子計畫六:肺正常和癌症組織中的轉移酵素表現和受
curcumin 的影響(2/3)
計畫類別: 整合型計畫 計畫編號: NSC94-2745-B-039-002-URD 執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 中國醫藥大學微生物學科 計畫主持人: 鍾景光 共同主持人: 張建國 計畫參與人員: 鍾景光,張建國 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 95 年 10 月 25 日
GADD153 mediated curcumin induced cell cycle arrest and apoptosis
in human lung carcinoma A-549 Cells through the elevation
of Ca
2+productions, decreasing of mitochondrial membrane potential and
promoted caspase-3 activation
Song-Shei Lin1, Hsuan-Pang Huang2, Jeng-Yuan Wu3, Shuw-Yuan Lin4, Te-Chun Hsia5, Jing-G.ung Chung6,7*
1Department of Radiological Technology, Central Taiwan Unuiversity of Science and Technology, No 11, Buzih Lane, Buzih District, Taichung 40605, Taiwan 3Department of Surgery, Armed Forces Taichung General Hospital, No.348, Sec. 2,
Chungshan Road, Taiping City, Taichung County 41168, Taiwan
4Department of Food and Nutrition, Hung-Kuang University, 34 Chung-Chie Road, Sha Lu, Taichung Hsien 433, Taiwan
5Department of Internal Medicine, China Medical University Hospital; Taichung, Taiwan
6Departments of Microbiology, 2School of Medical Laboratory Science and Biotechnology, 7School of Biological Science and Biotechnology, China Medical
University, No 91, Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C.
Lin et al: Curcumin induced G2/M arrest and apoptosis in human lung carcinoma cells
*Corresponding to Jing-Gung Chung, Ph. D., Department of Microbiology, School of
Medicine, China Medical University, No 91, Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C. Tel.: +886-4-2205-3366; fax: +886-4-2205-3764.
Abstract
Curcumin, a major component of the Curcuma species, is known to have antioxidant, anti-inflammatory properties and can induce cancer cells’ apoptosis. However, the precise molecular mechanisms of these effects on apoptosis in vitro are unclear. In this study, we showed that curcumin, a plant product containing the phenolic phytochemical, is a potent enhancer of Gadd153, and can cause mitochondria-dependent-induced apoptosis through the activation caspase-3 at treatment concentration of 30 µM in human lung cancer A-549 cells. In contrast, treatment with 5-10 µM of curcumin does not induce significant apoptosis, but rather triggers induced G2/M phase arrest in A-549 cells. We use flow cytometric analysis which indicated that curcumin directly increased intracellular oxidative stress based on the cell permeable dye 20,70-dichlorofluorescin diacetate (DCFH-DA) acting as an indicator of reactive oxygen species (ROS) generation. Using the dye Indo-1/AM as an indicator of Ca2+ generation, it showed that curcumin increase the levels of Ca2+ from ER. Using the dye DEioC6 as an indicator of mitochondrial membrane potential (∆Ψm), it demonstrated that curcumin decreased the levels of ∆Ψm in A-549 cells.
The result from Western blotting demonstrated that curcumin induced apoptosis from mitochondria-dependent through the activation of caspase 3- activation. Ca2+ also plays important role in the apoptosis pathway in curcumin treated cells because the A-549 cells were pretreated with the Ca2+ chelator (BAPTA) before leading to decreasing the apoptosis. Curcumin induced the production of ROS which caused ER stress based on the gene expression of Gadd153 from A-549 cells. Overall, our results demonstrated that curcumin dosage treatment determines the possible effect on ROS generation, ER stress, Ca2+ levels, ∆Ψm levels, caspase-3 activation and cell
Keywords: Curcumin; reactive oxygen species (ROS), Ca2+, mitochondrial membrane potential (∆Ψm), caspase-3, cell cycle arrest, apoptosis
1. Introduction
Cell cycle is a series of tightly integrated events which allow the cell to grow, proliferate and die, and it serves to protect the cell from genotoxic stress. Cell cycle can be divided into G0/G1-, S- and G2/M- phase based on the DNA content. The cyclins, cyclin-dependent kinases (CDKs) and even some of the inhibitors of cyclins and CDKs are involved in cell cycle. CDKs promoted the cell to move from one phase to the next pharse (G0/G1 to S, S to G2/M or G2 to M) of the cell cycle. If agents affect the cyclin and/or CDKs, it may lead to cell cycle arrest. If a cell continues to cycle with its damaged DNA intact, the apoptotic machinery is triggered and the cell will undergo apoptosis.
