator and in turn activates protein kinase A (PKA) that mediates its downstream signal via Efg1. Expression of the C. albicans
HMX1 gene, which encodes a heme oxygenase required for utilization of exogenous
heme and hemoglobin, is shown to be strongly de-regulated in an EFG1-null mutant (Santos et al., 2003). In C. albicans, the pH response is governed in part by the transcription factor Rim101. Rim101 promotes alkaline responses by repressing expression of Nrg1, itself a transcriptional repressor (Bensen et al., 2004). Rim101 and Nrg1 are also shown to act in parallel pathways to control hyphae morphogenesis. Interestingly, an alkaline pH condition induces expression of subsets of genes,
including several iron acquisition genes also mediated by Rim101 (Bensen et al., 2004). Moreover, in the yeast model system, Saccharomyces cerevisiae, cAMP controls the activity of ferrireductases, components of a high-affinity iron uptake system. Another example is the Tpk2, a catalytic subunit of protein kinase A, whose expression negatively regulates iron uptake genes (Lesuisse et al., 1991; Robertson et al., 2000). And the complication does not stop here. Recently, we have reported that in addition to being a virulence factor, Efg1 is also involved in drug resistance (Lo et al., 2005). Together, these studies implicate that the decision-making presides over the onset of pathogenesis is made of a complex signaling and regulatory network, which includes multiple components and each of them may controls subsets of gene expression. However, those studies also point out the fact that situating among this complex network, there is the key regulator Efg1, which through coordination of different components/pathways, regulates various cell functions (ie. drug resistance, morphogenesis, and gene expression) in response to different environmental cues (e.g.
iron availability, serum, and temperature). Hence, we are interested in what is the networking centering at Efg1.
Recently, we have reported that Efg1 and its downstream target CaEno1 have multiple functions in C. calbicans (Lo et al., 2005; Yang et al., 2006). CaENO1
encodes enolase and is under the control of Efg1 (Nantel et al., 2002). Enolase is a highly conserved protein throughout the phylogenetic tree (Van der et al., 1991). In addition to the known function in the glycolytic pathway, enolase is a major
glucan-associated protein found in the fungal cell wall. It also serves as a receptor for human plasmin/plasminogen (Jong et al., 2003) and the major immunogen of Candida infection. In different cellular locations, its function varies. Interestingly, the locality of enolase in cancer cells is associated with the ability for metastasis. Therefore, it is our interest to known the function of CaEno1 in connection to pathogenesis and the locality-function relationship.
The adaptation of C. albicans to the environments and its relation to the pathogenesis factors.
The ability of C. albicans to sense and adapt to alterations in host environments is integrated in its survival and pathogenicity (Soll, 2002). As it has been mentioned, several environmental conditions are known to affect cellular growth and
morphogenesis. In fact, they can even have direct impact on pathogenictiy. Those factors include various forms of stress and the availability of nutrients, for example, iron. The iron-free forms of host lactoferrin and transferrin inhibit C. albicans growth and render it more susceptible to damage by neutrophils (Okutomi et al., 1998). Iron deprivation affects the adhesive properties and cell wall compositions of
C. albicans (Paul et al., 1989; Sweet and Douglas, 1991) and studies on suspension
cultures and biofilms indicate that drug resistance of C. albicans is affected by iron availability (Baillie and Douglas, 1998; Paul et al., 1991). The high-affinity iron permease (CaFtr1) is required for systemic infection in a mouse model whereas the siderophore transporter (CaArn1) is required for epithelial invasion. Besides the endothelial cell injury caused by C. albicans is iron-dependent (Fratti et al., 1998; Heymann et al., 2002; Ramanan and Wang, 2000). Moreover, in the human host, iron is mostly bound to high-affinity ligands (e.g. transferrin, hemoglobin, lactoferrin and ferritin), and there is virtually no free iron available. This iron-withholding is an important defense mechanism for the host; the availability of iron has shown to be a common signal to induce the expression of virulence factors of pathogens (Paul et al., 1991). But how do the signals from those environmental cues link to the
pathogenesis pathway? Do they achieve this by sending signal to one of the virulence factors? To study the molecular mechanism of stimulus-responsive gene
regulation in C. albicans using iron availability as the model, we have identified iron-regulated genes and a potential DNA-binding protein, Sfu1, which negatively regulates gene expression under iron-repletion conditions (Lan et al., 2004).
Recently, the cellular levels of iron are shown to be crucial for the mode of action of a topical antifungal agent ciclopirox olamine, while the detail mechanism is most unknown (Sigle et al., 2005). Therefore, we are interested in how does the iron availability affect the pathogenesis of C. albicans, especially regarding to the Efg1 pathway.
Another system we are interested to study is the response of Candida albicans to the stimuli from the host. In addition to growth and proliferation on mucosal
surfaces, ingestion by human immune cells exposes C. albicans to novel
environments. The cells respond to phagocytosis of neutrophils by inducing genes related to arginine and methionine pathways, suggesting that the phagosome of the neutrophil is an amino acid-deficient environment; however, neither pathway is induced upon phagocytosis by monocytes. An analysis of its transcriptional response upon internalization by macrophages reveals that C. albicans activates alternate metabolic pathways, represses translation and induces genes related to the oxidative stress response, DNA damage repair, arginine biosynthesis and peptide utilization (Lorenz et al., 2004). These results suggest that the environment of the macrophage phagosome lacks usable nutrients (e.g. glucose) and contains reactive oxygen and nitrogen species, as part of its antimicrobial burst (Fang, 2004). Moreover, C. albicans encounters many antimicrobial proteins/peptides and innate defense molecules that act synergistically to combat infections. For example, some host proteins secreted onto the oral surface can directly inhibit Candida growth, morphogenesis, and adhesion through the action of antimicrobial peptides, including calprotectin (Sweet and Douglas, 1991), lysozyme (Laibe et al., 2003), low molecular weight salivary mucins (Satyanarayana et al., 2000), secretory leukocyte protease inhibitor (Chattopadhyay et al., 2004), lactoferrin (Samaranayake et al., 2001), and histatins (Lupetti et al., 2004). Finally, in the treatment of candidiasis, C. albicans also encounter various therapeutically antifungal drugs. All those stresses or cues induce the response of C. albicans cells and eventually via drug resistance and virulence/morphogenesis pathways send the signal to the controlling points of pathogenesis. Do all those signals eventually go to the well-known Efg1? Or do some of them go to other controlling points? For example, there are two potential candidates, Cph1 and Tup1. Therefore, we are interested in knowing which of those pathways linked to Efg1 and if not, where do those responses send their signals.
pathway and to expand the study by using genome-wide analysis to explore the networking and cross-talk between multiple components/pathways.
B. Specific Aims
The over-all long-term goal of this research is to elucidate the mechanisms of pathogenesis of Candida albicans. For current study, we propose to employ molecular genetics and functional genomics tools, particularly DNA microarray, to analyze and explore the signaling/regulatory pathways contributing to the
pathogenesis of C. albicans including the sensing of host environmental changes, iron availability, anti-microbial stresses, and morphogenesis/virulence. We are
particularly interested in the connection between those factors and the yeast to filament morphotypes transition and back, commonly believed to be associated with the commensal to pathogen transition. The first objective of this study is to focus on the functional study of the Efg1 pathway, known to involve in the regulation of this transition and the second objective is to establish a microarray platform to study the whole-genome profiling of the regulatory pathways and their global effects on
Candida albicans. In addition, this research may allow us to define particular gene products or signal transduction pathways that can be used as targets to block the transition or kill the fungal cells specifically. Therefore, the information obtained from this study would help us to understand the molecular mechanisms of fungal virulence/morphogenesis and in the next stage, to design and develop new antifungal drugs and/or new antifungal strategies on the molecular level.
