Elsevier Editorial System(tm) for Food and Chemical Toxicology Manuscript Draft
Manuscript Number: FCT-D-11-01547
Title: Antioxidant phenolic profile from ethyl acetate fraction of Fructus Ligustri Lucidi with protection against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells
Article Type: Full Length Article
Keywords: Fructus Ligustri Lucidi; hydroxytyrosol; salidroside; antioxidant activity; oxidative stress; antioxidant enzymes.
Corresponding Author: Dr. Chi-Rei Wu,
Corresponding Author's Institution: Institutent of Chinese Pharmaceutical Sciences, CHina Medical University
First Author: Heng-Yin Ju
Order of Authors: Heng-Yin Ju; Shiu-Ching Chen; Kuo-Jen Wu; Hui-Chun Wu; Hui Ching; Chi-Rei Wu Abstract: In this study, we demonstrated the antioxidant and protective properties of crude extract and fractions from Fructus Ligustri Lucidi (FLL) against hydrogen peroxide (H2O2)-induced oxidative damage in SH-SY5Y cells. The contents of the phytochemical profiles from FLL crude extracts and fractions were determined by spectrophotometric methods and high performance liquid
chromatography using a photodiode array detector. FLL crude extract possessed appreciable scavenging capacity against 1,1-diphenyl-2-picrylhydrazyl and H2O2. The ethyl acetate (EtOAc) fraction was the most active fraction in scavenging free radicals and H2O2. Following exposure of cells to H2O2, there was a marked decrease in cell survival and intracellular antioxidant enzymes.
Intracellular oxidative stress, the level of lipid peroxidation, and caspase-3 activity were increased when cells was exposed to H2O2. Simultaneous treatment with the EtOAc fraction blocked these H2O2-induced cellular events. Hydroxytyrosol and salidroside are the major components of the EtOAc fraction. These results show that the EtOAc fraction of FLL is enriched in phenol and contains tyrosol-related derivatives and that this fraction exerts protective effects against H2O2 toxicity via its free radical scavenging activity and ability to elevate the levels of antioxidant enzymes.
Suggested Reviewers: Antoni Camins
Unitat de Farmacologia i Farmacognòsia i Institut de Biomedicina (IBUB), Facultat de Farmàcia, Universitat de Barcelona, Nucli Universitari de Pedralbes, Barcelona, Spain
[email protected] Anat Elmann
Department of Food Science, Agricultural Research Organization, The Volcani Center, Bet Dagan, Israel [email protected]
Pinar Oztopcu-Vatan
Eskisehir Osmangazi University Department of Biology, Faculty of Art and Sciences Meselik, Eskisehir Turkey
Eduardo Madrigal-Bujaidar
Laboratorio de Genética, Escuela Nacional de Ciencias Biológicas, IPN, Unidad Profesional Adolfo López Mateos, Zacatenco, Wilfredo Massieu, Lindavista, 07738 México D.F., Mexico
[email protected] Opposed Reviewers:
September 07, 2011
Dear Editor-in-Chief:
We submit this manuscript which entitled "Antioxidant phenolic
profile from ethyl acetate fraction of Fructus Ligustrum Lucidi with protection against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells" for publication in the Food and Chemical Toxicology.
Fructus Ligustrum Lucidi (FLL) is commonly used for curing hepatitis and aging-associated symptoms by traditional Chinese physicians for centuries. This study evaluated the antioxidant properties and components of FLL, and further demonstrated their protective effects against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells. We found FLL partitioned with ethyl acetate has best antioxidant properties in vitro and enriched phenolic contents. It further protects oxidative damage caused by hydrogen peroxide in part from the upregulation of intracellular antioxidant status and the inhibition of caspase-3 activation. Hydroxytyrosol and salidroside are major components of ethyl acetate fraction of FLL. Six graphics (including one supplementary graph) and three tables are presented in this manuscript.
This paper is an original article and has not been submitted elsewhere for publications. I hope that it will be considered for publication in the Food and Chemical Toxicology. Thank you very much for your help and wish to
hear from you very soon.
Sincerely Yours,
Chi-Rei Wu
The School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University
91, Hsueh Shih Road, Taichung, 40424, Taiwan, R.O.C. [email protected]
中國醫藥大學
China Medical University台中市學士路91號,台灣
91, Hsueh Shih Road, Taichung Taiwan, R.O.C.
The School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources TEL: 886-4-2203366-5506 FAX:
E-mail: [email protected]
Food and Chemical Toxicology
Antioxidant phenolic profile from ethyl acetate fraction of Fructus Ligustrum Lucidi with protection against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells
Dear editor,
Thank you for your useful comments and suggestions on the language and the structure of our manuscript. We have modified the manuscript accordingly, and the detailed corrections are listed below point by point:
1) Research highlights provided should be indicated by bullet points, with a maximum of 85 characters (including spaces) pre bullet point.
We have corrected research highlights and re-upload the corrected “Research highlight” file.
2) Abstract and a list of keywords should be provided before the introduction section. We have added the abstract and keywords before the “introduction” section and re-upload the corrected “manuscript” file.
3) Figure 5 is not cited in the text.
We have cited Figure 5 in the corrected “manuscript” file.
4) Please check the suggested reviewers instructions - there should be 4 reviewers of which 2 should be from a different country to the corresponding author. E-mail addresses containing hotmail, gmail, and yahoo accounts should not be used.
We have changed two reviewers.
Thank you very much for your help and the manuscript has been resubmitted to your journal. We look forward to your positive response.
Sincerely Yours,
Chi-Rei Wu
The School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources
College of Pharmacy China Medical University
91, Hsueh Shih Road, Taichung, 40424, Taiwan, R.O.C. [email protected]
Food and Chemical Toxicology
Conflict of Interest PolicyDeclarations
Food and Chemical Toxicology requires that all authors sign a declaration of conflicting interests. If you have nothing to declare in any of these categories then this should be stated.
Conflict of Interest
A conflicting interest exists when professional judgement concerning a primary interest (such as patient’s welfare or the validity of research) may be influenced by a secondary interest (such as financial gain or personal rivalry). It may arise for the authors when they have financial interest that may influence their interpretation of their results or those of others. Examples of potential conflicts of interest include employment, consultancies, stock ownership, honoraria, paid expert testimony, patent applications/registrations, and grants or other funding.
Please state any competing interests
No competing interest.
Funding Source
All sources of funding should also be acknowledged and you should declare any involvement of study sponsors in the study design; collection, analysis and interpretation of data; the writing of the
manuscript; the decision to submit the manuscript for publication. If the study sponsors had no such involvement, this should be stated.
Please state any sources of funding for your research
We have described all funding in the “Acknowledgment” Section of our manuscript.
Signature (a scanned signature is acceptable, Print name
but each author must sign)
Heng-Yin Ju
Shiu Ching Chen
Kuo-Jen Wu
Hui-Chun Wu
Hui Ching
Chi-Rei Wu
Supplement:
Article Title: Antioxidant phenolic profile from ethyl acetate fraction of Fructus Ligustri Lucidi with protection against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells
Author name:Heng-Yin Ju, Shiu Ching Chen, Kuo-Jen Wu, Hui-Chun Wu, Hui Ching, Chi-Rei Wu
Original research paper
Antioxidant phenolic profile from ethyl acetate fraction of Fructus Ligustri Lucidi with protection against hydrogen peroxide-induced oxidative damage in SH-SY5Y cells
Heng-Yin Ju a,1, Shiu Ching Chen b,1, Kuo-Jen Wu a, Hui-Chun Wu a, Hui Ching b,
Chi-Rei Wu a,*
Affiliation
a The School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources,
College of Pharmacy, China Medical University, No.91, Hsueh Shih Road, Taichung, 40402, Taiwan, R.O.C.
b Taichung Hospital, Department of Health, The Executive Yuan, No.199, San Min
Rd., 40403, Taichung, Taiwan, R.O.C.
