行政院國家科學委員會專題研究計畫 成果報告
巨噬細胞在子宮腺肌症致病機轉中之角色
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-002-159- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院婦產科 計畫主持人: 楊政憲 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 10 月 21 日
中英文摘要 背景:曾有研究顯示子宮腺肌症的子宮內膜存在較多的巨噬細胞,因此巨噬細胞可能在子 宮腺肌症的致病過程中扮演重要角色。本研究嘗試探討在巨噬細胞作用下,子宮腺肌症子 宮的內膜基質細胞(ESCs)可能產生之反應。 方法:收集 10 位罹患子宮腺肌症婦女(實驗組)及 11 位罹患子宮肌瘤婦女(對照組)的 子宮內膜,將其分離成單一 ESC 後,與巨噬細胞共同培養 24 小時。 結果:免疫組織染色顯示子宮內膜中有 IL-6、IL-8、IL-10 蛋白質的存在。利用即時反轉錄 聚合酵素鏈鎖反應(real-time RT-PCR)發現 IL-6 mRNA 在子宮腺肌症的 ESCs 與巨噬細胞 共同培養後有明顯增高的表現量,但 IL-8 及 IL-10 mRNA 的表現量則在實驗組與對照組之 間無差異。
結論:子宮腺肌症的 ESCs 與巨噬細胞共同培養後有明顯增高的 IL-6 mRNA 表現量。此異 常表現可能在在子宮腺肌症的致病機轉中扮演重要角色。
BACKGROUND: An increased number of macrophages have been found in eutopic
endometrium of adenomyosis, suggesting possible involvement of macrophages in the development of adenomyosis. In this study, we tried to determine the effects of macrophage on endometrial stromal cells (ESCs) in women with adenomyosis.
METHODS: Eutopic endometrium was obtained and separated into single ESC in 10 women
with adenomyosis (study group) and 11 without adenomyosis (control group). ESCs were then cultured alone or with macrophage for 24h.
RESULTS: Immunohistochemistry identified the presence of IL-6, IL-8, and IL-10 in ESCs.
Quantitative real-time RT-PCR revealed that the IL-6 mRNA was significantly expressed in macrophage-cocultured ESCs in adenomyosis than that in the controls, but was not different in ESCs cultured alone between the two groups. The levels of IL-8 and IL-10 mRNA were similar in ESCs either cultured alone or with macrophage between women with and without adenomyosis.
CONCLUSIONS: IL-6 mRNA was significantly expressed in ESCs after in vitro coculture with
macrophage in adenomyosis. This aberrant behavior of ESCs might play a role in the formation of ectopic endometrial implants in adenomyosis.
報告內容
前言
Adenomyosis refers to the presence of endometrial glands and stroma that is haphazardly deep within the myometrium. Our previous study found that the expression of killer cell immunoglobulin (Ig)-like receptors (KIRs) on natural killer (NK) cells was decreased in eutopic endometrium in women with adenomyosis.1 It may be a compensatory effect in which the NK cytotoxicity is activated in order to eradicate the abnormal endometrial cells that might exit of the eutopic site of endometrium, suggesting that abnormal endometrial cells, but not aberrant immunological phenomenon, possibly account for the development of adenomyosis.
An increased number of macrophages has been found in eutopic and ectopic endometrium in adenomyosis,2,3 implying possible involvement of macrophage in the development of adenomyosis. In endometriosis, which is generally considered to have a similar pathophysiology with adenomyosis, blood monocytes were found to enhance autologous endometrial cell proliferation.4 Accordingly, macrophages appeared to play an important role in the pathogenesis of adenomyosis.5
Cytokines are protein mediators of extra- and intracellular communications within the immune system. A number of cytokines and growth factors are synthesized in the endometrium in response to sex steroid hormones and act to regulate endometrial function.6 Using an in vitro coculture system, macrophages have been shown to activate cytokine expression in tumor cells and thus enhance tumor proliferation.7 Similarly,
cytokines may also contribute to the proliferation of ectopic endometrial implants in adenomyosis after the activation by macrophage.
研究目的
In this study, endometrial stromal cells (ESCs) were obtained from women with and without adenomyosis, and were cultured alone or with macrophage. We investigated messenger RNA expression of interleukin-6 (IL-6), interleukin-8 (IL-8) and interleukin-10 (IL-10) in ESCs, in an attempt to elucidate the possible role of macrophage in the pathogenesis of adenomyosis.
