European Journal of Plant Pathology 00:000-000
Verification of Serological Relationship between Two Phylogenetically
Related Peanut-Infecting Tospovirus Species
Ya-Chi Kang1,2, Shyi-Dong Yeh2,3, Chih-Hsuan Liao1, Wan-Chen Chou1, Fang-Lin Liu2,
Jia-Hong Dong4, and Tsung-Chi Chen1,5*
1 Department of Biotechnology, Asia University, Wufeng, Taichung 41354, Taiwan
2 Department of Plant Pathology, National Chung Hsing University, Taichung 40227, Taiwan 3 NCHU-UCD Plant and Food Biotechnology Program, Biotechnology Center, National
Chung Hsing University, Taichung, Taiwan
4 Yunnan Provincial Key Laboratory of Agricultural Biotechnology, Key Laboratory of
Southwestern Crop Gene Resources and Germplasm Innovation of Ministry of Agriculture, Biotechnology and Germplasm Resources Institute, Yunnan Academy of Agricultural Sciences, Kunming 650223, China
5 Department of Medical Research, China Medical University Hospital, China Medical
University, Taichung 40402, Taiwan
*Correspondence: Dr. Tsung-Chi Chen
E-mail: [email protected] Tel: 886-4-23394362 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Abstract
Kang, Y.-C., Yeh, S.-D., Liao, C.-H., Chou, W.-C., Liu, F.-L., Dong, J.-H., & Chen, T.-C. (2014). Verification of serological relationship between two phylogenetically related peanut-infecting Tospovirus species. European Journal of Plant Pathology, 00, 000-000.
Based on the serological relationships of nucleocapsid proteins (NPs), a tospovirus species can be classified as a member of a serogroup or a distinct serotype, which greatly
helps virus identification and disease diagnosis. Recent studies reported that distinct tospovirus species sharing above 51.8% amino acid (aa) identity in their NPs may be serologically related. Two phylogenetically related peanut-infecting tospovirus species, Peanut chlorotic fan-spot virus (PCFV) in Taiwan and Peanut yellow spot virus (PYSV) in India, were previously considered as distinct serotypes, since no serological relationship has been established. To verify the serological relationship of PCFV and PYSV, the NP of PCFV was purified from leaf tissues of the infected Chenopodium quinoa plants and used to produce
polyclonal antiserum (RAs-PCFV NP) and a monoclonal antibody (MAb-PCFV NP).
Polyclonal antiserum to the bacterially expressed NP of PYSV (RAs-PYSV NP) was also
prepared. RAs-PCFV NP reacted with the homologous PCFV NP and the bacterial-expressed
PYSV NP and tissue extracts of PYSV-infected plants. Reciprocally, RAs-PYSV NP reacted with the homologous bacterial-expressed PYSV NP and the tissue extracts of PYSV-infected or PCFV-infected plants. In addition, MAb-PCFV NP reacted only with the tissue extracts of PCFV-infected plants. Our results demonstrate that PCFV NP is serologically related to PYSV NP and they should be classified as members of a unique serogroup.
Keywords: Tospovirus; nucleocapsid protein; antibody; serogroup
25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49
Introduction
Tospovirus, designated from the type species Tomato spotted wilt virus (TSWV), is the only genus with plant-infecting viruses inthe family Bunyaviridae. Tospoviruses have quasi-spherical and lipoprotein-enveloped particles with diameters of 80 to 110 nm and possess a tripartite single-stranded RNA (ssRNA) genome, denoted large (L), middle (M) and small (S) segments based on the molecular size (King et al. 2011). All three RNA segments have a panhandle conformation formed by base paring of complementary nucleotides (nt) at the 5′- and 3′-ends (Mohamed 1981). Tospoviral RNA genome associates with the RNA-dependent RNA polymerase (RdRp) and the nucleocapsid protein (NP) to form infectious
ribonucleoproteins (RNPs) (Goldbach and Peters 1996; Mohamed et al. 1973). The L RNA is of negative polarity, coding the RdRp from the viral complementary (vc) strand for virus replication and transcription (de Haan et al. 1991; van Knippenberg et al. 2002). Each of M and S RNAs contains two open reading frames (ORFs) in an ambisense organization (de Haan et al. 1990; Kormelink et al. 1992; Law et al. 1992). The NP is encoded from the vc strand of S RNA to encapsidate viral RNAs (de Haan et al. 1990). Although vector specificity and host range of viruses are essential criteria for demarcating tospoviruses, sequence homology of NP with the threshold of 90% amino acid (aa) identity is the most important criterion for
establishment of a distinct species (King et al. 2011).
Tospoviruses cause serious losses in many economic crops, such as tomato, peanut, onion and melon, which are globally important (Pappu et al. 2009; Peng et al. 2011). The persistent transmissibility and pesticide resistance of thrips vectors and the insufficiency of natural resistance genes for breeding of crops make control of tospoviruses difficult. Virus diagnosis will help growers and researchers to adopt the measures, such as early removal of infected plants from cultivated farms and application of proper insecticides, for preventing the
50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74
epidemics of tospovirus diseases. Accurate virus detection is also crucial for the inspection and quarantine of tospoviruses in international trade.
NP is usually used as the target for diagnosis of a tospovirus due to its abundance in infected plant tissues. Antisera against NPs are commonly used for diagnosis and
identification of tospoviruses. Although the International Committee on Taxonomy of Viruses (ICTV) only recognizes family, genus and species as formal taxa, a more practical classification of tospoviral species is based on serological relationships among their NPs (Jan et al. 2003; Pappu et al. 2009). According to the serological relatedness, tospoviruses
clustered in a serogroup are difficult to be distinguished by NP-specific antisera because of cross reactions, but they usually can be identified by monoclonal antibodies (MAbs) (Chen et al. 2010; Lin et al. 2005). Originally, the numeric system, such as serogroups I, II, III and IV, was used to describe grouping of tospovirus species (de Ávila et al. 1990; de Ávila et al. 1993; Goldbach and Kuo 1996). However, the numeric system becomes confusing after more tospoviruses were discovered. A new serological classification system using type member to assign serogroup has been accepted up to now (Chu et al. 2001; Pappu et al. 2009). Most of the characterized tospovirus species are classified into three major serogroups, each with type member of TSWV, Watermelon silver mottle virus (WSMoV) or Iris yellow spot virus
(IYSV) (Chen et al. 2010; Chu et al. 2001; Ciuffo et al. 2008). Although some newly identified tospoviruses, such as Bean necrosis mosaic virus (BeNMV) (de Oliveira et al. 2011) and Soybean vein necrosis-associated virus (SVNaV) (Zhou et al. 2011), have been phylogenetically clustered in a group (de Oliveira et al. 2012), their serological status still need to be clarified. In addition, Impatiens necrotic spot virus (INSV) (Law and Moyer 1990), Peanut yellow spot virus (PYSV) (Satyanarayana et al. 1998) and Peanut chlorotic fan-spot virus (PCFV) (Chu et al. 2001) were considered as distinct monospecies serotypes because their serological relationships with other tospovirus species have not been reported 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99
(Chu et al. 2001; Jan et al. 2003).