Correspondence
Dr. Chi-Rei Wu, The School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, College of Pharmacy, China Medical University, 91, Hsueh Shih Road, Taichung, 40402, Taiwan, R.O.C. E-mail: [email protected] Phone: + 886 4 22053366 5506
1 Both authors contribute equally *Manuscript Title Page
Fructus Ligustri Lucidi is commonly used for aging-associated symptoms Its phenolic-enriched fraction has the best antioxidant activity
It protects oxidative damage via elevating intracellular antioxidant status Hydroxytyrosol and salidroside are its major active components
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Abstract
In this study, we demonstrated the antioxidant and protective properties of crude extract and fractions from Fructus Ligustri Lucidi (FLL) against hydrogen peroxide (H2O2)-induced oxidative damage in SH-SY5Y cells. The contents of the
phytochemical profiles from FLL crude extracts and fractions were determined by spectrophotometric methods and high performance liquid chromatography using a photodiode array detector. FLL crude extract possessed appreciable scavenging capacity against 1,1-diphenyl-2-picrylhydrazyl and H2O2. The ethyl acetate (EtOAc)
fraction was the most active fraction in scavenging free radicals and H2O2. Following
exposure of cells to H2O2, there was a marked decrease in cell survival and
intracellular antioxidant enzymes. Intracellular oxidative stress, the level of lipid peroxidation, and caspase-3 activity were increased when cells was exposed to H2O2.
Simultaneous treatment with the EtOAc fraction blocked these H2O2-induced cellular
events. Hydroxytyrosol and salidroside are the major components of the EtOAc fraction. These results show that the EtOAc fraction of FLL is enriched in phenol and contains tyrosol-related derivatives and that this fraction exerts protective effects against H2O2 toxicity via its free radical scavenging activity and ability to elevate the
levels of antioxidant enzymes.
Keywords: Fructus Ligustri Lucidi; hydroxytyrosol; salidroside; antioxidant activity; oxidative stress; antioxidant enzymes
*Manuscript
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 Main text 1. Introduction
For centuries, Fructus Ligustri Lucidi (FLL) has been commonly used by traditional Chinese physicians for curing hepatitis and aging-associated symptoms. Harman (2006) suggested that the aging process is due to oxidative stress cascades, including attack from reactive oxygen species (ROS) and deficiency in intracellular antioxidative defense. In fact, many chronic diseases, including neurodegenerative diseases and cardiovascular diseases, may be caused by intracellular oxidative damage to biomolecules, such as carbohydrates, proteins, lipids and DNA, via ROS (Butterfield and Sultana, 2008; Fearon and Faux, 2009). Minimizing the cellular redox imbalance may be one of the most important approaches to the prevention of these aging-associated diseases. Antioxidants scavenge free radicals, which initiate and propagate oxidative chain reactions, and thus prevent intracellular oxidative damage (Valko et al., 2007). Natural antioxidants, such as phenols and flavonoids, have been revealed to possess multiple pharmacological activities, including
neuroprotective and anti-aging activities, via their antioxidant properties (Uttara et al., 2009).
Recent pharmacological studies have indicated that FLL possesses
hepatoprotective and antidiabetic activities (Gao et al., 2009; Yim et al., 2001), and some reports have indicated that FLL possesses antioxidant activities (He et al., 2001; Li et al., 2007). Therefore, its hepatoprotective and antidiabetic activities may be due to improvements intracellular antioxidative status, including glutathione regeneration and upregulation of antioxidant enzymes (Gao et al., 2009; Yim et al., 2001). Its active components have been recognized as triterpenoids, secoiridoids and the glycosides of these compounds (Gao et al., 2009; Li et al., 2007; Yim et al., 2001).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
In this paper, we evaluated the antioxidant and radical scavenging activities of FLL crude extract and fractions by several different in vitro antioxidant test systems, such as the 1-diphenyl-2-picrylhydrazyl (DPPH) radical-scavenging assay, the trolox equivalent antioxidant capacity (TEAC) assay, the reducing power assay and the iron-chelating activity (ICA) assay. Because ROS play an important role in
intracellular oxidative damage, we also evaluated the effects of FLL crude extract and fractions on scavenging hydrogen peroxide (H2O2) and hydroxyl radical and
inhibiting lipid peroxidation by the Fenton reaction. We further attempted to
demonstrate the possible protective effects of FLL crude extract and fractions against H2O2-induced oxidative damage in SH-SY5Y cells. Based on earlier reports on the
bioactive components of FLL (Gao et al., 2009; He et al., 2001; Li et al., 2007; Yim et al., 2001), the contents of all phytochemical profiles, including phenols, flavonoids, phenylpropanoids, iridoids and triterpenoids, from FLL crude extract and fractions were determined by spectrophotometry and high performance liquid
chromatography-photodiode array detector (HPLC-DAD).
2. Materials and methods
2.1. Preparation of herb extract and fractionation
FLL was purchased from an herb supplier in Taiwan. FLL (6 kg) was extracted five times with 50% ethanol. The resultant extract was combined and concentrated under reduced pressure to obtain 1,330 g of residue (22.7%). The crude extract was suspended in water and then partitioned successively with four-times-volume amounts of chloroform, ethyl acetate and n-butanol. Each fraction was evaporated in a vacuum to obtain the chloroform (CHCl3, 31.3 g, 2.35%), ethyl acetate (EtOAc, 61.3 g,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 2.2. Chemicals
2-deoxyribose, 2,2’-azino-bis-3-ethylbenzthiazoline-6-sulfonic acid (ABTS), 2’, 7’-dichlorofluorescein diacetate (DCFH-DA), 2-vinylpyridine (2-VP),
3-(2-pyridyl)-5,6-diphenyl-1,2,4-triazine-4’,4”-disulfonic acid (ferrozine), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT),
5,5’-dithio-bis(2-nitrobenzoic) acid (DTNB),
6-hydroxy-2,5,7,8-tetramethy-chroman-2-carboxylic acid (trolox), acridine orange (AO), aluminum nitrate, ascorbic acid, betulin, (+)-catechin, DPPH, ferric chloride, ferrous sulfate heptahydrate, Folin-Ciocalteu’s reagent, reduced glutathione (GSH), glutathione peroxidase (GPx), glutathione reductase (GR), homovanillic acid (HVA), horseradish peroxidase (HRPase), p-hydroxybenzoic acid (HBA), malodialdehyde (MDA), nitroblue tetrazolium chloride (NBT), oxidized glutathione (GSSG), potassium acetate, potassium ferricyanide, potassium persulfate, quercetin, sodium carbonate, superoxide dismutase (SOD), thiobarbituric acid (TBA), triethanolamine and trichloroacetic acid (TCA) were purchased from Sigma-Aldrich Chem. Corp. (St. Louis, MO, USA). Aucubin, betulinic acid, hydroxytyrosol, nuhenzide, oleanolic acid, oleuropein, protocatechuic acid (PCA), quercitrin, salidroside, tyrosol, ursolic acid and verbascoside standards were purchased from Extrasynthese (Lyon Nord, GENAY, France). H2O2 and o-phosphoric acid were purchased from Merck
(Darmstadt, Germany). All solvents were HPLC grade and also purchased from Merck.