Numerous naturally occurring substances are recognized to be an antioxidants and cancer preventive agents or even as a cancer therapy drug such as taxol [1]. Those bioactive substances exert their anti-cancer activity through blocking cell cycle progression and triggering tumor cell apoptosis. Therefore, the best strategy for anticancer activity of agents is causing cell cycle arrest and the induction of apoptosis in tumor cells [2, 3].
Curcumin (diferuloylmethane), a phenolic compound from the plant Curcuma longa (Linn), is a widely used flavoring agent in food [4]. Curcumin inhibits cell proliferation and induces apoptosis in human leukemia, prostate cancer, and non-small cell lung cancer cell lines [5-7]. Curcumin has been shown to have antitumor activity in the colon, skin, stomach, duodenum, soft palate, and breasts of
rodents [8-10]. Recently published phase I clinical trial of oral curcumin in colon cancer patients showed high tolerability of the drug in humans and proposed a possibility of developing curcumin as an oral cancer preventive or therapeutic agent [11]. TNF and/or cigarette smoke were shown to activate NF- kappaB in human non-small cell lung cancer cells by phosphorylation and degradation of IkBalpha and the translocation of p65 subunit [12]. The anti-tumor activity of curcumin is attributed to its ability to induce apoptosis via caspase-3 activation [13].
Although there have been extensive investigations to elucidate the mechanism of action of curcumin as an antitumor agent, the exact mechanism is yet to be established. Therefore, in the present study, we focused on the effect of curcumin on lung cancer cells and also investigated the exact mechanism of curcumin induced apoptosis in human lung cancer A549 cells.
2. Materials and Methods
2.1 Chemical and reagents
Curcumin, propidium iodide (PI), triton X-100, Tris-HCl, trypan blue, ribonuclease-A were obtained from Sigma Chemical Co. (St. Louis, MO, USA). Potassium phosphates, dimethyl sulfoxide (DMSO), and TE buffer were purchased from Merck Co. (Darmstadt, Germany). 90% Ham’s F12K medium with 2 mM L-glutamine adjusted to contain 1.5 g/L sodium bicarbonate + 10% fetal bovine serum, penicillin-streptomycin, and trypsin-EDTA were obtained from Gibco BRL (Grand Island, NY, USA).
2.2 Human lung carcinoma A-549 cell line
Human lung carcinoma A-549 cell line was obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan). The A-549 cells were placed into 75 cm3 tissue culture flasks and grown at 37°C under a humidified 5% CO2 atmosphere in 90% Ham’s F12K medium with 2 mM L-glutamine ajusted to
contain 1.5 g/L sodium bicarbonate and supplemented with 10% fetal bovine serum (Gibco BRL, Grand Island, NY), and 2% penicillin-streptomycin (10,000 U/ml penicillin and 10 mg/ml streptomycin) [14].
2.3 Morphological changes and cell viability of A-549 cells treated with curcumin were determined by using pharse-contrast microscope and flow cytometry
The A-549 cells were plated in 12-well plates at a density of 2x105 cells/well and grown for 24 h. Different concentrations of curcumin were then added to cells for final concentration 0, 5, 10, 20, 30 and 50 µM, while only adding DMSO (solvent) for the control regimen and grown at 37°C, 5% CO2 and 95% air for a different period of time. To determine morphological changes and cell viability, the pharse-contrast microscope and the flow cytometric assay was used as described previously [14, 15].
2.4 Cell cycle and apoptosis of A-549 cells treated with different concentrations of curcumin were examined by using Flow cytometry analysis.
Approximately 5x105 cells/well of A-549 cells in 12-well plate with various concentrations (0, 5, 10, 20 30 and 50 µM, respectively) of curcumin were incubated in an incubator for different time periods. Cells were harvested by centrifugation
and were fixed gently (drop by drop) by putting 70% ethanol (in PBS) in 4°C overnight and then re-suspended in PBS containing 40 µg/mL PI and 0.1 mg/mL RNase and 0.1% Triton X-100 in dark room. After 30 minutes at 37°C, the cells were analyzed with a flow-cytometry (Becton-Dickinson, San Jose, CA, USA) equipped with an argon ion laser at 488 nm wavelength. Then the cell cycle was determined and analyzed [13]. Annexin V-FITC and PI double staining kit from PharMingen (San Diego, CA) were used for apoptotic cells quantified [14-16].
2.5 Apoptosis of A-549 cells treated with different concentrations of curcumin were examined by using DAPI staining
Approximately 2x105 cells/well of A-549 cells in 12-well plate with 30 µΜ curcumin were incubated in an incubator for 48 hours. Cells were stained by DAPI, then photographed by fluorescence microscope as described previously [14-16].
