克雷白氏肺炎桿菌CG43中yfiD基因表現的調控
67
0
0
全文
(2) 致謝. 我的碩士生涯就在此告一段落,對於這裡的人、事、物感到些許 不捨,卻必須朝向未來邁進。在這裡不只訓練邏輯、技術、報告與寫 作,挑戰著時間的管理、做事的態度、人們之間的相處溝通、面對緊 張與壓力的應對,也同時考驗身體的耐性、意志的堅定與情緒的掌 握。這看似辛苦又寂寞的歷程其實處處精采充實,讓以前的自己充滿 嚮往與熱情,也成就現在的自己。所以,回顧兩年的種種,除了不捨, 更是充滿感恩的心情。 最感謝的是我的指導教授,彭慧玲老師。她不只授予碩士學生的 訓練指導,更用鼓勵和肯定給予信心,耐心的教導我用平常心面對問 題以及用心去思考解決,還有給我發揮的自由與機會。更重要的是, 因為她的啟發,讓我有了與以往不同的自信和開闊的視野去接受新的 事物與挑戰,對於未來更加期待。 感謝清華大學的張晃猷老師以及張老師實驗室的夥伴們,在這過 程中給予許多建議及協助,指出自己的盲點,使研究方向和目標更明 確。 感謝中興大學的鄧文玲老師,提示我以往缺少的思考方向,讓自 己得以從更廣的角度切入問題並且加以探討。. i .
(3) 感謝交通大學的楊昀良老師,給予研究不同的討論觀點以及導正 論文中的錯誤。 感謝實驗室的夥伴們一路扶持和陪伴,使我在工作之餘還有搞笑 歡樂。親切的靖婷學姐給了初入實驗室的我許多研究方向,用過來人 的經驗解除我從大學生變成研究生的緊張與不安。活潑可愛的好鄰居 盈蓉學姐,是我討論和提問的好對象。除了分享興趣討論休閒以及處 世觀點,也給予許多一針見血的看法。美麗又堅強的靜柔學姐,雖然 面對種種壓力與挑戰,還能樂觀開朗的面對與努力,傾聽與體貼我的 憂慮,是一起打拼的好夥伴。健誠學長的動人歌聲還有新耀學長的熱 血和細心讓大家充滿朝氣,笑聲爽朗的登魁、開朗搞笑的格維和熱衷 研究的朝陽為實驗室增添色彩。嘉怡的善良純真、媺如的認真成熟、 雅淑的聰明美麗、雅雯的可愛美麗大方、顗峰的獨家幽默與細膩、純 珊的溫柔浪漫、志桓的聰明負責、振宇的積極向上、佳瑩的善良可愛、 哲充的努力執著還有朝彥的陽光開朗,都刻印在我腦海裡。承哲、佩 君、偉新、冠維和信文為實驗室注入新血與歡樂。感謝你們體貼我的 健忘,為我打點畢業口試的事務,陪我一起努力、一起搞笑、一起沉 澱、一起暢談理想、吃美食、唱歌、打球、跳舞、體驗新竹之美。因 為你們,所以這裡是個好地方,所以我愛新竹。感謝我的好室友崧甫 與維晉,與我分享每天的笑料和點滴,以及感謝遠在各地的朋友分享. ii .
(4) 你們的一切。 感謝我的父母與家人一路支持相挺,放手讓我在外地闖蕩,體諒 我無法陪在你們身邊。這份碩士學位是我努力的見證,也是我的驕 傲,在此獻予你們! 最後,我將踏上人生旅途的另一階段,在心裡一直給我們祝福, 為我們加油,希望我們都能實現願望!. 秉熹 民國 97 年 8 月 18 日筆於新竹交通大學竹銘館分子調控實驗室. iii .
(5) Contents Abbreviation----------------------------------------------- v Abstract (in Chinese) ------------------------------------ vi Abstract (in English) ------------------------------------- viii Introduction------------------------------------------------ 1 Materials and Methods----------------------------------- 8 Results------------------------------------------------------ 16 Discussion------------------------------------------------- 22 References------------------------------------------------- 27 Tables------------------------------------------------------- 37 Figures----------------------------------------------------- 41 Appendix--------------------------------------------------- 55. . iv .
(6) Abbreviation AR. acid resistance. BCIP. 5-bromo-4-chloro-3-indolyl phosphate. BSA. bovine serum albumin. CFU. colony forming unit. DTT. dithiothreitol. EDTA. ethylenediamine-tetraacetic acid. EMSA. electrophoretic mobility shift assay. Evg. Escherichia virulence gene. HK. histidine kinase. IPTG. isopropyl-β-D-thiogalactopyranoside. kvg. Klebsiella virulence gene. kvh. Klebsiella virulence gene homologue. LB. Luria-Bertani medium. MES. morpholineethanesulfonic acid. NBT. nitro blue tetrazolium. PEG. polyethylene glycol. PNK. T4 polynucleotide kinase. PCR. polymerase chain reaction. RBS. ribosome binding sequence. Rcs. regulator of capsular polysaccharide. RR. response regulator. SDS. sodium dodecyl sulfate. SDS-PAGE. SDS polyacrylamide gel electrophoresis. v .
(7) 中文摘要 我們利用蛋白質體分析方法探討高致病性克雷白氏肺炎桿菌 CG43 中 KvhAS 雙分子系統的調控角色時發現:在弱酸培養後的細菌 CG43S3U9451 的二維電泳膠片中,有一蛋白質點的強度較 CG43S3 U9451kvhAS-膠片中的相對蛋白質點高出許多,經截取、純化此蛋白, 再經質譜分析確認此蛋白質點為 YfiD。接著以 LacZ 為報導基因分析 yfiD 啟動子活性,結果顯示只有在微氧培養的狀態下,yfiD 啟動子在 ΔkvhA 突變株中的活性減少;有趣的是,ΔrpoS 突變株之 yfiD 啟動子 活性也相對減少;而在截短 yfiD 啟動子上的一個 RpoS 結合序列後, 受 RpoS 基因缺損影響的現象即消失。我也利用膠體遲滯電泳分析, 證實 KvhA 直接黏附 yfiD 啟動子而達到正向轉錄調控的作用。進一 步,我還選殖了 yfiD 基因並將 yfiD 殖入大腸桿菌表現系統來大量表 現 YfiD 蛋白,並將蛋白純化後注射 Balb/c 老鼠來取得多株抗體。以 西方墨點法分析的結果顯示 kvhA 的缺損造成 YfiD 蛋白量的下降,而 轉型入 KvhA 表現質體時,YfiD 蛋白表現量即回升;另外,以帶有 定點突變的 KvhAD52A (持續去磷酸化)或 KvhAD52E (持續磷酸化)的 質體轉入 kvhA 的缺損株,我發現 KvhA 是否被磷酸化會影響 yfiD 基 因的表現調控。yfiD 基因缺損株在有氧條件之下,對於酸處理後的感 受性提高,而 yfiD 缺損對細菌的生長並沒有顯著的影響。最後,我 vi .
(8) 構築了 YfiDG102A 點突變株,證實 YfiD 的酵素活性參與其酸性逆境表 現型的改變。. vii .
(9) Abstract. A proteomic approach has been employed to unravel the regulatory role of the two-component system KvhAS in the highly virulent strain K. pneumoniae CG43. In comparing the two dimensional protein profiles derived from mild acid-growth bacteria, a protein spot highly expressed in the gel of K. pneumoniae CG43S3U9451 but not in that of CG43S3U9451kvhAS- mutant was isolated and later identified as YfiD by mass spectrum analysis. Promoter activity measurement using LacZ as reporter revealed a reduction of PyfiD activity in KvhA- mutant only under a microaerobic environment. Interestingly, deletion of rpoS also decreased yfiD expression. The rpoS deletion effect was no more observed while the putative RpoS-binding element was truncated. Furthermore, EMSA demonstrated a direct binding of KvhA to PyfiD DNA which supporting a positive transcriptional regulation on the expression of yfiD. In the meantime, yfiD encoding gene was cloned and overexpressed in E. coli, and the recombinant YfiD purified to homogeneity to inject Balb/c mice for polyclonal antibody preparation. The analysis of western blotting hybridization against the YfiD polyclonal antibody indicated that the deletion of kvhA reduced the expression of YfiD. The introduction of the plasmid carrying KvhA into the kvhA deletion mutant appeared to enhance the expression of yfiD. The transformation of the plasmid carrying KvhAD52A (constitutively dephosphorylated),. or. KvhAD52E. (constitutively. phosphorylated). confirmed the phosphorylation of KvhA is required for the regulation of viii .
(10) yfiD expression. Finally, deletion of yfiD was found to increase the bacterial susceptibility to acid treatment under aerobic condition while no effect on the bacterial growth. The YfiDG102A was generated and the involvement of its enzymatic activity in the acid-shock resistance was also demonstrated.. ix .
(11) Introduction. Klebsiella pneumoniae is a gram-negative bacillus of the family Enterobacteriaceae with a world-wide distribution and is an important cause of human disease resulting in significant morbidity and mortality (54).. On. the. basis. of. Voges-Proskauer. (VP). reaction. (38),. Enterobacteriaceae are divided into two groups. K. pneumoniae excretes acetoin (3-hydroxy-2-butanone) into outer cellular environments whereas E. coli does not. Acetoin is an important physiological metabolic product excreted by various microorganisms when they grow in environmental niche containing glucose or other fermentable carbon sources (24, 43). This neutral product allows the bacteria to ferment large amounts of carbohydrates without self-inhibition due to the overacidification of intercellular environments (16, 70). Acetoin can also be reutilized when glucose has been depleted (21). Moreover, owing to the reversible transformation between acetoin and 2,3-butanediol coupled with the NAD/NADH conversion, the pathway has been regarded as participating in regulation of the NAD/NADH ratio in bacteria (29). K. pneumoniae is also a commonly reported nosocomial pathogen, which often causes pneumonia, septicemia, bacteremia, suppurative lesion, wound infection, burn infection, and urinary or respiratory tract infections in chronic alcoholics and immunocompromised patients (54). In Taiwan, K. pneumoniae has been found to be closely associated with liver abscess leading to serious complications among diabetic patients (10,. 1 .