2.3. Determination of antioxidant and radical scavenging activities 2.3.1. Radical scavenging activity assay
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
The DPPH radical scavenging capacity assay was performed as described
previously (Wu et al., 2011b). Briefly, 25 L of sample solution or catechin standard was pipetted into each well, and then 175 L of 300 M DPPH solution was added. The absorbance of the mixture was read at 517 nm with a spectrophotometric microplate reader (Bio-Tek, PowerwaveX340, Winooski, VT) after a 30-min
incubation at 25 °C. The results were expressed as mmol of (+)-catechin equivalents per gram of sample.
2.3.2. TEAC assay
TEAC was measured by the ABTS radical scavenging assay (Wu et al., 2011b). First, ABTS radical was prepared by reaction of 8 mM ABTS solution and 8.4 mM potassium persulfate solution at a ratio of 2:1, respectively. After storage in the dark at RT for 12-16 h, the radical solution was further diluted with ethanol until an initial absorbance value of 0.70 ± 0.05 at 734 nm was reached. One hundred seventy-five L of diluted ABTS solution was mixed with 25 L of sample solution or trolox
standard. The results were expressed as TEAC values.
2.3.3. Reducing power assay
Reducing power was determined spectrophotometrically by the formation of Perl’s Prussian blue colored complex (Wu et al., 2011b). First, 25 L of sample solution was loaded into each well. Then 100 L of 50 M phosphate buffer (pH 6.6) and 0.1% (w/v) potassium ferricyanide was added. After incubation at 50 C for 20 min, 100 L of 1% (w/v) TCA was added. Finally, 200 L of the mixture solution was transferred into a new 96-well plate and mixed with 25 L of 5 mM ferric chloride. Absorbance
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
of the reaction mixture was read spectrophotometrically at 700 nm. The results were expressed as relative reducing equivalents of ascorbate.
2.3.4. H2O2 scavenging activity assay
H2O2 scavenging activity was performed as described previously (Wu et al.,
2011b). The reaction mixture contained 25 mM phosphate buffer (pH 7.4), 0.5 mM H2O2 and sample solution or Trolox standard in a total volume of 150 L. After
incubation for 5 min at RT, 50 L of 5 mM HVA and 8 U/mL HRPase were added, mixed and incubated for 25 min at RT. The fluorescence intensity was measured at an excitation of 315 nm and an emission of 425 nm using a fluorescence microplate reader (Bio-Tek, FLX800, Winooski, VT). The results were expressed as mol of trolox equivalents per gram of sample.
2.3.5. Hydroxyl radical scavenging activity assay
Hydroxyl radical scavenging activity was monitored using the 2-deoxyribose method (Halliwell et al., 1987). Briefly, the ferrous iron solution and EDTA were premixed, and then added to the assay mixture solution. The assay mixture solution contained 2.8 mM 2-deoxyribose, 20 M ferrous ion solution, 100 M EDTA and sample in a total volume of 1 mL of potassium phosphate buffer (10 mM, pH 7.4). The reaction was initiated by the addition of 1.42 M H2O2 and 100 M ascorbate,
and then incubated at 37°C for 30 min. At the end of the incubation period, 1% (w/v) TBA and 2.8% (w/v) TCA were added. This solution was heated for 30 min in a boiling water bath, cooled and measured at an absorbance of 532 nm, which is a maker for deoxyribose damage (Wu et al., 2011b). The reciprocal absorption values obtained for different concentrations were plotted against the concentrations of the
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
sample solution, and, from the graph, the second order rate constants were calculated with a rate constant of 3 ×109 M-1 S-1 (Halliwell et al., 1987).
2.4. ICA assay
This method is based on the formation of the ferrozine-Fe+2 colored complex which has a strong absorbance at 562 nm. The reaction mixture solution contained sample, 50 M ferrous sulfate and 300 M ferrozine. After incubation at 25 °C for 10 min, the absorbance was recorded. The percentage of inhibition of the ferronize-Fe+2 complex formation was calculated in the same way as in our previous report (Wu et al., 2011b).
2.5. Lipid peroxidation inhibition assay
Rat brain homogenate was used as a source of polyunsaturated fatty acids for determining the extent of lipid peroxidation. Whole brain was homogenized (100 mg/mL) in ice-cold 0.1 M phosphate buffer (pH 7.4), and then brain homogenate was centrifuged at 10,000 rpm for 15 min at 4 °C. The reaction mixture consisted of brain homogenate, 1 mM ferrous sulfate, 5 mM ascorbic acid and sample solution. The reaction solution was incubated in a shaking water bath in an open tube. After incubating at 37 °C for 30 min, the thiobarbituric acid reactive substance (TBARS) test was performed by rapidly adding 1.2% (w/v) TBA and 10% TCA. Then, the tubes were incubated at 90 °C for 60 min. After cooling, the system was centrifuged at 3,000 rpm for 10 min and the absorbance of the supernatant was determined at 532 nm (Wu et al., 2011b). The absorbance was recorded against blanks prepared in the same way as the experimental samples but without homogenate.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
2.6. Determination of the amount of antioxidant compounds 2.6.1. Determination of total phenol content
The total phenol content was assayed as described in our previous paper (Wu et al., 2011b). This method is based on the formation of blue-colored products by redox reaction with Folin-Ciocalteu’s reagent. The absorbance of the colored solution at 725 nm is proportional to the total phenol concentration. The total phenol concentration of the samples was expressed as mg of catechin equivalents per gram of sample.
2.6.2. Determination of flavonoid content
The flavonoid content was also assayed by a 96-well microtiter spectrophotometric method at 415 nm (Wu et al., 2011b). This method is based on the formation of colored products by flavonoids with aluminum salt. The absorbance of the colored solution is proportional to the total flavonoid concentration. The total flavonoid concentration of the samples was expressed as mg of quercetin equivalents per gram of sample.
2.6.3. Determination of phenylpropanoid content
The total phenylpropanoid content was determined by a 96-well microtiter spectrophotometric method at 525 nm (Wu et al., 2011b). Briefly, 150 L of sample solution was pipetted into each well, and then 50 L of 0.5 N HCl, 50 L of Arnow reagent (containing 5% (w/v) sodium nitrate and 5% sodium molybdate) and 50 L of 2N NaOH were added. After incubation at 25 C for 10 min, the absorbance was read. The absorbance of the colored solution is proportional to the total phenylpropanoid concentration. The total phenylpropanoid concentration of the samples was expressed as mg of verbascoside equivalents per g of sample.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
2.6.4. Determination of total iridoid content
The total iridoid concentration was determined by a 96-well microtiter
spectrophotometric method at 609 nm (Wu et al., 2011b). Twenty-five l of sample solution was pipetted into each well, and then 250 L of Trim-Hill reagent (acetic acid:0.2% CuSO4:HCl at a ratio of 10:1:0.5, respectively) was added. After heating at
100 C for 5 min, the absorbance was read. The absorbance of the colored solution is proportional to total iridoid concentration. The total iridoid concentration of the samples was expressed as mg of aucubin equivalents per g of sample.
2.6.5. Determination of triterpenoids and phenolic compounds by HPLC-DAD
FLL crude extract and fractions were dissolved in methanol and then filtered with a 0.22 m filter. Stock solutions of the standards were prepared in methanol to final concentrations of 1 mg/mL. All standard and sample solutions were injected into 10
L in triplicate. The Shimadzu VP series HPLC system and Shimadzu Class-VPTM chromatography data system were used. All chromatographic operations were carried out at 25 C. The chromatographic peaks of triterpenoids and phenolic compounds were confirmed by comparing their retention times and UV spectra.