研究方法
Subjects and specimens
This study consisted of 10 women who suffered from adenomyosis (study group) and 11 women in whom leiomyoma was found (control group). These women underwent hysterectomy, either via abdominal or vaginal route, at our hospital due to intolerable symptoms, e.g. low abdominal pain, dysmenorrhea, hypermenorrhea, etc. All the participating women were at pre-menopausal status. Since endometrial cells might have different behaviors between follicular and secretory phases and that an increased number of macrophage has been found in the late-secretory phase,8,9 only women at early- to mid-secretory phases were included in this study. The diagnosis of adenomyosis was made by histopathologic examination without exception. Informed consent was obtained from each woman before surgery, and this study protocol was approved by the institutional review board (IRB) at our hospital.
Purification of ESCs
Endometrium was obtained immediately after the uterus was removed away from the women, and was placed immediately in an ice-cold 1:1 mixture of Dulbecco’s Modified Eagle’s Medium and Ham’s F-12 (DMEM/F-12) for transport to the laboratory. The tissue was gently dissected into small pieces (1 to 2 mm3) and washed by centrifugation (400 x g) in fresh medium to remove any debris or excess blood cells. The tissue was then incubated for 2 hours at 37°C in a shaking water bath in DMEM/F-12 containing 0.5% collagenase and 0.02% DNAase. The dispersed cells were filtered through a 70-µm nylon mesh to remove the undigested tissue pieces containing the glandular epithelium.
The filtered fraction was separated further from epithelial cell clumps by differential sedimentation at unit gravity, as follows. The cells were resuspended in 2 mL of culture medium and layered slowly over 10 mL of the medium in a centrifuge tube. Sealed tubes were placed in an upright position at 37°C in 5% CO2 in air for 30 minutes. After
sedimentation, the top 8mL of medium was collected. The medium containing stromal cells was filtered through a 40-µm nylon mesh. Final purification was achieved by allowing stromal cells to adhere selectively to culture dishes for 30 minutes at 37°C in 5% CO2 in
air. Nonadhering epithelial cells were removed.
The stromal cells were plated in plastic flasks (75 cm2) containing DMEM/F-12 supplemented with 100 IU/mL penicillin G, 50 mg/mL of streptomycin, 2.5 µg/mL of amphotericin B, and 10% fetal bovine serum (FBS, vol/vol) at 37°C in 5% CO2 in air, and
allowed to replicate to confluence. Thereafter,
stromal cells were passed by standard methods of trypsinization and were plated in 6-well plates. Cells became preconfluent (covering 80% of the culture well) in approximately 5-7 days.
Coculture of ESCs with macrophage
The human monocyte cell line THP-1 (ATCC TIB202; ATCC, Manassas, VA) was stimulated to differentiate into macrophage by the addition of 1.6 x 10-7 M phorbol myristate acetate (Sigma Chemical, St. Louis, MO) for 48 hours. All cells were incubated at 37°C in 5% CO2 in air. Cultured macrophages were
detached by exposure to versene (1:5000 dilution; Life Technologies, Paisley, UK), washed with 15 ml of PBS for three times to avoid any effect of phorbol myristate acetate, then incubated in 20 ml of serum-free media for 24 hours.
The ESCs, seeded in 6-well plates, were allowed to grow to confluence for 24 hours and then washed twice with serum-free RPMI 1640 and incubated in 2 mL of serum-free RPMI. Macrophages (5 x 105 in 2 mL of serum-free RPMI) were seeded into the top chamber of a transwell apparatus (Costar, Cambridge, MA), which was then placed directly on top of the 6-well plates containing ESCs. Two mL of serum-free RPMI without macrophage was added to another confluent well of ESCs.
After incubation for 24 hours, the ESCs were washed twice with PBS and collected. Total RNAs were extracted using 1 mL of Trizol (Invitrogen, Carlsbad, CA).