(12) 51, 66). There are five virulence factors identified to be involved in K. pneumoniae infections, which include capsular polysaccharides (23), lipopolysaccharides (64), adhesins (63), iron-acquisition systems (53), and antibiotic resistance (59). In addition to these virulence factors, very little is known about the other factors that may participate in the pathogenesis.. The. increasing. prevalence. of. extended. spectrum. β-lactamase, which leads to ESBL-producing K pneumoniae, has promoted the development of new drugs to overcome drug-resistance difficulties (47). Much of the success of infections depends on the bacterial ability to survive and thrive in adverse environments. K. pneumoniae has evolved mechanisms to cope with diverse conditions from human guts to soil (54). These different environments present ever changing challenges such as oxygen availability, pH shift, and osmolarity change and the adaptation ability is ultimately rooted in altering patterns of gene expression. Two-component systems (2CSs) are widely distributed in gram positive and negative bacteria, which play diverse regulatory roles in bacterial physiology, such as stress resistance, virulence, energy metabolism, nutrient acquisition, and quorum sensing (9, 18, 52, 58, 62). 2CSs consist of a transmembrane sensor histidine kinase (HK) and a cytosolic response regulator (RR). Upon sensing input signals, the sensor HK catalyzes an autophosphorylation which transfers a phosphate from ATP to a conserved histidine residue on the transmitter domain. The phosphate group is subsequently transferred to a specific aspartate residue on the receiver domain of the cognate RR. The phosphorylated RR is thus. 2 .
(13) activated and begins to regulate the expression of downstream genes (4). Recently, drug development targeting 2CSs to intervene the signal transduction involved in microbial infections has been established (49). The specific aspartate residue in RcsB, the RR of RcsCDB signal transduction system, has been demonstrated by site-directed mutagenesis showing its pivotal role in the His-Asp phosphorelay for the regulation of downstream genes (6). Since signal transduction in mammals occurs by a different mechanism, inhibition of histidine kinases provides another potential target for antimicrobial agents (46). Besides 2CSs which are responsible for the regulation to adapt to environmental changes, RpoS (σs), an alternative sigma subunit of RNA polymerase, has been identified to regulate bacterial general stress response (68). RpoS controls more than 80 genes (16, 30). Some of the gene expression has been found to be involved in the acid resistance mechanism (8, 67), which is important for enterobacterial infection. For gastrointestinal pathogens such as K. pneumoniae and E. coli, penetrating the barrier of gastric acid is crucial for the colonization and infection. This requires either a large number of organisms or a powerful acid resistance (AR) mechanism that allows small numbers of bacteria to survive until the stomach empties its contents into the more alkaline intestine. Four AR systems have been identified in bacteria to resist acidic environments. AR system 1 is repressed by glucose evident in stationary-phase cells, and protects cells in minimal medium. It has been shown in E. coli that RpoS and cyclic AMP receptor protein (CRP) play a role in control of this pathway (8, 67). The other systems are not. 3 .
(14) repressed by glucose and require the addition of glutamic acid (AR system 2), arginine (AR system 3), or lysine (AR system 4) in pH 2.5 acid challenge medium (8, 26, 42). These amino acid-dependent systems utilize matched decarboxylases and antiporters to protect the cell. The decarboxylases are glutamic acid decarboxylase isozymes GadA and GadB for system 2, the arginine decarboxylase AdiA for system 3, and presumably, the inducible lysine decarboxylase CadA for system 4 (8, 14, 22, 26). However, whether these AR systems exist in K. pneumoniae remains to be investigated. We have previously identified a 2CS, KvhAS, by PCR-based cloning from K. pneumoniae CG43 (40). The analysis using dot-blotting hybridization revealed that kvhAS is presented in all strains collected in the laboratory, suggesting an important role of the 2CS in K. pneumoniae (40). According to the sequence analysis, kvhAS shows high homology to evgAS (Escherichia virulence gene). KvhA shares 66.7% protein identity with EvgA, while KvhS shares 48% protein identity with EvgS. EvgAS has been demonstrated to regulate multi-drug resistance (48), acid resistance (44) and heat-resistance (12). Therefore, a similar role of KvhAS as EvgAS was proposed. The subsequent studies showed that the overexpression of KvhA altered the susceptibility to fosfomycin, and deletion of kvhS reduced kvhAS promoter activity in the presence of 300 mM NaCl in M9 medium (41). A heat shock (44℃) activated promoter activity using GalU as reporter has been demonstrated (25). Moreover, an interacting regulation of KvhAS with KvgAS and KvhR for the expressions of cps gene cluster was also demonstrated (40), but the. 4 .
(15) molecular mechanism of the regulation remains unknown. A proteomic approach has been employed to uncover if KvhAS is involved in acid resistance in K. pneumoniae (74). In comparison of the two dimensional protein profiles that were extracted from the bacteria incubated in mild acid (pH 5.5), a protein spot was found highly expressed in K. pneumoniae CG43S3U9451 but not in that of CG43S3U9451ΔkvhAS (Appendix I). The protein was then isolated, subjected to trypsin digestion and mass spectrum analysis and identified as YfiD. YfiD is a homolog to the C-terminal region of pyruvate formate-lyase (PFL), the product of pflB gene. For a functional activity, YfiD and PFL have to be post-translationally activated in anaerobic environment by PFL activase (encoded by pflA) through inserting a radical to them. The radical which is crucial for the catalytic activity is susceptible to oxidative cleavage thereafter leading to inactivation of the enzyme. The inserted radical in PFL can be quenched by PFL deactivase AdhE microaerobically to prevent oxidative cleavage, however, YfiD does not respond to the deactivated AdhE (69). It has also been shown that YfiD acts as a “spare part” by providing an active glycyl radical domain to restore activity of the oxygenolytically cleaved PFL (65) for balance of the carbon flux in microaerobic environment. Upon the activation, a glycyl radical forms around a glycine residue (Glycine 102) contained in the highly conserved pentapeptide motif (RVXGY) of YfiD (Appendix III). The activated YfiD, via catalyzing the reaction to convert pyruvate into acetyl-coenzyme A, is able to reduce the accumulation of 5 .
(16) acidic metabolic end product thereby to modulate the intracellular pH (69, 73). A phosphorylated form of YfiD has been demonstrated in vitro in stagnant growth of E. coli L form NC-7 (19), suggesting a phosphorylation regulation for the YfiD activity. The expression of E. coli yfiD has been shown to be under the regulation of two well-known global regulators FNR (fumarate-nitrate reduction regulator) and ArcA (aerobic respiration control regulator) (69), and a pyruvate sensor PdhR (55) (Appendix II). In oxygen deprived condition, FNR, which contains a Fe-S cluster as a redox sensor, upregulated the yfiD expression at FNR element I (high affinity FNR binding element). While increased expression of FNR under low oxygen level allowed it to occupy the FNR element II (low affinity FNR binding element) to prevent from the yfiD expression. In contrast, ArcA competes with FNR for binding the FNR element II under the anaerobic condition to increase the expression of yfiD. Consequently, the yfiD predominantly expressed in microaerobic environment (20). While PdhR was shown to mediate repression of yfiD indicating a link to pyruvate/central metabolism. Previous reports have also indicated that intracellular YfiD levels are enhanced by growth in the presence of pyruvate (5). YfiD protein has been identified in a wide range of Gram-negative and positive bacteria including Serratia sp., S. liquefaciens and Haemophilus infuenzae, as well as in T4 and T5 bacteriophages (69). Sequence analysis revealed that K. pneumoniae yfiD is located in the srmB-ung intergenic region, which is similar to that of E. coli and H. influenzae yfiD gene. However, the T4 yfiD is located next to the 6 .
(17) thymidine kinase gene while the T5 yfiD is next to a tRNA gene cluster. In Serratia, the yfiD gene is associated with the phlA gene encoding an extracellular phospholipase A1 (69). In this study, we investigate if KvhAS plays a regulatory role in the expression of yfiD and the functional role of YfiD in K. pneumoniae CG43. Firstly, PyfiD activity measurement has been used to investigate the regulation of KvhA on yfiD expression at transcriptional level. In the meantime, rpoS deletion mutant has been utilized to inspect if RpoS is involved in yfiD regulation. Secondly, EMSA has been employed to demonstrate the specific binding of KvhA to yfiD promoter. Thirdly, anti-YfiD polyclonal antibody, YfiD expression plasmid and yfiD deletion mutant have been generated to demonstrate if the decreased YfiD expression in K. pneumoniae CG43S3ΔkvhA mutant could be restored by introducing a KvhA expression plasmid. The involvement of KvhAS phosphorelay in the regulation of yfiD expression is also investigated. Finally, the deletion effect of yfiD on bacterial growth in acidic environment and the acid stress response were studied and shown.. 7 .
(18) Materials and Methods Bacterial strains, plasmids and growth conditions The bacterial strains and plasmids used in this study are listed in Table 1 and 2. K. pneumoniae CG43S3 (36), CG43S3kvhA-, CG43S3rpoS-, CG43S3yfiD-,. CG43S3Z01,. CG43S3Z01kvhA-,. CG43S3Z01rpoS-,. CG43S3U9451 and CG43S3U9451kvhA- are derivative strains of CG43 which is a highly virulent strain of K2 serotype with an LD50 of 10 cfu in laboratory mouse (11). Tryptone, yeast extract, and agar were purchased from Difco Laboratories (Detroit, Mi). Unless otherwise indicated, bacterial cultures were grown in Luria-Bertani broth (LB) supplemented with appropriate antibiotics at 37°C. Microaerobic growth was carried out essentially as described by Croinin (13).. Recombinant DNA technique The recombinant DNA experiment was carried out by standard procedures (30). Primers used in PCR are listed in Table 3. Plasmid DNA was prepared by VIOGENE Miniprep kit. Restriction endonucleases and DNA modifying enzymes were purchased from New England Biolab (Beverly, MA), MBI (Fermentas, Hanover, MD), or Roche Molecular Biochemicals (Mannheim, Germany), and were used according to the recommendations of the suppliers.. 8 .
(19) Promoter activity assessment The putative promoter of yfiD and the derived truncation regions were PCR amplified from K. pneumoniae CG43S3 by the designed primers yfiD01, yfiD02, yfiD05, yfiD07, yfiD08, and yfiD09 (Table 2) and subcloned into placZ15 (40) to fuse them with the promoterless lacZ reporter gene. One-hundredth overnight culture of the bacteria carrying each of the plasmids were subcultured and freshly grown in LB medium in microaerobic or aerobic condition for 4 h. β-galactosidase activity assay was carried out essentially as described by Miller (47). The data presented were derived from a single experiment which was representative of at least three independent experiments. Every sample was assayed in triplicate, and the average activity and standard deviation were presented.. Construction of yfiD mutant Two 1.1 kb DNA fragments flanking the yfiD gene were PCR amplified from. K.. pneumoniae. CG43S3. using. yfiDm06/yfiDm07. and. yfiDm08/yfiDm09. The generated DNA fragments were ligated and subcloned into a suicide vector pKAS46 (61). The resulting plasmid pYm2KAS, containing a 308-bp deletion in yfiD, was transformed into E. coli S17-1 λ-pir by electroporation. The E. coli S17-1λ-pir and K. pneumoniae CG43S3 were cultured to mid-logarithmical phase and mixed at a ratio of 1:2, and pelleted by centrifugation. The mating mixture was then suspended in 100 μl of saline and spread onto a 9 .