Our specific aims for this proposal are:
Specific Aim 1. DNA microarrays for gene expression profiling concerning pathogenesis of C. albicans
a. Constructing DNA microarrays and optimizing them for analyzing gene expression profile of C. albicans. Data pre-processing and normalization will be initiated shortly after the construction of DNA microarrays such that it can be used to compare expression patterns across different experimental conditions and different strains. Linear mixed model is powerful for longitudinal studies and will provide statistical significance for the patterns observed and can select target features for cluster analysis.
b. Revealing the connection between different environmental cues known to affect the pathogenesis and the virulence/morphogenesis of C. albicans. We will employ the DNA microarray to compare responses of C. albicans to different stresses and conditions mimicing that of selected host and physical environments, including iron availability, oxidative stress, nitric oxide, and antimicrobial mediators.
c. Using DNA microarray to identify potential target genes and compare expression profiles mediated by different environmental cue and stress-responsive transcriptional regulators.
Specific Aim 2. Unveiling the molecular involvement of Efg1 pathway in the pathogenesis of C. albicans
Efg1 is a known key transcriptional control point for virulence, morphogenesis, and drug resistance in C. albicans, which establishes its position as a key player in the pathogenesis of Candida albicans. Therefore, we would like to determine the role of Efg1 pathway in the pathogenesis of C. albicans, starting from investigating the functions of CaEno1, a multi-functional glycolytic protein regulated by Efg1 and it is also a major component of cell wall as well as a secreted antigen in Candida infection. We will approach this goal in the following steps:
a. Understanding the molecular function of the Efg1-regulated CaEno1 with genetics study.
(1). By constructing Caeno1/Caeno1 homozygous mutant and to determine the effects on C. albicans.
(2). By investigating the signals on CaEno1 responsible for its various localities.
b. Identifying target genes of the Efg1 transcription factor by DNA microarray. (1). By comparing the gene expression patterns between the wild-type cells and the efg1/efg1 mutant cells using microarray assay.
(2). By identifying interested target genes involved in either virulence/morphological switch or drug resistance, and/or both.
Specific Aim 3. Functional characterization of genes of interest to map the gene networks and regulatory/signaling pathways.
a. Construction of stains containing mutations on each selected gene to determine the effects of mutations on host cells to elucidate the functions of the genes.
b. Elucidation of the signal transduction pathways regulating the control of pathogenesis, especially those associated with virulence/morphological switch, environmental stress, or drug resistance by mapping and correlating the functional of the selected genes.
C. Research Design and Methods
1. DNA microarrays for gene expression profiling of C. albicans
To construct DNA microarrays of C. albicans, the QIAGEN Operon 70mer
probe sets (Array-Ready Genome Oligo Set and Candida Genome AROS Upgrade Set) will be used. This oligo set contains 7,925 optimized probes that represent the entire genome of C. albicans and 10 different controls. The oligo set has been successfully used in studies of C. albicans biology and pathogenesis (Cao et al., 2006; Chen et al., 2004; Magee et al., 2003; Zhao et al., 2005).
a. RNA isolation, sample labeling and microarray hybridization.
Recognizing that one of the most important aspects of microarray analysis is the source and quality of the RNA used in these experiments, we will use standardization of protocols for RNA isolation as previously described (Lan et al., 2002; Lan et al., 2004). In general, cells grown at different conditions will be collected throughout lag, log and stationary phases of growth. RNA isolation, sample labeling and microarray hybridization will be performed using established protocols
(http://microarrays.org). Briefly, cells are harvested by centrifugation immediately after sampling; pellets are either snap-frozen in dry ice/ethanol or extracted in the presence of 15% SDS and buffered phenol:chloroform (1:1). Total RNA is
precipitated with absolute ETOH. After centrifugation, the RNA pellet is air dried and suspended in DEPC-treated water. For labeling, cDNA containing a T7 RNA polymerase recognition sequence are prepared from total RNA amplified with T7 RNA polymerase. After processing, blunt-ended cDNAs are used as templates to produce antisense RNA using a T7 Ampliscribe kit from Epicentre Technologies (Phillips and Eberwine, 1996). A second round of double-stranded cDNA synthesis is performed in the presence of random hexamer primers, reverse transcriptase and aminoallyl-dUTP. Cy3-/Cy5- dyes are incorporated into single-stranded cDNAs with monofunctional NHS esters that bind to free amino groups. Un-incorporated dye is removed by ultrafiltration using a Centricon 30 unit (Amicon). DNA microarrays will be analyzed using GenePix 4000B scanner and GenePix Pro 6.0 software (Molecular Device).
b. Experimental design and statistical analysis for expression profiling.
To derive accurate signals for the gene expression profiling, the first issue is the proper experimental design. Since the main purpose of this proposal is to understand the signaling/regulatory network involved in C. albicans response to environmental stimuli, especially the host environments, the number of time points measured is
crucial to the characterization for the upstream and downstream components. For most studies, at least five time points will be measured. Some effort will also be made in meta-analysis of the existing data to understand the variation and to decide the number of replicates needed for the expected significance level. At least five replicates are considered at this point and the number is subjected to change. With the high cost of microarray experiments, properly arranging the samples hybridized together with loop design in oligo spotted arrays can reduce the arrays needed without loss of too much information (Churchill, 2002). If not enough samples or arrays are available, pooling the samples is considered as a choice to get more reliable signals (Kendziorski et al., 2005).
Microarray data are known to be very noisy and no single statistical method has been recognized as standard approach for the normalization. The data derived will be analyzed with several different statistically solid algorithms. Image segmentation resultsfrom GenePix™,Spot(Jain et al., 2002), and model-based approaches will be compared and the one with the most reproducibility of replicates will be chosen for the next step. For the spotted oligo arrays proposed in this proposal, we will use ANOVA model (Wolfinger et al., 2001) to adjust for systematic noises. Also, global intensity-based patterns will be corrected with loess or quantile normalization when needed.
Gene expression profiles will be compared across a variety of conditions. Comparisons will include, for example, wild type and mutants strains, drug-treated and un-treated, and various environmental conditions. Since the experiment is designed according to the statistical significance needed, we will follow the longitudinal model decided well ahead of the experiment and to choose genes that show differential expression among the control and experimental group. Given the large number of genes compared, care will also be exercised to avoid proportionately large numbers of false positives. To properly assess differential gene expression, we will employ at least two approaches: (1) Significance Assessment for Microarrays (SAM) (Tusher et al., 2001) to control the false discovery rate (2) the one controls the family wise error rate, but avoids the conservatism of Bonferroni correlation by utilizing a step-down method (Dudoit and Speed, 2000).
