利用分子標誌輔助選拔水稻低白堊質品系
郭婷玫
1、林順福
1、簡禎佑
2、丁文彥
3、林家玉
3、陳榮坤
4、廖大經
5,*
1國立臺灣大學農藝學系
2行政院農委會桃園區農業改良場作物改良課
3行政院農委會臺東區農業改良場作物改良課
4行政院農委會臺南區農業改良場嘉義分場
5行政院農委會農業試驗所嘉義農業試驗分所農藝系
摘要
水稻近年的育種趨勢除了追求穩定高產
的目標外,提升米質亦為重點方向之一。白
堊質米粒不但容易造成碾米時米粒斷裂,白
米外觀亦為消費者在選購小包裝米的重要參
考依據;然白堊質常受到環境的影響,在不
同品種間也存在著差異。本研究利用分子標
誌 輔 助 選 拔
(marker-assisted-selection;
MAS)之技術,以白堊質率低的水稻品種「越
光」及高產品種「台農
67 號」作為雜交親本,
建立
F2
衍生系並區分為兩個族群進行。其
中,先行利用
150 個衍生系以 112 個 SSR 及
InDel 分子標誌建立連鎖群圖譜,並評估該族
群
F2
和
F2:3
之白米白堊質粒率、白堊質面積
與白堊質程度,以及株高、穗數與抽穗期等
農藝性狀,以區間定位法定位得到三個重要
數量性狀基因,分別位於第
5、7 及 12 對染
色體處,其表現相關的分子標誌為
RM1248、
CH0701 及 RM6306;另外再結合一組與「越
光」食味品質表現相關的分子標誌
RM4108,
共同作為另一族群
(860 個 F2
衍生系
)的篩
選。由以上
4 個分子標誌篩選得到的 23 個優
良品系,自
2011 年第 2 期作起分別種植於嘉
義、桃園及臺東,進行產量、米質及其他農
藝性狀的綜合評估,再配合利用分子標誌針
對這些獲選的優良品系中白堊質相關性狀之
分離基因座進行純化和單株選拔,最後推薦
19 個米質佳、豐產的優質品系做為各參試單
位命名及推廣之參考。
關鍵詞︰水稻、分子標誌輔助選拔、低白堊
質。
Marker-assisted Selection for Less
Grain Chalkiness Lines of Rice
Ting-Mei Kuo
1, Shun-Fu Lin
1, Jen-You
Jian
2, Wen-Yen Ting
3, Chia-Yu Lin
3,
Rong-Kuen Chen
4and Dah-Jing Liao
5,*
1 Department of Agronomy, National TaiwanUniversity, Taipei 10617, Taiwan ROC
2 Department of Crop Improvement, Taoyuan
District Agricultural Research and Extension Station, Shinwu, Taoyuan 32745, Taiwan ROC
3 Department of Crop Improvement, Taitung
District Agricultural Research and Extension Station, Taitung 95055, Taiwan ROC
4Chiayi Branch Station, Tainan District
Agricultural Research and Extension Station, Lucao, Chiayi 61151, Taiwan ROC
5 Department of Agronomy, Chiayi Agricultural
Experiment Branch, Taiwan Agricultural Research Institute, Chiayi 60044, Taiwan ROC
ABSTRACT
Recently the trend in rice breeding is not only focused on higher and stable yield of grains, but also the quality of grains. The chalky grains are easily broken during the process of milling, and the appearance of grains is the important * 通信作者, [email protected]
投稿日期:2014 年 8 月 7 日 接受日期:2014 年 11 月 24 日
作物、環境與生物資訊 11:230-242 (2014)
Crop, Environment & Bioinformatics 11:230-242 (2014) 189 Chung-Cheng Rd., Wufeng, Taichung 41362, Taiwan ROC
reference for consumers when they are buying a small pack of rice. However, the trait of chalkiness is easily affected by environment, and it also varies among rice varieties. In this study, we used marker-assisted-selection (MAS) to select superior lines with less chalky grain. The F2 derivative
lines developed from a cross between varieties ‘Koshihikari’ (less chalkiness) and ‘TNG67’ (high yield) were divided into mapping population and MAS population. A total of 150 lines were used to establish linkage map with 112 SSR and InDel markers, and map QTLs (quantitative trait loci) related to the percentage of grains with chalkiness, area of chalky endosperm, degree of endosperm chalkiness, and some agronomic traits. Three QTLs mapped around markers RM1248, CH0701 and RM6306 were distributed on 5th, 7th and 12th chromosomes, respectively. These markers were applied to screen 860 F2 derivative lines, and 23
superior lines were selected. These superior lines were planted in Taoyuan, Chiayi and Taitung to assess grain yield, grain quality and other agronomic traits, and were purified with SSR markers. Further characterization of 19 lines with excellent grain quality and high grain yield were recommended as the reference of extension and registration of plant variety properties for all participating breeding stations or research institutes.
Key words: Rice, Marker-assisted selection, Chalkiness.