Previous studies reported that Melon severe mosaic virus (MeSMV) and Tomato
chlorotic spot virus (TCSV) share 59.7% aa identity in their NPs and they are serologically
related (Ciuffo et al. 2009). Melon yellow spot virus (MYSV) sharing 58% aa identity with WSMoV in NP was previously classified as a distinct serotype (Kato et al. 2000), but has been recently rearranged into WSMoV serogroup because it has strong serological cross-reactions with the members of WSMoV serogroup (Chen et al. 2010). The NP of newly characterized Tomato necrotic ringspot virus (TNRV) shares 58.2%, 56.0% and 51.8% identity, respectively, with those of Capsicum chlorosis virus (CaCV), WSMoV and MYSV, and all these tospoviruses reacted with the polyclonal antibody “PAb MYSV6”made against the NP of MYSV (Seepiban et al. 2011). Noteworthily, the NP of PCFV, a peanut-infecting tospovirus discovered in Taiwan, shares 67.5% aa identity with that of PYSV from India (Chu et al. 2001). Although PCFV and PYSV have been clustered in an evolutionary group based on the phylogenetic analysis (Chu et al. 2001), their serological relationship has not been clarified yet. In this investigation, polyclonal and monoclonal antibodies against the respective NPs of PCFV and PYSV were produced to verify the serological relatedness of these two tospoviruses. Our results showed that PCFV and PYSV are serologically related and should be classified as a single serogroup.
Materials and methods
Virus sources
PCFV isolated from peanut (Chu et al. 2001), WSMoV (Yeh et al. 1988) and MYSV (Chen et al. 2008) from watermelon, and Calla lily chlorotic spot virus (CCSV) from calla lily (Chen et al. 2005a) were collected in Taiwan. A high temperature-recovered gloxinia isolate 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125
(HT-1) of CaCV from the United States was provided by H.-T. Hsu (Hsu et al. 2000). TSWV isolated from tomato in New York (TSWV-NY) was provided by R. Provvidenti (Chu et al. 2001). A tomato isolate of Groundnut ringspot virus (GRSV) collected from Brazil was gifted from D. Gonsalves (Pang et al. 1993). INSV originated from impatiens in United States (INSV-M) was provided by J. Moyer (Law and Moyer 1990). The typical isolate of TCSV (de Ávila et al. 1993) was obtained from German Collection of Microorganisms and Cell Cultures, Germany (Import Permit No.: 96-V-56). The original isolates of IYSV (Cortes et al. 1998) and Tomato yellow ring virus (TYRV) (Hassani-Mehraban et al. 2005) were given by R. Kormelink at Wageningen University, The Netherlands (Import Permit No.: 96-V-54). Peanut bud necrosis virus (PBNV) from tomato and Watermelon bud necrosis virus (WBNV) (Li et al. 2011) from watermelon were obtained from P. A. Rajagopalan of Mahyco Seeds Company, India (Import Permit No.: 97-V-41). All virus cultures were maintained in the local lesion host Chenopodium quinoa Willd. and the systemic host Nicotiana
benthamiana Domin in a temperature controlled greenhouse (25-28oC) under isolated
conditions and used for this study.
Purification of PCFV NP from plant tissue
Leaves of PCFV-infected C. quinoa plants were used for the purification of viral nucleocapsid according to the previously described method with modifications (Yeh et al. 1996). Briefly, the infected leaves 4 days post-inoculation were homogenized with TB buffer (10 mM Tris-HCl, pH 8.0, containing 10 mM Na2SO3 and 0.1% cysteine; 300 ml/100 g tissue)
by a blender, and the supernatant was collected by centrifugation in a Beckman JA-14 fixed angle rotor at 15300 ×g for 10 min. The supernatant was treated with 1% Triton X-100 at 4oC
for 30 min, and then centrifuged at 79700 ×g for 2.5 hr through a 20% sucrose cushion in a Beckman Type 35Ti rotor to collect the pellet. The pellet was resuspended in TBG buffer 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150
(TB buffer containing 10 mM glycine) for isopycnic centrifugation through 35% Cs2SO4 at
83500 ×g for 16 hr in a Beckman SW41 rotor. The opalescent zone containing viral
nucleocapsid was collected and centrifuged at 163600 ×g for 1 hr in a Beckman 70Ti rotor. The pellet was resuspended in TBG buffer, and the included proteins were separated by 12% SDS-PAGE. The desired bands corresponding to NP were cut and eluted from gel by a Model 442 Electro-Eluter (Bio-Rad, Hercules, CA). The yield of the purified NP was estimated by the software Spot Density of AlphaInnotech IS2000 (AlphaInnotech
Corporation, San Leandro, CA) by comparison with the quantified bovine serum albumin (BSA) (Chen et al. 2005b).
Expression of PYSV NP in bacterial pET system
The nt sequence corresponding to the reported full-length NP ORF of PYSV (accession number AF013994) carrying an NcoI site and a XhoI site at the 5′- and 3′-ends, respectively, was artificially synthesized and cloned in a pUC57 vector (GMbiolab Co, Ltd., Taichung, Taiwan). The nt sequence of the synthetic PYSV N gene was confirmed by sequencing. The DNA fragment corresponding to the NP ORF of PYSV was released from the pUC57 vector by digestion with NcoI and XhoI and then ligated with the pET-28b(+) vector (Novagen, Madison, WI) digested with the same enzymes. The recombinant plasmid was introduced into competent cells of Escherichia coli DH5α for multiplication. The plasmid was then isolated from E. coli DH5α and transferred into E. coli Rosetta BL21(DE3) cells (Novagen, Madison, WI) for protein expression.
Expression of the pET-derived PYSV NP was induced following the manufacturer’s instruction with modifications for purification. Briefly, 0.5-ml overnight culture starters (OD600≦ 0.5) were added into 50 ml LB medium containing kanamycin (15 g/ml) and
chloramphenicol (34 g/ml), and incubated at 37oC for 2 hr with shaking at 220 rpm (the cell
151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175
number reaches OD600=1.0). Isopropyl β-D-1-thiogalactopyranoside (IPTG) was then added
to final concentration of 1 mM to induce the expression of fusion proteins. After a 4 hr-induction, bacterial cells were collected by centrifugation at 8000 ×g at 4oC for 10 min. The
pellets were resuspended in 5 ml lysozyme buffer (40 mM Tris-HCl, pH 7.5, 1 mM EDTA and 150 mM NaCl) containing 1 mg/ml lysozyme (Bio Basic, Canada) and incubated at 37oC
for 30 min. The lysates were then sonicated with 100% amplitude on ice for 1 hr. After sonication, the lysates were centrifuged at 8000 ×g for 10 min. The supernatants and pellets were separately analyzed by 12% SDS-polyacrylamide gel electrophoresis (PAGE) and western blotting detected by rabbit antiserum (RAs-His) against the histidine tag (Viogene, Taipei, Taiwan) to evaluate the solubility of the expressed fusion proteins.