A LiChrospher® RP-18e (250 × 4 mm, 5 m) column (Merck KGaA, Darmstadt, Germany) was used. Chromatographic separation of phenolic compounds, including HBA, hydroxytyrosol, oleuropein, PCA, salidroside, tyrosol, quercitrin and
verbascoside, was carried out using a two-solvent system. Solvent A is 100%
methanol, and solvent B is 0.2% acetic acid at pH=3.23. The analyses were performed by a gradient program. The conditions were as follows: initial condition of 90%
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
16-22 min changed to 60% solvent B, 22-30 min changed to 57% solvent B, 30-40 min changed to 60% solvent B and 40-42 min changed to 65% solvent B. Signals were detected at 280 nm. The concentrations used for the calibration of reference phenolic compounds were between 10 and 200 g/mL.
A Supelco Discovery® C18 (150 × 4.6 mm, 5 m) column (Sigma-Aldrich Co., USA) was used for triterpenoids, including betulin, betulinic acid, oleanolic acid and ursolic acid. The HPLC-DAD conditions were the same as in our previous report (Wu et al., 2011a). The concentrations used for the calibration of reference triterpenoid compounds were between 10 and 250 g/mL.
2.7. Protective effects of H2O2-induced neuronal damage in SH-SY5Y cells 2.7.1. Cell culture and treatment
Human neuroblastoma SH-SY5Y cells were cultured in DMEM supplemented with 10% fetal bovine serum, 100 units/mL penicillin and 100 g/mL streptomycin in a water-saturated atmosphere of 5% CO2 at 37 C. Experiments were carried out 24 h
after cells were seeded in 96-well sterile clear-bottom plates, 60 mm or 90 mm dish. Cells were plated at an appropriate density according to the scale of each experiment. H2O2 (1 mM) or vehicle, as control, was used in the study, described below, after a
16-h exposure. All samples were simultaneously treated with H2O2. The stock
solutions of FLL crude extract, EtOAc fraction and trolox were prepared using DMSO, filtered with a 0.22 M sterile filter and then diluted with DMEM without phenol red.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
The MTT assay was performed to study cell survival. This assay is based on the ability of living cells to reduce MTT to insoluble formazan. The amount of formazan produced reflects cell viability. Briefly, 16 h after treatment with H2O2, the medium
was replaced and MTT was added to each well. The cells were incubated for 2 h at 37C under a 5% CO2 atmosphere. The cells were washed with PBS, and DMSO was
added. After 30 min of stirring, the absorbance was measured at 570 nm using a microplate reader. The inhibition of MTT reduction indicates the degree of H2O2-induced toxicity to cells. Experiments were performed in triplicate in five
independent experiments. Cell viability was expressed as percentage relative to H2O2-untreated cells, which served as the control group (designated as 100% viable).
In other world, the results were expressed as percentage of the control.
2.7.3. Observation of morphologic changes and AO staining
SH-SY5Y cells were seeded at appropriate densities in a 60-mm dish and incubated overnight in a water-saturated atmosphere of 5% CO2 at 37C. All samples
were treated simultaneously with H2O2. Cellular morphology was observed 16 h after
H2O2 exposure using a phase-contrast microscope (Nikon, Tokyo). AO staining was
observed by fluorescence microscopy (Nikon, Tokyo). Viable cells have a bright green nucleus with an intact structure, while apoptotic cells exhibit a bright green nucleus showing chromatin condensation as dense green areas.
2.7.4. Measurement of caspase-3 activity
The activity of caspase-3 was determined using a caspase-3 fluorometric assay kit, and all measurements were carried out in triplicate in 96-well clear-bottom plates. Cells, at a density of 2 × 106 cells per 90 mm dish, treated with or without H
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
h were collected and centrifuged at 800 rpm for 5 min at 4 C. The pellet was
collected, washed with ice-cold PBS and lysed with 100 L of lysis buffer. The lysate was collected, sonicated and centrifuged at 12,000 rpm for 10 min at 4 C. Cell lysate (50 g protein) was added to each well. The peptide substrate for caspase-3
(Ac-DEVD-AMC) and assay buffer (20 mM HEPES, pH 7.5, 10% glycerol and 2 mM dithiothreitol) were added to start the reaction. Fluorescence was measured and
recorded at 360 nm excitation and 460 nm emission. Caspase activity was calculated as follows: caspase activity = [(mean AMC fluorescence from triplicate wells) – (background fluorescence)]/ g of protein.
2.7.5. Measurement of intracellular ROS levels
The intracellular ROS level was estimated using the fluorescent probe DCFH-DA. DCFH-DA readily diffuses through the cell membrane and is enzymatically
hydrolyzed by intracellular esterases to form non-fluorescent 2’,
7’-dichlorofluorescein (DCFH), which is then rapidly oxidized to form highly fluorescent 2’ ,7’-dichlorofluorescin (DCF) in the presence of intracellular ROS, especially H2O2. On the day of the experiment, SH-SY5Y cells, at a density of 2 × 104
cells/well, were washed 3 times with Krebs-Ringer-HEPES (KRH) buffer (120 mM NaCl, 4.7 mM KCl, 2.2 mM CaCl2, 10 mM HEPES, 1.2 mM KH2PO4 and 1.2 mM
MgSO4, pH 7.4) after removal of the medium, and then were incubated with
DCFH-DA in serum-free medium for 30 min at 37C under a 5% atmosphere. After DCFH-DA was removed, the cells were washed 3 times with KRH buffer again and incubated with DMEM without phenol red containing H2O2, FLL crude extract (300
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
cells in each well was measured and recorded at 485 nm excitation and 530 nm emission (Tsai et al., 2009). Data points were taken every 30 min for 2 h.
2.7.6. Measurement of intracellular GSH and GSSG levels
The GSH and GSSG levels were determined described previously (Tsai et al., 2009). Briefly, the lysates (20 g/50 L) and GSH standards were pipetted into each well of a 96-well plate. The reaction solution included 660 M DTNB, 900 M NADPH and 4.5 U/mL GR and, after addition to each well, was recorded at 405 nm for 5 min in a microplate reader. GSSG levels in cells were measured by the above method, except the lysates were pretreated with 2-VP and neutralized with
triethylamine to conjugate cellular GSH. The ratio of GSH/GSSG was calculated as follows: GSH/GSSG = [(total glutathione levels/ g of protein)-(GSSG levels/ g of protein)] / (GSSG/ g of protein)
2.7.7. Measurement of cellular antioxidant enzymes.
Following incubation for 16 h with H2O2, SH-SY5Y cells were collected from
culture dishes and sonicated on ice. The solution was centrifuged for 15 min at 4°C to eliminate cell debris, and the supernatant was used in enzyme activity assays.
Antioxidant enzyme activities including catalase, GPx, GR and SOD were measured as in our previous report (Tsai et al., 2009). Catalase and SOD activities were
expressed as units per mg of protein. The activities of GPx and GR were expressed as mU/mg of protein.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
Lipid peroxidation was measured with the TBARS assay in cell cultures. Briefly the lysates (200 g/100 L) and MDA standard were pipetted into 1.5 mL tubes, and a TBA test was performed by rapidly adding 1.2% (w/v) TBA and 10% TCA. Then, the tubes were incubated at 90 °C for 60 min. After cooling, the system was
centrifuged at 3,000 rpm for 10 min, and the absorbance of the supernatant was determined at 532 nm (Tsai et al., 2009). The experiments were performed in triplicate in three independent trials.
2.8. Statistical Analyses
All results were expressed as mean ± standard deviation (SD). Significant
differences were calculated in SPSS software with a one-way ANOVA followed by Scheffe’s test, and P values < 0.05 were considered to be significant.