2.6. Comet assay examining the curcumin-induced DNA damage of A-549 cells.
Approximately 5x103 cells/ml cells of A-549 were treated with or without various concentrations of curcumin at 0, 5, 10, 20, 30 and 50 µM for 24 hours then isolated for examining DNA damage by using the Comet assay described previously [14].
2.7 Caspase-3 activity determination of A-549 cells treated with curcumin and/or cells were pretreated with caspase-3 inhibitor (Ac-DEVD-CHO) then with curcumin.
Approximately 5x105 cells/well of A-549 cells in 12-well plate with various concentrations (0, 5, 10, 20 30 and 50 µM) of curcumin were incubated in an incubator for different time periods. Cells were harvested by centrifugation and the mediums were removed. Then, 50 µL of 10 µM substrate solution (PhiPhilux is a unique class of substrates for caspase-3) was added to the cell pellet (1x105 cells per
sample) or cells were pretreated with caspase-3 inhibitor (Ac-DEVD-CHO) for 3 hours, then treated with 30 µM curcumin. Cells then were incubated at 37°C for 60 minutes. Then the cells were washed once by adding 1 mL of ice cold PBS and re-suspend in fresh 1 mL. Cells were analyzed with a flow-cytometry (Becton-Dickinson, San Jose, CA, USA) equipped with an argonion laser at 488 nm wavelength. Then the caspase-3 activity was determined and analyzed [14]. The MMP and apoptosis were determinated [17].
2.8. Detection of reactive oxygen species (ROS) in A-549 cells after treated with curcumin by flow cytometry
Approximately 5x105 cells/well of A-549 cells in 12-well plate with various 30 µM curcumin were incubated for 15, 30, 45, 60 and 120 minutes to detect the changes of ROS. The cells were harvested and washed twice, re-suspended in 500 µL of 2,7-Dichlorodihydrofluorescein diacetate (DCFH-DA)(10 µM) and incubated at 37°C for 30 min and analyzed by flow cytometry [14-16].
2.9. Detection of mitochondrial membrane potential (∆Ψm) in A-549 cells after treated
with curcumin by flow cytometry
Approximately 5x105 cells/well of A-549 cells in 12-well plate with various concentrations (0, 5, 10, 20, 30 or 40 µM, respectively) of curcumin were incubated for 24 h to detect the changes of ∆Ψm. The cells were harvested and washed twice,
re-suspended in 500 µL of DiOC6 (4 mol/L) and incubated at 37°C for 30 min and
2.10. Detection of Ca2+ concentrations in A-549 cells after treated with curcumin by flow cytometry
Approximately 2x105 cells/well of A-549 cells in 12-well plate with 30 µM curcumin were incubated for 6, 12, 24 and 48 h to detect the changes of Ca2+ concentration. The cells were harvested and washed twice, once for apoptosis analysis and the other underwent re-suspendion in Indo 1/AM (3 µg/ml) and was incubated at 37°C for 30 min and analyzed by flow cytometry [14-16].
2.11. Detection of Ca2+ concentrations and MMP levels and apoptosis in A-549 cells
after pre-treated with BAPTA then treated with curcumin.
The level of Ca2+, ∆Ψm and apoptosis of the A-549 cells was determined by flow
cytometry (Becton Dickinson FACS Calibur), using the Indo 1/AM and DiOC6
(Calbiochem; La Jolla, CA) and PI, respectively. About 5x105 cells/well of A-549 cells in 12-well plate were pre-treated with BAPTA before adding 30 µM of curcumin for incubation for 24 h to detect the changes of Ca+2 concentration and the levels of
∆Ψm and percentage of apoptosis. The cells were harvested and washed twice, once
for apoptosis analysis and the other for re-suspension in Indo 1/AM (3 µg/ml) and DiOC6(4 mol/L) before being incubated at 37°C for 30 min and analyzed by flow
cytometry [15-16].
2.12. Curcumin affected the expressions of Wee1, CHK1, Cdc25c, p53, p21, Cdc2, Cyclin B1,cspase-3 and -9, Bcl-2, COX-1 and -2, Naf-kB, catalase, SOD(Zn and Cu), MMP-2 and -9 in A-549 cells.
The total proteins were collected from A-549 cells treated with 30 µM for 0, 6, 12, 24 and 48 h before Wee1, CHK1, Cdc25c, p53, p21, Cdc2, Cyclin B1,cspase-3 and -9, Bcl-2, COX-1 and -2, Naf-kB, catalase, SOD(Zn and Cu), MMP-2 and -9 were measured by sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot as described previously [14-17].