前言
稻米是全球重要的糧食之一,近年隨著
國家經濟與國民生活水平的提高,稻米生產
從早期以高產為目標,近年亦重視品質的改
良。稻米白堊質除了影響米粒透明度而不利
於外觀品質外,也由於碎米粒增加而導致完
整米比率降低,造成整體商品價值的損失。
遺傳與環境因素同時影響米粒的白堊率以及
白堊面積大小(Takeda et al. 1983, Kuo and
Liu 1986),而白堊質屬於數量性狀遺傳(Tan
et al. 2000, Shi et al. 2002),因此白堊質主要
是 數 量 性 狀 基 因 (quantitative trait loci;
QTLs)與環境因素的交感效果,所以傳統育
種 方 法 對 於 選 育 低 白 堊 質 品 系 有 其 困 難 之
處 。 近 年 來 隨 著 水 稻 分 子 標 誌 的 開 發 , 如
RFLP (restriction fragment length
polymorphism) 、 RAPD (random
amplified polymorphic DNA) 、 AFLP
(amplified fragment length
polymorphism) 、 SSR (simple sequence
repeat) 及
SNP (single nucleotide
polymorphism)等,以及統計方法的進步與
電腦軟體運算能力的提升,使得
QTLs 的定
位較以往更為精準,而利用分子標誌技術偵
測影響米粒白堊質性狀
QTLs 的研究也日益
增加。
目前已發表偵測白堊質性狀
QTLs 的連
鎖圖譜族群大多以秈、稉雜交為主,例如
He
et al. (1999)以秈稻品系‘ZYQ8’與稉稻品系
‘JX17’ 雜 交 後 代 F1
之 單 倍 體 倍 加
(doubled-haploid; DH)族群為材料,測得影
響白堊質
QTLs 主要位於第 8 及第 12 對染色
體 ;
Wan et al. (2005) 則 以 稉 稻 品 種
‘Asominori’與秈稻品種‘IR24’雜交組合之染
色 體 置 換 品 系 (chromosome segment
substitution lines; CSSLs)為對象,在 8 個不
同栽培環境下,一致測得影響稻米白堊質率
之
QTLs 主要位於第 8 及第 9 對染色體。然
而
Ebitani et al. (2008)及 Zhou et al. (2009)
分 別 以 稉 稻 品 種 越 光 與 秈 稻 品 種
’Kasalath’
之雜交組合,以及兩系雜交稻品種‘PA64s’與
秈 稻 品 系
‘9311’之雜交組合的 CSSLs 為材
料,卻發現主要影響白堊質之
QTLs 分別位
於第
1 及第 7 對(Ebitani et al. 2008),或者第
6 及第 7 對染色體上(Zhou et al. 2009)。而以
稉、稉雜交後代族群分析稻米白堊質
QTLs
之研究則有
Kobayashi et al. (2007)以稉稻品
種
‘Hana-echizen’與‘Niigatawase’的雜交組
合後代
F2
及
F3
族群為對象,測得影響稻米白
堊質之
QTLs 位於第 3、第 4 及第 6 (主要)對
染 色 體 ;
Tabata et al. (2007) 以 稉 稻 品
種’Chiyonishiki’與’Koshijiwase’雜交組合之
F6
重 組 自 交 系
(recombinant inbred lines;
RILs)為材料,分析稻米白堊質有關之 QTLs
位於第
1、第 2 及第 8 對染色體上;而 Kepiro
et al. (2008) 亦 以 稉 稻 品 種 ’Cypress’
與’Panda’雜交組合之 F11 RILs 為材料,獲得
控制稻米白堊質之
QTLs 位於第 2 及第 6(主
要)對染色體上。
歸納以上文獻的研究結果顯示,影響稻
米 白堊 質之
QTLs 位置因不同雜交組合而
異,且易受栽培環境影響。臺灣以稉稻品種
栽培為主,因此本研究以具有極少稻米白堊
質且食味佳之優良稉稻品種「越光」
,和廣適
應性之穩定高產稉稻品種「台農
67 號」為雜
交親本,產生
F2
雜交族群,並建立分子標誌
遺傳圖譜。並依據
F2:3
品系之性狀調查資料
定位
QTLs,以進行分子標誌輔助選種工作,
藉此探討影響稻米白堊質之基因表現,選育
適合台灣栽培之高產且外觀品質優良之新品
種。
材料與方法
一、試驗材料
本研究以稉稻品種「越光」為母本,以
及另一稉稻品種「臺農
67 號」為父本,於 2008
年第
2 期作於臺南區農業改良場(以下簡稱臺
南場)進行雜交獲得 F1
世代種子,種植
F1
世
代植株經自交後獲得
F2
世代共計
1010 粒種
子。其中,150 個 F2
單株種植及雜交親本於
2010 年第 1 期作在行政院農委會農業試驗所
嘉義農業試驗分所(以下簡稱嘉義分所)溫室
種 植 做 為 基 因 定 位 用
F2
族 群
(mapping
population),其他 860 個單株則種植於桃園
區農業改良場(以下簡稱桃園場)之田區中,成
為 分 子 標 誌 輔 助 選 種 用
F2
族 群
(MAS
population),田間管理與病蟲防治則參照良
質米推薦方式。
二、建立
F2
連鎖群圖譜
2010 年第 1 期作種植於嘉義分所溫室之
150 株 F2
單株種植及雜交親本,約於
4 葉齡
苗期時分別予以編號,並且剪取新鮮葉片樣
本送交國立臺灣大學抽取
DNA 用以建立 F2
連鎖群圖譜。本研究參考
Gramene database
設計引子來篩選多型性之
SSR 或 InDel 分子
標誌,使用軟體
MAPMAKER 3.0 (Lincoln et
al. 1993),以 LOD 為 3.0 門檻值及最大遺傳
距離
40 cM 之條件,進行分子標誌連鎖分析
與遺傳距離估算,完成建立分子標誌連鎖圖
譜,並以
MapChart 軟體進行遺傳圖譜之繪
製。
三、QTLs 定位
本研究進行
QTLs 定位所使用族群為種
植於嘉義分所溫室之
F2
族群,以及由單株收
穫之
F2:3
族群。F2:3
族群於
2010 年第 2 期作
種植分別在嘉義分所、桃園場與臺東區農業
改良場(以下簡稱臺東場)種植,採隨機完全區
集設計(RCBD),2 重複,每重複種植 1 短行,
每行有
10 株,單本植,雜交親本各種植 2 短
行。F2:3
族群調查性狀包括株高、抽穗期、白
堊質率與白堊質面積等項目,以供
QTLs 定
位分析使用。QTLs 定位係採用 Lander and
Botstein (1989)提出之區間定位法(interval
mapping; IM),使用軟體 Windows QTLs
Cartographer 2.5 版(Wang et al. 2011)進行
定位分析,其中分子標誌距離若大於
40 cM
之區間則不用於
QTLs 的定位分析,LOD 門
檻值則設定為
2.0。
四、利用分子標誌輔助選拔低白堊質品系
根據
QTLs 定位分析結果,選出與白堊
質率
QTLs 緊密連鎖之分子標誌,以及已知
位於品種「越光」第
3 對染色體與食味品質
相關
QTLs 緊密連鎖之分子標誌(Wada et al.