Exploratory data analyses using cluster methods can allow us to find patterns embedded in the data set and provide us some clues of the major targets. There exist a number of publicly available programs that allow one to cluster genes on the basis of similarity of their expression patterns across multiple experiments. This type of analysis has proven a very useful predictor of the function of unknown genes, and determined previously unknown linkages between different signaling/regulatory
pathways. Analysis of the promoter region of members of a cluster is another criterion that has been used to strengthen a case for possible physiological connections between its members (Bussemaker et al., 2001). Datasets from our microarray analysis will be analyzed using various methods such as hierarchical cluster analysis (Eisen et al., 1998), self-organizing maps (Tamayo et al., 1999), k-means clustering (Tavazoie et al., 1999) and principal component analysis (Raychaudhuri et al., 2000).
2. Understanding the molecular involvement of Efg1-CaEno1 pathway in the pathogenesis of C. albicans
Studies of C. albicans and its potential mechanisms of pathogenesis have relied heavily on the expression of various phenotypes induced by environmental changes or by its morphogenetic transitions. The relationship of these conditions/pathways to one another is complex. Here, we propose to study the molecular mechanism of the Efg1-CaEno1 pathway in drug resistance and virulence using molecular genetic tools and to identify Efg1 target genes using gene expression profiling.
a. Construction of a Caeno1/Caeno1 mutant
Since there is no known plasmid of C. albicans, homology recombination will be the main approach to generating knockout and knock-in mutations and other genetic manipulation. And since C. albicans is a diploid organism without known sexual cycle, both alleles of a given gene will have to be replaced at the same time to generate mutations. A homozygous Caenol/Caeno1 mutant will be constructed by gene disruption method based on the homology recombination strategy described previously (Gerami-Nejad et al., 2001; Wilson et al., 2000; Wilson et al., 1999). Both copies of CaENO1 gene will be replaced by SAT1 cassette. A DNA fragment containing the SAT1 construct flanked with homology regions of CaENO1 at two extremities will be transformed into the C. albicans strain SC5314. The
transformants will be selected for the drug resistance marker and then screened for lose of the marker for the pup-out of the SAT1. Then, PCR product containing the
SAT1 sequence flanking with the CaENO1 homology regions at two extremities will
be transformed again into the CaENO1/Caeno1 strain to generate the Caeno1/Caeno1 homozygous mutant via homology recombination. Since CaEno1 is required for cell growth in the presence of glucose (Yang et al., 2006), the Caeno1/Caeno1
homozygous mutant will be constructed by selecting for SAT1+ transformants on the selective medium using glycerol as the carbon source. After selection for STA1-again, a DNA fragment containing the wild-type CaENO1 will be transformed into the Caeno1/Caeno1 homozygous mutant to generate the Caeno1/Caeno1::CaENO1
strain.
b. Determining whether CaENO1 is involved in morphogenesis/virulence, drug resistance in Candida albicans and other characterization of the Caeno1/Caeno1 mutant
To determine if CaENO1 regulates the morphogenesis of C. albicans, we will compare the wild-type and the Caeno1/Caeno1 mutant about their morphology, germ tube formation, colony formation, and cellular growth under filament-inducing condition including the addition of serum, temperature, pH, and other environment cues. Since the Efg1 also involved in drug resistance, we will also test the drug susceptibility of the Caeno1/Caeno1 mutant by Etest and/or agar dilution to unveil the connection between those pathways.
c. Characterization of new interested genes obtained by DNA microarray.
DNA microarray will be also employ to find out other genes regulated by Efg1. The interested candidates will be subjected to mutagenesis by homologous
replacement as described in (1). Heterozygous and homozygous null mutants will be functional characterized by comparing their phenotypes with that of the wild type strain. The phenotypes to be studied include cell growth, cell morphogenesis, susceptibility to antifungal drugs and sensitivity to different stress conditions.
Finally, the correlation of the genes of interest with virulence will be assessed using a mouse model of systemic infection.
d. Elucidation of the relationship between CaENO1 and other genes
Making double mutations on two genes to assess whether they have related function is an approach commonly used for studying gene functions (Lo et al., 1997). Thus, first of all, we will construct Caeno1/Caeno1 and new target gene double mutant. A PCR product containing the URA3-dpl200 sequence with the short homology regions (70 bps) flanking at the two extremities of another interested gene (NEW) will be transformed into Caena/Caeno1 to generate
NEW/new::dpl200-URA3-dpl200 Caeno1::ARG4/Caeno1::dpl200 strain. The
NEW/new::dpl200 Caeno1::ARG4/Caeno1::dpl200 cells will be selected by growing
the NEW/new::dpl200-URA3-dpl200 Caeno1::ARG4/Caeno1::dpl200 cells into a medium containing 5FOA.
The presence of 5FOA will select for the recombinants that have lost URA3. Again, the same PCR product containing the URA3-dpl200 sequence with the NEW short homology regions (70 bps) at two extremities will be transformed into
new::dpl200-URA3-dpl200/new::dpl200 Caeno1::ARG4/Caeno1::dpl200 double
mutant by selecting for Ura+ transformants.
3. The response of C. albicans to the environmental stimuli
Since the relationship of conditions/pathways of pathogenesis to one another is complex, we intend to study the possible cross-talk between the Efg1 pathway and other pathways of our interest.
a. Cell morphogenesis.
Efg1, Rim101, Nrg1 have been indicated to control cell morphogenesis. As described above, we will identify target genes of Efg1. To reveal genes that are commonly regulated by all three transcriptional factors or subsets of genes that are specifically regulated by one of the three factors, the target genes of Rim101 and Nrg1 will be also studied. Experiments will be performed by comparing expression
profiles between wild-type and mutants lacking functions of each transcriptional factor, and that between cell growth of yeast and hyphal forms. The mutant strains will be generated using the SAT1-flipper method (as described below) or the methods described above.
b. Iron-responsive gene regulation.
In the regulation of morphogenesis, Efg1 receives its upstream signal via a cAMP/PKA (protein kinase A)-dependent pathway (Ernst, 2000). Although that has not been studied in C. albicans, components of the cAMP/PKA pathway not only affect morphogenesis, but also affect iron-acqusition gene expression in S. cerevisiae. To explore the possibility of the cAMP/PKA and Efg1 pathway to control
iron-responsive gene expression, we will also generate null mutant of PKA. We will compare patterns of gene expression between wild-type and mutants lacking functions of PKA and Efg1, and that between cell grown in iron-limiting and iron-repletion conditions. In addition to cAMP/PKA and Efg1 pathway, other potential
transcriptional factors controlling gene expression in response to iron availability will also be examined. We are generating deletion mutation of C. albicans Orf19.2272, which encodes a protein with a high homology with S. cerevisiae Aft1p. In S.
cerevisiae, Aft1p is an activator for iron acquisition and many other iron-responsive
genes. The media representing iron-limiting and iron repletion conditions are used as previously described (Lan et al., 2004).
c. Other stress responses.
immune system, including the microbial killing mechanisms of phagocytosis.
Macrophages and neutrophils are the main components of the innate immune system and use reactive oxygen and nitrogen species to protect the host (Nathan and Shiloh, 2000). Superoxide readily dismutates to hydrogen peroxide or combines with nitric oxide to form strong oxidant peroxynitrite, which is fungicidal (Vazquez-Torres and Balish, 1997). In the presence of transition metals such as iron, hydrogen peroxide can even break down to form the highly reactive hydroxyl radical. Therefore, the regulation of iron may be of great importance to C. albicans to deal with oxidative stress.