Real-time quantitative PCR
A real-time quantitative PCR system (ABI PRISM 7700 Sequence Detection System; PE Applied Biosystems) was used as
previously described (10, 11). Briefly, RNA was prepared and subsequently cDNA was obtained by random hexamer priming. The primers and probes used in the PCR were designed according to the TaqMan primer and probe design system. Based on the manufacturer's protocol, the FAM (6-carboxyfluorescein) and VIC were used as the reporter dyes and TAMRA (6-carboxy-tetramethyl-rhodamine) as the quencher dye. The probes were labeled with both reporter dye and quencher dye on the 5’- and 3’-ends, respectively. During PCR, the reporter dye was released and the resultant fluorescence was detected and could be quantified. The PCR was carried out in a thermal cycler (ABI PRISM 7700 Sequence Detection System) in the following sequence: reaction at 50°C for 2 min, at 95°C for 10 min, and subsequently the PCR was repeated for 45 cycles at denaturation at 95°C for 15 s and annealing and extension at 60°C for 1 min. Each sample was examined in triplicate and a mean value was obtained. The relative concentration of each mRNA was subsequently calculated according to the Manufacturer's User Manual. Briefly, the threshold cycle (CT) values of the target gene
(IL-6, IL-8 and IL-10) and the internal control gene (β-actin) mRNA in the studied sample were first measured. The ∆CT value of the
studied sample was calculated by the following formula: ∆CT = CT of target gene -
CT of β-actin (this ∆CT is designated as
“sample ∆CT”). Similarly, the CT values of the
target gene and its respective β-actin of a positive control were also obtained and the ∆CT was calculated (designated as “calibrator
∆CT”). ∆∆CT was then calculated using the
following formula: ∆∆CT = ∆CT (sample) -
∆CT (calibrator) and finally the relative value
of each mRNA was calculated by the formula: 2-∆∆CT. PCR without template was used as a negative control (called no template control) to verify experimental results.
Immunohistochemistry
The immunohistochemistry protocol has been described previously.10,11 Briefly, formalin-fixed, paraffin-embedded tissue sections were deparaffinized with xylene, and hydrated with step-down concentrations of ethanol. The sections were then incubated with 30% H2O2/70% methanol solution at RT
for 5 min, treated with normal horse serum (Pierce Biotechnology, Rockford, IL) at RT for 5 min to reduce non-specific binding of antibodies, and incubated overnight at 4°C with primary antibodies (for IL-6 detection, mouse monoclonal anti-human IL-6 antibody, 1:50 dilution; Chemicon, Temecula, CA; for IL-8 detection, goat polyclonal anti-human IL-8 antibody, 1:50 dilution; R&D; for IL-10 detection, mouse monoclonal anti-human IL-10 antibody, 1:50 dilution; R&D). On the next day, the sections were washed with PBS, treated with polyvalent biotinylated secondary antibody (Dako, Carpinteria, CA), which bound to primary antibody at RT for 10 min, and then treated with streptavidin-peroxidase reagent, which bound to the secondary antibody, at RT for 10 min. The color was developed using 3,3’ diaminobenzidine (DAB) chromogen at RT for 15 min and counterstained with Mayer’s hematoxylin for 1 min. Experiments with the omission of primary antibodies were used as negative controls.
Statistical analysis
All values are expressed as mean ± standard error (SEM). As the data were not
normally distributed, the Mann-Whitney U test for non-parametric data was used to compare the difference between the groups. A probability value less than 0.05 was considered statistically significant.
結果與討論
The mean age was 41.3 years for women with adenomyosis and 43.8 for those without adenomyosis. The body mass indices were similar between the two groups (mean = 24.7 vs. 23.8). All the participating women were at early- to mid-secretory phases.
Immunohistochemical study for IL-6, IL-8, and IL-10 proteins in ESCs
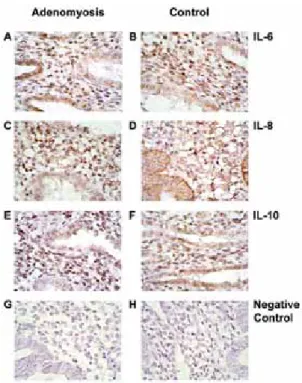
Immunohistochemical study identified that the immunoreactive IL-6, IL-8, and IL-10 were found in ESCs. The staining was also detectable in the glandular epithelium. As expected, the negative controls did not show any immunoreactive staining (Figure 1).