Purification of bacterial expressed PYSV NP
Because the NP of PYSV was expressed as soluble fusion protein, the supernatants collected from centrifugation after sonication were filtered through 0.45-μm filters
(Millex®HA, Millipore) to remove large impurities. Subsequently, the lysates were loaded
onto columns stuffed with Ni2+-NTA resins (Ni-NTA SUPERFLOW, Qiagen, Germany)
which were pre-equilibrated with binding buffer (20 mM Tris-HCl, pH 7.9, and 500 mM NaCl). The resins were washed with twofold volume of binding buffer containing 5 mM imidazole. The fusion proteins bound to the resins were eluted with 10 ml of binding buffer containing 500 mM imidazole. Each fraction of the purification steps was monitored by RAs-His (Viogene) in western blotting. The recombinant PYSV NP was further purified by gel electrophoresis method and quantified as described above.
Preparation of rabbit antisera
Antisera were produced from New Zealand white rabbits as described previously with modifications (Yeh and Gonsalves 1984). The purified PCFV and PYSV NPs (500 μg in 1 ml 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201
of PBS were separately emulsified with an equal volume of Freund’s complete adjuvant (Difco Laboratories, BD, NJ) and injected subcutaneously into individual rabbits.
Subsequently, 500 μg of immunogens in 1 ml of PBS emulsified with an equal volume of Freund’s incomplete adjuvant (Difco Laboratories) were administered weekly for 3 times. The rabbits were bled weekly from the ear marginal veins 1 week after the fourth injection.
Preparation of mice monoclonal antibody
The 6 to 8 weeks old female BALB/cByJ mice (Academia Sinica, Taipei) were
immunized by the first intraperitoneal injection with 50 μg of the purified PCFV NP in 250 μl PBS emulsified with an equal volume of Freund’s complete adjuvant (Difco Laboratories), and the same amount of protein emulsified with Freund’s incomplete adjuvant (Difco
Laboratories) was used for the following two intraperitoneal injections at weekly intervals. A final intraperitoneal injection using the same antigen without adjuvant was conducted 3 days before sacrifice of mice. Spleen cells of the sacrificed mice were harvested for fusion with Fox-NY myeloma cells (American Type Culture Collection, Manassas, VA20108) to obtain hybridoma cells following the method described previously (Hsu et al. 1984). Cultured media were used for screening by indirect enzyme-linked immunosorbent assay (ELISA) using crude antigen from leaves of PCFV-infected N. benthamiana plants. The selected hybridoma cells were further cloned by limiting dilution to obtain stable hybridoma cell lines. The Pristane-primed BALB/cByJ mice were intraperitoneally injected with 1.0× 106 hybridoma
cells to produce antibody-containing ascitic fluids.
Western blotting
Leaves of N. benthamiana plants infected with individual tospoviruses and the frozen leaves of PYSV-infected chili pepper plants collected from fields in Yunnan, China in 2008 (isolate SP-C, GenBank accession code: HQ402596) were ground in protein sample buffer 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227
[50 mM Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate (SDS), 12% glycerol, 0.01%
bromophenol blue and 2% -mercaptoethanol] at a 1/50 dilution. The extracts were boiled for 3 min and centrifuged at 17000 ×g for 3 min. The supernatants were separated in 12% SDS-PAGE and then transferred onto nitrocellulose (NC) membranes in transfer buffer (25 mM Tris-HCl, pH 7.4, 192 mM glycine and 20% methanol) on ice using a Bio-Rad mini transblot cell at 120 V for 30 min. The produced rabbit antisera and mice MAb were diluted at proper dilutions for determining their reactivity. RAs-His (Viogene) was used at a 1/2000 dilution for detecting the presence of the bacterial expressed PYSV NP. Alkaline phosphatase (AP)-conjugated goat anti-rabbit IgG and AP-(AP)-conjugated goat anti-mouse IgG (Jackson Immuno Research Laboratories, Inc., West Grove, PA), each diluted in TSW buffer (10 mM Tris-HCl, pH 7.4, 154 mM NaCl, 4% gelatin, 0.1% Triton X-100 and 2% SDS; 1/5000 dilution), was used for detecting rabbit antisera or mice MAb, respectively. The reactions for primary and secondary antibodies were performed at room temperature for 40 min. Color development was conducted by adding 50 μl of 5% NBT (nitro blue tetrazolium chloride) and 25 μl of 5% BCIP (5-bromo-4-chloro-3-indoyl phosphate) in 7.5 ml substrate buffer (100 mM Tris-HCl, pH 9.5, 100 mM NaCl and 5 mM MgCl2). The reaction was stopped by submerging the NC
membranes in water.
Indirect ELISA
Indirect ELISA was conducted according to the method described previously with modifications (Yeh and Gonsalves 1984) and used to determine the titers of the produced rabbit antisera and mice MAb, and the serological relationships of tospoviruses. Crude extracts (200 μl) of virus-infected tissues of N. benthamiana plants at a 1/50 dilution in coating buffer (15 mM Na2CO3, 34 mM NaHCO3 and 3 mM NaN3) were used for coating.
Rabbit antisera to PFCV NP (1/2000) and PYSV bacteria-expressed NP (1/4000) and mice 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252
MAb to PCFV NP (1/10000) were diluted in the conjugate buffer (PBST containing 2% PVP-40 and 0.2% ovalbumin) and used as the primary antibodies. AP-conjugated goat anti-rabbit IgG and AP-conjugated goat anti-mouse IgG (Jackson Immuno Research Laboratories), each diluted at a 1/5000 dilution, were used for detecting rabbit antisera and mouse MAb,
respectively. Color-developing solution was prepared by dissolving ρ-nitrophenyl phosphate disodium hexahydrate (ρ-NPP) in substrate buffer (9.7% diethanolamine and 3 mM NaN3) to
a final concentration of 1 mg/ml and 180 μl of solution was loaded to each well. The
absorbance at 405 nm (A405) was recorded by Bio-Rad Model 680 microplate reader for 1-2 hr
after the addition of enzyme substrate.
Phylogenetic analysis
Nucleotide sequences of the S RNAs or N genes of individual tospoviruses were obtained from GenBank (National Center for Biotechnology Information (NCBI) website http://www.ncbi.nlm.nih.gov/), and their accession codes are listed in Table 1. Multiple sequence alignments were performed by the ClustalW program (Biology Workbench, San Diego Supercomputer Center (SDSC), http://workbench.sdsc.edu/). Phylogenetic analyses were conducted using Phylip 3.69 (Felsenstein 1989). Bootstrapping was produced based on 1,000 replicates, and versions of the input data sets were resampled by the Seqboot program. Distance matrixes of amino acid sequences were produced by the Protdist program using PAM matrixes of Dayhoff model (Dayhoff et al. 1979). Phylogenetic branches were set by the Neighbor program using the Neighbor-Joining method (Saitou and Nei 1987).