To study the cross-talk between cell responses to iron availability and to oxidative stress, C. albicans will be treated with 0.4 mM H2O2or 0.5 mM menadione (a
superoxide generating agent) that allows the organism to tolerate ordinarily lethal levels of these oxidants (Jamieson et al., 1996). A comparison of S. cerevisiae and C.
albicans indicate that the latter can adapt much higher levels of reactive oxygen
species (Jamieson et al., 1996). This analysis will be the identification of genes that may functionally confer this ability. Revealing the overlap or difference between cell responses to iron and oxidative stresses will be important for our understanding to the survival/persistence of C. albicans in its host environment.
d. Functional characterization of genes of interest
It is the expectation that DNA microarray data obtained will allow us to identify significantly expressed ORFs of both known and unknown functions. From these data, we will be possible to identify a limited number of ORFs which are related to general functions of interest. To analyze functions of these genes of interest (GOI), we will construct target gene disruptions using the SAT1-flipper method (Reuss et al., 2004). This method relies on the use of a cassette that contains a dominant
nourseothricin-resistance marker (CaSAT1) for the selection of integrative
transformants and an inducible FLP recombinase system for subsequent excision of the cassette (Reuss et al., 2004). Briefly, the flanking sequences of GOI are located at the both sides of the cassette. Following integration of the marker cassette by homologous recombination of the GOI flanking sequences, transformants were grown in a medium containing 10% BSA (bovine serum albumin) to induce the recombinase for marker construct excision. Cells were plated at a low nourseothricin concentration (25g/ml) to identify SAT1-negative colonies which grew to a smaller size under these conditions than the colonies from SAT1-positive cells. Selecting the small colonies and plating on the high drug concentration (100g/ml) dish to make sure it is
SAT1-negative indeed. Two rounds of integration/excision result in the disruption of both alleles of the GOI.
Heterozygous and homozygous null mutants will be functional characterized by comparing their phenotypes with that of the wild type strain. The phenotypes to be studied include cell growth, cell morphogenesis, and susceptibility to antifungal drugs and sensitivity to different stress conditions. Finally, the correlation of the genes of interest with virulence will be assessed using a mouse model of systemic infection.
D. Results and Discussions
1: Construction of DNA microarrays for gene expression profiling concerning the pathogenesis of C. albicans
a. We have completed the construction of a set of DNA microarray of Candida
albicans as well as the tuning of noise and normalization of the background
of the data obtained from the array.
b. We have established a standard experimental procedure and operation platform for employment of the DNA microarray.
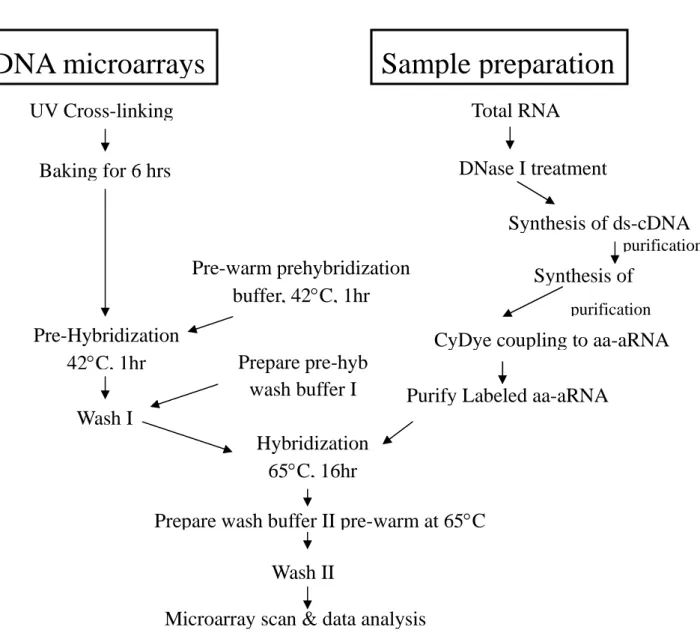
In order to study the response of C. albicans to its host environments and the functions of virulence factors of Candida albicans, we have set out to generate DNA microarrays representative of the whole genome of this pathogen. In the first year of this grant period, we have focused on the construction of DNA microarrays, optimization of sample preparation and microarray processing and facilitating practice for data analysis (an overview see Fig. 1). We have reached several goals that would provide important foundation for the continuing experiments in the next stages of the study.
(1) Construction C. albicans DNA microarrays. We have constructed the spotter
arrays for C. albicans. The QIAGEN Operon C. albicans Array-Ready Oligo Set (AROS) (v. 1.2) containing 6,266 probes and C. albicans AROS upgrade set (v. 1.1) containing 1,659 probes has been used. These 70mer oligo sets contain a total of 7,925 optimized probes that represent the entire genome of C. albicans and more than 10 different controls. The oligo sets have been also successfully used in the studies of different aspects of C. albicans biology and pathogenesis.
The C. albicans oligos were spotted onto the UltraGAPS coated slides (Corning, New York, NY) with the use of the OmniGrid 100 microarrayer (Genomic Solutions, Ann Arbor,MI),according to themanufacturer’sinstructions.To ensurethequality of the microarray printing, one or two microarrays of each batch are randomly chosen and are labed with the Cy5 fluorescent dye. The quality of the test spotted microarrays is determined by visualization. At this stage, we have generated less than one hundred such C. albicans spotted arrays.
(2) Optimization of experimental protocols for RNA isolation, sample labeling and microarray hybridization. In addition to the quality of microarray per se,
another important issues in microarray studies is the source and quality of the RNA used. To this end, we have examined various protocols for RNA isolation from either the yeast (balstspore) or filamentous cells of C. albicans. For the yeast form of C.
albicans, a single colony of the clinically isolated wild-type strain SC5314 was
inoculated into 10 ml of YPD medium and grown overnight at 30°C. The overnight cultures were harvested by centrifugation and the cell pellets were washed twice with sterile water. The same amount of cells (OD600= 0.05) were inoculated into YPD liquid medium and incubated at 30°C. To obtain hyphael cells, equivalent overnight cultures (OD600= 0.1) were inoculated into Lee's and YPD + 10% bovine serum and incubated at 37°C. Both yeast and filament forms of C. albicans were grown until the cells reached early to mid-exponential phase. At each sampling, each culture was immediately collected by centrifugation at 4°C, and the cell pellets were snap-frozen in an ethanol-dry ice bath and stored at −80°C. Total RNA was then extracted as previously described (Lan et al., 2004). Briefly, the cell pellets were resuspended in 3 ml of extraction buffer (0.1M LiCl, 0.02M dithiothreitol, 0.1M Tris-HCl, pH 7.5) and vortexed. Then, the cell suspension was added to an ice-cold tube containing 6g of acid-washed glass beads, 0.5 ml of 10% SDS, 5ml of phenol-holoroform. The mixture was vortexed for five minutes to disrupt the cells. After centrifugation, the upper aqueous phase of the mixture was transferred to a fresh tube. Finally, the samples were extracted by phenol/cholorform twice, ethanol precipitation, treated with RQ1 DNase (Promega, Madison, WI) for 1 h and cleaned by using an RNeasy Column (QIAGEN, Valencia, CA). For the hyphal cells, the protocol for cell disruption were slightly modified: the lysed cells were disrupted by five cycles or vortexing (3 min each); between cycles the tubes were incubated alternatively for 3 min at 42oC and then at 0oC. The quality and quantity of total RNA are determined using Bioanalyzer 2100 (Agilent Technologies) and/or ND-1000 spectrophotometer (NanoDrop Inc).