2008),利用這些分子標誌從 860 個 F2:3
品系
中篩選出具有優良基因型品系
50–60 個,也
同時利用其他與食味品質及抽穗期有關之分
子標誌進行篩選,最後篩選出
30–40 個同結
合基因型的
F2:3
品系。
五、
選獲優良 F2:3
品系在多地區進行白
堊質及其他農藝性狀評估
2011 年第 1 期作篩選之優良 F2:3
品系,
於同年第
2 期作分別於嘉義分所、桃園場及
臺東場進行白堊質及其他農藝性狀評估,採
RCBD 設計,2 重複,每品系種植 3 行,行長
度
2 公尺。調查性狀有抽穗期、株高、穗數、
千粒重及穀粒產量等一般農藝性狀,以及白
堊 質 率
(percentage of grains with
chalkiness; PGWC) 、 白 堊 質 面 積 (area of
chalky endosperm; ACE) 與 白 堊 質 程 度
(degree of endosperm chalkiness; DEC)等
白堊質性狀。本研究供分析使用之糙米樣本
係碾製
1 次,分析米粒白堊質相關性狀係使
用日本
SATAKE 公司製造之穀粒判別器(型
號
RGQI20A)。白堊質性狀調查項目及調查
方式說明如下:
白堊質率:計算
100 粒米粒中,含有白
堊質米粒之比例。
白堊質面積:自
100 粒米粒中挑選白堊
質米粒,並計算白堊質面積佔該米粒總面積
的比例。
白堊質程度:由白堊質率與白堊質面積
相乘而得。
六、統計分析
獲選品系間之白堊質率與白堊質面積使
用變方分析比較其差異,以上統計分析使用
SAS/STAT 9.1 完成。
結果
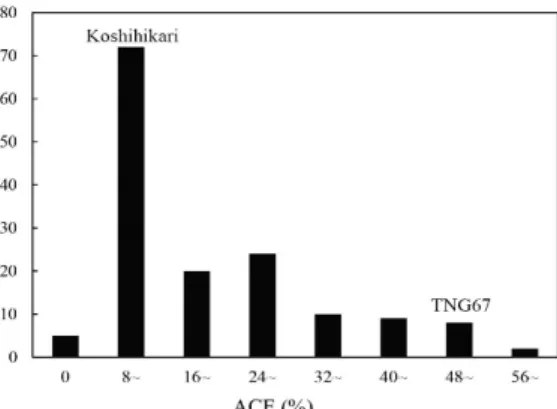
一、
F2
族群之白堊質性狀分佈
種植於嘉義分所溫室之
150 株 F2
族群之
白堊質率與白堊質面積分佈如
Fig. 1、Fig. 2
顯示,雜交親本「越光」的白堊質率為
26.9%,
另一親本「台農
67」號則為 95.0%,各單株
之白堊質率範圍介於
0–100%之間,而以 20–
30%為最多數。F2
族群米粒白堊質面積百分
比變異範圍介於
0–62.5%之間,以 8–15%為
最多,而雜交親本「越光」為
12.9 %,「台農
67」號則為 47.4 %。
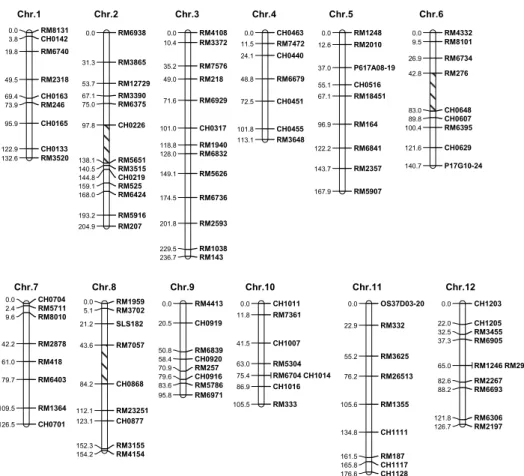
二、連鎖圖譜之建立
本研究以
112 個 SSR 與 InDel 分子標誌
所建立之連鎖圖譜,共分為
15 個連鎖群,分
別位於
12 對染色體上,總長為 1781.2 cM,
相鄰分子標誌平均距離為
16.2 cM (Fig. 3)。
三、
QTLs 定位
本研究以區間定位法分析
F2
族群之白堊
質率、白堊質面積與白堊質程度等白堊質相
關性狀與抽穗期性狀基因的表現,如
Table
1、Table 2 所示;F2:3
族群白堊質相關性狀與
株高、穗數及抽穗期等一般農藝性狀基因的
表現如
Table 3、Table 4 所示 。F2
族群與
F2:3
族群共分別偵測到
8 個及 6 個與白堊質性狀
相關的
QTLs,其中影響白堊質率的 QTLs,
F2
族群有
3 個,F2:3
族群有
1 個;影響白堊質
面積的
QTLs,F2
族群有
3 個,F2:3
族群有
3
個;影響白堊質程度的
QTLs,F2
族群有
2
個,F2:3
族群有
2 個。另外,F2:3
族群也偵測到
影響一般農藝性狀的
QTLs 共計 12 個,包括了
Fig. 1. Frequency distribution for PGWC in F2 lines.
Fig. 3. Linkage map of a rice F2 population, ‘Koshihikari’ ‘TNG67’, constructed with 112 SSR and InDel
DNA markers.
Table 1. Identification of QTLs for grain chalkiness related traits of a rice F2 population, ‘Koshihikari’
‘TNG67’ .
Trait QTLs Chr. Interval Position(cM) LOD Additive effect (%) R2
PGWC z 2 RM525 0 1.8 3.97 7.4
q-PGWC3 5 P617A08-19 - CH0516 4 2.0 -2.28 11.7
6 CH0629 0 1.6 -4.12 6.5
q-PGWC7a 7 RM8010 - RM2878 28 2.2 5.03 13.0
q-PGWC7b 7 RM6403 - RM1364 12 3.1 13.11 31.2 ACE y q-ACE5 5 RM2010 - P617A08-19 4 4.6 -6.94 22.2
7 RM6403 - RM1364 16 1.5 5.86 14.2
q-ACE8 8 RM1959 0 2.0 4.72 8.1
10 CH1016 0 1.5 4.69 6.3
q-ACE11 11 RM26513 - RM1355 20 2.3 6.89 22.5 DEC x q-DEC5 5 RM2010 - P617A08-19 12 2.2 -302.54 13.2
q-DEC7a 7 RM6403 - RM1364 14 2.8 569.72 30.6
z percentage of grains with chalkiness. y area of chalky endosperm.
x degree of endosperm chalkiness.