(20) sterilized nitrocellulose membrane on a prewarmed LB plate. After overnight incubation at 37 ℃ , the cells were washed off from the membrane with 5 ml of LB. The transconjugants were selected by plating with a 105-fold dilution of the culture on M9 minimal medium containing 30 μg/ml kanamycin and 100 μg/ml ampicillin for the integration of the suicide vector. Subsequently, several of the transconjugants were cultured in LB without selection to late logarithmic phase followed by selection on LB plates containing 500 μg/ml streptomycin for the loss of vector plasmid. The deletion was further demonstrated by PCR analysis.. Expression and purification of the histidine-tagged YfiD The recombinant plasmid pY34B containing the His6-tagged YfiD was constructed and the plasmid was transformed into E. coli BL21(DE3). The transformants carrying pY34B were cultured in LB to log phase, and expression of the His6-YfiD recombinant protein was induced with 0.1 mM IPTG for 1 h. The IPTG-induced bacterial cells were harvested by centrifugation at 4℃, 3300 Xg for 15 min, resuspended in 1 X binding buffer (5 mM imidazole, 0.5 M NaCl and 20 mM Tris-HCl pH 7.9), and then disrupted by sonication. Subsequently, the cell lysate was centrifuged at 20,400 Xg for 25 min at 4℃, and the supernatant was applied to a column containing His6-Bind Resin (Novagen, Madison, WI). After washing the column with 1 X binding buffer and then 1 X wash buffer (60 mM imidazole, 0.5 M NaCl, and 20 mM Tris-HCl pH7.9), the. 10 .
(21) His6-YfiD protein was eluted with 1 X elution buffer (1 M imidazloe, 0.5 M NaCl, and 20 mM Tris-HCl pH 7.9). The eluted His6-YfiD protein was subjected to dialysis with 1 X phosphate-buffered saline (PBS) by CelluSep Dialysis Membranes purchased from MFPI (Seguin, TX). Finally, the dialyzed His6-YfiD protein was concentrated by covering with polyethylene glycol 20,000 on dialysis membranes.. Preparation of YfiD antisera To obtain anti-YfiD antibody, 5 μg of the purified YfiD was injected intraperitoneally to five-week old female BALB/c mice which were purchased from the animal center at National Taiwan University. Ten days later, the mice were immunized again with 5 μg of the protein and the antisera obtained by intra-cardiac puncture.. Western blotting analysis Total cellular lysates from K. pneumoniae CG43S3 grown overnight in LB medium were resolved by SDS-PAGE. The proteins were then electrophoretically transferred onto a PVDF membrane (Millipore, Billerica, MA, USA). After incubation with 5% skim milk at room temperature overnight, the membrane was washed 3 times with 1 X phosphate-buffered saline (PBS). Subsequently, the membrane was incubated at room temperature for 2 h with diluted anti-YfiD serum. After washes three times with 1 X PBS, a 3000-fold diluted alkaline. 11 .
(22) phosphatase-conjugated anti-mouse immunoglobulin G was added and the incubation continued for one more hour. The blot was again washed and the bound antibodies were detected by using the chromogenic reagent BCIP (5-bromo-4-chloro-3-indolyl phosphate) and NBT (Nitro blue tetrazolium).. Determination of protein intensity The protein intensity was determined by analyzing the Western blot image using Gel-Pro AnalyzerTM (Media Cybernetics, Inc.) according to the instruction.. Generation of a kvhA expression plasmid A DNA fragment containing the kvhA coding region and a RBS sequence was PCR amplified from K. pneumoniae CG43S3 using its primers kvhAc01/ kvhAc02 and cloned into pCPP45. The resulting plasmid was designated as pKvhAcPP.. Generation of yfiD expression plasmids A DNA fragment containing the yfiD coding region and a RBS sequence was PCR amplified from K. pneumoniae CG43S3 using primers yfiDc01/yfiDc02 and cloned into pRK415 (32) and pCPP45. The resulting plasmids were designated as pYcRK and pYcPP respectively.. 12 .
(23) Construction of KvhAD52A, KvhAD52E and YfiDG102A expression plasmids by site-directed mutagenesis Site-directed mutagenesis in KvhA and YfiD was carried out essentially as described by QuikChange® Site-Directed Mutagenesis Kit (Stratagene, USA). Asp52 in KvhA is responsible for receiving an orthophosphate, which was changed into alanine and glutamic acid residues in pKvhAcPP by primer sets hAd52a-F3/hAd52a-R3 and hAd52e-F2/hAd52e-R2, and the resulting plasmids were designated as pHAcAPP and pHAcEPP respectively. Meanwhile, gly102 in the YfiD active site was changed into an alanine in pYcPP by primer set yfiDg102a-F3/ yfiDg102a-R3, and the resulting plasmid was named pYc102PP.. Electrophoretic mobility shift assay (EMSA) The probes used in the electrophoretic mobility shift assay were the restriction fragments of the putative yfiD promoter-containing region including PyfiD-2 and PyfiD-5, putative kvhA promoter-containing region and kvhA coding sequence. The restriction fragments were then separated on a 2% agarose gel and recovered. They were 5’ end-labelled with [γ-32P]-ATP (25 mCi/ml) using T4 polynucleotide kinase. The His6-KvhA was phosphorylated by incubating in 1 X gel shift binding buffer [20 mM Tris-HCl, pH 8.0, 0.4 mM MgCl2, 50 mM KCl, 1 mM CaCl2, 1 mM DTT, and 10% glycerol] (Karimova et al. 1996) in the presence of 50 mM. 13 .
(24) acetylphosphate and 1μg of acetylated bovine serum albumin for 30 min at 25℃. And then the radioactively-labeled DNA fragments was added and incubated at 37℃ for 30 min. The samples were applied to 5% non-denaturing polyacrylamide (acrylamide: bis-acrylamide, 19: 1) gels that were pre-electrophoresed at a constant voltage of 100 V for 30 min at 4℃ before the samples were loaded. Electrophoresis was performed at 25 mA for 40 to 80 min at 4℃. The gels were read by InstantImagerTM (Packard Instrument Company) or visualized by autoradiography.. Bacterial growth analysis One-hundredth overnight grown bacteria were inoculated to LB or pH 5.5 LBMES medium (1, 56) and incubated at 37°C in aerobic or microaerobic condition. The optical density at the wavelength of 600 nm (OD600) was measured every hour, which was representative of at least three independent experiments.. Acid survival assessment One-hundredth overnight cultured bacteria were inoculated to pH 5.5 LBMES medium and grown at 37°C aerobically to OD600 of about 0.7. Aliquots of the bacteria were then collected by centrifugation and resuspended in pH 3 LB medium, and subjected to 37°C incubation aerobically for 35 min. CFU of the bacteria before and after the stress treatment were counted, and the survival rate was determined by the ratio 14 .
(25) of the CFU after to before the acid stress treatment. The representative data of at least three independent experiments were presented. Every sample was assayed in triplicate, and the average activity and standard deviation were presented.. 15 .
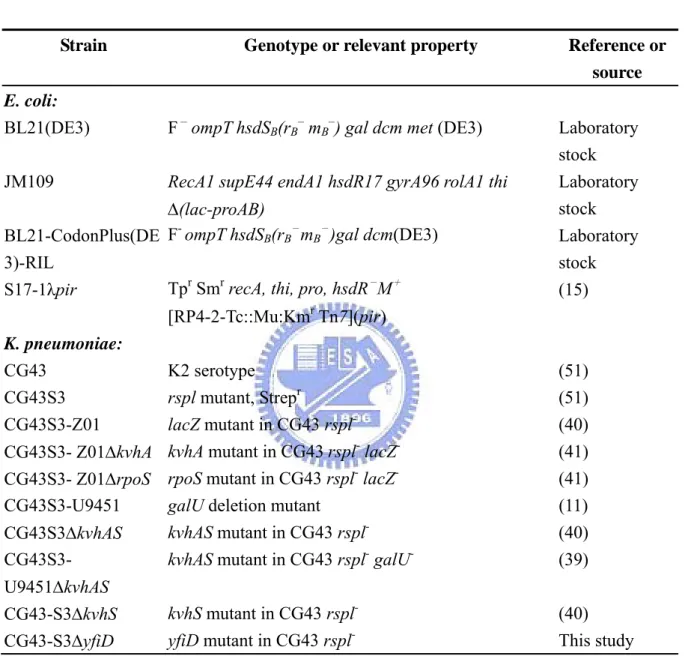
(26) Results Sequence analysis of the yfiD putative promoter revealed conserved binding sequences of FNR, ArcA, RpoS and EvgA DNA sequence upstream to the yfiD coding region was analyzed by BPROM of Softberry (http://www.softberry.com/) and -10 and -35 boxes were identified. As shown in Fig. 1, FNR and ArcA binding elements (69), a putative EvgA binding box (31), and two putative RpoS elements (34, 37) were also found.. Deletion of kvhA or rpoS reduced PyfiD activity As shown in Fig. 2A, the respective LacZ reporter plasmids containing the putative promoters PyfiD-1, PyfiD-2, PyfiD-3 of yfiD were constructed and transformed into K. pneumoniae CG43Z01 and derived bacteria. The bacteria were cultivated in aerobic or microaerobic conditions in LB at 37 ℃ for 4 hours and then subjected to activity assessment. In aerobic. condition, the activity of PyfiD-1, PyfiD-2, PyfiD-3 in Z01, Z01ΔkvhA or Z01ΔrpoS showed no apparent difference (Fig. 2C). Under microaerobic condition, PyfiD-1 activity was slightly reduced in Z01ΔkvhA. The kvhA deletion appeared to have more profound effect on the activity of PyfiD-2, suggesting the presence of KvhA binding site after the truncation from PyfiD-1 to PyfiD-2, and the negative effects of rpoS deletion apparently reduced the activity of PyfiD-1 and PyfiD-2. As shown in. 16 .