Phylogenetic trees were produced by the Consense program of Phylip 3.69. Comparison of the NP sequences of PCFV and PYSV was carried out by the BL2SEQ program (Biology Workbench, SDSC). 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278
Results
Titration of the produced antisera and MAb
The rabbit antiserum against PCFV NP, denoted RAs-PCFV NP, was produced and its dilution end-point was determined as 1/16000 by indirect ELISA, based on the average reading more than twofold higher than the average reading of the healthy negative control, (Fig. 1a). Accordingly, RAs-PCFV NP was used at a1/2000 dilution for further tests. For MAb production, a stable hybridoma cell line 162H5H1 was selected and injected into mice to produce ascitic fluids. The ascitic fluids containing MAb against the PCFV NP were designated as MAb-PCFV NP. The dilution end-point of MAb-PCFV NP was determined as10-6 by indirect ELISA (Fig. 1b). Consequently, the10-4-diluted ascitic fluid was used for
further assays. The produced rabbit antiserum against the bacterial expressed NP of PYSV was named RAs-PYSV NP. The purified bacterial expressed immunogen was used to react with RAs-PYSV NP in this study because of the unavailability of fresh PYSV-infected plant material. The dilution end-point of RAs-PYSV NP was determined as 1/128000 by indirect ELISA (Fig. 1c). Therefore, RAs-PYSV NP was used at a 1/4000 dilution for further assays.
Serological relationship between PCFV NP and PYSV NP analyzed by reciprocal reactions with the prepared antisera and MAb
The crude extract of PCFV-infected N. benthamiana leaves and the purified PCFV NP were used for assays in western blotting (Fig. 2a). Both crude and purified antigens of PCFV, with an estimated molecular size of 31 kDa, strongly reacted with RAs-PCFV NP, MAb-PCFV NP and RAs-PYSV NP. However, no responses were observed when RAs-MAb-PCFV NP, MAb-PCFV NP and RAs-PYSV NP were used to incubate with the crude extract from the healthy control (Fig. 2b-d).
In addition, when the total proteins of the IPTG-induced bacterial cells and the eluted 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304
proteins from Ni2+-NTA affinity chromatography were used to incubate with the produced
antisera and MAb, an estimated 31 kDa of the recombinant PYSV NP was noticed (Fig. 3a). The recombinant PYSV NP containing a histidine tag at the C terminus was verified by RAs-His in western blotting (Fig. 3b). Thus, both crude and purified bacteria-expressed PYSV NP strongly reacted with RAs-PYSV NP and RAs-PCFV NP (Fig. 3c and d).
When total protein of the IPTG-induced bacterial culture containing the empty vector pET-28b(+) was used to incubate with the tested antisera and MAb, no signals were noticed. Moreover, the frozen leaf samples of PYSV-infected chili pepper plants collected in Yunnan Province, Southern China, were used to incubate with the produced antisera and MAb in western blotting. Our results revealed that the PYSV-infected samples positively reacted with the two antisera of RAs-PCFV NP and RAs-PYSV NP, but did not react with MAb-PCFV NP (Fig. 4).
Serological reactions of the produced antisera and MAb with other tospoviruses
Indirect ELISA was conducted to evaluate the serological reactions of the produced antisera and MAb with other tospoviruses. Crude leaf extracts of N. benthamiana plants infected with individual tospoviruses, including CaCV, CCSV, GRSV, INSV, IYSV, MYSV, TCSV, TSWV, TYRV, PBNV,PCFV, WBNV and WSMoV, and the purified bacterial expressed PYSV NP were used for assays. The crude extract of healthy N. benthamiana leaf was used as the negative control. All ELISA tests were duplicated, results showed that RAs-PYSV NP strongly reacted to the crude antigen of PCFV (the mean of readings =
2.546±0.113) and the homologous PYSV antigen (the mean of readings = 1.807±0.013), but did not react with antigens of other tospoviruses and the negative healthy control (Fig. 5a). RAs-PCFV NP strongly reacted with the homologous crude antigen (the mean of readings = 2.014±0.056) and moderately reacted with the bacteria-expressed PYSV NP (the mean of 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329
readings = 0.395±0.005). No positive reaction was noticed when RAs-PCFV NP was used to react with the crude antigens of other tospoviruses and the negative control (Fig. 5b). MAb-PCFV NP only specifically reacted with the crude sap of N. benthamiana infected with MAb-PCFV (the reading mean of 2.283±0.102) (Fig. 5c). Taken together, the serological relationship between PCFV NP and PYSV NP is reciprocally evidenced by the produced polyclonal antibodies.
Phylogenetic relationship among tospoviruses
Phylogenetic analysis of the NPs showed that the current tospoviruses are divided to five clusters (Fig. 6). PCFV and PYSV are closely related, belonging to the same cluster that is genetically distinct from all other known tospoviruses. Alignment of the NP sequences of PCFV and PYSV revealed that the contiguous consensus sequences are present around both NPs (Fig. 7).
Discussion
The serological sub-classification of tospoviruses is important for virus detection. Using the NP antiserum or MAb in ELISA is the most convenient and cost saving method for the identification of a tospovirus. However, tospovirus species clustered in a serogroup are usually misidentified by antisera. For instance, MYSV was previously mistaken for WSMoV in Taiwan by its cross reactions with the antisera against the NP of CaCV or WSMoV (Chen et al. 2008; Chen et al. 2010). Although MAbs can be used to distinguish most different tospovirus species, WSMoV, CaCV, PBNV and WBNV are still indistinguishable by the available MAbs (Chen et al. 2005b; Chen et al. 2010). The aa identities and similarities of above 85% and 90%, respectively, in tospoviral NPs are considered unable to distinguish the 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355
corresponding viruses using antisera or MAbs prepared from the native viral proteins (Chen et al. 2010). Similar results were also reported in the serological responses of the antisera and MAbs against the NSs protein. The rabbit antisera produced from a 24-aa peptide,
YFLSKTLEVLPKNLQTMSYLDSIQC, conserved region of the tospoviral NSs proteins can be used to detect TSWV, GRSV, TCSV and INSV (Heinze et al. 2000), which were clustered as a geographic group (Pappu et al. 2009). The MAbs targeting the consensus sequence VRKPGVKNTGCKFTMHNQIFNPN of the NSs proteins of tospoviruses classified in the WSMoV and IYSV serogroups, which are collected as another geographic group (Pappu et al. 2009), can broadly react with the members of these two serogroups (Chen et al. 2006; Chen et al. 2011). Thus, serogroup-specific MAbs could be used for diagnosis of tospoviruses.