3. Results and Discussion
3.1. Antioxidant and radical-scavenging activities
In this study, we first evaluated the free radical scavenging activity of FLL crude extract and fractions using the DPPH and TEAC assays because these assays are quick and common in vitro antioxidant activity assays. The IC50 values for FLL crude
extract in the DPPH and TEAC assays were 126.16 and 52.32 g/mL, respectively. The relative radical scavenging potency of FLL crude extract and all fractions in the DPPH method (using (+)-catechin) and TEAC assay (using trolox) is shown in Fig.
1(A) and 1 (B). The greatest scavenging capacity for both radicals was observed in
the EtOAc fraction, with the other fractions following in decreasing order: BuOH ≧ water > CHCl3. Therefore, the EtOAc fraction possessed the best free radical
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
scavenging properties. It is evident in the literature that the antioxidant activity of plants is associated with their reducing power, which terminates free radical chain reactions (Wu et al., 2011b; Zhu et al., 2009). Our results also showed that the most reducing power among all the fractions was observed in the EtOAc fraction, which is consistent with its free radical scavenging capacity (Fig. 1(C)). Therefore, our results are consistent with other reports that have found that the antioxidant capacity of natural plants is associated with their reducing power (Wu et al., 2011b; Zhu et al., 2009).
3.2. ROS scavenging and lipid peroxidation inhibiting activities
ROS, major pathological factors, cause many serious diseases, including
neurodegenerative and cardiovascular disorders (Fearon and Faux, 2009; Uttara et al., 2009; Valko et al., 2007). Among ROS, H2O2 readily crosses cellular membranes and
gives rise to the highly reactive hydroxyl radical, which is the most reactive and induces severe damage to adjacent biomolecules (Valko et al., 2007). Thus, H2O2 and
the hydroxyl radical scavenging activity of FLL crude extract and all fractions were investigated. The IC50 value for FLL crude extract in the H2O2 scavenging activity
assay was 140.59 g/mL. The highest H2O2 scavenging activity was observed in the
EtOAc fraction, followed by the BuOH, CHCl3 and water fractions (Table 1).
However, the highest second order rate constant for scavenging hydroxyl radical generated from the Fenton reaction was observed in the CHCl3 fraction (Table 1).
Because hydroxyl radicals cause lipid peroxidation by attacking polyunsaturated fatty acids such as brain tissues, we used rat brain homogenate as the oxidizable
biomolecule target for the Fe2+/ascorbate method to investigate the lipid
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
that the lowest IC50 value for the inhibition of Fe2+/ascorbate-induced lipid
peroxidation was also observed in the CHCl3 fraction. Because the activity of some
antioxidant compounds may be correlated with iron-chelation (Moran et al., 1997) when iron acts as a catalyst during lipid peroxidation (Valko et al., 2007), further investigation of the ICA activity of FLL crude extract and all fractions was carried out. FLL crude extract at the concentrations used in the hydroxyl radical scavenging and lipid peroxidation-inhibiting assay showed a 20.9% ion-chelating capacity. Among all the fractions, the water and CHCl3 fractions exhibited the highest ion-chelating
capacities (about 49.94 and 36.84%, Fig. 1(D)). These results indicate that the EtOAc fraction inhibited lipid peroxidation in brain homogenate systems by terminating oxidative chain reactions through its radical scavenging capacity; however, the inhibitory effects of the water and CHCl3 fractions on lipid peroxidation may be
partially related to iron chelation. Hence, we suggest that the antioxidant mechanism of the EtOAc fraction, a major antioxidant fraction from FLL, is possibly due to oxidative chain termination by radical-scavenging but not iron chelation.
3.3. Antioxidant components
Phenolic compounds, the main class of natural antioxidants, have been shown to be correlated with the antioxidant activities of plants (Wu et al., 2011b; Zhu et al., 2009). The most widespread and diverse phenolic compounds are flavonoids (Hernandez et al., 2009). Phenylpropanoids, intermediates in the biosynthesis of flavonoids and other phenolic compounds, also possess antioxidant activities (Korkina, 2007). Moreover, some researchers have indicated that secoiridoids and their glycosides are the active components of FLL that scavenge DPPH and APPH radicals (He et al., 2001; Li et al., 2007). Thus, by spectrophotometric methods, we first measured the
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
amount of total phenols, flavonoids, phenylpropanoid glycosides and iridoid
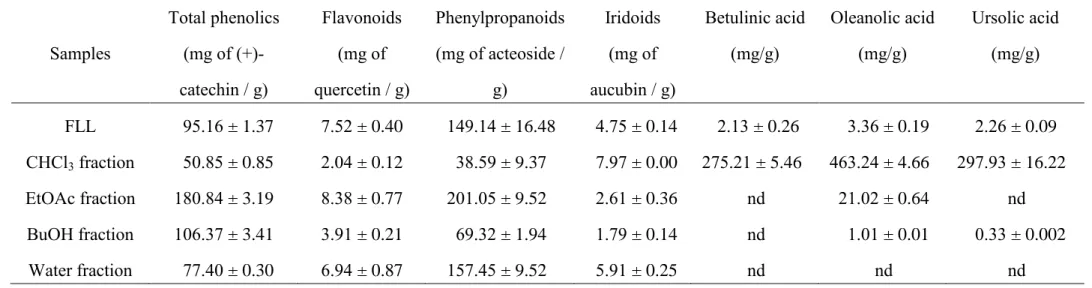
glycosides in FLL crude extract, which were equivalent to 95.16 mg of catechin, 7.52 mg of quercetin, 149.14 mg of verbascoside and 4.75 mg of aucubin, respectively. The EtOAc fraction had the highest content of phenolic compounds, including flavonoids and phenylpropanoid glycosides; however, the CHCl3 fraction had the
highest iridoid content among all the fractions (Table 2). Some reports have indicated that oleanolic acid may be an active component of FLL with respect to its antidiabetic and hepatoprotective effects (Gao et al., 2009; Yim et al., 2001). We also found that FLL crude extract contained 2.13 mg/g of betulinic acid, 3.36 mg/g of oleanolic acid and 2.26 mg/g of ursolic acid (Table 2). There is lack of betulin in FLL crude extract, which is inconsistent with our previous result from the leaves of Ligustrum lucidum (Wu et al., 2011a). Thus, we suggest that the various parts of Ligustrum lucidum contain different types of triterpenoids. The CHCl3 fraction had the higher amounts of
three triterpenoids than other fractions (Table 2). The relationship between
antioxidant potency parameters and the amount of antioxidants was calculated for all the FLL fractions by Pearson’s test. Among all the antioxidant assays, only DPPH, TEAC and reducing power showed high correlation coefficients (r=0.95, p < 0.05 for DPPH vs TEAC; r=0.98, p < 0.01 for DPPH vs reducing power; r=0.98, p < 0.01 for reducing power vs TEAC), suggesting that the antioxidant activities of FLL are closely related to their hydrogen-donor capacity. In agreement with other reports (Wu et al., 2011b; Zhu et al., 2009), our assay found total phenol content among all
phytochemical profiles to be positively and highly correlated with DPPH (r = 0.96), TEAC (r = 0.95) and reducing power (r = 0.96). However, the tendency in the FLL fractions to contain triterpenoids and iridoids is inconsistent with their antioxidant activities. Hence, we suggest that phenolic compounds (mainly flavonoids and
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
phenylpropanoids) are major determinants of these antioxidant activities from FLL, although some secoiridoid compounds isolated from FLL possess radical scavenging activity (He et al., 2001; Li et al., 2007). In fact, some flavonoid and phenylpropanoid compounds isolated from other Ligustrum plants possess antioxidant and
antimutagenic effects in vitro (Nagy et al., 2009; Zhu et al., 2009). Furthermore, our chromatographic result from HPLC-DAD showed that the EtOAc fraction mainly contained flavonoid derivatives, such as quercitrin (4.82±0.07 mg/g dry weight of EtOAc fraction), and phenol derivatives, such as hydroxytyrosol, PCA, salidroside, tyrosol, oleuropein and verbascoside (27.95±0.75, 9.03±0.24, 20.08±0.61, 12.28±0.32, 10.69±0.19, and 9.31±0.21 mg/g dry weight of EtOAc fraction, respectively) (Figure
2). Hydroxytyrosol and PCA were first identified in FLL, but all of the above
compounds identified in FLL with HPLC-DAD have been shown to possess
antioxidant activities (Di Benedetto et al., 2007; O'Dowd et al., 2004; Yu et al., 2007; Zhu et al., 2009). Hydroxytyrosol and salidroside content was highest in the EtOAc fraction. Therefore, the EtOAc fraction may be enriched in phenol, especially tyrosol-related derivatives, and possess the better antioxidant activity.