Quantitation of IL-6, IL-8 and IL-10 mRNA by real-time quantitative PCR
In ESCs cultured alone for 24 hours, the relative levels of IL-6, IL-8 and IL-10 mRNA, using β-actin as a reference gene, were 0.5 ± 0.2, 6.4 ± 2.0, and 0.2 ± 0.1 (mean ± SEM) in women with adenomyosis, similar to those (0.5 ± 0.2, 3.2 ± 1.1 and 0.05 ± 0.04) in women without adenomyosis. In ESCs cultured with macrophage for 24 hours, however, the relative level of IL-6 mRNA was much higher in women with than that in women without adenomyosis (2.2 ± 0.8 vs. 0.3 ± 0.1, P = 0.006). The relative levels of IL-8 (24.6 ± 6.9 vs. 16.9 ± 4.4) and IL-10 (0.07 ± 0.04 vs. 0.05 ± 0.04) mRNA remained similar between women with and without adenomyosis in ESCs cultured with
macrophage (Figure 2).
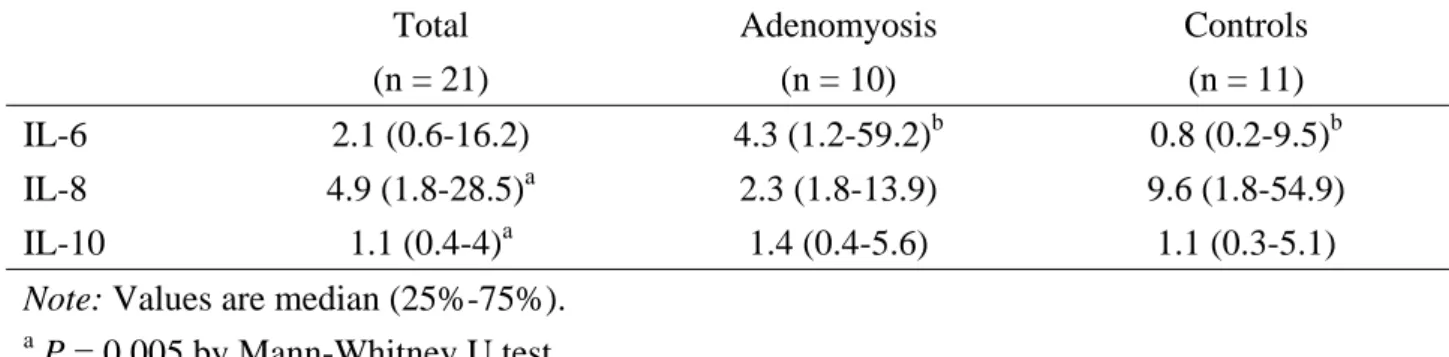
Without regard to women suffering from adenomyosis or not, the IL-8 mRNA expression in ESCs cultured with macrophage was 4.9 times higher than that in ESCs cultured alone. This magnification was significantly higher than that (1.1 times) of IL-10 mRNA (P = 0.005). On the other hand, the magnification of IL-6 mRNA expression in women with adenomyosis had a higher value than that in women without adenomyosis (4.3 vs. 0.8, P = 0.041), but it was not different between women with and without adenomyosis in terms of IL-8 (2.3 vs. 9.6) and IL-10 (1.4 vs. 1.1) mRNA expression (Table I).
The etiology and pathologic mechanism responsible for adenomyosis are not yet well known by far. Our results, using a coculture system in which the cells were physically separated, revealed an increased IL-6 mRNA expression in ESCs after culture with macrophage in women with adenomyosis. Although macrophages are able to mediate the cytolysis of eutopic and ectopic endometrial cells,12,13 they could also enhance cell growth and proliferation through the amplification of cytokine secretion.7,14 In this study, macrophage appeared to play an important role in the regulation of IL-6 expression in ESCs. This IL-6 mRNA expression might be enhanced by some kinds of factors in the conditioned medium that was secreted after macrophage-ESC communication. Although it is unknown how many and which factors are involved in this regulatory process, TNF-α is a possible one that has been found to be secreted by macrophage and induce the proliferation of ESCs.15,16
line THP-1 was stimulated to differentiate into macrophage, and was then cocultured with ESCs. The reason for using THP-1 instead of endometrial macrophage was that the isolation of tissue macrophage is not technically available by far. Tissue macrophages are reported to be phenotypically heterogeneous in different tissues.17,18 Although peripheral blood monocyte and peritoneal macrophage have been employed in in vitro cytolysis of endometrial cells,12,13 their biological behaviors are not fully consistent with that of endometrial macrophage. Therefore, THP-1 served as a surrogate of endometrial macrophage in this in vitro experiment.