The serological relationship of tospoviruses can be deduced by homology and
phylogenetic analysis of NPs (Chen et al. 2010; Seepiban et al. 2011). The previous study reported that tospovirus species sharing the NP homology above 51.8% identity is
serologically related (Seepiban et al. 2011). Due to the presence of contiguous consensus sequences, the generation of broad-spectrum antibodies against the common epitopes is possible even the aa identity of two tospoviral NPs is low. Nevertheless, the reactions of NP-specific antibodies could be diverse resulting from the different epitope-binding
characteristics. Hence, it is necessary to investigate the serological relationship of phylogenetically related tospoviruses.
PCFV and PYSV are phylogenetically related (Fig. 6), which share 67.5% identity and 86% similarity of NP, and have the similar biological properties such as peanut infecting and transmission by thrips vector Scirtothrips dorsalis Hood (Chen and Chiu 1996; Reddy et al. 1991). However, the two viruses have different geographic distribution (Chu et al. 2001; Satyanarayana et al. 1998). Although the contiguous consensus sequences are present between the NPs of PCFV and PYSV (Fig. 7), both viruses were considered as distinct 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374 375 376 377 378 379 380
serotypes due to the lack of a direct evidence of serological comparison (Chu et al. 2001; Satyanarayana et al. 1998).
In this investigation, the serological relationship between PCFV and PYSV is reciprocally verified by the cross reactions using the two rabbit antisera against the
corresponding NPs. Because the infectious PYSV is unavailable, its NP ORF was expressed in E. coli for the preparation of the antiserum. However, the MAb to the PYSV NP was not produced because the lack of PYSV-infected plant tissue for screening of hybridoma lines.
Some issues are usually concerned when the pET-expressed proteins were used for production
of antibodies. For example, a protein expressed in bacteria is normally produced as fusion form with a histidine tag at the N or C terminal end for detection and purification. On the other hand, the gene expression of prokaryotic cells is insufficient in post-translational modifications and the fused tag may cause conformational change of the expressed proteins. Both the antiserum against bacterial-expressed PYSV NP and the antiserum against the native PCFV NP showed high specificity to the corresponding homologous antigens, with low backgrounds to the crude extracts from the healthy control plant in indirect ELISA (Fig. 1), indicating that the bacterially expressed NP of PYSV has serological reactions similar to the native NP of PCFV. Therefore, the positive reactions of RAs-PYSV NP with the native PCFV NP and RAs-PCFV NP with the pET-expressed PYSV NP in both western blotting and indirect ELISA demonstrate the serological relationship of PYSV and PCFV.
To verify the serological reactions of the antisera and MAb with the native PYSV antigen, the chili pepper isolates of PYSV, such as SP-C, collected from Yunnan, China were recently provided by Yunnan Key Laboratory of Agricultural Biotechnology, Biotechnology and Genetic Germplasm Institute, Yunnan Academy of Agricultural Sciences for assays. The NP sequence of PYSV SP-C (GenBank accession code HQ402596) shares 94.3% aa identity with that of the original PYSV isolate (accession code AF013994) and thus the assignment as 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396 397 398 399 400 401 402 403 404 405
PYSV is validated. The serological relationship between PCFV and PYSV was directly confirmed by the positive reactions of PYSV SP-C with the homologous and heterologous antisera prepared in this study (Fig. 4). Since the instability of tospovirus, our attempts to isolate the infectious PYSV from the frozen samples were not successful. However, our western blot analysis using the field samples of PYSV clearly verified the serological cross-reactions of PYSV and PCFV by the antisera produced in this study.
Two obvious protein bands were noticed from the western blotting profile of the pET-PYSV-expressing protein and the eluted recombinant PYSV NP (Fig. 3a). These two protein bands can be recognized by RAs-His, RAs-PYSV NP and RAs-PCFV NP, suggesting that the bacterial expressed PYSV NP may be truncated at the N-terminal end. A smaller third protein less than 30 kDa can be detected by PYSV NP and PCFV NP, but not RAs-His, implying the cleavage at the C-terminal end containing the histidine tag of the expressed PYSV NP (Fig. 3b-d). This may be the reason that a lower reading mean (0.395±0.005) was obtained in indirect ELISA when RAs-PCFV NP reacted with the pET-expressed PYSV NP (Fig. 5b). Alternatively, the conformational folding of the PYSV NP may also affect ELISA results. In contrast to the bacterial-expressed NP, no degraded proteins were observed from the native PCFV NP (Fig. 2), suggesting that the bacterial-expressed NP shows less stability but does not affect its antigenicity in western blotting. The epitopes of the antisera are expected at the central regions of the NPs of PYSV and PCFV, which contain highly conserved sequences (Fig. 7). MAb-PCFV NP produced in this study specifically reacted with the homologous antigen, indicating that it can be used to distinguish PCFV from PYSV. The epitope of MAb-PCFV NP is deduced that locates at the N- or C-terminal region of the PCFV NP, where more diverse sequences between the NPs of PCFV and PYSV exist (Fig. 7).
RAs-PCFV NP, MAb-PCFV NP and RAs-PYSV NP did not react with the crude
antigens from plant tissues infected with other individual tospoviruses in indirect ELISA (Fig. 406 407 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430
5) and western blotting (data not shown), indicating that PCFV and PYSV are serologically distinct from other tospoviruses as previously predicted (Chen et al. 2010; Chu et al. 2001; Satyanarayana et al. 1998). Taken together, our results indicate that PCFV is closely related to PYSV phylogenetically and serologically, and these two tospoviruses should be classified in the same serogroup assigned by PYSV, since it was published prior to PCFV (Chen and Chiu 1996; Reddy et al. 1991). The antisera can be used to detect PCFV and PYSV in crops, and MAb-PCFV NP is a useful tool for differentiation of PCFV from PYSV.
Acknowledgements
We thank the funding from the Bureau of Animal and Plant Health Inspection and Quarantine, Council of Agriculture, Executive Yuan [101AS-10.3.1-BQ-B1(3)], the National Science Council (NSC-101-2911-I-005-301 and NSC-102-2911-I-005-301), and the Ministry of Education, Taiwan, R.O.C. under the ATU plan.
References
Chen, C. C., Chen, T. C., Lin, Y. H., Yeh, S. D., & Hsu, H. T. (2005a). A chlorotic spot
disease on calla lilies (Zantedeschia spp.) is caused by a tospovirus serologically but
distantly related to Watermelon silver mottle virus. Plant Disease, 89, 440-445. Chen, C. C., & Chiu, R. J. (1996). A tospovirus infecting peanut in Taiwan. Acta
Horticulturae, 431, 57-67.
Chen, T. C., Hsu, H. T., Jain, R. K., Huang, C. W., Lin, C. H., Liu, F. L., & Yeh, S. D. 431 432 433 434 435 436 437 438 439 440 441 442 443 444 445 446 447 448 449 450 451 452 453 454 455
(2005b). Purification and serological analyses of tospoviral nucleocapsid proteins
expressed by Zucchini yellow mosaic virus vector in squash. Journal of Virological
Methods, 129, 113-124.