Chr.1 Chr.2 Chr.3 Chr.4 Chr.5 Chr.6 RM8131 0.0 CH0142 3.8 RM6740 19.8 RM2318 49.5 CH0163 69.4 RM246 73.9 CH0165 95.9 CH0133 122.9 RM3520 132.6 RM6938 0.0 RM3865 31.3 RM12729 53.7 RM3390 67.1 RM6375 75.0 CH0226 97.8 RM5651 138.1 RM3515 140.5 CH0219 144.8 RM525 159.1 RM6424 168.0 RM5916 193.2 RM207 204.9 RM4108 0.0 RM3372 10.4 RM7576 35.2 RM218 49.0 RM6929 71.6 CH0317 101.0 RM1940 118.8 RM6832 128.0 RM5626 149.1 RM6736 174.5 RM2593 201.8 RM1038 229.5 RM143 236.7 CH0463 0.0 RM7472 11.5 CH0440 24.1 RM6679 48.8 CH0451 72.5 CH0455 101.8 RM3648 113.1 RM1248 0.0 RM2010 12.6 P617A08-19 37.0 CH0516 55.1 RM18451 67.1 RM164 96.9 RM6841 122.2 RM2357 143.7 RM5907 167.9 RM4332 0.0 RM8101 9.5 RM6734 26.9 RM276 42.8 CH0648 83.0 CH0607 89.8 RM6395 100.4 CH0629 121.6 P17G10-24 140.7 Chr.7 Chr.8 Chr.9 Chr.10 Chr.11 Chr.12 CH0704 0.0 RM5711 2.4 RM8010 9.6 RM2878 42.2 RM418 61.0 RM6403 79.7 RM1364 109.5 CH0701 126.5 RM1959 0.0 RM3702 5.1 SLS182 21.2 RM7057 43.6 CH0868 84.2 RM23251 112.1 CH0877 123.1 RM3155 152.3 RM4154 154.2 RM4413 0.0 CH0919 20.5 RM6839 50.8 CH0920 58.4 RM257 70.9 CH0916 79.6 RM5786 83.6 RM6971 95.8 CH1011 0.0 RM7361 11.8 CH1007 41.5 RM5304 63.0 RM6704 CH1014 75.4 CH1016 86.9 RM333 105.5 OS37D03-20 0.0 RM332 22.9 RM3625 55.2 RM26513 76.2 RM1355 105.6 CH1111 134.8 RM187 161.5 CH1117 165.8 CH1128 176.6 CH1203 0.0 CH1205 22.0 RM3455 32.5 RM6905 37.3 RM1246 RM2972 65.0 RM2267 82.6 RM6693 88.2 RM6306 121.8 RM2197 126.7
Table 2. Identification of QTLs for grain chalkiness related traits of a rice F2:3 population, ‘Koshihikari’
’TNG67’ .
Trait QTLs Chr. Interval Position (cM) LOD Additive effect R2 (%)
PGWC z 1 CH0165 - CH0133 16 1.6 -0.08 15.1 2 RM6938 0 1.5 -1.37 6.7 3 RM7576 - RM218 8 1.9 2.93 12.7 4 CH0451 - CH0455 14 1.8 -2.21 14.8 5 RM1248 0 1.8 -1.58 7.7 7 RM1364 - CH0701 16 1.6 -0.39 7.6 q-PGWC12 12 RM6693 - RM6306 18 2.3 0.04 24.7 ACE y 1 RM8131 0 1.8 0.84 8.0 q-ACE2 2 RM6938 0 2.2 -2.53 9.3 q-ACE3 3 CH0317 - RM1940 6 2.5 4.04 13.9 q-ACE6 6 RM4332 0 2.0 -3.40 8.8 7 RM2878 0 1.6 2.68 6.8 7 RM1364 - CH0701 8 1.9 0.38 14.1 10 CH1016 0 1.9 -0.58 8.1 DEC x q-DEC2 2 RM6938 0 2.2 -58.36 9.6 q-DEC7b 7 RM1364 - CH0701 12 2.0 -5.78 12.6
z percentage of grains with chalkiness. y area of chalky endosperm.
x degree of endosperm chalkiness.
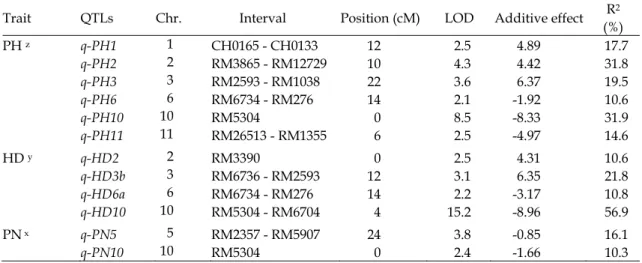
Table 3. Identification of QTLs for days to heading of a rice F2 population, ‘Koshihikari’ ‘TNG67’ .
Trait QTLs Chr. Interval Position (cM) LOD Additive effect R2 (%)
HD z q-HD3a 3 RM1038 0 5.7 6.09 22.0 q-HD6a 6 RM6734 – RM276 14 5.3 -5.93 22.7 q-HD6b 6 CH0629 – P17G10-24 10 2.1 -0.98 16.1 q-HD7 7 RM0810 0 2.2 3.95 9.4 q-HD10 10 RM5304 – RM6704 4 9.3 -6.75 36.6 z days to heading.
Table 4. Identification of QTLs for plant height, days to heading and panicle number of a rice F2:3
population, ‘Koshihikari’ ‘TNG67’ .
Trait QTLs Chr. Interval Position (cM) LOD Additive effect R2 (%) PH z q-PH1 1 CH0165 - CH0133 12 2.5 4.89 17.7 q-PH2 2 RM3865 - RM12729 10 4.3 4.42 31.8 q-PH3 3 RM2593 - RM1038 22 3.6 6.37 19.5 q-PH6 6 RM6734 - RM276 14 2.1 -1.92 10.6 q-PH10 10 RM5304 0 8.5 -8.33 31.9 q-PH11 11 RM26513 - RM1355 6 2.5 -4.97 14.6 HD y q-HD2 2 RM3390 0 2.5 4.31 10.6 q-HD3b 3 RM6736 - RM2593 12 3.1 6.35 21.8 q-HD6a 6 RM6734 - RM276 14 2.2 -3.17 10.8 q-HD10 10 RM5304 - RM6704 4 15.2 -8.96 56.9 PN x q-PN5 5 RM2357 - RM5907 24 3.8 -0.85 16.1 q-PN10 10 RM5304 0 2.4 -1.66 10.3 z plant height. y days to heading. x panicle number.