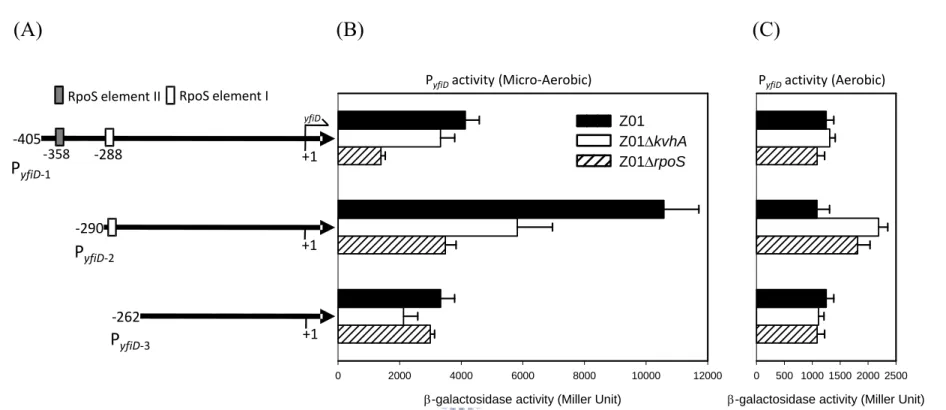
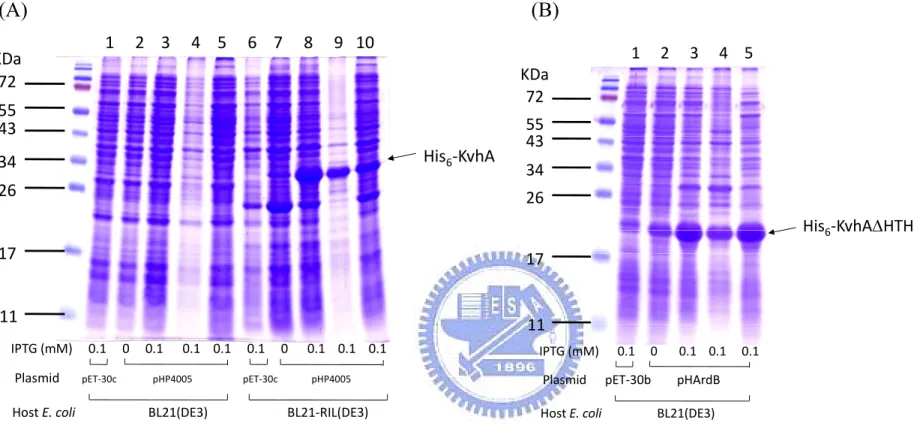
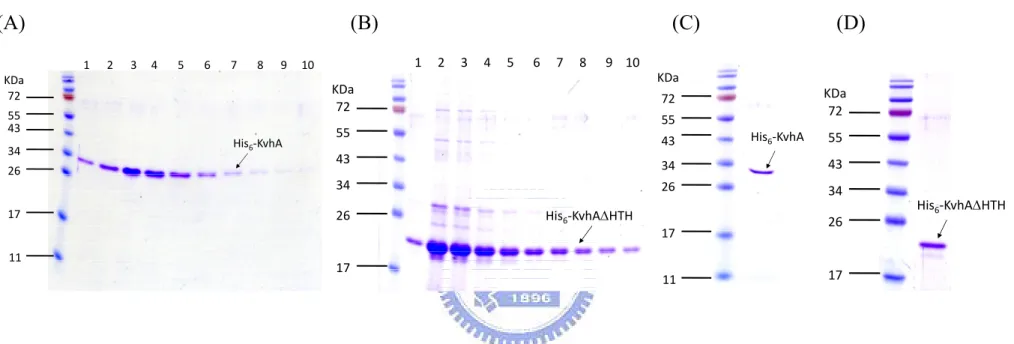
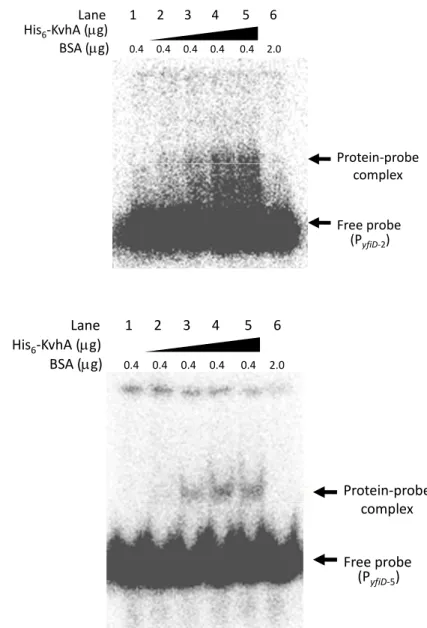
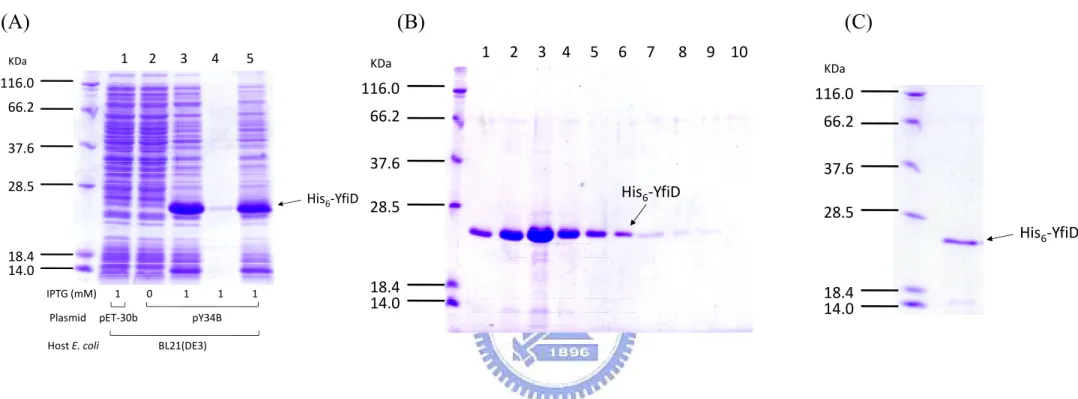
(27) Fig. 2B, the negative effect of rpoS deleltion was alleviated in the expression of PyfiD-3 that contains no RpoS binding element. This implied the RpoS element I as shown in Fig. 2A is the major regulatory sequence for the RpoS dependent expression.. Preparation of His6-KvhA and His6-KvhAΔHTH proteins His6-KvhA expression plasmid pHP4005 (40) and His6-KvhAΔHTH expression. plasmid. pHArdB. were. introduced. into. E.. coli. BL21-CodonPlus(DE3)-RIL and BL21(ED3), respectively. One-tenth of the overnight bacterial cultures were freshly grown in LB at 37℃ for 2 h, then 0.1 mM IPTG was added for another 5 h incubation followed by SDS-PAGE resolution of the total proteins. Overexpression of the proteins with expected molecular mass of KvhA (Fig. 3A) and the truncation form of KvhA (Fig. 3B) could be observed.. After disruption. by sonication, large amounts of His6-KvhA and His6-KvhAΔHTH proteins were found in the supernatant fraction (Figure 3A, B). After purification by His6-Bind Resin column, the collected fractions (Figure 4A, B) were subjected to dialysis and concentration. The purified His6-KvhA and His6-KvhAΔHTH proteins were then confirm by SDS-PAGE (Figure 4C, D).. The phosphorylated His6-KvhA directly bound to PyfiD-2 and PyfiD-5 DNA fragments. 17 .
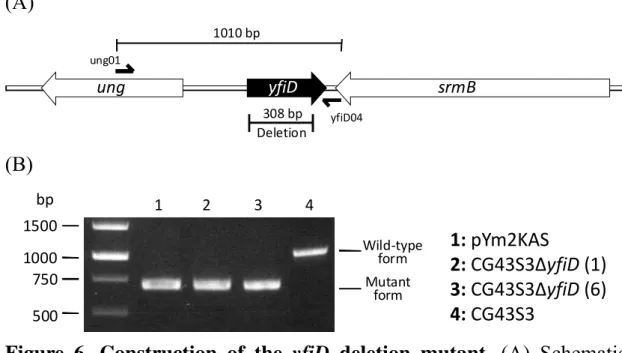
(28) The response regulator, KvhA, like many other response regulator of the two-component system, possesses a helix-turn-helix motif at the C-terminus to bind to specific DNA for its regulatory function. In order to demonstrate direct binding of KvhA to PyfiD-2 (containing a putative EvgA binding box shown in Fig. 1 and PyfiD-5 (Fig. 1), EMSA was carried out. As shown in Fig.5, the recombinant protein His6-KvhA directly bound to PyfiD-2 (Fig.5A) and PyfiD-5 (Fig.5B) DNA fragments. While the presence of excess BSA, the retarded DNA was no more detected, indicating the DNA shift was not resulted from the BSA addition.. Construction of yfiD deletion mutant To construct an yfiD deletion mutant, the DNA fragments flanking the deleted region were amplified by PCR and cloned into the suicide vector, pKAS46 (61). The resulting plasmid pYm2KAS was introduced into K. pneumoniae CG43S3 by conjugation. PCR using the primer set ung01/yfiD04 was employed to confirm the deletion of the resulting mutant (Figure 6).. Expression and purification of His6-YfiD protein The plasmid pY34B, a derivative of pET-30b containing the yfiD coding sequence, was transformed into E. coli BL21(DE3) for protein expression. As shown in Fig. 7A, overexpression of YfiD could be observed after 5 h induction with IPTG. After disruption by sonication, a large amount of. 18 .
(29) His6-YfiD protein was found in the supernatant fraction (Figure 7A). After purification by His6-Bind Resin column, the collected fractions (Figure 7B) were subjected to dialysis and concentration. The purified His6-YfiD was then confirmed by SDS-PAGE (Figure 7C).. YfiD protein content was reduced in ΔkvhA mutant strain One-hundred dilution of the overnight grown bacteria including K. pneumoniae CG43S3, CG43S3ΔkvhA, CG43S3ΔyfiD, CG43S3ΔkvhA [pHAm] and CG43S3ΔyfiD [pYcRK], were refreshly and aerobically cultured in LBMES (pH 5.5) or M9MES (pH 5.5) at 37℃ for 6 h. Total protein was then harvested and the concentration was determined as described (7). Five micrograms of each sample was incubated at 100℃ for 10 min before loaded onto SDS-polyacrylamide gel. Western-blot analysis using anti-YfiD polyclonal antibody was then performed. As shown in Fig. 8A, in comparing with that of the wild type bacteria, a reduced YfiD protein content was observed for the total protein extracts from the kvhA deletion mutant grown in LBMES (Fig. 8A) or M9MES (Fig. 8B).. KvhAS phosphorelay is required for the regulation of YfiD Expression Upon. sensing. extracellular. signals,. 19 . a. sensor. protein. is.
(30) auto-phosphorylated, the orthophosphate then transferred to the cognate response regulator (RR), and finally the activated RR regulates the expression of. downstream genes (4). To answer whether the. phosphorylation of KvhA is essential for yfiD regulation, two mutant forms KvhAD52A (constitutively dephosphorylated) and KvhAD52E (constitutively. phosphorylated). were. generated. by. site-directed. mutagenesis. As shown in Fig. 9, the reduced expression of YfiD in ΔkvhA strain was slightly restored by introducing a KvhA expression plasmid pKvhAcPP, while introducing pHAcAPP (expressing KvhAD52A) did not appear to increase YfiD content. The YfiD protein content was apparently increased by introducing with pHAcEPP (expressing KvhAD52E), suggesting the KvhAS phosphorelay is required for the regulation of YfiD expression.. Deletion of yfiD had no apparent effect on growth Overnight cultures of K. pneumoniae CG43S3 or CG43S3ΔyfiD were refreshly grown in LB (pH 7.0) or LBMES (pH 5.5) at 37℃ under aerobic and microaerobic conditions, and the OD600 was measured each hour. As shown in Fig. 10, no obvious effect of yfiD deletion on the bacterial growth was observed.. YfiD affected the acid-shock resistance As shown in Fig. 11A, the deletion of yfiD apparently reduced the. 20 .