3.4. Protective effects of FLL crude extract, the EtOAc and CHCl3 fractions against H2O2-induced oxidative damage in SH-SY5Y cells
From the above results, we found that the EtOAc and CHCl3 fractions possessed
a higher H2O2 and hydroxyl radical scavenging potency than the other fractions. H2O2
is used extensively as an inducer of oxidative stress in vitro because its cellular actions and pathophysiological roles have been well studied. SH-SY5Y human neuroblastoma cells are highly sensitive to oxidative stressors such as H2O2.Hence,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
we evaluated the protective effects of FLL crude extract, the EtOAc and CHCl3
fractions against H2O2-induced cell injury in SH-SY5Y cells with the MTT assay. The
cell viability of SH-SY5Y cells incubated with 1 mM H2O2 for 16 h decreased to
12.4% compared with the control (Fig. 3(A)). FLL crude extract, the EtOAc and CHCl3 fractions were able to increase cell viability in a concentration-dependent
manner, acting against H2O2. The EtOAc fraction had the best protective effect
against H2O2, and the potency at 25 g/mL is equal to that of trolox at 200 M.
Furthermore, to confirm the protective effect of FLL crude extract and the EtOAc fraction against H2O2-induced cell injury, we observed morphological alterations by
phase-contrast microscopy. Incubation with 1 mM H2O2 for 16 h induced cell
shrinkage and a decrease in cell number. AO staining showed apoptotic
morphological changes, including chromatin condensation and nuclear fragmentation. FLL crude extract (300 g/mL) and the EtOAc fraction (50 g/mL) significantly alleviated these morphological manifestations and then reversed the apoptotic morphological changes (please see supplementary data). Because H2O2-induced
apoptosis occurs mainly through the apoptotic caspase pathway and includes
caspase-3 activation (Kim et al., 2000), we assayed the alteration of caspase-3 activity in SH-SY5Y cells treated with H2O2 plus either FLL crude extract or the EtOAc
fraction. Incubation with 1 mM H2O2 for 16 h increased intracellular caspase-3
activity. Only the EtOAc fraction significantly decreased intracellular capase-3 activity that was increased by H2O2 (Fig. 3(B)). Therefore, we suggest that the EtOAc
fraction protects against H2O2-induced cell injury and apoptosis in SH-SY5Y cells.
The active compounds within the EtOAc fraction acting against H2O2-induced
oxidative injury may be tyrosol-related compounds, mainly hydroxytyrosol,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
can scavenge H2O2 and protect against H2O2-induced oxidative cell death (O'Dowd et
al., 2004; Zhang et al., 2007; Zrelli et al., 2011b).
Cell oxidative injury is mainly due to an intracellular imbalance between the generation and elimination of free radicals, caused by the elevation of intracellular radical levels or a decrease in intracellular antioxidant defenses (Butterfield and Sultana, 2008; Valko et al., 2007). GSH, a major intracellular antioxidant, is depleted in intracellular oxidative stress caused by H2O2 and other oxidants (Han et al., 2003).
To clarify the protective mechanism of FLL crude extract and the EtOAc fraction against H2O2-induced cell injury in SH-SY5Y cells, we measured intracellular
oxidative stress by DCFH-DA and cellular antioxidant defenses such as GSH and antioxidant enzymes. In DCFH-DA-loaded SH-SY5Y cells, fluorescence intensity gradually increased after 1 mM H2O2 treatment, suggesting an increase in the
generation of intracellular ROS. When simultaneously treating with FLL crude extract or the EtOAc fraction, the oxidant burden of SH-SY5Y cells rapidly decreased (Fig.
3(C)). Otherwise, incubation with 1 mM H2O2 for 16 h decreased intracellular GSH
levels and the ratio of GSH/GSSG in SH-SY5Y cells (Fig. 4(A)-4(C)). It also
decreased the levels of cellular antioxidant enzymes, such as catalase, SOD, GPx and GR (Table 3), and caused an increase in the level of lipid peroxidation in SH-SY5Y cells (Fig. 3(D)). This phenomenon is consistent with our previous report (Tsai et al., 2009), which also identified the activation of the caspase pathway and apoptosis (Kim et al., 2000). FLL crude extract and the EtOAc fraction restored intracellular GSH levels, and the ratio of GSH/GSSG decreased by 1 mM H2O2 in SH-SY5Y cells (Fig.
3(A)-3(C)). They also reversed the activities of catalase, SOD, GPx and GR, which
decreased by 1 mM H2O2 in SH-SY5Y cells (Table 3). Only the EtOAc fraction
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
It is well known that the protective effects of antioxidants in biological systems are mainly due to their free radical-scavenging capacity, metal chelating activities and activation of biological antioxidant enzymes (Kulkarni et al., 2004). Hence, we suggest that FLL crude extract alleviated H2O2-induced oxidative damage via a
decrease in intracellular oxidative peroxide levels and an increase in the intracellular GSH levels and cellular antioxidant enzymes. This protective mechanism is the same as that of reports that demonstrate antidiabetic and hepatoprotective effects of FLL, which may be mediated by intracellular GSH regeneration and upregulation of
antioxidant status (Gao et al., 2009). The EtOAc fraction, a major active fraction from FLL, protected H2O2-induced oxidative damage and apoptosis in SH-SY5Y cells by
reversing the redox imbalance and blocking caspase-3 activation. Its major active compounds, such as hydroxytyrosol and salidroside, also have been shown to protect against H2O2-induced oxidative damage and apoptosis via the modulation of the
mitochondrial membrane, the downregulation of the pro-apoptotic gene Bax, the upregulation of the anti-apoptotic genes Bcl-2 and Bcl-X(L) and the activation of AKT, ERK and Nrf2 (Yu et al., 2010; Zhang et al., 2007; Zhu et al., 2011; Zrelli et al., 2011a).
4. Conclusion
In conclusion, EtOAc fraction from FLL has the highest phenol content,
especially hydroxytyrosol and salidroside, and exhibits the highest radical scavenging potency. Its antioxidant activity can be correlated with its reducing power but is not due to iron chelation, and this antioxidant activity depends on the phenolic
antioxidants but not the triterpenoids or secoiridoids in the fraction, as described in other reports (He et al., 2001; Li et al., 2007; Yim et al., 2001). The EtOAc fraction
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
has a protective effect against H2O2-induced oxidative injury and apoptosis in vitro,
and this protective mechanism might be related to its ability to decrease intracellular peroxides (via H2O2 scavenging) and caspase-3 activity, and to its ability to elevate
intracellular GSH levels and antioxidant enzyme activities (Fig. 5). Tyrosol-related compounds, including hydroxytyrosol, salidroside, tyrosol and oleuropein, are its major active compounds. Furthermore, these compounds could further protect neuronal damage caused by the amyloid peptide and MPP(+) (Li et al., 2011; St-Laurent-Thibault et al., 2011). Hence, we and others (Li et al., 2011;
St-Laurent-Thibault et al., 2011) have found evidence for the potential therapeutical benefits of FLL, especially its phenol enriched fraction, in treating aging-associated symptoms and neurodegenerative disorders.