影響株高的
QTLs 有 6 個,影響穗數的 QTLs
有
2 個,以及影響抽穗期的 QTLs 有 4 個。
(一)白堊質率
F2族群偵測到影響白堊質率的
QTLs 有 3
個
(Table 1) , 分 別 位 於 第 5 對 染 色 體
P617A08-1CH0516 之區間內(q-PGWC3)、
第
7 對染色體 RM8010─RM2878 之區間內
(q-PGWC7a) 、 及 第
7
對 染 色 體
RM8010RM2878 之區間內(q-PGWC7b)。加
性效應分別為-2.28、5.03 與 13.11,外表型貢
獻量分別為
11.7、13.0 及 31.2%。
F2:3
族 群 偵 測 到
1 個影響白堊質率的
QTLs (Table 2) , 位 於 第 12 對 染 色 體
RM6693–RM6306 之區間內(q-PGWC12),加
性效應為
0.04%,外表型貢獻量為 24.7%。
(二)白堊質面積
F2
族群偵測到影響白堊質面積的
QTLs
有
3 個(Table 1),分別位於第 5 對染色體
RM2010–P617A08-19 之區間內(q-ACE5)、第
8 對染色體 RM1959 上(q-ACE8)、及第 11 對
染 色 體
RM26513–RM1355 之 區 間 內
(q-ACE11)。加性效應分別為-6.94、4.72 與
6.89,而外表型貢獻量則分別為 22.2、8.1 及
22.5%。
F2:3
族群偵測到
3 個影響白堊質面積的
QTLs (Table 2),分別位於第 2 對染色體
RM6938 上(q-ACE2)、第 3 對染色體 CH0317–
RM1940 之區間內(q-ACE3)、及第 6 對染色
體
RM4332 上(q-ACE6)。加性效應分別為
-2.53、4.04 與-3.40,外表型貢獻量分別為
9.3、13.9 及 8.8%。
(三)白堊質程度
F2
族群偵測到影響白堊質程度的
QTLs
有
2 個(Table 1),分別位於第 5 對染色體
RM2010–P617A08-19 之區間內 (q-DEC5)及
第
7 對染色體 RM6403–RM1364 之區間內
(q-DEC7) 。 加 性 效 應 分 別 為 -302.54 與
569.72,外表型貢獻量分別為 13.2 %與 30.6 %。
F2:3
族群偵測到
2 個影響白堊質程度的
QTLs (Table 2),分別位於第 2 對染色體
RM6938 上 (q-DEC2) 及 第 7 對 染 色 體
RM1364–CH0701 之區間內(q-DEC7b)。加性
效應分別為-58.36 與-5.78,外表型貢獻量分
別為
9.6 %與 12.6 %。
(四)株高
F2:3
族群偵測到影響株高的
QTLs 有 6 個
(Table 4),分別位於第 1 對染色體 CH0165–
CH0133 之區間內(q-PH1)、第 2 對染色體
RM3865–RM12729 之區間內(q- PH2)、第 3
對 染 色 體
RM2593–RM1038 之 區 間 內 (q-
PH3)、第 6 對染色體 RM6734–RM276 之區
間內(q- PH6)、第 10 對染色體 RM5304 上(q-
PH10)、及第 11 對染色體 RM26513RM1355
之區間內(q- PH11)。加性效應分別為 4.89、
4.42、6.37、-1.92、-8.33 與-4.97 cm,外表型
貢獻量分別為
17.7、31.8、19.5、10.6、31.9
及
14.6%。
(五)穗數
F2:3
族群偵測到影響穗數的
QTLs 有 2 個
(Table 4),分別位於第 5 對染色體 RM2357–
RM5907 之區間內(q-PN5)、及第 10 對染色體
RM5304 上(q- PN10)。加性效應分別為-0.85
穗與-1.66 穗,外表型貢獻量分別為 16.1%與
10.3%。
(六)抽穗期
F2
族群偵測到影響抽穗期的
QTLs 有 5
個(Table 3),分別位於第 3 對染色體 RM1038
上(q-HD3a)、第 6 對染色體 RM6734–RM276
之區間內(q-HD6a)、第 6 對染色體 CH0629–
P17G10-24 之區間內(q-HD6b)、第 7 對染色
體
RM8010 上(q-HD7)、及第 10 對染色體
RM5304-RM6704 之區間內(q-HD10)。加性
效應分別為
6.09、-5.93、-0.98、3.95 與-6.75
d,外表型貢獻量分別為 22.0、22.7、16.1、
9.4 及 36.6%。F2:3
族群偵測到
4 個影響抽穗
期的
QTLs (Table 4),分別位於第 2 對染色
體
RM3390 上 (q-HD2) 、 第 3 對 染 色 體
RM6736–RM2593 之區間內(q-HD3b)、第 6
對 染 色 體
RM6734–RM276 之 區 間 內
(q-HD6a) 、 及 第 10 對 染 色 體 RM5304–
RM6704 之區間內(q-HD10)。加性效應分別
為
4.31、6.35、
-3.14 與-8.96 d,外表型貢獻量分別為 10.6、
21.8、10.8 及 56.9%。
三、
利用分子標誌輔助選拔優良基因型
品系
綜合
F2
與
F2:3
兩代結果從中選出與白堊
質率表現相關的
QTLs,分別位於第 5、7 與
12 對染色體處,並且以與之連鎖的分子標誌
RM1248、CH0701 和 RM6306,以及另一與
「 越 光 」 食 味 品 質 表 現 相 關 的 分 子 標 誌
RM4108,作為後續輔助選拔之用。利用以上
4 個 分 子 標 誌 , 以 及 4 個 與 食 味 品 質
(RM4332)及 抽 穗 期 (RM6704、 RM1038 和
RM6734)相關的分子標誌,針對相同雜交組
合的
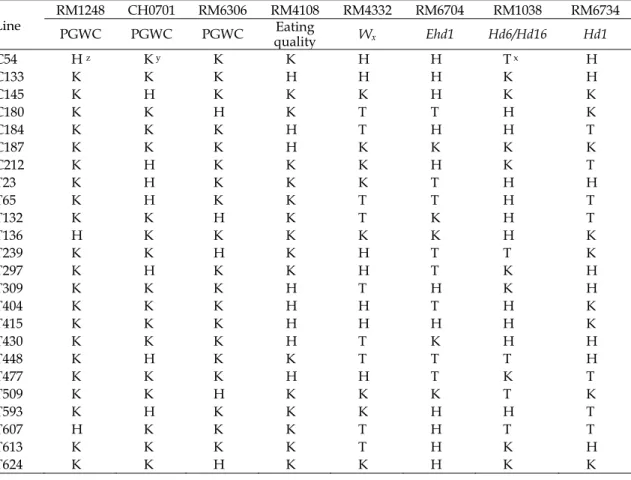
860 個 MAS 族群,共篩選出 24 個具有
優良基因型的水稻品系(Table 5)。除了品系
T613 之 4 組分子標誌皆與「越光」基因型相
同,為同質結合基因型外,其餘
23 個品系則
有
1 組分子標誌為異質結合基因型,而其餘 3
組分子標誌皆為同質結合基因型。
四、
優良基因型品系於不同地區之白堊
質與一般農藝性狀表現
23 個具有優良基因型的水稻 F2:4
品系,
於
2011 年第 2 期作分別於嘉義分所、桃園場
及臺東場種植,並且調查白堊質率、白堊質
面積與白堊質程度,以及株高、穗數與抽穗
期等一般農藝性狀表現(Tables 6 and 7),其
Table 5. Genotypes of DNA makers associated with grain chalkiness, eating quality and days to heading traits for 24 rice superior lines.