Chen, T. C., Huang, C. W., Kuo, Y. W., Liu, F. L., Hsuan Yuan, C. H., Hsu, H. T., & Yeh, S.
D. (2006). Identification of common epitopes on a conserved region of NSs proteins
among tospoviruses of Watermelon silver mottle virus serogroup. Phytopathology, 96,
1296-1304.
Chen, T. C., Lu, Y. Y., Cheng, Y. H., Chang, C. A., & Yeh, S. D. (2008). Melon yellow spot
virus in watermelon: a first record from Taiwan. Plant Pathology, 57, 765.
Chen, T. C., Lu, Y. Y., Cheng, Y. H., Li, J. T., Yeh, Y. C., Kang, Y. C., Chang, C. P.,
Huang, L. H., Peng, J. C., & Yeh, S. D. (2010). Serological relationship between
Melon yellow spot virus and Watermelon silver mottle virus and differential detection
of the two viruses in cucurbits. Archives of Virology, 155, 1085-1095.
Chen, T. C., Lu, Y. Y., Kang, Y. C., Li, J. T., Yeh, Y. C., Kormelink, R., & Yeh, S. D.
(2011). Detection of eight different Tospovirus species by a monoclonal antibody
against the common epitope of NSs protein. Acta Horticulturae, 901, 61-66.
Chu, F. H., Chao, C. H., Peng, Y. C., Lin, S. S., Chen, C. C., & Yeh, S. D. (2001). Serological
and molecular characterization of Peanut chlorotic fan-spot virus, a new species of the
genus Tospovirus. Phytopathology, 91, 856-863.
Ciuffo, M., Kurowski, C., Vivoda, E., Copes, B., Masenga, V., Falk, B. W., & Turina, M. 456 457 458 459 460 461 462 463 464 465 466 467 468 469 470 471 472 473 474 475
(2009). A new Tospovirus sp. in cucurbit crops in Mexico. Plant Disease, 93,
467-474.
Ciuffo, M., Tavella, L., Pacifico, D., Masenga, V., & Turina, M. (2008). A member of a new
Tospovirus species isolated in Italy from wild buckwheat (Polygonum convolvulus). Archives of Virology, 153, 2059-2068.
Cortes, I., Livieratos, I. C., Derks, A., Peters, D., & Kormelink, R. (1998). Molecular and
serological characterization of Iris yellow spot virus, a new and distinct tospovirus
species. Phytopathology, 88, 1276-1282.
Dayhoff, M. O., Schwartz, R. M., & Orcutt, B. C. (1979). In Atlas of protein sequence and
structure, Vol. 5, pp. 345-352. Ed M.O. Dayhoff. National Biomedical Research
Foundation: Silver Spring.
de Avila, A. C., de Haan, P., Kormelink, R., Resende, R.O., Goldbach, R. W., & Peters, D.
(1993). Classification of tospoviruses based on phylogeny of nucleoprotein gene
sequences. Journal of General Virology, 74, 153-159.
de Ávila, A. C., Huguenot, C., Resende, R. O., Kitajima, E. W., Goldbach, R. W., & Peters,
D. (1990). Serological differentiation of 20 isolates of tomato spotted wilt virus.
Journal of General Virology, 71, 2801-2807.
de Haan, P., Kormelink, R., de Oliveira Resende, R., van Poelwijk, F., Peters, D., &
Goldbach, R. (1991). Tomato spotted wilt virus L RNA encodes a putative RNA
polymerase. Journal of General Virology, 72, 2207-2216.
de Haan, P., Wagemakers, L., Peters, D., & Goldbach, R. (1990). The S RNA segment of 476 477 478 479 480 481 482 483 484 485 486 487 488 489 490 491 492 493 494 495 496
tomato spotted wilt virus has an ambisense character. Journal of General Virology, 71,
1001-1007.
de Oliveira, A. S., Bertran, A. G., Inoue-Nagata, A. K., Nagata, T., Kitajima, E. W., & Oliveira Resende, R. (2011). An RNA-dependent RNA polymerase gene of a distinct Brazilian tospovirus. Virus Genes, 43, 385-389.
de Oliveira, A. S., Melo, F. L., Inoue-Nagata, A. K., Nagata, T., Kitajima, E. W., & Oliveira
Resende, R. (2012). Characterization of Bean necrotic mosaic virus: A member of a
novel evolutionary lineage within the genus Tospovirus. PLoS ONE, 7, e38634.
Felsenstein, J. (1989). PHYLIP – Phylogeny Inference Package (Version 3.2). Cladistics, 5, 164-166.
Goldbach, R., & Kuo, G. (1996). Introduction. Acta Horticulturae, 431, 21-26.
Goldbach, R., & Peters, D. (1996). Molecular and biological aspects of tospoviruses. In The
Bunyaviridae, pp. 129-157. Ed R.M. Elliott. New York, NY: Plenum Press.
Hassani-Mehraban, A., Saaijer, J., Peters, D., Goldbach, R., & Kormelink, R. (2005). A new
tomato-infecting tospovirus from Iran. Phytopathology, 95, 852-858.
Heinze, C., Roggero, P., Sohn, M., Vaira, A. M., Masenga, V., & Adam, G. (2000).
Peptide-derived broad-reacting antisera against tospovirus NSs-protein. Journal of Virological
Methods, 89, 137-146.
Hsu, H. T., Aebig, J., Rochow, W. F. (1984). Differences among monoclonal antibodies to
Barley yellow dwarf viruses. Phytopathology, 74, 600-605.
497 498 499 500 501 502 503 504 505 506 507 508 509 510 511 512 513 514 515 516
Hsu, H. T., Ueng, P. P., Chu, F. H., Ye, Z., & Yeh, S. D. (2000). Serological and molecular
characterization of a high temperature-recovered virus belonging to Tospovirus
serogroup IV. Journal of General Plant Pathology, 66, 167-175.
Jan, F. J., Chen, T. C., & Yeh, S. D. (2003). Occurrence, importance, taxonomy, and control
of thrips-borne tospoviruses. In Advances in Plant disease Management, pp. 399-421.
Eds H.C. Huang and S.N. Acharya. Trivandrum, India:
ResearchSignpost.
Kato, K., Hanada, K., & Kameya-Iwaki, M. (2000). Melon yellow spot virus: A distinct
species of the genus Tospovirus isolated from melon. Phytopathology, 90, 422-426. King, A. M. Q., Adams, M. J., Carstens, E. B., & Lefkowitz, E. J. (2011). Virus Taxonomy:
Ninth Report of the International Committeeon Taxonomy of Viruses. Amsterdam:
Elsevier Academic Press.
Kormelink, R., de Haan, P., Meurs, C., Peters, D., & Goldbach, R. (1992). The nucleotide
sequencw of the M RNA segments of Tomato spotted wilt virus, a bunyavirus with
two ambisense RNA segments. Journal of General Virology, 73, 2795-2804. Law, M. D., & Moyer, J. W. (1990). A tomato spotted wilt-like virus with a serologically
distinct N protein. Journal of General Virology, 71, 933-938.