The optimal conditions for sample labeling and microarray hybridization are currently working in-progress. Briefly, cDNA containing a T7 RNA polymerase recognition sequence are prepared from total RNA amplified with T7 RNA polymerase. After processing, blunt-ended cDNAs are used as templates to produce antisense RNA using a T7 Ampliscribe kit. A second round of double-stranded cDNA synthesis is performed in the presence of random hexamer primers and reverse transcriptase; aminoallyl-dUTP. The Cy3-/Cy5-dyes are incorporated into single-stranded cDNAs with monofunctional NHS esters that bind to free amino groups. Un-incorporated dyes are removed by ultrafiltration using a Centricon 30 unit
(Amicon). DNA microarrays will be analyzed using Axon 4000B scanner (Axon Technologies). A representative of these tested microarrays is shown in Fig 2. In this figure, the overall good distributions, fine shapes of each oligo spots and fine hybridization are demonstrated.
(3) Practice toward statistical analysis for expression profiles. Microarray data
could be noisy and no single statistical method has been chosen as a standard approach for the data normalization. We have collected some of the data sets in the public domain and used these available data to practice for the statistical analysis. We have practiced for the three major steps of data analysis: (1) image processing (2) statistical analysis for differential expressed genes, and (3) clustering genes with similar time profiles. For image processing, we were focused on the most widely adopted software, GenePix Pro. With these practice, we try to extract reliable signals even though our budget can only support for the spotted microarrays, which may not be the best platform comparing to the commercially available ones. For statistical analysis of significant genes, we have used classical linear model approach in the JMP®Genomics software. Several results of our practices are shown in Fig 3.
Figure 1. An overview for working on the C. albicans oligo microarrays and data analysis.
Microarray scan & data analysis
Total RNA
Synthesis of ds-cDNA
CyDye coupling to aa-aRNA
Purify Labeled aa-aRNA
UV Cross-linking
Baking for 6 hrs
Pre-Hybridization
42C, 1hr
Hybridization
65C, 16hr
Wash II
Pre-warm prehybridization
buffer, 42C, 1hr
Wash I
Prepare pre-hyb
wash buffer I
DNase I treatment
Synthesis of
purification purificationPrepare wash buffer II pre-warm at 65C
Sample preparation
DNA microarrays
(A)
(B)
(A)
(B)
Figure 2. A representative of the C. albicans spotted oligo microarrays. (A) The
whole view of a test microarray with sample hybridized. (B) A portion of the microarray is enlarged and shown.
(A)
(B)
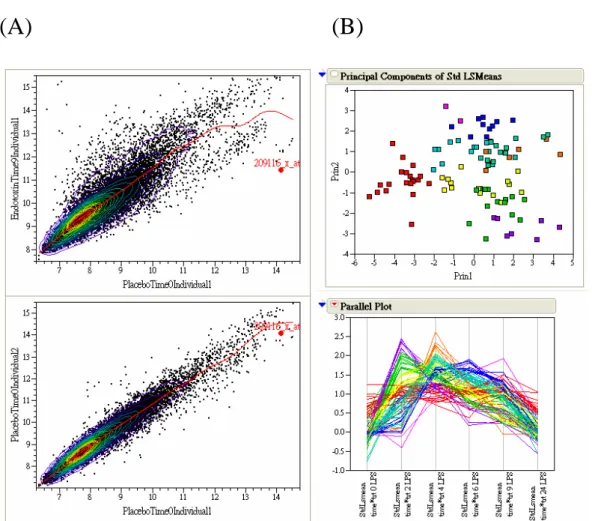
Figure 3. A parctice for the DNA microarray data analysis. (A) Scatter plot for two
pairs of arrays. The upper panel shows a scatter plot for two arrays that belong to different treatments and the lower panel shows a scatter plot for two arrays that belong to biological replicates of the same treatment. (B) Principal component analysis for time course data. In the upper panel, the first two principal components clearly separate the time profiles of a few hundreds of genes. The lower panel shows the clustering results with different groups indicated by different colors.
2: Unveiling the molecular involvement of Efg1 pathway in the pathogenesis of
C. albicans
a. We have constructed Caeno1/Caeno1 homozygous mutant for the purpose of understand the effects of the null mutation on C. albicans.
b. We have performed deletion analysis on CaENO1 for the purpose of determination of the signals on CaEno1 responsible for its various localities.
CaEno1 is a multi-functional glycolytic protein regulated by Efg1 and it is also a major component of cell wall as well as a secreted antigen in Candida infection. Its location in the cell is related to its function and subjected to the regulation of Efg1. Hence, understand the signal and mechanism will help to reveal the role of the Efg1 pathway and the controlling mechanism of pathogenesis. The approach we used is the mutagenesis analysis to unveil its functions.
In the first year of this research, our effort is to construct CaENO1 mutants for functional study. We started by constructing the knockout mutation of CaENO1 to understand the role of CaEno1p in morphogenesis/virulence. At the same time, we also fused various sequence fragments of CaENO1 to reporter GFP to monitor the location of the GFP to determine the sequences necessary for the various locations of CaEno1p, which is in connection to the virulence and pathogenesis of C. albicans.
(1). Constructing Caeno1/Caeno1 homozygous mutant and to determine the effects on C. albicans.
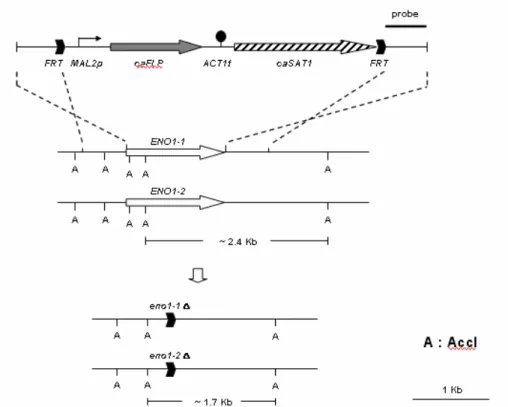
Due to the diploid-only nature of C. albicans and the lack of known plasmid, the homozygous mutation on CaENO1 is in fact a null mutation constructed by knock-out procedure based on homologous replacement. An SAT1 expression cassette was flanked by the 5’and 3’CaENO1 sequences of CaENO1, introduced by restriction
cloning (Fig. 4). This recombinant DNA fragment was then introduced into C.
albicans SC5314. Transformants were selected for nourseothricin resistance expressed by SAT1. In those cells, the SAT1 cassette replaces the center portion of
CaENO1 ORF after homologous recombination. However, the SAT1 cassette contains two FRT sequences, which will then recombine to pup out the SAT1
sequence, leaving truncated CaENO1 with only the 5’and 3’sequences. After PCR and Southern blot confirmation (Fig. 5), this heterozygous mutant is subjected for second round knock-out to remove the second allele of CaENO1. The resultants were assessed again by PCR and Southern analysis (Fig. 5). Since CaENO1 is known to be the sole gene encoding enolase in C. albicans, its null mutant will not be able to grow on media containing glucose, which is what has been observed (Fig. 6). In short, we have obtained the homozygous mutant strain and are in the process of
characterizing the effects on the phenotypes involved in morphogenesis and virulence.