Line RM1248 CH0701PGWC PGWC PGWC EatingRM6306 RM4108 RM4332 RM6704 RM1038 RM6734 quality Wx Ehd1 Hd6/Hd16 Hd1 C54 H z K y K K H H T x H C133 K K K H H H K H C145 K H K K K H K K C180 K K H K T T H K C184 K K K H T H H T C187 K K K H K K K K C212 K H K K K H K T T23 K H K K K T H H T65 K H K K T T H T T132 K K H K T K H T T136 H K K K K K H K T239 K K H K H T T K T297 K H K K H T K H T309 K K K H T H K H T404 K K K H H T H K T415 K K K H H H H K T430 K K K H T K H H T448 K H K K T T T H T477 K K K H H T K T T509 K K H K K K T K T593 K H K K K H H T T607 H K K K T H T T T613 K K K K T H K H T624 K K H K K H K K
z K: same genotypes as ‘Koshihikari’ . y T: same genotypes as ‘TNG67’ . x H: heterozygous genotypes.
中桃園場及臺東場之白堊質相關性狀調查因
缺植較多故未列入統計分析。針對嘉義分所
調查之白堊質率與白堊質面積進行變方分析
顯示,不同品系間白堊質率呈極顯著差異,
而白堊質面積則不存在顯著差異(Table 8)。
其中除
C54 品系之平均白堊質率 29.2%大於
親本「台農
67 號」之 27.8%外,其他品系之
平均白堊質率均小於「台農
67 號」。而所有
品系之白堊質面積均超過
50%,介於 54-90%
之間。三個地區之一般農藝性狀,除了
T477
品系的平均抽穗期
68.3 d,大於親本「台農
67 號」之 65 d 以外,其他品系均小於 65 d。
株高除了
C133 與 T477 品系高於「台農 67
號」之平均值
109.9 cm 外,其他品係均低於
「台農
67 號」。
五、優良品系之基因型純化及單株選拔
以五組分子標誌
RM1248、CH0701、
RM6306、RM4108 及 RM4332,針對 23 個優
良品系中與白堊質粒率及食味品質相關基因
座於前世代為異結合基因型者進行純化,在
F2:5
中篩選同結合型單株,可避免該基因座於
後代繼續分離。其中
13 個品系僅有 1 組分子
標誌分離如
Table 9 所示,各品系可選取與越
光相同之同型結合個體,以避免後代基因型
繼續分離;另有
6 個品系有 2 組分子標誌分
離如
Table 10 所示,各品系需選取兩分子標
誌基因型均與越光相同的同型結合個體。理
論上單一基因座分離出任一種同結合基因型
之比例為
3/8,在 95%之信心水準下,若種
Table 6. Grain chalkiness related traits for 23 rice superior lines (F2:4) obtained by marker assisted selection.
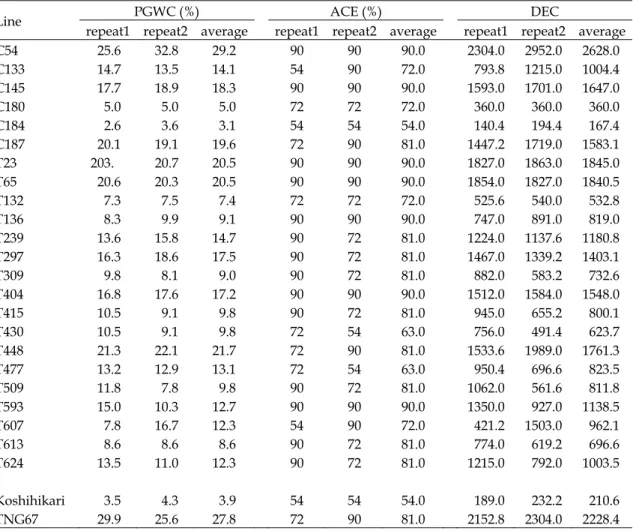
Line PGWC (%) ACE (%) DEC
repeat1 repeat2 average repeat1 repeat2 average repeat1 repeat2 average
C54 25.6 32.8 29.2 90 90 90.0 2304.0 2952.0 2628.0 C133 14.7 13.5 14.1 54 90 72.0 793.8 1215.0 1004.4 C145 17.7 18.9 18.3 90 90 90.0 1593.0 1701.0 1647.0 C180 5.0 5.0 5.0 72 72 72.0 360.0 360.0 360.0 C184 2.6 3.6 3.1 54 54 54.0 140.4 194.4 167.4 C187 20.1 19.1 19.6 72 90 81.0 1447.2 1719.0 1583.1 T23 203. 20.7 20.5 90 90 90.0 1827.0 1863.0 1845.0 T65 20.6 20.3 20.5 90 90 90.0 1854.0 1827.0 1840.5 T132 7.3 7.5 7.4 72 72 72.0 525.6 540.0 532.8 T136 8.3 9.9 9.1 90 90 90.0 747.0 891.0 819.0 T239 13.6 15.8 14.7 90 72 81.0 1224.0 1137.6 1180.8 T297 16.3 18.6 17.5 90 72 81.0 1467.0 1339.2 1403.1 T309 9.8 8.1 9.0 90 72 81.0 882.0 583.2 732.6 T404 16.8 17.6 17.2 90 90 90.0 1512.0 1584.0 1548.0 T415 10.5 9.1 9.8 90 72 81.0 945.0 655.2 800.1 T430 10.5 9.1 9.8 72 54 63.0 756.0 491.4 623.7 T448 21.3 22.1 21.7 72 90 81.0 1533.6 1989.0 1761.3 T477 13.2 12.9 13.1 72 54 63.0 950.4 696.6 823.5 T509 11.8 7.8 9.8 90 72 81.0 1062.0 561.6 811.8 T593 15.0 10.3 12.7 90 90 90.0 1350.0 927.0 1138.5 T607 7.8 16.7 12.3 54 90 72.0 421.2 1503.0 962.1 T613 8.6 8.6 8.6 90 72 81.0 774.0 619.2 696.6 T624 13.5 11.0 12.3 90 72 81.0 1215.0 792.0 1003.5 Koshihikari 3.5 4.3 3.9 54 54 54.0 189.0 232.2 210.6 TNG67 29.9 25.6 27.8 72 90 81.0 2152.8 2304.0 2228.4
植
8 株可望至少獲得一株預期之基因型,故
本試驗針對單一基因型分離之品系種植
10
株,而兩對基因分離之品系則種植
20 株。試
驗結果顯示,在
13 個單一分子標誌分離品系
中,除了
C187 品系未能獲得預期基因型個體
外,其餘
12 個品系均能獲選 1 株以上之預期
基因型個體。在
6 個 2 組分子標誌分離品系
中,除了
T239 品系未能獲得預期基因型個體
外,其他
5 個品系亦均能選獲 1 株以上之預
期基因型個體。而
C187 與 T239 品系再次分
析 10 株幼苗後,亦均能獲得 1 株預期基因型
個體。以上
19 個獲選品系除了可增加遺傳純
度外,亦可降低白堊質米粒比例,可同時提
高米粒外觀與食味品質。
Table 7. Days to heading and plant height for 23 rice superior lines (F2:4) obtained by marker assisted
selection.