IL-6 contains angiogenic and mitogenic effects and plays a pivotal role in mediating numerous physiologic and pathogenic processes.19 The production of IL-6 may be modulated by several factors, such as ovarian steroids and inflammation-associated cytokines including TNF-α, IL-1β, and interferon-γ.20,21
It has been suggested that IL-6 has important functions in reproductive physiology, including the regulation of ovarian steroid production, folliculogenesis, and early events related to implantation.22,23 Both eutopic and ectopic endometrium were proved to produce IL-6.13,24 It is consistent with that found in the present study, in which immunohistochemistry demonstrated the presence of IL-6 in ESCs.
In this study, IL-8 mRNA expression in ESCs was increased by 4.9-fold after interaction with macrophage, which was much higher than that of IL-10 (1.1-fold). This marked expression of IL-8 mRNA after the activation of macrophage was also reported in lung cancer cells, in which a 270-fold increase was found.7 Although IL-8 has been shown to enhance the proliferation of eutopic and
ectopic ESCs in endometriosis,17,25 we did not found difference in the expression of IL-8 mRNA in ESCs, either cultured alone or with macrophage, between women with and without adenomyosis. Therefore, IL-8 did not appear to play an important role in the pathogenesis of adenomyosis, which is different from that found in endometriosis.25,26 Similarly, IL-10 was not differently expressed at RNA level in ESCs between adenomyosis and controls in this study, although it has been reported to have an increased production in peritoneal fluid in endometriosis.27,28
There is only limited literature investigating the angiogenic and growth factors that might encourage adenomyosis. Besides IL-6 noted in this study, vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) are two other possible factors accounting for the growth of adenomyosis.29 The array technology might be a good tool to explore more angiogenic and growth factors in adenomyosis. It deserves further investigation.
Dilemma exists in the clinical management of adenomyosis, i.e., between the effectiveness of treatment and the preservation of fertility. Hysterectomy is the most common and effective treatment for adenomyosis as it nearly ensures cure. However, desire for a future pregnancy contraindicates hysterectomy. On the contrary, drug therapy including oral contraceptive pill, progestogen, danazol, and gonadotropin releasing hormone analogue (GnRHa) preserves female fertility, but is relatively ineffective in treating adenomyosis. Our study provides an idea in the possible prevention of adenomyosis. For women at the risk of development of adenomyosis, such as prior uterine surgery30 and prior endometrial curettage or resection,
31,32
the future occurrence of adenomyosis might be potentially prevented through the prophylactic management of their eutopic endometrium. Although the precise manner of prevention remains unclear, it deserves further investigation.
Our results appeared to suggest that IL-6 mRNA was significantly expressed in ESCs after in vitro coculture with macrophage in adenomyosis. This aberrant behavior of ESCs might play a role in the formation of ectopic endometrial implants in adenomyosis.
參考文獻
1. Yang JH, Chen MJ, Chen HF, Lee TH, Ho HN, Yang YS: Decreased expression of killer cell inhibitory receptors on natural killer cells in eutopic endometrium in women with adenomyosis. Hum Reprod 2004;19:1974-1978.
2. Ferenczy: A. Pathophysiology of adenomyosis. Hum Reprod Update 1998;4:312-322.
3. Ota H, Igarashi S, Hatazawa J, Tanaka T: Is adenomyosis an immune disease? Hum Reprod Update 1998;4:360-367. 4. Braun DP, Muriana A, Gebel H, Rotman
C, Rana N, Dmowski WP: Monocyte-mediated enhancement of endometrial cell proliferation in women with endometriosis. Fertil Steril 1994;61:78-84.
5. Wu MY, Ho HN: The role of cytokines in endometriosis. Am J Reprod Immunol 2003;49:285-296.
6. Zarmakoupis PN, Rier SE, Maroulis GB, Becker JL: Inhibition of human endometrial stromal cell proliferation by interleukin 6. Hum Reprod 1995;10:2395-2399.