Fig. 4. Genomic construction of CaENO1 null mutation. Acc1 is used to treat genomic DNA for Southern analysis.
Fig. 6. CaENO1 null mutant can not grow on YPD. YCH78: wild-type
CaENO1/CaENO1 strain; YCH61: eno1Δ::FRT/eno1Δ::FRT; YCH81: wild-type CaENO1/CaENO1 strain.
(2). Investigating the signals on CaEno1 responsible for its various cellular localizations.
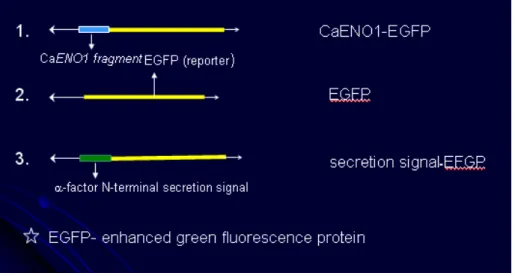
The attempt is to perform deletion analysis to narrow the sequences responsible for the locations of CaENO1 till the minimal sequences are defined. However, due to the difficulty of molecular manipulation in C. albicans, our strategy is to perform the deletion analysis in the baker yeast first, and then assess the result in C. albicans. Therefore, we have fused the sequence of CaENO1 to EFGP in a S. cerevisiae expression vector (Fig. 7) and then determined whether the CaEno1p expressed in S.
cerevisiae could still be retained in cytoplasm as well as secreted into media.
Plasmids carrying the secretion signal of-factor fused to the 5’of EGFP and plasmids carrying the EGFP gene alone serve as controls. The cells transformed with plasmids carrying the sequence of CaENO1-EGFP fusion can be seen to express EGFP (Fig. 8A), so does the plasmids carrying EGFP (Fig. 8B). Hence, the fusion of EGFP and CaENO1 can be expressed and the EGFP can serve as the reporter. In addition, cells transformed with plasmids carrying-factor secretion signal fused to the 5’of EGFP was also expressed but a lower intensity (Fig. 8C). One possible reason for the lower intensity is that the fusion proteins were indeed secreted. Hence, we decided to collect the media from the culture and analyzed with Western blot against EGFP. As it is shown in figure 9, Western analysis was able to reveal a band with the size of about 75 kD in the culture media of cells carrying the sequence of
CaENO1-EGFP fusion. Hence, the CaEno1p-EGFP is secreted into media as the wild-type CaEno1p shall have. We are now completed the construction of various truncated CaENO1 fused with EGFP to determine whether different portions of the sequence can direct EGFP to different cellular location.
Fig 7. Schematic representation of fusion construct of CaENO1 on baker yeast expression vector YEP363.
Fig 8. Plasmid-based CaENO1-EGFP can express in S. cerevisiae. Cells carrying different plasmids were observed under fluorescence microscope. A, cells contain plasmid carrying CaENO1-EGFP; B, cells contain plasmid carrying EGFP alone; C, cells contain plasmid carrying secretion signal-EGFP.
Fig. 9. Western analysis revealed that CaENO1-EGFP can be detected in media.
五、參考文獻
Baillie,G.S. and Douglas,L.J. (1998). Iron-limited biofilms of Candida albicans and their susceptibility to amphotericin B. Antimicrob. Agents Chemother. 42, 2146-2149.
Beck-Sague,C. and Jarvis,W.R. (1993). Secular trends in the epidemiology of nosocomial fungal infections in the United States, 1980-1990. National Nosocomial Infections Surveillance System. J. Infect. Dis. 167, 1247-1251. Bensen,E.S., Martin,S.J., Li,M., Berman,J., and Davis,D.A. (2004). Transcriptional
profiling in Candida albicans reveals new adaptive responses to extracellular pH and functions for Rim101p. Mol. Microbiol. 54, 1335-1351.
Berman,J. and Sudbery,P.E. (2002). Candida Albicans: a molecular revolution built on lessons from budding yeast. Nat. Rev. Genet. 3, 918-930.
Bussemaker,H.J., Li,H., and Siggia,E.D. (2001). Regulatory element detection using correlation with expression. Nat. Genet. 27, 167-171.
Calderone,R.A. and Fonzi,W.A. (2001). Virulence factors of Candida albicans. Trends Microbiol. 9, 327-335.
Cao,F., Lane,S., Raniga,P.P., Lu,Y., Zhou,Z., Ramon,K., Chen,J., and Liu,H. (2006). The Flo8 transcription factor is essential for hyphal development and virulence in Candida albicans. Mol. Biol. Cell. 17, 295-307.
Chattopadhyay,A., Gray,L.R., Patton,L.L., Caplan,D.J., Slade,G.D., Tien,H.C., and Shugars,D.C. (2004). Salivary secretory leukocyte protease inhibitor and oral
candidiasis in human immunodeficiency virus type 1-infected persons. Infect. Immun. 72, 1956-1963.
Chen,C.G., Yang,Y.L., Cheng,H.H., Su,C.L., Huang,S.F., Chen,C.T., Liu,Y.T., Su,I.J., and Lo,H.J. (2006). Non-lethal Candida albicans cph1/cph1 efg1/efg1
transcription factor mutant establishing restricted zone of infection in a mouse model of systemic infection. International Journal of Immunopathology and Pharmacology In press.
Chen,C.G., Yang,Y.L., Shih,H.I., Su,C.L., Lo,H.J., and . (2004). CaNdt80 is involved in drug resistance in Candida albicans by regulating CDR1. Antimicrob. Agents Chemother. 48, 4505-4512.
Churchill,G.A. (2002). Fundamentals of experimental design for cDNA microarrays. Nat. Genet. 32 Suppl:490-5., 490-495.
Csank,C., Costanzo,M.C., Hirschman,J., Hodges,P., Kranz,J.E., Mangan,M., O'Neill,K., Robertson,L.S., Skrzypek,M.S., Brooks,J., and Garrels,J.I. (2002). Three yeast proteome databases: YPD, PombePD, and CalPD (MycoPathPD). Methods Enzymol. 350:347-73., 347-373.
De,B., Magee,P.T., and Pla,J. (2000). Recent developments in molecular genetics of
Candida albicans. Annu. Rev. Microbiol. 54:463-98., 463-498.
Dudoit,S. and Speed,T.P. (2000). A score test for the linkage analysis of qualitative and quantitative traits based on identity by descent data from sib-pairs.
Biostatistics. 1, 1-26.
Edwards,E.J.J. (1990). Candida species. In Prinicples and Practice of Infectious Diseases. G.L.Mandell, R.G.douglas, and J.E.Bennett, eds. (New York: pp. 1943-1958.
Eisen,M.B., Spellman,P.T., Brown,P.O., and Botstein,D. (1998). Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci U. S. A. 95, 14863-14868.
Ernst,J.F. (2000). Transcription factors in Candida albicans - environmental control of morphogenesis. Microbiology. 146, 1763-1774.