Law, M. D., Speck, J., & Moyer, J. W. (1992). The M RNA of Impatiens necrotic spot
Tospovirus (Bunyaviridae) has an ambisense genomic organization. Virology, 188,
732-741. 517 518 519 520 521 522 523 524 525 526 527 528 529 530 531 532 533 534 535 536
Li, J. T., Yeh, Y. C., Yeh, S. D., Raja, J. A. J., Rajagopalan, P. A., Liu, L. Y., & Chen, T. C.
(2011). Complete genomic sequence of watermelon bud necrosis virus. Archives of
Virology, 156, 359-362.
Lin, Y. H., Chen, T. C., Hsu, H. T., Liu, F. L., Chu, F. H., Chen, C. C., Lin, Y. Z., & Yeh, S.
D. (2005). Serological comparison and molecular characterization for verification of
Calla lily chlorotic spot virus as a new tospovirus species belonging to Watermelon
silver mottle virus serogroup. Phytopathology, 95, 1482-1488.
Mohamed, N. A. (1981). Isolation and characterization of subviral structures from Tomato
spotted wilt virus. Journal of General Virology, 53, 197-208.
Mohamed, N. A., Randles, J. W., & Francki, R. I. B. (1973). Protein composition of Tomato
spotted wilt virus. Virology, 56, 12-21.
Pang, S. Z., Slightom, J. L., & Gonsalves, D. (1993). The biological properties of a distinct
tospovirus and sequence analysis of its S RNA. Phytopathology, 83, 728-733.
Pappu, H. R., Jones, R. A. C., & Jain, R. K. (2009). Global status of tospovirus epidemics in
diverse cropping systems: Successes achieved and challenges ahead. Virus Research,
141, 219-236.
Peng, J. C., Yeh, S. D., Huang, L. H., Li, J. T., Cheng, Y. F., & Chen, T. C. (2011). Emerging
threat of thrips-borne Melon yellow spot virus on melon and watermelon in Taiwan.
European Journal of Plant Pathology, 130, 205-214.
Reddy, D. V. R., Sudarshana, M. R., Ratna, A. S., Reddy, A. S., Amin, P. W., Kumar, I. K., & 537 538 539 540 541 542 543 544 545 546 547 548 549 550 551 552 553 554 555 556
Murthy, A. K. (1991). The occurrence of yellow spot virus, a member of tomato
spotted wilt virus group, on peanut (Arachis hypogaea L) in India. In
Virus-Thrips-Plant interactions of tomato spotted wilt virus, pp. 77-88. Eds H.T. Hsu and R.H.
Lawson. Proceedings of the USDA Agricultural Research Service, ARS-87.
Saitou, N., & Nei, M. (1987). The neighbor-joining method: a new method for reconstructing
phylogenetic trees. Molecular Biology and Evolution, 4, 406-425.
Satyanarayana, T., Gowda, S., Reddy, K. L., Mitchell, S. E., Dawson, W. O., & Reddy, D. V.
R. (1998). Peanut yellow spot virus is a member of a new serogroup of Tospovirus
genus based on small (S) RNA sequence and organization. Archives of Virology, 143,
353-364.
Seepiban, C., Gajanandana, O., Attathom, T., & Attathom, S. (2011). Tomato necrotic
ringspot virus, a new tospovirus isolated in Thailand. Archives of Virology, 156,
263-274.
van Knippenberg, I., Goldbach, R., & Peters, D. (2002). Purified Tomato spotted wilt virus
particles support both genome replication and transcription in vitro. Virology, 303,
278-286.
Yeh, S. D., Chao, C. H., Cheng, Y. H., & Chen, C. C. (1996). Serological comparison of four
distinct tospoviruses by polyclonal antibodies to purified nucleocapsid proteins. Acta
Horticulturae, 431, 122-134.
Yeh, S. D., Cheng, Y. H., Jih, C. L., Chen, C. C., & Chen, M. J. (1988). Identification of 557 558 559 560 561 562 563 564 565 566 567 568 569 570 571 572 573 574 575 576
tomato spotted infecting horn melon and watermelon. Plant Protection Bulletin, 30,
319-320.
Yeh, S. D., & Gonsalves, D. (1984). Purification and immunological analysis of
cylindrical-inclusion protein induced by Papaya ringspot virus and Watermelon mosaic virus I.
Phytopathology, 74, 1273-1278.
Zhou, J., Kantartzi, S. K., Wen, R. H., Newman, M., Hajimorad, M. R., Rupe, J. C., &
Tzanetakis, I. E. (2011). Molecular characterization of a new tospovirus infecting
soybean. Virus Genes, 43, 289-295. 577 578 579 580 581 582 583 584
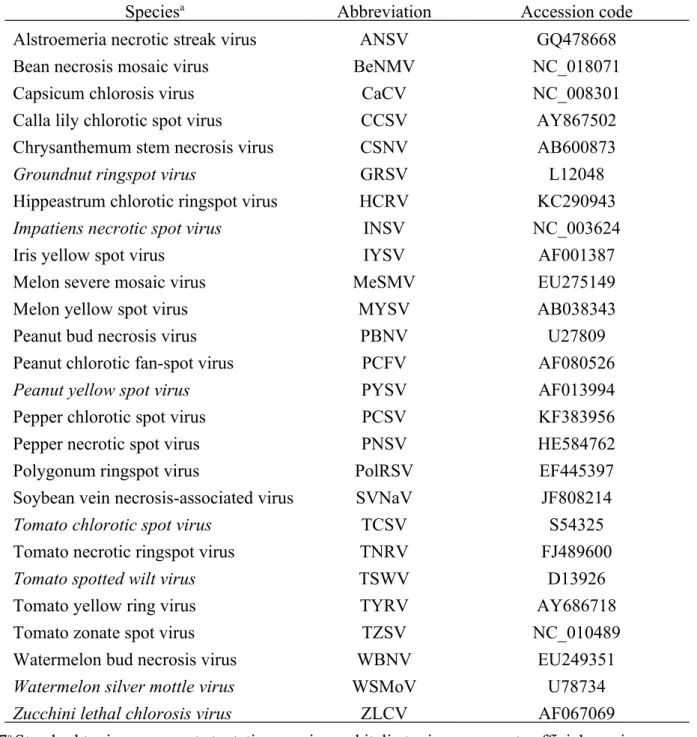
Table 1. Accession codes of the small (S) RNA or nucleocapsid (N) gene sequences of
tospoviruses analyzed in this study
Speciesa Abbreviation Accession code
Alstroemeria necrotic streak virus ANSV GQ478668
Bean necrosis mosaic virus BeNMV NC_018071
Capsicum chlorosis virus CaCV NC_008301
Calla lily chlorotic spot virus CCSV AY867502
Chrysanthemum stem necrosis virus CSNV AB600873
Groundnut ringspot virus GRSV L12048
Hippeastrum chlorotic ringspot virus HCRV KC290943
Impatiens necrotic spot virus INSV NC_003624
Iris yellow spot virus IYSV AF001387
Melon severe mosaic virus MeSMV EU275149
Melon yellow spot virus MYSV AB038343
Peanut bud necrosis virus PBNV U27809
Peanut chlorotic fan-spot virus PCFV AF080526
Peanut yellow spot virus PYSV AF013994
Pepper chlorotic spot virus PCSV KF383956
Pepper necrotic spot virus PNSV HE584762
Polygonum ringspot virus PolRSV EF445397
Soybean vein necrosis-associated virus SVNaV JF808214
Tomato chlorotic spot virus TCSV S54325
Tomato necrotic ringspot virus TNRV FJ489600
Tomato spotted wilt virus TSWV D13926
Tomato yellow ring virus TYRV AY686718
Tomato zonate spot virus TZSV NC_010489
Watermelon bud necrosis virus WBNV EU249351
Watermelon silver mottle virus WSMoV U78734
Zucchini lethal chlorosis virus ZLCV AF067069
a Standard typing represents tentative species and italic typing represents official species.