(31) survival of K. pneumoniae CG43S3 while the bacteria in LBMES (pH 5.5) changed to pH 3.0 medium. The deletion effect could be complemented by introducing into the mutant with the YfiD expression plasmid pYcRK. As shown in Fig. 11B, the survival analysis of CG43S3ΔyfiD [pYcRK] showing no full restoration may be resulted from the loss of pYcRK plasmid. Furthermore, the introduction of pRK415 into CG43S3 led to apparent decrease of the survival. An yfiD expression plasmid pYcPP contain yfiD coding gene and its RBS was thus cloned into pCPP45 (Dr. David Bauer at Cornell University), carrying the partition region from RK2 plasmid and hence leading to high stability, to resolve the problem. As shown in Fig. 11(C), the survival rate still remained low when introducing pCPP45 into CG43S3. Although the vast majority of the studies on YfiD focuses on the microaerobic regulation and activation, YfiD was found to be phosphorylated in vitro in stagnant growth of E. coli L form NC-7 (19), suggesting an functional role of YfiD under aerobic environment. Sequence analysis has shown that glycine 102 (g102) is contained in the active site to execute the YfiD enzymatic function (57, 69), pYc102PP containing an YfiDG102A coding region and an RBS sequence has thus been generated to determine if YfiD enzyme activity is involved in the regulation of the acid stress response. As shown in Fig. 11(C), the survival rate of CG43S3ΔyfiD [pYc102PP] apparently decreased comparing to that of CG43S3ΔyfiD [pYcPP], suggesting an involvement of YfiD enzyme activity in phenotypic alteration.. 21 .
(32) Discussion Two-component signal transduction systems, which are widely distributed in gram positive and negative bacteria to sense extracellular signals, render bacteria to respond environmental changes and thus to survive and thrive in the environments or hosts. KvhAS has previously been identified and found to regulate the synthesis of CPS (40) , which is a crucial virulence factor for K. pneumoniae infection (23). We have also shown that the overexpression of KvhA resulted in an alteration of bacterial susceptibility to a small number of antibiotics; the deletion of kvhS reduced kvhAS promoter activity in the presence of 300 mM NaCl (41), and 44℃-heat shock treatment increased kvhAS promoter activity (25). Sequence analysis revealed that KvhAS has high sequence homology to EvgAS that has been demonstrated to regulate multi-drug resistance (48), heat-resistance (12) and acid resistance (44) in E. coli. A similar role was hence proposed. Herein we show a positive role of KvhAS on the expression of YfiD by 2D-PAGE comparative analysis of the. proteome. patterns. of. K.. pneumoniae. CG43U9435. and. CG43U9435ΔkvhA. Upon analyzing the regulation in aerobic culture, the kvhA deletion showed no apparent effect on yfiD expression in mild acid (pH 5.5) condition (74). The activity measurement was thus carried out under microaerobic condition which has been shown to be optimal for yfiD expression E. coli (69). Under the microaerobic condition, both KvhA and RpoS were found to play positive roles in the control of the. 22 .
(33) expression of yfiD. Activity analysis of PyfiD-1, PyfiD-2 and PyfiD-3 containing different sizes of the yfiD promoter suggesting RpoS element I is for RpoS binding to regulate yfiD expression. Unknown repressive element may be located near RpoS element II since an apparent increase of PyfiD-2 activity. While the reduced activity of PyfiD-3 suggested unknown up-regulatory element exists around RpoS element I. Nevertheless, further work have to be carried out for an insight view of the regulation of RpoS on yfiD expression. Interestingly, the previous study has shown that RpoS negatively regulates kvhAS expression (41). Herein a model of RpoS-KvhAS-YfiD regulatory pathway is proposed. As shown in Fig. 12, we speculate that the RpoS-dependent regulation is much stronger than KvhAS for the expression of yfiD. Acetate stress simultaneously induced the expression of YfiD and RpoS growth phase regulon in E. coli W3110 (33), also supporting a role of RpoS in yfiD regulation. Nevertheless, a regulatory role of KvhA in yfiD expression at transcriptional level could be supported by EMSA. The putative EvgA box in PyfiD-2 was not required for the binding by the recombinant KvhA, suggesting a different regulatory pattern of KvhAS from that of EvgAS. The non-identical residues in the highly homologous (63.8% sequence identity) DNA binding domain possibly contribute to the different DNA binding specificity. By comparing YfiD content of mild acid-grown bacteria, it was shown that ΔkvhA mutant strain had a reduced YfiD protein expression. However, kvhA complement plasmid using pHAm (41) carrying a kvhA 23 .
(34) coding region did not increase the expression of YfiD protein, possibly due to low expression of KvhA. Introducing the plasmid pKvhAcPP with kvhA coding gene and RBS into ΔkvhA mutant showed a small increase of YfiD expression. The small increase may be due to the interference of some non-phosphorylated KvhA protein and thus no fully-complementing effect was observed. The possibility was further confirmed by transformation of pHAcAPP or pHAcEPP to the kvhA mutant. The substitution of the receiver residue by glutamate KvhAD52E, which was able to mimic a continuously phosphorylated status of KvhA, led to apparent increase of YfiD expression. Acid-resistant role was proposed since YfiD was demonstrated to reduce acidic-metabolite accumulation in cytosol under microaerobic environment (69). In addition, expression of yfiD has been found to be activated by acid either in aerobic and anaerobic environments (5). Using acid-shock resistance assay, we have also shown an involvement of yfiD at aerobic condition and the glycine residue 102 determines the catalytic activity in acid-shock resistance. Although YfiD was demonstrated to be highly expressed and activated in microaerobic condition (69), the expression level remains high under aerobic environment suggesting a role in aerobic environment. The complementation assay using pRK415 and pCPP45 with or without the addition of the antibiotics tetracycline resulted in a steep drop of the survival rate, an involvement of tetracycline-resistant gene was hence proposed. TetA is a membrane-spanning H+-[tc-Mg]+ antiporter embedded in the cytoplasmic membrane (71, 72). Overexpression of TetA 24 .
(35) has been shown to be lethal for E. coli (3), which has been attributed to membrane potential collapse (17). Moreover, expression of TetA in the absence of tetracycline was also harmful to bacterial cells (2). It is hence other selection markers containing plasmids should be used to resolve the problem. E. coli yfiD mutant also revealed growth defect upon acid treatment (50). However, no apparent growth retardation of the ΔyfiD strain was observed in the growth curve comparison under pH 7.0, pH 5.5, aerobic and microaerobic environments. The yfiD null mutant of E. coli W3110 showed an apparent increase of acid-metabolite accumulation including pyruvate, formate, succinate and lactate (69). No growth defect was observed for K. pneumoniae CG43S3ΔyfiD in microaerobic growth could be attributed to the reutilization of the neutral metabolite acetoin to prevent for the accumulation of acidic metabolic end products in the environment (70). Analysis of K. pneumoniae CG43 genome sequence (http://140.113.173.58) revealed two proteins TdcE and YbiW which may compensate YfiD activity since a conserved radical-enzyme domain was identified. However, whether the radical-enzyme domain in these proteins is crucial for bacterial growth remains to be investigated. No study has indicated a regulatory role of EvgAS on the expression of yfiD in E. coli. The acid-shock resistance assay indicated a reduced survival of ΔyfiD mutant comparing to the parental strain implying a role of YfiD in the acid shock resistance in K. pneumoniae CG43. EvgAS has been demonstrated to regulate the expression of gadA and gadBC (45) which have been identified to be the pivotal components of AR system 2 25 .
(36) (42). The AR 2 assay condition (28) could also be employed to investigate if the phenotypic alteration of yfiD deletion is related to the acid resistance pathway. The cDNA subtractive hybridization analysis (39) revealed a KvhA-activated gene dcuB, encoding an anaerobic C4-dicarboxylate transporter (60). KvhAS to sense extracellular status or to cross-react with other regulator to execute the YfiD- and DcuB- mediated metabolic pathway is hence proposed. As shown in Fig. 13, the energy-source materials are ingested and flow through glycolysis and TCA cycle to derive energy, and finally go through an electronic transport chain using O2 as a terminal e- acceptor. While under microaerobic environment where fumarate respiration dominates (27), pyruvate, the end product of glycolysis, is introduced into further oxidation pathway by YfiD with fumarate as a terminal e- acceptor. It is then DcuB transports fumarate into the bacterium to replenish the fumarate pool, and pull succinate, a reduced form of fumarate, out to the environment (60).. 26 .
(37) References. 1.. Barua, S., T. Yamashino, T. Hasegawa, K. Yokoyama, K. Torii, and M. Ohta. 2002. Involvement of surface polysaccharides in the organic acid resistance of Shiga Toxin-producing Escherichia coli O157:H7. Mol Microbiol 43:629-40.. 2.. Berens, C., and W. Hillen. 2003. Gene regulation by tetracyclines. Constraints of resistance regulation in bacteria shape TetR for application in eukaryotes. Eur J Biochem 270:3109-21.. 3.. Berg, C. M., L. Liu, B. Wang, and M. D. Wang. 1988. Rapid identification of bacterial genes that are lethal when cloned on multicopy plasmids. J Bacteriol 170:468-70.. 4.. Bijlsma, J. J., and E. A. Groisman. 2003. Making informed decisions: regulatory interactions between two-component systems. Trends Microbiol 11:359-66.. 5.. Blankenhorn, D., J. Phillips, and J. L. Slonczewski. 1999. Acidand base-induced proteins during aerobic and anaerobic growth of Escherichia coli revealed by two-dimensional gel electrophoresis. J Bacteriol 181:2209-16.. 6.. Boulanger, A., A. Francez-Charlot, A. Conter, M. P. Castanie-Cornet, K. Cam, and C. Gutierrez. 2005. Multistress regulation in Escherichia coli: expression of osmB involves two independent promoters responding either to sigmaS or to the RcsCDB His-Asp phosphorelay. J Bacteriol 187:3282-6.. 7.. Bradford, M. M. 1976. A rapid and sensitive method for the 27 . .