Fang,F.C. (2004). Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nat. Rev. Microbiol. 2, 820-832.
Fratti,R.A., Belanger,P.H., Ghannoum,M.A., Edwards,J.E., Jr., and Filler,S.G. (1998). Endothelial cell injury caused by Candida albicans is dependent on iron. Infect. Immun. 66, 191-196.
Gerami-Nejad,M., Berman,J., and Gale,C.A. (2001). Cassettes for PCR-mediated construction of green, yellow, and cyan fluorescent protein fusions in Candida albicans. Yeast 18, 859-864.
invasion. Curr. Opin. Microbiol. 5, 366-371.
Heymann,P., Gerads,M., Schaller,M., Dromer,F., Winkelmann,G., and Ernst,J.F. (2002). The siderophore iron transporter of Candida albicans (Sit1p/Arn1p) mediates uptake of ferrichrome-type siderophores and is required for epithelial invasion. Infect. Immun. 70, 5246-5255.
Jain,A.N., Tokuyasu,T.A., Snijders,A.M., Segraves,R., Albertson,D.G., and Pinkel,D. (2002). Fully automatic quantification of microarray image data. Genome Res.
12, 325-332.
Jamieson,D.J., Stephen,D.W., and Terriere,E.C. (1996). Analysis of the adaptive oxidative stress response of Candida albicans. FEMS Microbiol. Lett. 138, 83-88.
Jones,T., Federspiel,N.A., Chibana,H., Dungan,J., Kalman,S., Magee,B.B., Newport,G., Thorstenson,Y.R., Agabian,N., Magee,P.T., Davis,R.W., and
Scherer,S. (2004). The diploid genome sequence of Candida albicans. Proc. Natl. Acad. Sci U. S. A. 101, 7329-7334.
Jong,A.Y., Chen,S.H., Stins,M.F., Kim,K.S., Tuan,T.L., and Huang,S.H. (2003). Binding of Candida albicans enolase to plasmin(ogen) results in enhanced invasion of human brain microvascular endothelial cells. J Med. Microbiol. 52, 615-622.
Kendziorski,C., Irizarry,R.A., Chen,K.S., Haag,J.D., and Gould,M.N. (2005). On the utility of pooling biological samples in microarray experiments. Proc. Natl. Acad. Sci U. S. A. 102, 4252-4257.
Laibe,S., Bard,E., Biichle,S., Vielle,J., Millon,L., Drobacheff,C., Seilles,E., and Meillet,D. (2003). New sensitive method for the measurement of lysozyme and lactoferrin to explore mucosal innate immunity. Part II: time-resolved
immunofluorometric assay used in HIV patients with oral candidiasis. Clin. Chem. Lab Med. 41, 134-138.
Lan,C.Y., Newport,G., Murillo,L.A., Jones,T., Scherer,S., Davis,R.W., and Agabian,N. (2002). Metabolic specialization associated with phenotypic
switching in Candida albicans. Proc. Natl. Acad. Sci. U. S. A 99, 14907-14912. Lan,C.Y., Rodarte,G., Murillo,L.A., Jones,T., Davis,R.W., Dungan,J., Newport,G.,
and Agabian,N. (2004). Regulatory networks affected by iron availability in
Candida albicans. Mol. Microbiol. 53, 1451-1469.
Lesuisse,E., Horion,B., Labbe,P., and Hilger,F. (1991). The plasma membrane ferrireductase activity of Saccharomyces cerevisiae is partially controlled by cyclic AMP. Biochem. J. 280, 545-548.
Lo,H.J., Kohler,J.R., DiDomenico,B., Loebenberg,D., Cacciapuoti,A., and Fink,G.R. (1997). Nonfilamentous C. albicans mutants are avirulent. Cell 90, 939-949.
Lo,H.J., Wang,J.S., Lin,C.Y., Chen,C.G., Hsiao,T.Y., Hsu,C.T., Su,C.L., Fann,M.J., Ching,Y.T., and Yang,Y.L. (2005). Efg1 involved in drug resistance by
regulating the expression of ERG3 in Candida albicans. Antimicrob. Agents Chemother. 49, 1213-1215.
Lorenz,M.C., Bender,J.A., and Fink,G.R. (2004). Transcriptional response of Candida
albicans upon internalization by macrophages. Eukaryot. Cell. 3, 1076-1087.
Lupetti,A., Brouwer,C.P., Dogterom-Ballering,H.E., Senesi,S., Campa,M., Van Dissel,J.T., and Nibbering,P.H. (2004). Release of calcium from intracellular stores and subsequent uptake by mitochondria are essential for the candidacidal activity of an N-terminal peptide of human lactoferrin. J Antimicrob. Chemother.
54, 603-608.
Magee,P.T., Gale,C., Berman,J., and Davis,D. (2003). Molecular genetic and genomic approaches to the study of medically important fungi. Infect. Immun. 71,
2299-2309.
Miller,L.G., Hajjeh,R.A., and Edwards,J.E., Jr. (2001). Estimating the cost of nosocomial candidemia in the united states. Clin. Infect. Dis. 32, 1110. Murillo,L.A., Newport,G., Lan,C.Y., Habelitz,S., Dungan,J., and Agabian,N.M.
(2005). Genome-wide transcription profiling of the early phase of biofilm formation by Candida albicans. Eukaryot. Cell. 4, 1562-1573.
Naglik,J., Albrecht,A., Bader,O., and Hube,B. (2004). Candida albicans proteinases and host/pathogen interactions. Cell Microbiol. 6, 915-926.
Nantel,A., Dignard,D., Bachewich,C., Harcus,D., Marcil,A., Bouin,A.P., Sensen,C.W., Hogues,H., van het,H.M., Gordon,P., Rigby,T., Benoit,F., Tessier,D.C.,
Thomas,D.Y., and Whiteway,M. (2002). Transcription profiling of Candida
albicans cells undergoing the yeast-to-hyphal transition. Mol. Biol. Cell 13,
3452-3465.
Nathan,C. and Shiloh,M.U. (2000). Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci U. S. A. 97, 8841-8848.
Niimi,M., Cannon,R.D., and Monk,B.C. (1999). Candida albicans pathogenicity: a proteomic perspective. Electrophoresis. 20, 2299-2308.
Okutomi,T., Tanaka,T., Yui,S., Mikami,M., Yamazaki,M., Abe,S., and Yamaguchi,H. (1998). Anti-Candida activity of calprotectin in combination with neutrophils or lactoferrin. Microbiol. Immunol. 42, 789-793.
Paul,T.R., Smith,S.N., and Brown,M.R. (1989). Effect of iron depletion on cell-wall antigens of Candida albicans. J Med. Microbiol. 28, 93-100.
Paul,T.R., Smith,S.N., and Brown,M.R.W. (1991). Influence of iron depletion and antifungal antibiotics on cell surface hydrophobicity of Candida albicans. Mycol.
Res. 95, 1312-1314.
Ramanan,N. and Wang,Y. (2000). A high-affinity iron permease essential for
Candida albicans virulence. Science. 288, 1062-1064.
Raychaudhuri,S., Stuart,J.M., and Altman,R.B. (2000). Principal components analysis to summarize microarray experiments: application to sporulation time series. Pac. Symp. Biocomput. :455-66., 455-466.