2.13. Curcumin affects the levels of Gadd153 mRNA in A-549 cells.
The total RNA was extracted from the A-549 cells after treatment 30 µM curcumin for 1, 3, 6 and 12 h, using the Qiagen RNeasy Mini Kit as described previously [18-20]. The PCR reactions were performed with the following primers, as described previously (Chung et al., 2001, Tang et al., 2006). The sequences of the primers were: Gadd153: 5V-GAAACGGAAACAGAGTGGTCATTCCCC-3V (sense) and 5V-GTGGGATTGAGGGTCACATCATTGGCA-3V (antisense) to produce a 301-bp DNA fragment of Gadd153. In each PCR, an internal control using the primers was: GAPDH: 5V-CTCAGACACCATGGGGAAGGTGA-3V (sense) and 5V-ATGATCTTGAGGCTGTTGTCATA-3V (antisense) to produce a 450-bp fragment of the GAPDH gene that was included. Each assay was conducted at least twice to ensure reproducibility [18-20].
2.14. Curcumin affects the migration of A-549 cells.
The 2x105 cells/well of A-549 cells in 10 cm-plate with 30 µM curcumin were incubated for 12 and 21 h to detect the migration of A-549 cell as described previously [14] .
Student t-test was used to analyze the statistical analysis between the capsaicin treated and control groups.
3. Results
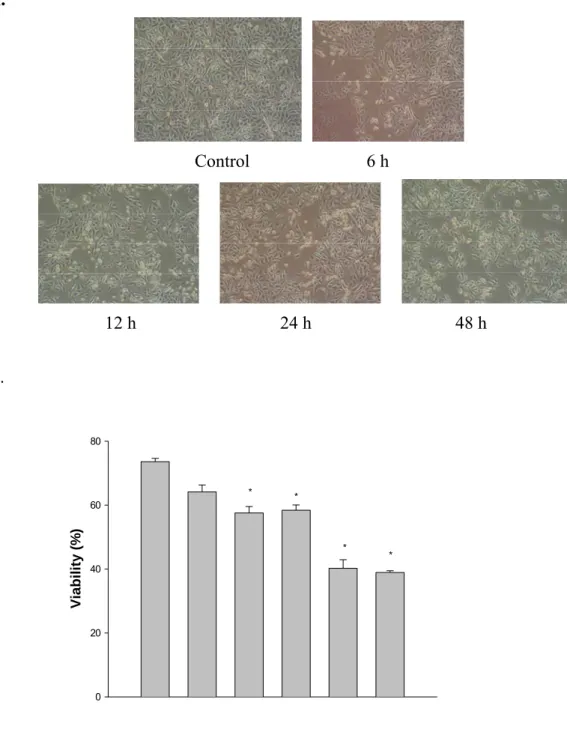
3.1 Ccurcumin induced morphological changes and decreased the percentage of viable A-549 cells.
After A-549 cells were treated with or without various concentrations of curcumin for various time periods, morphological changes were photographed by using pharse-contrast microscope (Fig. 1A) and viable cells were examined by flow cytometric methods. The results demonstrated that curcumin induced morphological changes (Fig. 1A) and decreased the percentage of viable cells (Fig. 1B) and is of a time-dependent (Fig. 1A) and dose-dependent (Fig. 1B) manner.
3.2 Curcumin induced cell cycle arrest and apoptosis in A-549 cells.
A-549 cells were treated with or without various concentrations of curcumin for various time periods before cell cycle and apoptosis (sub-G1 group) were examined and analyzed by flow cytometric methods. The results demonstrated that curcumin induced G2/M phase arrest (Fig. 2A) (dose-dependent effects) and apoptosis (Fig. 2B) (time-dependent manner).
DAPI staining.