(38) quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248-54. 8.. Castanie-Cornet, M. P., T. A. Penfound, D. Smith, J. F. Elliott, and J. W. Foster. 1999. Control of acid resistance in Escherichia coli. J Bacteriol 181:3525-35.. 9.. Chang, C., S. F. Kwok, A. B. Bleecker, and E. M. Meyerowitz. 1993. Arabidopsis ethylene-response gene ETR1: similarity of product to two-component regulators. Science 262:539-44.. 10.. Chang, F. Y., and M. Y. Chou. 1995. Comparison of pyogenic liver abscesses caused by Klebsiella pneumoniae and non-K. pneumoniae pathogens. J Formos Med Assoc 94:232-7.. 11.. Chang, H. Y., J. H. Lee, W. L. Deng, T. F. Fu, and H. L. Peng. 1996. Virulence and outer membrane properties of a galU mutant of Klebsiella pneumoniae CG43. Microb Pathog 20:255-61.. 12.. Christ, D., and J. W. Chin. 2008. Engineering Escherichia coli heat-resistance by synthetic gene amplification. Protein Eng Des Sel 21:121-5.. 13.. Croinin, T. O., and C. J. Dorman. 2007. Expression of the Fis protein is sustained in late-exponential- and stationary-phase cultures of Salmonella enterica serovar Typhimurium grown in the absence of aeration. Mol Microbiol 66:237-51.. 14.. De Biase, D., A. Tramonti, F. Bossa, and P. Visca. 1999. The response to stationary-phase stress conditions in Escherichia coli: role and regulation of the glutamic acid decarboxylase system. Mol Microbiol 32:1198-211.. 15.. de Lorenzo, V., and K. N. Timmis. 1994. Analysis and 28 . .
(39) construction of stable phenotypes in gram-negative bacteria with Tn5- and Tn10-derived minitransposons. Methods Enzymol 235:386-405. 16.. Deng, W. L., H. Y. Chang, and H. L. Peng. 1994. Acetoin catabolic system of Klebsiella pneumoniae CG43: sequence, expression, and organization of the aco operon. J Bacteriol 176:3527-35.. 17.. Eckert, B., and C. F. Beck. 1989. Overproduction of transposon Tn10-encoded tetracycline resistance protein results in cell death and loss of membrane potential. J Bacteriol 171:3557-9.. 18.. Fabret, C., V. A. Feher, and J. A. Hoch. 1999. Two-component signal transduction in Bacillus subtilis: how one organism sees its world. J Bacteriol 181:1975-83.. 19.. Freestone, P., S. Grant, M. Trinei, T. Onoda, and V. Norris. 1998. Protein phosphorylation in Escherichia coli L. form NC-7. Microbiology 144 ( Pt 12):3289-95.. 20.. Green, J., M. L. Baldwin, and J. Richardson. 1998. Downregulation of Escherichia coli yfiD expression by FNR occupying a site at -93.5 involves the AR1-containing face of FNR. Mol Microbiol 29:1113-23.. 21.. Grundy, F. J., D. A. Waters, T. Y. Takova, and T. M. Henkin. 1993. Identification of genes involved in utilization of acetate and acetoin in Bacillus subtilis. Mol Microbiol 10:259-71.. 22.. Hersh, B. M., F. T. Farooq, D. N. Barstad, D. L. Blankenhorn, and J. L. Slonczewski. 1996. A glutamate-dependent acid resistance gene in Escherichia coli. J Bacteriol 178:3978-81. 29 . .
(40) 23.. Highsmith, A. K., and W. R. Jarvis. 1985. Klebsiella pneumoniae: selected virulence factors that contribute to pathogenicity. Infect Control 6:75-7.. 24.. Huang, M., F. B. Oppermann-Sanio, and A. Steinbuchel. 1999. Biochemical and molecular characterization of the Bacillus subtilis acetoin catabolic pathway. J Bacteriol 181:3837-41.. 25.. Huang, T. Y. 2002. Characterization of the Two-component System KvhAS in Klebsiella pneumoniae CG43. National Chiao-Tung university master dissertation.. 26.. Iyer, R., C. Williams, and C. Miller. 2003. Arginine-agmatine antiporter in extreme acid resistance in Escherichia coli. J Bacteriol 185:6556-61.. 27.. Janausch, I. G., E. Zientz, Q. H. Tran, A. Kroger, and G. Unden. 2002. C4-dicarboxylate carriers and sensors in bacteria. Biochim Biophys Acta 1553:39-56.. 28.. Jennison, A. V., and N. K. Verma. 2007. The acid-resistance pathways of Shigella flexneri 2457T. Microbiology 153:2593-602.. 29.. Johansen, L., K. Bryn, and F. C. Stormer. 1975. Physiological and biochemical role of the butanediol pathway in Aerobacter (Enterobacter) aerogenes. J Bacteriol 123:1124-30.. 30.. Joseph, S., and D. W. Russell. 2001. Molecular Cloning: a laboratory manual—3rd edition. Cold Spring Harbor Laboratory Press.. 31.. Kato, A., H. Ohnishi, K. Yamamoto, E. Furuta, H. Tanabe, and R. Utsumi. 2000. Transcription of emrKY is regulated by the EvgA-EvgS two-component system in Escherichia coli K-12. 30 . .
(41) Biosci Biotechnol Biochem 64:1203-9. 32.. Keen, N. T., S. Tamaki, D. Kobayashi, and D. Trollinger. 1988. Improved broad-host-range plasmids for DNA cloning in gram-negative bacteria. Gene 70:191-7.. 33.. Kirkpatrick, C., L. M. Maurer, N. E. Oyelakin, Y. N. Yoncheva, R. Maurer, and J. L. Slonczewski. 2001. Acetate and formate stress: opposite responses in the proteome of Escherichia coli. J Bacteriol 183:6466-77.. 34.. Lacour, S., A. Kolb, and P. Landini. 2003. Nucleotides from -16 to -12 determine specific promoter recognition by bacterial sigmaS-RNA polymerase. J Biol Chem 278:37160-8.. 35.. Lai, M., C. 2000. Cloning, Expression and Analysis of the kvgAII Gene of Klebsiella pneumoniae CG43. National Chiao-Tung university master dissertation.. 36.. Lai, Y. C., H. L. Peng, and H. Y. Chang. 2001. Identification of genes induced in vivo during Klebsiella pneumoniae CG43 infection. Infect Immun 69:7140-5.. 37.. Lee, S. J., and J. D. Gralla. 2001. Sigma38 (rpoS) RNA polymerase promoter engagement via -10 region nucleotides. J Biol Chem 276:30064-71.. 38.. Levine, M. 1916. On the Significance of the Voges-Proskauer Reaction. J Bacteriol 1:153-64.. 39.. Lin, C. T. 2006. Functional analysis of the two-component systems KvgAS, KvhR and KvhAS in Klebsiella pneumoniae CG43. National Chiao-Tung university Ph. D. dissertation.. 40.. Lin, C. T., T. Y. Huang, W. C. Liang, and H. L. Peng. 2006. 31 . .
(42) Homologous response regulators KvgA, KvhA and KvhR regulate the synthesis of capsular polysaccharide in Klebsiella pneumoniae CG43 in a coordinated manner. J Biochem 140:429-38. 41.. Lin, C. T., and H. L. Peng. 2006. Regulation of the homologous two-component systems KvgAS and KvhAS in Klebsiella pneumoniae CG43. J Biochem 140:639-48.. 42.. Lin, J., I. S. Lee, J. Frey, J. L. Slonczewski, and J. W. Foster. 1995. Comparative analysis of extreme acid survival in Salmonella typhimurium, Shigella flexneri, and Escherichia coli. J Bacteriol 177:4097-104.. 43.. Lopez, J. M., B. Thoms, and H. Rehbein. 1975. Acetoin degradation in Bacillus subtilis by direct oxidative cleavage. Eur J Biochem 57:425-30.. 44.. Ma, Z., N. Masuda, and J. W. Foster. 2004. Characterization of EvgAS-YdeO-GadE branched regulatory circuit governing glutamate-dependent acid resistance in Escherichia coli. J Bacteriol 186:7378-89.. 45.. Masuda, N., and G. M. Church. 2003. Regulatory network of acid resistance genes in Escherichia coli. Mol Microbiol 48:699-712.. 46.. Matsushita, M., and K. D. Janda. 2002. Histidine kinases as targets for new antimicrobial agents. Bioorg Med Chem 10:855-67.. 47.. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring Harbor Laboratory Press.. 48.. Nishino, K., and A. Yamaguchi. 2001. Overexpression of the response regulator evgA of the two-component signal transduction system modulates multidrug resistance conferred by multidrug 32 . .
(43) resistance transporters. J Bacteriol 183:1455-8. 49.. Okada, A., Y. Gotoh, T. Watanabe, E. Furuta, K. Yamamoto, and R. Utsumi. 2007. Targeting two-component signal transduction: a novel drug discovery system. Methods Enzymol 422:386-95.. 50.. Partridge, J. D., G. Sanguinetti, D. P. Dibden, R. E. Roberts, R. K. Poole, and J. Green. 2007. Transition of Escherichia coli from aerobic to micro-aerobic conditions involves fast and slow reacting regulatory components. J Biol Chem 282:11230-7.. 51.. Peng, H. L., P. Y. Wang, J. L. Wu, C. T. Chiu, and H. Y. Chang. 1991. Molecular epidemiology of Klebsiella pneumoniae. Chinese journal of microbiology and immunology 24:264-71.. 52.. Perraud, A. L., V. Weiss, and R. Gross. 1999. Signalling pathways in two-component phosphorelay systems. Trends Microbiol 7:115-20.. 53.. Podschun, R., A. Fischer, and U. Ullmann. 1992. Siderophore production of Klebsiella species isolated from different sources. Zentralbl Bakteriol 276:481-6.. 54.. Podschun, R., and U. Ullmann. 1998. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev 11:589-603.. 55.. Quail, M. A., and J. R. Guest. 1995. Purification, characterization and mode of action of PdhR, the transcriptional repressor of the pdhR-aceEF-lpd operon of Escherichia coli. Mol Microbiol 15:519-29.. 56.. Sanchez, J. C., T. Powell, H. M. Staines, and R. J. Wilkins. 2006. 33 . .
(44) Electrophysiological demonstration of voltage- activated H+ channels in bovine articular chondrocytes. Cell Physiol Biochem 18:85-90. 57.. Sawers, G., and G. Watson. 1998. A glycyl radical solution: oxygen-dependent interconversion of pyruvate formate-lyase. Mol Microbiol 29:945-54.. 58.. Scarlato, V., B. Arico, M. Domenighini, and R. Rappuoli. 1993. Environmental regulation of virulence factors in Bordetella species. Bioessays 15:99-104.. 59.. Sirot, D. 1995. Extended-spectrum plasmid-mediated beta-lactamases. J Antimicrob Chemother 36 Suppl A:19-34.. 60.. Six, S., S. C. Andrews, G. Unden, and J. R. Guest. 1994. Escherichia coli possesses two homologous anaerobic C4-dicarboxylate membrane transporters (DcuA and DcuB) distinct from the aerobic dicarboxylate transport system (Dct). J Bacteriol 176:6470-8.. 61.. Skorupski, K., and R. K. Taylor. 1996. Positive selection vectors for allelic exchange. Gene 169:47-52.. 62.. Stock, J. B., A. M. Stock, and J. M. Mottonen. 1990. Signal transduction in bacteria. Nature 344:395-400.. 63.. Svanborg-Eden, C., L. Hagberg, R. Hull, S. Hull, K. E. Magnusson, and L. Ohman. 1987. Bacterial virulence versus host resistance in the urinary tracts of mice. Infect Immun 55:1224-32.. 64.. Tomas, J. M., S. Camprubi, S. Merino, M. R. Davey, and P. Williams. 1991. Surface exposure of O1 serotype lipopolysaccharide in Klebsiella pneumoniae strains expressing 34 . .