Acknowledgments
We thank the financial support of the China Medical University CMU94-019 and CMU95-PH-02, Chinese Medicine Committee CCMP94-RD-013, and National Sciences Council NSC97-2320-B-039-041 and NSC98-2320-B-039-041 MY3.
References
Butterfield, D.A., Sultana, R., 2008. Redox proteomics: understanding oxidative stress in the progression of age-related neurodegenerative disorders. Expert Rev. Proteomics 5, 157-160.
Di Benedetto, R., Vari, R., Scazzocchio, B., Filesi, C., Santangelo, C., Giovannini, C., Matarrese, P., D'Archivio, M., Masella, R., 2007. Tyrosol, the major extra virgin
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
olive oil compound, restored intracellular antioxidant defences in spite of its weak antioxidative effectiveness. Nutr Metab Cardiovasc Dis 17, 535-545.
Fearon, I.M., Faux, S.P., 2009. Oxidative stress and cardiovascular disease: novel tools give (free) radical insight. J. Mol. Cell. Cardiol. 47, 372-381.
Gao, D., Li, Q., Li, Y., Liu, Z., Fan, Y., Zhao, H., Li, J., Han, Z., 2009. Antidiabetic and antioxidant effects of oleanolic acid from Ligustrum lucidum Ait in alloxan-induced diabetic rats. Phytother. Res. 23, 1257-1262.
Halliwell, B., Gutteridge, J.M., Aruoma, O.I., 1987. The deoxyribose method: a simple "test-tube" assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 165, 215-219.
Han, D., Canali, R., Rettori, D., Kaplowitz, N., 2003. Effect of glutathione depletion on sites and topology of superoxide and hydrogen peroxide production in mitochondria. Mol. Pharmacol. 64, 1136-1144.
Harman, D., 2006. Free radical theory of aging: an update: increasing the functional life span. Ann. N. Y. Acad. Sci. 1067, 10-21.
He, Z.D., But, P.P.H., Chan, T.W., Dong, H., Xu, H.X., Lau, C.P., Sun, H.D., 2001. Antioxidative glucosides from the fruits of Ligustrum lucidum. Chem. Pharm. Bull. 49, 780-784.
Hernandez, I., Alegre, L., Van Breusegem, F., Munne-Bosch, S., 2009. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 14, 125-132.
Kim, D.K., Cho, E.S., Um, H.D., 2000. Caspase-dependent and -independent events in apoptosis induced by hydrogen peroxide. Exp. Cell Res. 257, 82-88.
Korkina, L.G., 2007. Phenylpropanoids as naturally occurring antioxidants: from plant defense to human health. Cell Mol. Biol. 53, 15-25.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
Kulkarni, A.P., Aradhya, S.M., Divakar, S., 2004. Isolation and identification of a radical scavenging antioxidant - punicalagin from the pith and carpellary membrane of pomegranate fruit. Food Chem. 87, 551-557.
Li, X., Ye, X., Sun, X., Liang, Q., Tao, L., Kang, X., Chen, J., 2011. Salidroside protects against MPP(+)-induced apoptosis in PC12 cells by inhibiting the NO pathway. Brain Res 1382, 9-18.
Li, Y., Zuo, Y., Sun, W.J., 2007. Study on antioxidant activity of two major secoiridoid glucosides in the fruits of Ligustrum lucidum Ait. J. Chin. Med. Mater. 30, 543-546.
Moran, J.F., Klucas, R.V., Grayer, R.J., Abian, J., Becana, M., 1997. Complexes of iron with phenolic compounds from soybean nodules and other legume tissues: prooxidant and antioxidant properties. Free Radic. Biol. Med. 22, 861-870.
Nagy, M., Krizkova, L., Mucaji, P., Kontsekova, Z., Sersen, F., Krajcovic, J., 2009. Antimutagenic activity and radical scavenging activity of water infusions and phenolics from ligustrum plants leaves. Molecules 14, 509-518.
O'Dowd, Y., Driss, F., Dang, P.M., Elbim, C., Gougerot-Pocidalo, M.A., Pasquier, C., El-Benna, J., 2004. Antioxidant effect of hydroxytyrosol, a polyphenol from olive oil: scavenging of hydrogen peroxide but not superoxide anion produced by human neutrophils. Biochem Pharmacol 68, 2003-2008.
St-Laurent-Thibault, C., Arseneault, M., Longpre, F., Ramassamy, C., 2011. Tyrosol and Hydroxytyrosol, Two Main Components of Olive Oil, Protect N2a Cells against Amyloid-beta-Induced Toxicity. Involvement of the NF-kappaB Signaling. Curr Alzheimer Res.
Tsai, F.H., Lien, J.C., Lin, L.W., Chen, H.Y., Ching, H., Wu, C.R., 2009. Protective effect of Broussonetia papyrifera against hydrogen peroxide-induced oxidative stress in SH-SY5Y cells. Biosci. Biotechnol. Biochem. 73, 1933-1939.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Uttara, B., Singh, A.V., Zamboni, P., Mahajan, R.T., 2009. Oxidative stress and neurodegenerative diseases: a review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 7, 65-74.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M.T., Mazur, M., Telser, J., 2007. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 39, 44-84.
Wu, C.R., Hseu, Y.C., Lien, J.C., Lin, L.W., Lin, Y.T., Ching, H., 2011a. Triterpenoid contents and anti-inflammatory properties of the methanol extracts of ligustrum species leaves. Molecules 16, 1-15.
Wu, C.R., Lin, W.H., Hseu, Y.C., Lien, J.C., Lin, Y.T., Kuo, T.P., Ching, H., 2011b. Evaluation of the antioxidant activity of five endemic Ligustrum species leaves from Taiwan flora in vitro. Food Chem. 127, 564-571.
Yim, T.K., Wu, W.K., Pak, W.F., Ko, K.M., 2001. Hepatoprotective action of an oleanolic acid-enriched extract of Ligustrum lucidum fruits is mediated through an enhancement on hepatic glutathione regeneration capacity in mice. Phytother. Res. 15, 589-592.
Yu, P., Hu, C., Meehan, E.J., Chen, L., 2007. X-ray crystal structure and antioxidant activity of salidroside, a phenylethanoid glycoside. Chem Biodivers 4, 508-513. Yu, S., Shen, Y., Liu, J., Ding, F., 2010. Involvement of ERK1/2 pathway in
neuroprotection by salidroside against hydrogen peroxide-induced apoptotic cell death. J Mol Neurosci 40, 321-331.
Zhang, L., Yu, H., Sun, Y., Lin, X., Chen, B., Tan, C., Cao, G., Wang, Z., 2007. Protective effects of salidroside on hydrogen peroxide-induced apoptosis in SH-SY5Y human neuroblastoma cells. Eur J Pharmacol 564, 18-25.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
genotypes from Ilex kudingcha, Ilex cornuta, and Ligustrum robustum. J. Agric. Food Chem. 57, 6082-6089.
Zhu, Y., Shi, Y.P., Wu, D., Ji, Y.J., Wang, X., Chen, H.L., Wu, S.S., Huang, D.J., Jiang, W., 2011. Salidroside Protects Against Hydrogen Peroxide-Induced Injury in Cardiac H9c2 Cells via PI3K-Akt Dependent Pathway. DNA Cell Biol.