585
586
587 588 589
Figure legends
Fig. 1 Titration of the antisera and monoclonal antibody produced in this study. Fifty-fold
diluted crude extracts of the healthy (H) and Peanut chlorotic fan-spot virus (PCFV)-infected
Nicotiana benthamiana plants were used to react with the rabbit antiserum (RAs-PCFV NP)
(a) and mice monoclonal antibody MAb-PCFV NP (b) against the nucleocapsid protein (NP)
of PCFV. One hundred nanogram of the purified NP of Peanut yellow spot virus (PYSV) expressed by bacteria was used to react with the rabbit antiserum (RAs-PYSV NP) (c) to PYSV NP. The dilutions of antisera and ascitic fluid used for titration assays are indicated.
Fig. 2 Serological reactions of the nucleocapsid protein (NP) of Peanut chlorotic fan-spot
virus (PCFV) with the produced rabbit antisera and mice monoclonal antibody. The leaf extract of a healthy (H) N. benthamiana plant was used as the negative control. The leaf crude extract (CE) of a PCFV-infected Nicotiana benthamiana plant (1/50 dilution) and the NP purified from Chenopodium quinoa plants (NP, 2 g) were analyzed after separation by 12% SDS-PAGE and stained by Coomassie blue (a). Rabbit antiserum against the PCFV NP (RAs-PCFV NP) (b), mice monoclonal antibody against the PCFV NP (MAb-PCFV NP) (c), and rabbit antiserum against the bacteria-expressed PYSV NP (RAs-PYSV NP) (d) were used to analyze the serological reactions.
Fig. 3 Serological reactions of the bacterial-expressed nucleocapsid protein (NP) of Peanut
yellow spot virus (PYSV) with the produced rabbit antisera in western blotting. Total protein
of the IPTG-induced bacterial culture containing the empty vector pET-28b(+) (EV) was used as a control. Total protein (TP) of the bacterial lysate (5 l of 1 ml IPTG-induced culture) containing pET-expressed PYSV NP and the eluted protein (EP) (2 g) obtained from Ni2+
-590 591 592 593 594 595 596 597 598 599 600 601 602 603 604 605 606 607 608 609 610 611 612 613 614
NTA affinity chromatography were separated by 12% SDS-PAGE and stained by Coomassie blue (a). The rabbit antiserum against the histidine tag (RAs-His) was used to confirm the expression of PYSV NP (b). The rabbit antisera to the bacteria-expressed PYSV NP (RAs-PYSV NP) (c) and to the PCFV NP (RAs-PCFV NP) (d) were used for assay.
Fig. 4 Serological reactions of Peanut yellow spot virus (PYSV) with the produced antisera
and monoclonal antibody in western blotting. The crude saps extracted from the frozen leaves of chili plants infected with PYSV (1/50 dilution, sample number 1-4), which were collected in Yunnan, China (the SP-C isolate is assigned as sample 1, GenBank accession code: HQ402596), were used to incubate with the individual rabbit antisera against the bacteria-expressed nucleocapsid protein (NP) of PYSV (RAs-PYSV NP) (a) and the NP of Peanut chlorotic fan-spot virus (PCFV) (RAs-PCFV NP), (b) and the mice monoclonal antibody against the PCFV NP (MAb-PCFV NP) (c). The crude extracts from leaves of healthy
Chenopodium quinoa (Cq) and pepper (Pep) plants were used as the negative controls. The
eluted pET-expressed PYSV NP (EP) and the crude extract of PCFV-infected C. quinoa were used as the positive controls of corresponding antibodies.
Fig. 5 Serological reactions of the produced rabbit antisera and mice monoclonal antibody
(MAb) with crude nucleocapsid protein (NP) antigens of different tospoviruses, analyzed by indirect enzyme-linked immunosorbent assay. RAs-PYSV NP (a) against the NP of Peanut
yellow spot virus (PYSV), and RAs-PCFV NP (b) and MAb-PCFV NP (c) against the NP of
Peanut chlorotic fan-spot virus (PCFV) were used to react with leaf extracts of Nicotiana
benthamiana plants infected with individual tospoviruses, including PCFV, Impatiens necrotic spot virus (INSV), Tomato spotted wilt virus (TSWV), Groundnut ringspot virus
(GRSV), Tomato chlorotic spot virus (TCSV), Watermelon silver mottle virus (WSMoV), 615 616 617 618 619 620 621 622 623 624 625 626 627 628 629 630 631 632 633 634 635 636 637 638 639
Peanut bud necrosis virus (PBNV), Watermelon bud necrosis virus (WBNV), Capsicum
chlorosis virus (CaCV), Calla lily chlorotic spot virus (CCSV), Melon yellow spot virus (MYSV), Iris yellow spot virus (IYSV) and Tomato yellow ring virus (TYRV). The bacteria-expressed NP of PYSV (100 ng) was also used for assay. The non-inoculated healthy N.
benthamiana plant (H) was used as the negative control.
Fig. 6 Phylogenetic tree of tospoviruses based on the amino acid sequence of the
nucleocapsid protein encoded by S RNA. The dendropraph was produced using the Neghbour-Joining algorithm based on 1000 bootstrap replicates. The tospovirus species, which are potentially serological-related based on experimental results and literatures, are circularized. See Table 1 for full virus names of the acronyms.
Fig. 7 Alignment of both nucleocapsid proteins of Peanut yellow spot virus (PYSV) and
Peanut chlorotic fan-spot virus (PCFV). Sequence comparison was performed by BL2SEQ program of Biology Workbench, San Diego Supercomputer Center (SDSC)
(http://workbench.sdsc.edu/). Sequences were obtained from GenBank under the following accession codes: PYSV, AF013994 and PCFV, AF080526. The identical nucleotides are highlighted by black boxes.
640 641 642 643 644 645 646 647 648 649 650 651 652 653 654 655 656 657 658