7. Chen JJ, Yao PL, Yuan A, Hong TM, Shun CT, Kuo ML, Lee YC, Yang PC: Up-regulation of tumor interleukin-8 expression by infiltrating macrophages: its correlation with tumor angiogenesis and patient survival in non-small cell lung cancer. Clin Cancer Res 2003;9:729-737.
8. Braun DP, Ding J, Shen J, Rana N, Fernandez BB, Dmowski WP: Relationship between apoptosis and the number of macrophages in eutopic endometrium from women with and without endometriosis. Fertil Steril 2002;78:830-835.
9. Eidukaite A, Tamosiunas V: Endometrial and peritoneal macrophages: expression of activation and adhesion molecules. Am J Reprod Immunol 2004;52:113-117. 10. Chen HF, Lin CY, Chao KH, Wu MY,
Yang YS, Ho HN: Defective production of interleukin-11 by decidua and chorionic villi in human anembryonic pregnancy. J Clin Endocrinol Metab 2002;87:2320-2328.
11. Chen HF, Chao KH, Shew JY, Yang YS, Ho HN: Expression of leukemia inhibitory factor and its receptor is not altered in the decidua and chorionic villi of human anembryonic pregnancy. Hum Reprod 2004;19:1647-1654.
12. Braun DP, Gebel H, Rana N, Dmowski WP: Cytolysis of eutopic and ectopic endometrial cells by peripheral blood monocytes and peritoneal macrophages in women with endometriosis. Fertil Steril 1998;69:1103-1108.
13. Dmowski WP, Gebel H, Braun DP: Decreased apoptosis and sensitivity to macrophage mediated cytolysis of endometrial cells in endometriosis. Hum
Reprod Update 1998;4:696-701.
14. Leek RD, Lewis CE, Whitehouse R, Greenall M, Clarke J, Harris AL: Association of macrophage infiltration with angiogenesis and prognosis in invasive breast carcinoma. Cancer Res 1996;56:4625-4629.
15. Iwabe T, Harada T, Tsudo T, Nagano Y, Yoshida S, Tanikawa M, Terakawa N: Tumor necrosis factor-alpha promotes proliferation of endometriotic stromal cells by inducing interleukin-8 gene and protein expression. J Clin Endocrinol Metab 2000;85:824-829.
16. Tsudo T, Harada T, Iwabe T, Tanikawa M, Nagano Y, Ito M, Taniguchi F, Terakawa N: Altered gene expression and secretion of interleukin-6 in stromal cells derived from endometriotic tissues. Fertil Steril 2000;73:205-211.
17. Rutherford MS, Witsell A, Schook LB: Mechanisms generating functionally heterogeneous macrophages: chaos revisited. J Leukocyte Biol 1993;53:602-618.
18. Hunt JS, Robertson SA: Uterine macrophages and environmental programming for pregnancy success. J Reprod Immunol 1996;32:l-25.
19. Motro B, Itin A, Sachs L, Keshet E: Pattern of interleukin 6 gene expression in vivo suggests a role for this cytokine in angiogenesis. Proc Natl Acad Sci USA 1990;87:3092-3096.
20. Tabibzadeh SS, Santhanam U, Sehgal PB, May LT: Cytokine-induced production of IFN-beta 2/IL-6 by freshly explanted human endometrial stromal cells. Modulation by estradiol-17 beta. J Immunol 1989;142:3134-3139.
21. Akoum A, Lemay A, Paradis I, Rheault N,
Maheux R: Secretion of interleukin-6 by human endometriotic cells and regulation by proinflammatory cytokines and sex
steroids. Hum Reprod 1996;11:2269-2275.
22. Gorospe WC, Hughes FM Jr, Spangelo BL: Interleukin-6: effects on and production by rat granulosa cells in vitro. Endocrinology 1992;130:1750-1752. 23. Jacobs AL, Sehgal PB, Julian J, Carson
DD: Secretion and hormonal regulation of interleukin-6 production by mouse uterine stromal and polarized epithelial cells cultured in vitro. Endocrinology 1992;131:1037-1046.