(45) different K antigens. Infect Immun 59:2006-11. 65.. Wagner, A. F., S. Schultz, J. Bomke, T. Pils, W. D. Lehmann, and J. Knappe. 2001. YfiD of Escherichia coli and Y06I of bacteriophage T4 as autonomous glycyl radical cofactors reconstituting the catalytic center of oxygen-fragmented pyruvate formate-lyase. Biochem Biophys Res Commun 285:456-62.. 66.. Wang, J. H., Y. C. Liu, S. S. Lee, M. Y. Yen, Y. S. Chen, J. H. Wang, S. R. Wann, and H. H. Lin. 1998. Primary liver abscess due to Klebsiella pneumoniae in Taiwan. Clin Infect Dis 26:1434-8.. 67.. Waterman, S. R., and P. L. Small. 1996. Identification of sigma S-dependent genes associated with the stationary-phase acid-resistance phenotype of Shigella flexneri. Mol Microbiol 21:925-40.. 68.. Weber, H., T. Polen, J. Heuveling, V. F. Wendisch, and R. Hengge. 2005. Genome-wide analysis of the general stress response network in Escherichia coli: sigmaS-dependent genes, promoters, and sigma factor selectivity. J Bacteriol 187:1591-603.. 69.. Wyborn, N. R., S. L. Messenger, R. A. Henderson, G. Sawers, R. E. Roberts, M. M. Attwood, and J. Green. 2002. Expression of the Escherichia coli yfiD gene responds to intracellular pH and reduces the accumulation of acidic metabolic end products. Microbiology 148:1015-26.. 70.. Xiao, Z., and P. Xu. 2007. Acetoin metabolism in bacteria. Crit Rev Microbiol 33:127-40.. 71.. Yamaguchi, A., Y. Iwasaki-Ohba, N. Ono, M. Kaneko-Ohdera, 35 . .
(46) and T. Sawai. 1991. Stoichiometry of metal-tetracycline/H+ antiport mediated by transposon Tn10-encoded tetracycline resistance protein in Escherichia coli. FEBS Lett 282:415-8. 72.. Yamaguchi, A., T. Udagawa, and T. Sawai. 1990. Transport of divalent cations with tetracycline as mediated by the transposon Tn10-encoded tetracycline resistance protein. J Biol Chem 265:4809-13.. 73.. Zhu, J., S. Shalel-Levanon, G. Bennett, and K. Y. San. 2007. The YfiD protein contributes to the pyruvate formate-lyase flux in an Escherichia coli arcA mutant strain. Biotechnol Bioeng 97:138-43.. 74.. 林佳融,大專生參與專題研究計畫,核定計畫編號 94-2815-C-009-045-B,探討 KvhAS 雙分子系統在克雷白氏肺炎 桿菌抗酸能力上所扮演的調控角色,中華民國九十五年六月。. 36 .
(47) Tables Table 1. Bacterial strains used in this study Strain. Genotype or relevant property. E. coli: BL21(DE3). F – ompT hsdSB(rB– mB–) gal dcm met (DE3). JM109. RecA1 supE44 endA1 hsdR17 gyrA96 rolA1 thi. Δ(lac-proAB) - - BL21-CodonPlus(DE F ompT hsdSB(rB mB )gal dcm(DE3) 3)-RIL Tpr Smr recA, thi, pro, hsdR-M+ S17-1λpir. Reference or source Laboratory stock Laboratory stock Laboratory stock (15). [RP4-2-Tc::Mu:Kmr Tn7](pir) K. pneumoniae: CG43 CG43S3 CG43S3-Z01. K2 serotype rspl mutant, Strepr lacZ mutant in CG43 rsplCG43S3- Z01ΔkvhA kvhA mutant in CG43 rspl- lacZCG43S3- Z01ΔrpoS rpoS mutant in CG43 rspl- lacZCG43S3-U9451 galU deletion mutant kvhAS mutant in CG43 rsplCG43S3ΔkvhAS CG43S3kvhAS mutant in CG43 rspl- galU-. (51) (51) (40) (41) (41) (11) (40) (39). U9451ΔkvhAS CG43-S3ΔkvhS CG43-S3ΔyfiD. (40) This study. kvhS mutant in CG43 rsplyfiD mutant in CG43 rspl-. 37 .
(48) Table 2. Plasmids used in this study Plasmid. Relevant characteristic. Source or reference. yT&A pET-30b pKAS46 pHP4001 pHP4005 pHAm pRK415 pLacZ15. Yeastern Biotech Co. Novagene (61) (35) (40) (41) (32) (40). pCPP45. PCR cloning vector, Apr His6-tagging protein expression vector, Kmr Suicide vector, rpsL, Kmr putative kvhA promoter region cloned into pUC-T pET-30c carrying kvhA gene BamHI digested fragment of pHP4005 cloned into BamHI site of pETm-c broad-host-range IncP cloning vector, Tcr A derivative of pYC016 (36), containing a promoterless lacZ from K. pneumoniae CG43S3 as the reporter, Cmr broad-host-range cloning vector with the partition region from RK2 plasmid, Tcr. pYm67TA pYm89TA pYm2KTA pYm2KAS pKvhAcTA pKvhAcPP pHAcATA pHAcAPP pHAcETA pHAcEPP. 1081-bp PCR product of the region upstream of yfiD cloned into yT&A 1148-bp PCR product of the region downstream of yfiD cloned into yT&A BamHI digested fragment of pYm89TA cloned into pYm67TA EcoRI/XbaI digested fragment of pYm2KTA cloned into pKAS46 698-bp PCR product carrying kvhA cloned into yT&A BamHI/EcoRI digested fragment of pKvhAcTA cloned into pCPP45 698-bp PCR product expressing KvhAD52A cloned into yT&A BamHI/EcoRI digested fragment of pHAcATA cloned into pCPP45 698-bp PCR product expressing KvhAD52E cloned into yT&A BamHI/EcoRI digested fragment of pHAcETA cloned into pCPP45 38 . . Dr. David Bauer at Cornell University This study This study This study This study This study This study This study This study This study This study.
(49) Plasmid. Relevant characteristic. Source or reference. pYcTA pYcPP pYcRK pYc102TA pYc102PP pYc102RK pY34TA pY34B pHArdTA pHArdB pYfiD01 pYfiD02 pY25TA pY25Z pY27TA pY27Z pY28TA pY28Z pY29TA pY29Z pUNY. 502-bp PCR product carrying yfiD cloned into yT&A BamHI/EcoRI digested fragment of pYcTA cloned into pCPP45 BamHI/EcoRI digested fragment of pYcTA cloned into pRK415 502-bp PCR product expressing YfiDG102A cloned into yT&A BamHI/EcoRI digested fragment of pYc102TA cloned into pCPP45 BamHI/EcoRI digested fragment of pYc102TA cloned into pRK415 429-bp PCR product carrying yfiD cloned into yT&A 441-bp BamHI/HindIII digested fragment of pY34TA cloned into pET-30b 337-bp PCR product BamHI/EcoRI digested fragment of pHArdTA cloned into pET-30b 302-bp PCR product carrying PyfiD-2 cloned into yT&A BamHI/BglII digested fragment of pYfiD01 cloned into the BamHI site of pLacZ15 417-bp PCR product carrying PyfiD-1 cloned into yT&A BamHI/BglII digested fragment of pY25TA cloned into the BamHI site of pLacZ15 274-bp PCR product carrying PyfiD-3 cloned into yT&A BamHI/BglII digested fragment of pY27TA cloned into the BamHI site of pLacZ15 204-bp PCR product carrying PyfiD-4 cloned into yT&A BamHI/BglII digested fragment of pY28TA cloned into the BamHI site of pLacZ15 130-bp PCR product carrying PyfiD-5 cloned into yT&A BamHI/BglII digested fragment of pY29TA cloned into the BamHI site of pLacZ15 1010-bp PCR product carrying yfiD and PyfiD cloned into yT&A. This study This study This study This study This study This study This study This study This study This study Laboratory Stock Laboratory Stock This study This study This study This study This study This study This study This study This study. 39 .
(50) Table 3. Primers used in this study Primer. Sequence. yfiDm06 yfiDm07 yfiDm08 yfiDm09 kvhAc01 kvhAc02 hAd52a-F3 hAd52a-R3 hAd52e-F2 hAd52e-R2 yfiDc01 yfiDc02 yfiDg102a-F3 yfiDg102a-R3 kvhA-rd-F kvhA-rd-R yfiD01 yfiD02 yfiD03 yfiD04 yfiD05 yfiD07 yfiD08 yfiD09. 5’- GAATGGTGGCGTCAGAACAAGGCG -3’ 5’- GGATCCCAGGCAGTGTTTATTCGA -3’ 5’- GGATCCGCGGTACGCTTCAACTCT -3’ 5’- TCTAGACATCGCCACCATTACCGG -3’ 5’- CGCGGATCCTGCACCAGGC -3’ 5’- GATAGAATTCGACGAACCGCCATG -3’ 5’- CCTGCTCATCGTCGCTGTCGATATTCCG -3’ 5’- CGGAATATCGACAGCGACGATGAGCAGG -3’ 5’- CCTGCTCATCGTCGAAGTCGATATTCCGGA -3’ 5’- TCCGGAATATCGACTTCGACGATGAGCAGG -3’ 5’- GGATCCGGCTGATATATATACAC -3’ 5’- GAATTCACATCTGGCGTTAGCCT -3’ 5’- CAGCTGACCATTCGTGTTTCCGCATACGCG -3’ 5’- CGCGTATGCGGAAACACGAATGGTCAGCTG -3’ 5’- TCTGCAGCCCATGAATGCGATAAT -3’ 5’- GATTCAATTCATTCCCTCTTTTTT -3’ 5’- CATCGCTAGATCTTAACGGC -3’ 5’- CCAGTAATCATTGGATCCCTC -3’ 5’- CGAATTCACACTGCCTGGGA -3’ 5’- CACATCTGGCGTTAGCCTTA -3’ 5’- GGAGATCTTCGGCGACGGTC -3’ 5’- ATAGATCTAAATCGGCACGG -3’ 5’- AGATCTCACAATGGCGTAAC -3’ 5’- AGATCTATATCAAGGAATGCCG -3’. 40 .