Zrelli, H., Matsuoka, M., Kitazaki, S., Araki, M., Kusunoki, M., Zarrouk, M., Miyazaki, H., 2011a. Hydroxytyrosol induces proliferation and cytoprotection against oxidative injury in vascular endothelial cells: role of Nrf2 activation and HO-1 induction. J Agric Food Chem 59, 4473-4482.
Zrelli, H., Matsuoka, M., Kitazaki, S., Zarrouk, M., Miyazaki, H., 2011b. Hydroxytyrosol reduces intracellular reactive oxygen species levels in vascular endothelial cells by upregulating catalase expression through the AMPK-FOXO3a pathway. Eur J Pharmacol 660, 275-282.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Legends for Figures
Figure 1. Antioxidant activity of crude extract and fractions from Fructus Ligustri Lucidi (FLL) on (A) DPPH assay, (B) TEAC assay, (C) reducing power assay and (D) ICA assay. Data are expressed as mean ± SD (n = 4).
Figure 2. HPLC chromatograms of the ethyl acetate (EtOAc) fraction from Fructus Ligustri Lucidi (FLL) at 280 nm. Trace: (A) Standard, (B) EtOAc fraction. 4-HC: 4-hydroxycoumarin as an internal standard.
Figure 3. Effect of crude extract, ethyl acetate (EtOAc) and chloroform (CHCl3)
fractions from Fructus Ligustri Lucidi (FLL) and trolox on (A) cell viability (MTT assay), (B) the activation of caspase-3, (C) time-course changes in the formation of reactive oxygen species (ROS) caused by H2O2. The SH-SY5Y cells were treated
with 1 mM H2O2 in the presence of these above samples for 16 h at 37 °C. Data are
expressed as mean ± SEM (n = 3). ** p < 0.01, *** p < 0.001, compared with H2O2
group.
Figure 4. Effects of crude extract, ethyl acetate (EtOAc) fraction from Fructus Ligustri Lucidi (FLL) and trolox on (A) cellular glutathione (GSH) levels, (B) oxidized glutathione (GSSG) levels, (C) ratio of GSH/GSSG, and (D) MDA levels. The SH-SY5Y cells were treated with 1 mM H2O2 in the presence of these above
samples for 16 h at 37 °C. Data are expressed as mean ± SEM (n = 3). * p < 0.05, ** p < 0.01, *** p < 0.001, compared with H2O2 group.
Figure 5. The biological action of Fructus Ligustri Lucidi partitioned with ethyl acetate as a potential antioxidant and protective plant against oxidative stress caused by hydrogen peroxide. Prohibition sign indicates that the inhibitory effect of Fructus Ligustri Lucidi.
Table 1. The H2O2, hydroxyl radical scavenging and lipid peroxidation-inhibition activities of crude
extract and fractions of Fructus Ligustrum Lucidi (FLL).
Samples H2O2 scavenging capacity (mol Trolox / g)
Second order rate constant in scavenging hydroxyl radical (×1010) IC50 in lipid peroxidation (mg / L) FLL 450.79 ± 10.87 2.37 ± 0.25 17.09 ± 0.56 CHCl3 fraction 602.80 ± 11.78 447.68 ± 1.33 9.64 ± 0.77 EtOAc fraction 981.40 ± 8.75 124.25 ± 0.98 16.51 ± 0.28 BuOH fraction 620.69 ± 4.10 2.54 ± 0.20 13.95 ± 0.34 Water fraction 404.01 ± 1.06 1.10 ± 0.15 17.40 ± 0.54 Data were expressed as mean ± SD (n = 3).
Table 2. The phytochemical composition of crude extract and fractions of Fructus Ligustrum Lucidi (FLL). Samples Total phenolics (mg of (+)-catechin / g) Flavonoids (mg of quercetin / g) Phenylpropanoids (mg of acteoside / g) Iridoids (mg of aucubin / g) Betulinic acid (mg/g) Oleanolic acid (mg/g) Ursolic acid (mg/g) FLL 95.16 ± 1.37 7.52 ± 0.40 149.14 ± 16.48 4.75 ± 0.14 2.13 ± 0.26 3.36 ± 0.19 2.26 ± 0.09 CHCl3 fraction 50.85 ± 0.85 2.04 ± 0.12 38.59 ± 9.37 7.97 ± 0.00 275.21 ± 5.46 463.24 ± 4.66 297.93 ± 16.22 EtOAc fraction 180.84 ± 3.19 8.38 ± 0.77 201.05 ± 9.52 2.61 ± 0.36 nd 21.02 ± 0.64 nd BuOH fraction 106.37 ± 3.41 3.91 ± 0.21 69.32 ± 1.94 1.79 ± 0.14 nd 1.01 ± 0.01 0.33 ± 0.002 Water fraction 77.40 ± 0.30 6.94 ± 0.87 157.45 ± 9.52 5.91 ± 0.25 nd nd nd Data were expressed as mean ± SD (n = 3).
Table 3. Effects of crude extract and ethyl acetate fraction (EtOAc) of Fructus Ligustrum Lucidi (FLL) on SOD, catalase, glutathione peroxidase
(GPx), and glutathione reductase (GR) activity in SH-SY5Y Cells exposed to 1 mM H2O2
Samples SOD (U/mg of protein) Catalase (U/mg of protein) GPx (mU/mg of protein) GR (mU/mg of protein)
Control 87.37 ± 7.75*** 8.25 ± 0.29*** 126.57 ± 7.62** 1.24 ± 0.09**
1 mM H2O2 36.01 ± 5.58 3.68 ± 0.12 57.72 ± 12.69 0.67 ± 0.04
FLL 300 g/ml + H2O2 84.24 ± 6.28*** 6.86 ± 0.48** 124.97 ± 18.09** 1.09 ± 0.11**
EtOAc 50 g/ml + H2O2 72.18 ± 3.49** 4.57 ± 0.50 168.27 ± 7.21*** 1.15 ± 0.04**
SH-SY5Y cells were treated with the above samples plus 1 mM H2O2. Data are expressed as mean ± SEM (n = 4). ** p < 0.01, *** p < 0.001 as
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Supplementary Material
Abstract
In this study, we demonstrated the antioxidant and protective properties of crude extract and fractions from Fructus Ligustri Lucidi (FLL) against hydrogen peroxide (H2O2)-induced oxidative damage in SH-SY5Y cells. The contents of the
phytochemical profiles from FLL crude extracts and fractions were determined by spectrophotometric methods and high performance liquid chromatography using a photodiode array detector. FLL crude extract possessed appreciable scavenging capacity against 1,1-diphenyl-2-picrylhydrazyl and H2O2. The ethyl acetate (EtOAc)
fraction was the most active fraction in scavenging free radicals and H2O2. Following
exposure of cells to H2O2, there was a marked decrease in cell survival and
intracellular antioxidant enzymes. Intracellular oxidative stress, the level of lipid peroxidation, and caspase-3 activity were increased when cells was exposed to H2O2.
Simultaneous treatment with the EtOAc fraction blocked these H2O2-induced cellular
events. Hydroxytyrosol and salidroside are the major components of the EtOAc fraction. These results show that the EtOAc fraction of FLL is enriched in phenol and contains tyrosol-related derivatives and that this fraction exerts protective effects against H2O2 toxicity via its free radical scavenging activity and ability to elevate the
levels of antioxidant enzymes.
Keywords: Fructus Ligustri Lucidi; hydroxytyrosol; salidroside; antioxidant activity; oxidative stress; antioxidant enzymes