Days to heading Plant height (cm)
Line Chiayi Taoyuan Taitung average Chiayi Taoyuan Taitung average
C54 56 51 61 56.0 106.6 100.3 99.5 102.1 C133 56 55 63 58.0 122.0 107.8 101.7 110.5 C145 63 54 67 61.3 110.8 108.8 104.0 107.9 C180 43 47 46 45.3 83.4 68.5 78.1 76.7 C184 46 47 51 48.0 97.2 90.2 81.2 89.5 C187 43 51 54 49.3 96.8 91.1 85.0 91.0 T23 63 53 65 60.3 115.0 105.9 102.2 107.7 T65 63 52 65 60.0 116.6 105.7 102.5 108.3 T132 43 47 44 44.7 85.8 68.5 82.1 78.8 T136 50 50 53 51.0 92.2 88.4 89.8 90.1 T239 59 51 62 57.3 101.8 104.1 100.1 102.0 T297 69 53 66 62.7 102.0 98.2 102.1 100.8 T309 50 47 51 49.3 100.0 95.2 91.9 95.7 T404 59 51 61 57.0 98.0 93.9 87.0 93.0 T415 43 49 45 45.7 83.6 85.5 81.4 83.5 T430 46 47 50 47.7 97.8 86.7 88.6 91.0 T448 59 52 65 58.7 101.4 94.4 91.2 95.7 T477 73 56 76 68.3 134.0 106.9 101.7 114.2 T509 50 48 52 50.0 98.2 92.6 89.4 93.4 T593 50 51 51 50.7 104.8 101.2 92.2 99.4 T607 43 50 46 46.3 83.0 80.2 86.9 83.4 T613 43 48 46 45.7 80.2 88.3 89.5 86.0 T624 50 48 51 49.7 103.0 93.0 92.9 96.3 Koshihikari 46 46 - 46.0 96.4 79.8 - 88.1 TNG67 73 57 - 65.0 128.8 91.0 - 109.9
Table 8. ANOVA of chalkiness related traits of 23 rice superior lines (F2:4).
SOV PGWC ACE
df MS F-test df MS F-test
Line 22 75.30 16.69** 22 201.06 1.47
Block 1 1.43 0.32 1 63.39 0.46
Error 22 4.51 22 137.03
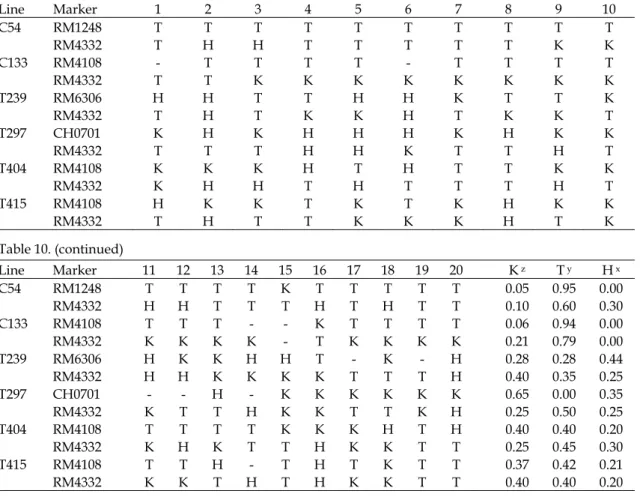
Table 9. Genotypic selection for 13 rice superior lines F2:5 by one marker. Line Marker 1 2 3 4 5 6 7 8 9 10 K z T y H x C145 CH0701 K K K T T - K K K K 0.78 0.22 0.00 C184 RM4108 K K K T K K K K K T 0.80 0.20 0.00 C187 RM4108 T T T T T T T T T T 0.00 1.00 0.00 T23 CH0701 K K K K K K K K K K 1.00 0.00 0.00 T65 CH0701 T T T T K K K - T K 0.44 0.56 0.00 T132 RM6306 K K K K K K K K K K 1.00 0.00 0.00 T136 RM1248 K K K T K K K T K K 0.80 0.20 0.00 T309 RM4108 T H T K K K K K K H 0.60 0.20 0.20 T448 CH0701 T T H K T T K T T T 0.20 0.70 0.10 T477 RM4108 K T H K K K K H K T 0.60 0.20 0.20 T593 CH0701 H T H T T T K T H T 0.10 0.60 0.30 T613 RM4108 K K K H K K K K K K 0.90 0.00 0.10 T624 RM6306 H T T K K H K H H T 0.30 0.30 0.40
z K: same genotypes as ‘Koshihikari’ . y T: same genotypes as ‘TNG67’ . x H: heterozygous genotypes.
Table 10. Genotypic selection for 6 rice superior lines F2:5 by two markers.
Line Marker 1 2 3 4 5 6 7 8 9 10 C54 RM1248 T T T T T T T T T T RM4332 T H H T T T T T K K C133 RM4108 - T T T T - T T T T RM4332 T T K K K K K K K K T239 RM6306 H H T T H H K T T K RM4332 T H T K K H T K K T T297 CH0701 K H K H H H K H K K RM4332 T T T H H K T T H T T404 RM4108 K K K H T H T T K K RM4332 K H H T H T T T H T T415 RM4108 H K K T K T K H K K RM4332 T H T T K K K H T K Table 10. (continued) Line Marker 11 12 13 14 15 16 17 18 19 20 K z T y H x C54 RM1248 T T T T K T T T T T 0.05 0.95 0.00 RM4332 H H T T T H T H T T 0.10 0.60 0.30 C133 RM4108 T T T - - K T T T T 0.06 0.94 0.00 RM4332 K K K K - T K K K K 0.21 0.79 0.00 T239 RM6306 H K K H H T - K - H 0.28 0.28 0.44 RM4332 H H K K K K T T T H 0.40 0.35 0.25 T297 CH0701 - - H - K K K K K K 0.65 0.00 0.35 RM4332 K T T H K K T T K H 0.25 0.50 0.25 T404 RM4108 T T T T K K K H T H 0.40 0.40 0.20 RM4332 K H K T T H K K T T 0.25 0.45 0.30 T415 RM4108 T T H - T H T K T T 0.37 0.42 0.21 RM4332 K K T H T H K K T T 0.40 0.40 0.20
z K: same genotype as ‘Koshihikari’ . y T: same genotypes as ‘TNG67’ . x H: heterozygous genotypes.