Reuss,O., Vik,A., Kolter,R., and Morschhauser,J. (2004). The SAT1 flipper, an optimized tool for gene disruption in Candida albicans. Gene. 341:119-27., 119-127.
Robertson,L.S., Causton,H.C., Young,R.A., and Fink,G.R. (2000). The yeast A kinases differentially regulate iron uptake and respiratory function. Proc. Natl. Acad. Sci U. S. A. 97, 5984-5988.
Samaranayake,Y.H., Samaranayake,L.P., Pow,E.H., Beena,V.T., and Yeung,K.W. (2001). Antifungal effects of lysozyme and lactoferrin against genetically similar, sequential Candida albicans isolates from a human immunodeficiency
virus-infected southern Chinese cohort. J Clin. Microbiol. 39, 3296-3302. Santos,R., Buisson,N., Knight,S., Dancis,A., Camadro,J.M., and Lesuisse,E. (2003).
Haemin uptake and use as an iron source by Candida albicans: role of
CaHMX1-encoded haem oxygenase. Microbiology. 149, 579-588.
Satyanarayana,J., Situ,H., Narasimhamurthy,S., Bhayani,N., Bobek,L.A., and
Levine,M.J. (2000). Divergent solid-phase synthesis and candidacidal activity of
MUC7 D1, a 51-residue histidine-rich N-terminal domain of human salivary
mucin MUC7. J Pept. Res. 56, 275-282.
Sigle,H.C., Thewes,S., Niewerth,M., Korting,H.C., Schafer-Korting,M., and Hube,B. (2005). Oxygen accessibility and iron levels are critical factors for the antifungal action of ciclopirox against Candida albicans. J Antimicrob. Chemother. 55, 663-673.
Soll,D.R. (2002). Candida commensalism and virulence: the evolution of phenotypic plasticity. Acta Trop. 81, 101-110.
Soll,D.R. (1997). Gene regulation during high-frequency switching in Candida
albicans. Microbiology. 143, 279-288.
Stehr,F., Felk,A., Gacser,A., Kretschmar,M., Mahnss,B., Neuber,K., Hube,B., and Schafer,W. (2004). Expression analysis of the Candida albicans lipase gene family during experimental infections and in patient samples. FEMS Yeast Res.
4, 401-408.
Sundstrom,P. (2002). Adhesion in Candida spp. Cell Microbiol. 4, 461-469.
Sweet,S.P. and Douglas,L.J. (1991). Effect of iron deprivation on surface composition and virulence determinants of Candida albicans. J Gen. Microbiol. 137,
859-865.
Tamayo,P., Slonim,D., Mesirov,J., Zhu,Q., Kitareewan,S., Dmitrovsky,E.,
Lander,E.S., and Golub,T.R. (1999). Interpreting patterns of gene expression with self-organizing maps: methods and application to hematopoietic
differentiation. Proc. Natl. Acad. Sci U. S. A. 96, 2907-2912.
Tavazoie,S., Hughes,J.D., Campbell,M.J., Cho,R.J., and Church,G.M. (1999). Systematic determination of genetic network architecture. Nat. Genet. 22, 281-285.
Theiss,S., Ishdorj,G., Brenot,A., Kretschmar,M., Lan,C.Y., Nichterlein,T., Hacker,J., Nigam,S., Agabian,N., and Kohler,G.A. (2006). Inactivation of the
phospholipase B gene PLB5 in wild-type Candida albicans reduces
cell-associated phospholipase A(2) activity and attenuates virulence. Int. J Med. Microbiol. ..
Tusher,V.G., Tibshirani,R., and Chu,G. (2001). Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci U. S. A. 98, 5116-5121.
Van der,S.D., Rodrigues-Pousada,R.A., Goodman,H.M., and Van Montagu,M. (1991). Plant enolase: gene structure, expression, and evolution. Plant Cell 3, 719-735. Vazquez-Torres,A. and Balish,E. (1997). Macrophages in resistance to candidiasis.
Microbiol. Mol. Biol. Rev. 61, 170-192.
White,T.C., Marr,K.A., and Bowden,R.A. (1998). Clinical, cellular, and molecular factors that contribute to antifungal drug resistance. Clin. Microbiol. Rev. 11, 382-402.
Wilson,R.B., Davis,D., Enloe,B.M., and Mitchell,A.P. (2000). A recyclable Candida
albicans URA3 cassette for PCR product-directed gene disruptions. Yeast 16,
65-70.
Wilson,R.B., Davis,D., and Mitchell,A.P. (1999). Rapid hypothesis testing with
Candida albicans through gene disruption with short homology regions. J
Bacteriol. 181, 1868-1874.
Wolfinger,R.D., Gibson,G., Wolfinger,E.D., Bennett,L., Hamadeh,H., Bushel,P., Afshari,C., and Paules,R.S. (2001). Assessing gene significance from cDNA microarray expression data via mixed models. J Comput. Biol. 8, 625-637. Yang,Y.L. (2003). Virulence factors of Candida species. J Microbiol. Immunol.
Infect. 36, 223-228.
Yang,Y.L., Chen,H.F., Kuo,T.J., and Lin,C.Y. (2006). Mutations on CaENO1 in
Candida albicans inhibit cell growth in the presence of glucose. J Biomed. Sci. 13, 313-321.
Microbiol Immunol Infect 34, 79-86.
Zhao,R., Daniels,K.J., Lockhart,S.R., Yeater,K.M., Hoyer,L.L., and Soll,D.R. (2005). Unique aspects of gene expression during Candida albicans mating and possible G(1) dependency. Eukaryot. Cell. 4, 1175-1190.
六、自評
As described above, in the first year of this project, we have successfully generated spotted oligo microarrays representative of the entire C. albicans genome. Also, we have tested and established an standard protocol for RNA isolation, sample processing and hybridization. To familiar with the handling of large amount of data in the near future, we practiced the microarray data analysis using public available data. Although there is not many experimental data shown in this report, we have gone through many tedious tests and practices; these initial steps will be crucial for the next stage to start different experiments. In sum, we matched quite well with the planned schedules for the specific aim 1, in the original grant proposal.
For the specific aim 2, we have successfully completed the construction of Caeno1/Caeno1 knock-out strain and have assessed the mutant to our satisfaction based on the result of PCR and Southern analyses. We are now in the process of characterizing the phenotypes related to morphogenesis/virulence and will complete the task in one month. Although there is a one-month delay, this is still in good accord with the planned schedule.
Another focus in the specific aim 2 is to identify the various localization signals of CaEno1p. Here we have successfully established the baker yeast system for our investigation. The CaENO1-EGFP was able to express in the baker yeast. And the fusion proteins appeared to be retained inside the cells as well as secreted into the surround media, which is consist with the behavior in C. albicans. We have now constructed various deletions on the fusion proteins to define the signals response to various cellular locations of CaENO1. This is in good agreement with the planned schedule.
In conclusion, for all three projects in this year, the one with the goal of
establishing DNA microarray platform has completed accordingly, the one with the aim for identify localization signals of CaEno1p is in progress matching the timetable, and the one with CaENO1null mutation is about one-month behind the schedule, a minor delay that shall not have any significant consequence. Therefore, the whole research is in good agreement with the original planning and the results will be suitable for academic publication within months.