(51) Figures. ‐405. PyfiD‐1. tggtgatcccggactggcgctcttcggcgacggtcttcagagtattgagaaaatagggttg RpoS element II. tTGCTTTTCATccgccagcacgtcatgccaggttaaaggggtggtcatcttgctctcctgc ‐358 PyfiD‐3 PyfiD‐2 ‐290 ‐262 RpoS element I. aaaagggtcaTCGCTATAgcttaacggcttctttctatagaacaaaatcggcacggcttga PyfiD‐4 ‐288 ‐193 tgccggcttttatgcaaaaaaaattgaaaatttacaaaattatttatagctcacaatggcg PyfiD‐5 putative EvgA binding box ArcA elements. ‐119. TAACTTGATGTAAAACAATAAAaacctcctatacccacgttgggtgagtggttTTTATTGA FNR element II. ‐35 box. ‐10 box. TTTATATCAAGGAatgccgagggtgcggctgatatatatacactcatacaacaatggtttt FNR element I. +1. accaattggccgcgaagaggccaaaatcgaataaacactgcctgggaggcatcaaatg. yfiD. Figure 1. Sequences of the putative yfiD promoter. A putative -10 box and -35 box are highlighted. FNR element I and II, ArcA element, RpoS element I and II, and a putative EvgA binding box (31, 34, 37, 69) are also indicated. The promoter containing regions used in the study include PyfiD-1 (pY25Z), PyfiD-2 (pYfiD02), PyfiD-3 (pY27Z), PyfiD-4 (pY28Z) and PyfiD-5 (pY29Z).. 41 .
(52) (A). (B) RpoS element II. (C) PyfiD activity (Micro‐Aerobic). RpoS element I yfiD. ‐405. ‐358. PyfiD‐1. ‐288. PyfiD activity (Aerobic). Z01 Z01ΔkvhA Z01ΔrpoS. +1. ‐290. PyfiD‐2. +1. ‐262. PyfiD‐3. +1 0. 2000. 4000. 6000. 8000. β-galactosidase activity (Miller Unit). 10000. 12000. 0. 500 1000 1500 2000 2500. β-galactosidase activity (Miller Unit). Figure 2. The deletion of kvhA or rpoS reduced the expression of yfiD in microaerobic environment. (A) Two RpoS binding elements (I & II) and the putative promoters are shown. The promoters PyfiD-1, PyfiD-2, and PyfiD-3 respectively containing 405 bp, 290 bp, and 262 bp noncoding sequence upstream of the yfiD start codon were isolated and inserted in front of the promoterless lacZ gene of pLacZ15. (B) The bacteria were cultured for 4 h in 15 ml tightly sealed culture tube filled with LB and the activity of PyfiD-1, PyfiD-2 and PyfiD-3 in either of K. pneumonaeCG43S3-Z01, CG43S3-Z01ΔkvhA, or CG43S3-Z01ΔrpoS was measured.. 42 .
(53) (A) KDa 72. (B) 1 2 3 4 5 6 7 8 9 10. 1 2 3 4 5 KDa 72. 55 43. 55 43. His6‐KvhA. 34. 34. 26. 26 His6‐KvhAΔHTH. 17. 17. 11. 11. IPTG (mM) 0.1 0 0.1 0.1 0.1 0.1 0 0.1 0.1 0.1. IPTG (mM) 0.1 0 0.1 0.1 0.1. Plasmid pET‐30c pHP4005 pET‐30c pHP4005. Plasmid pET‐30b. Host E. coli. Host E. coli. BL21(DE3) BL21‐RIL(DE3). pHArdB BL21(DE3). Figure 3. Overexpression of His6-KvhA and His6-KvhAΔHTH proteins. (A) pHP4005 carrying kvhA coding sequence was transformed into E. coli BL21(DE3) and BL21-RIL respectively. After 5 h IPTG induction (lanes 1 to 3, 6 to 8), the induced bacteria (lanes 3, 8) were disrupted and centrifugated into pellet (Lanes 4, 9) and supernatant (Lanes 5, 10) forms. The supernatant containing soluble His6-KvhA (lane 10) was then subjected to purification. (B) pHArdB carrying the kvhA coding sequence deletion of HTH domain was transformed into E. coli BL21(DE3). After 5 h IPTG induction (lanes 1 to 3), the induced bacterium (lane 3) was disrupted and centrifugated into pellet (lanes 4) and supernatant (lanes 5) forms. The supernatant containing soluble His6-KvhA ΔHTH (lane 5) was also subjected to purification. 43 .
(54) (A). (B). 34 26 17. KDa. KDa 72. 55 43 His6‐KvhA. KDa 72. 72 55. 55. 43. His6‐KvhA. 55. 43. 34. 43. 34. 26. 34. 26. His6‐KvhAΔHTH. His6‐KvhAΔHTH 17. 11. (D). 1 2 3 4 5 6 7 8 9 10. 1 2 3 4 5 6 7 8 9 10 KDa 72. (C). 17. 11. 26. 17. Figure 4. Purification of His6-KvhA and His6-KvhAΔHTH proteins. Soluble fractions of the total cell lysates respectively containing His6-KvhA (A) and His6-KvhAΔHTH (B) were subjected to His6-Bind Resin column for purification. The protein purity of each eluted fractions containing His6-KvhA (lanes 1 to 10 of (A)) and His6-KvhAΔHTH (lanes 1 to 10 of (B)) were then analyzed by SDS-PAGE. The collected fractions were subjected to dialysis, concentration and the purity of His6-KvhA (C) and His6-KvhAΔHTH (D) confirmed by SDS-PAGE.. 44 .
(55) (A) Lane 1 2 3 4 5 6 His6‐KvhA (μg) BSA (μg) 0.4 0.4 0.4 0.4 0.4 2.0. Protein‐probe complex Free probe (PyfiD‐2). (B) Lane 1 2 3 4 5 6 His6‐KvhA (μg) BSA (μg) 0.4 0.4 0.4 0.4 0.4 2.0. Protein‐probe complex Free probe (PyfiD‐5). Figure 5. Direct binding of the phosphorylated His6-KvhA to PyfiD. The direct binding activity of His6-KvhA was assayed by electrophoretic mobility shift assay with 5 % (w/v) non-denature polyacrylamide gel. Different amount of the purified His6-KvhA were phosphorylated in 1 X binding buffer in the presence of 50 mM acetylphosphate and 1μg of acetylated bovine serum, and then the 32P-labelled PyfiD-2 (A) or PyfiD-5 (B) DNA fragments was added. The His6-KvhA added in lanes 1 to 6 respectively was 0 μg, 0.18 μg, 0.54 μg, 1.08 μg, 1.44 μg and 0 μg.. 45 .
(56) (A) 1010 bp ung01. ung. yfiD. srmB. 308 bp Deletion. yfiD04. (B) bp. 1 2 3 4. 1500 Wild‐type form. 1000 750. Mutant form. 500. 1: pYm2KAS 2: CG43S3ΔyfiD (1) 3: CG43S3ΔyfiD (6) 4: CG43S3. Figure 6. Construction of the yfiD deletion mutant. (A) Schematic representation of yfiD and the flanking genes. Primer set ung01/yfiD04 was designed to confirm yfiD mutant. (B) PCR analysis using ung01/yfiD04 primer to confirmed the deletion of yfiD. The plasmid pYm2KAS contains the flanking DNA of the yfiD for homologous recombination to generate the yfiD deletion mutant.. 46 .
(57) (A). (B) 1 2 3 4 5. KDa. KDa. 116.0 66.2 37.6 28.5. His6‐YfiD. (C) 1 2 3 4 5 6 7 8 9 10 KDa. 116.0. 116.0. 66.2. 66.2. 37.6. 37.6 His6‐YfiD. 28.5. 28.5 His6‐YfiD. 18.4 14.0 IPTG (mM) 1 0 1 1 1 Plasmid pET‐30b pY34B Host E. coli. 18.4 14.0. 18.4 14.0. BL21(DE3). Figure 7. Expression and purification of His6-YfiD protein. (A) The coding sequence of yfiD gene was cloned into pET-30b and the resulting plasmid pY34B transformed into E. coli BL21(DE3). After 5 h IPTG induction (lanes 1 to 3), the induced bacterium (lane 3) was disrupted and centrifugated into pellet (lane 4) and supernatant (lane 5) forms. The supernatant containing soluble His6-YfiD (lane 5) was then subjected to His6-Bind Resin column for purification. (B) The protein purity of each eluted fractions containing His6-YfiD (lanes 1 to 10) were then analyzed by SDS-PAGE. The collected fractions were subjected to dialysis, concentration and the purity of His6-YfiD (C) confirmed by SDS-PAGE.. 47 .
(58) (A) KDa. 1 2 3 4 5. 6 7 8. 9. 10. 72 55 43 34 26 17. YfiD 11. (B) KDa. 1 2 3 4 5. 6. 7. 8. 9. 10. 72 55 43 34 26 17. YfiD 11. Figure 8. Western blotting analysis of the YfiD protein. Overnight bacteria were refreshed grown in (A) LBMES (pH 5.5) and (B) M9MES (pH 5.5) medium at 37℃ aerobically for 6 h and then subjected to SDS-PAGE and western blot hybridization using the YfiD antiserum. Lanes 1 and 6 are the cell lysates from K. pneumoniae CG43S3; 2 and 7 from CG43S3ΔkvhA; 3 and 8 from CG43S3ΔyfiD; 4 and 9 from CG43S3 [pHAm]; 5 and 10 from CG43S3ΔyfiD [pYcRK].. 48 .
數據



+7
相關文件
In JSDZ, a model process in the modeling phase is treated as an active entity that requires an operation on its data store to add a new instance to the collection of
z The caller sent signaling information over TCP to an online Skype node which forwarded it to callee over TCP. z The online node also routed voice packets from caller to callee
In Case 1, we first deflate the zero eigenvalues to infinity and then apply the JD method to the deflated system to locate a small group of positive eigenvalues (15-20
As the above-mentioned 'An Outline to the Classified Books...' is an authorized and influental academic work in the bibliographical field, the late
The PE curriculum contributes greatly to enabling our students to lead a healthy lifestyle with an interest and active participation in physical and aesthetic
In 1990s, the research area of nonhomogeneous Harmonic analysis became very active due to three famous mathematicians Nazarov, Treil and Volberg published an important result in
According to the Heisenberg uncertainty principle, if the observed region has size L, an estimate of an individual Fourier mode with wavevector q will be a weighted average of
For the proposed algorithm, we establish its convergence properties, and also present a dual application to the SCLP, leading to an exponential multiplier method which is shown