討論
本研究以
112 組多型性分子標誌用於建
立連鎖圖譜,但是在第
2、第 6 及第 8 對染色
體仍有較大之區間,雖然對於這
3 個區間持
續篩選分子標誌,卻仍無法篩選出多型性分
子標誌,所以未來有必要增加該區間的分子
標誌數目。
應用於分子標誌輔助選拔主要依據
F2:3
族群所定位,與影響白堊質性狀之
QTLs 連
鎖的分子標誌,而
F2
族群的定位結果則供做
參考用。原因在於
F2
族群試驗材料係單株栽
培於溫室,可能受到高溫影響定位結果。F2:3
族群具有
2 重複之短行評估,並且在田間環
境下進行性狀調查,因此
QTLs 分析結果對
於分子標誌的選擇較具應用價值。
F2:3
族群
QTLs 定位顯示,影響白堊質面
積的
q-ACE2 與影響白堊質程度的 q-DEC2 均
位於第
2 對染色體 RM6938 上,外表型貢獻
量分別為
9.3%與 9.6%,均非主效 QTLs。而
影響白堊質性狀主效
QTLs,如白堊質率的
q-PGWC12、白堊質面積的 q-ACE3,與白堊
質程度的
q-DEC7b 則各別位於不同染色體
上,以上結果說明白堊質面積與白堊質程度
二者之間的關係密切,其中位於第
7 對染色
體
RM1364 分子標誌之鄰近區域為論在 F2
族
群或在
F2:3
族群之
PGWC、ACE 及 DEC 等
三個性狀均測得相關之
QTLs。
利用分子標誌輔助選拔
23 個優良基因品
系,除
C54 品系外,其他品系的白堊質率及
白堊質程度均小於親本「台農
67 號」。依據
「中華民國國家標準」(CNS 總號 2425,類
號
N1059)之規定,白米外觀品質為 CNS 一
等米者,白粉質粒在
5%以下,符合該項標準
有
C180 與 C184 等 2 個品系;CNS 二等米
者,白粉質粒在
10%以下,符合該項標準有
T132、T136、T309、T415、T430、T509 與
T613 等 7 個品系;CNS 三等米者,白粉質粒
在
15%以下,符合該項標準有 C133、T239、
T477、T593、T607 與 T624 等 6 個品系,總
計有
15 個品系符合 CNS 各級標準,佔全體
優良品系之
65%,顯示利用分子標誌輔助選
拔低白堊質米粒品系確有其效果。
本研究於早世代(F2
世代)即開始利用分
子標誌輔助選拔,可減少傳統育種改良過程
中所需之人力和時間,避免環境變動影響選
拔效果。由於遺傳圖譜建立及田間外表性狀
調查充份配合,故能偵測有關
QTLs 位置,
以利用緊密連鎖之分子標誌完成品系選拔及
純化。期望未來可利用分子標誌輔助選拔之
方式,改善不易進行傳統育種選拔之性狀,
藉此縮短育種年限,提高育種效率。
誌謝
本研究承行政院農業委員會計畫(100 農
科-9.2.1-糧-Z1 及 101 農科-9.2.1-糧-Z1)補助
經費,特此誌謝。
引用文獻
Ebitani T, Y Yamamoto, M Yano, M Funane (2008) Identification of quantitative trait loci for grain appearance using chromosome segment substitution lines in rice. Breed. Res. 10:91–99. He P, SG Li, Q Qian, TQ Ma, JZ Li, WM Wang, Y
Chen, LH Zhu (1999) Genetic analysis of rice grain quality. Theor. Appl. Genet. 98:502–508. Kepiro JL, AM McClung, MH Chen, KM Yeater,
RG Fjellstrom (2008) Mapping QTLs for milling yield and grain characteristics in a tropical japonica long grain cross. J. Cereal Sci. 48:477– 485.
Kobayashi A, B Genliang, Y Shenghai, K Tomita (2007) Detection of quantitative trait loci for white-back kernels under high temperature stress in japonica rice varieties. Breed. Sci. 57:107–116.
Kuo YC , C Liu (1986)Inheritance of Chalkiness of Rice Endosperm. (in Chinese) J. Agric. Res.
China. 35:129–138.
Lander ES, Botstein D (1989) Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics. 121:195–199. Lincoln SE, MJ Daly, ES Lander (1993)
Constructing genetic linkage maps with MAPMAKER/EXP Version 3.0: A Tutorial and
reference manual. 3rd ed. A Whitehead Institute
for Biomedical Research Technical Report. Shi CH, JG Wu, XB Lou, J Zhu, P Wu (2002)
Genetic analysis of transparency and chalkiness area at different filling stages of rice (Oryza sativa L.).Field Crops Res. 76:1–9. Tabata M, H Hirabayashi, Y Takeuchi, I Ando, Y
Lida, R Ohsawa (2007) Mapping of quantitative trait loci for the occurrence of white-back kernels associated with high temperatures during the ripening period of rice (Oryza sativa L.). Breed. Sci. 57:47–52.
Takeda K, K Saito (1983) Heritability of kernel weight and white belly frequency in rice and genetic correlation. Jpn. J. Breed. 33:468–480. Tan YF, YZ Xing, JX Li, SB Yu, CG Xu, QF Zhang
(2000) Genetic bases of appearance quality of rice grains in Shanyou 63, an elite rice hybride.
Theor. Appl .Genet. 101:823–829.
Wada T, T Ogata, M Tsubone, Y Uchimura, Y Matsu (2008) Mapping of QTLs for eating quality and physicochemical properties of the
japonica rice ‘Koshikari’. Breed. Sci. 58:427–435.
Wan XY, JM Wan, JF Weng, L Jiang, JC Bi, CM Wang, HQ Zhai (2005) Stability of QTLs for rice grain dimension and endosperm chalkiness characteristics across eight environments. Theor. Appl. Genet. 110:1334– 1346.
Wang S, CJ Basten, ZB Zeng (2011) Windows QTL Cartographer 2.5, Department of Statistics, North Carolina State University, Raleigh, N.C. Zhou L, L Chen, L Jiang, W Zhang, L Liu, X Liu, Z
Zhao, S Liu, L Zhang, J Wang, J Wan (2009) Fine mapping of the grain chalkiness QTL
qPGWC-7 in rice (Oryza sativa L.). Theor. Appl.
Genet. 118:581–590.