24. Tseng JF, Ryan IP, Milam TD, Murai JT, Schriock ED, Landers DV, Taylor RN: Interleukin-6 secretion in vitro is up-regulated in ectopic and eutopic endometrial stromal cells from women with endometriosis. J Clin Endocrinol Metab 1996;81:1118-1122.
25. Arici A, Seli E, Senturk LM, Gutierrez LS, Oral E, Taylor HS: Interleukin-8 in the human endometrium. J Clin Endocrinol Metab 1998;83:1783-1787. 26. Iwabe T, Harada T, Tsudo T, Tanikawa M,
Onohara Y, Terakawa N: Pathogenetic significance of increased levels of interleukin-8 in the peritoneal fluid of patients with endometriosis. Fertil Steril 1998;69:924-930.
27. Ho HN, Wu MY, Chao KH, Chen CD, Chen SU, Yang YS: Peritoneal interleukin-10 increases with decrease in activated CD4+ T lymphocytes in women with endometriosis. Hum Reprod 1997;12:2528-2533.
28. Rana N, Braun DP, House R, Gebel H, Rotman C, Dmowski WP: Basal and stimulated secretion of cytokines by
peritoneal macrophages in women with endometriosis. Fertil Steril 1996;65:925-930.
29. Reynolds LP, Grazul-Bilska AT, Redmer DA. Angiogenesis in the female reproductive organs: pathological implications. Int J Exp Pathol 2002;83:151-163.
30. Panganamamula UR, Harmanli OH, Isik-Akbay EF, Grotegut CA, Dandolu V, Gaughan JP: Is prior uterine surgery a risk factor for adenomyosis? Obstet Gynecol 2004;104:1034-1038.
31. Parazzini F, Vercellini P, Panazza S, Chatenoud L, Oldani S, Crosignani PG: Risk factors for adenomyosis. Hum Reprod 1997;12:1275-1279.
32. Curtis KM, Hillis SD, Marchbanks PA, Peterson HB: Disruption of the endometrial–myometrial border during pregnancy as a risk factor for adenomyosis. Am J Obstet Gynecol 2002;187:543-544.
計畫成果自評
本 研 究 內 容 與 原 計 畫 百 分 之 九 十 相 符,已達成預期目標,並已投稿至學術期刊
Am J Reprod Immunol (Impact factor =
1.808),目前 minor revision 中。在應用價值 方面,現有針對子宮腺肌症的治療方式,不 論藥物或手術,很難讓罹患子宮腺肌症婦女 同時達到懷孕與症狀緩解的目的,進一步瞭 解其免疫變化及致病機轉有助於未來尋求 更妥善的臨床處理方法。
附錄
Figure 1. Immunohistochemical study of IL-6 (A, B), IL-8 (C, D) and IL-10 (E, F) in eutopic
endometrium in women with and without adenomyosis. Experiments with the omission of primary antibodies were used as negative controls (G, H). Formalin-fixed, paraffin-embedded tissue samples were treated with anti-IL-6, anti-IL-8 and anti-IL-10 antibodies (1:50 dilution) using the Streptavidin-Biotin Universal Detection System as described in Materials and Methods. The color was developed using 3,3’ diaminobenzidine chromogen and counterstained by hematoxylin. Magnification, x400.
Figure 2. Comparison of IL-6, IL-8 and IL-10 mRNA levels in endometrial stromal cells (ESCs)
cultured (A) alone or (B) with macrophage between women with and without adenomyosis. It was done with real-time quantitative PCR, using β-actin as an internal control. Longitudinal axis indicates their relative values calculated by the formula: 2-∆∆CT, as described in the Materials and Methods. Bars represent the mean ± SEM.
* P = 0.006 by Mann-Whitney U-test.
TABLE I. Magnifying times of mRNA expression in endometrial stromal cells after coculture
with macrophage. Total (n = 21) Adenomyosis (n = 10) Controls (n = 11) IL-6 2.1 (0.6-16.2) 4.3 (1.2-59.2)b 0.8 (0.2-9.5)b IL-8 4.9 (1.8-28.5)a 2.3 (1.8-13.9) 9.6 (1.8-54.9) IL-10 1.1 (0.4-4)a 1.4 (0.4-5.6) 1.1 (0.3-5.1)
Note: Values are median (25%-75%).
a
P = 0.005 by Mann-Whitney U test
b