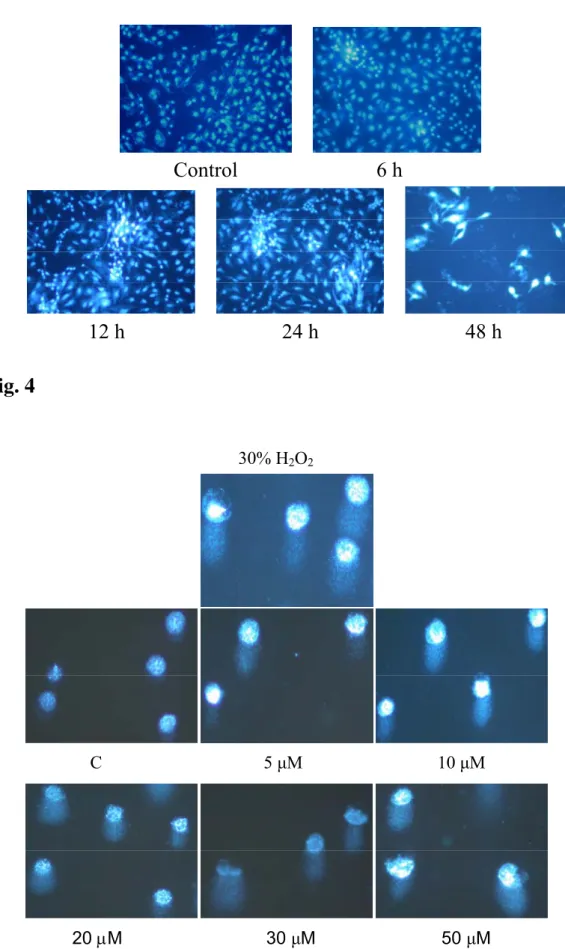
Induction of apoptosis from A-549 cells after treated with curcumin that were determined. Apoptosis was detected by DAPI staining method after 6, 12, 24 and 48 hours of continuous exposure to curcumin before being photographed by fluorescence microscope. As shown in Figure 3, curcumin induced apoptosis in a time-dependent manner.
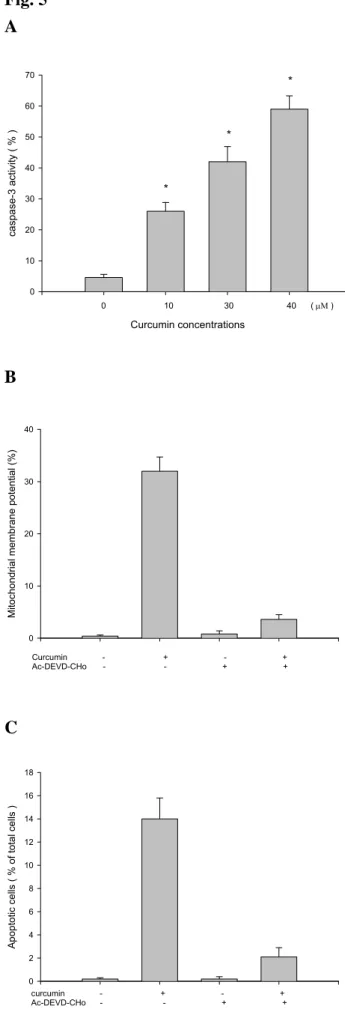
3.4. Curcumin-induced DNA damage in A-549 cells was examined by Comet assay.
The Comet assay examining showed that curcumin-induced DNA damage in A-549 cells based on the smear DNA migration. The higher concentrations of curcumin led to longer DNA migration smear (Fig. 4).
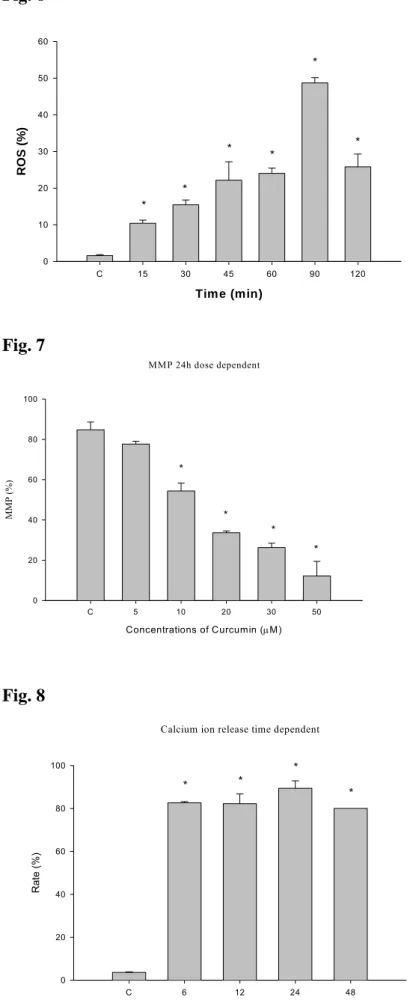
3.5 Curcumin promoted the caspase-3 activity and caspase-3 inhibitor
(Ac-DEVD-CHO) inhibits caspase-3 activity in A-549 cells.
After A-549 cells were pre-treated with or without caspase-3 inhibitor (Ac-DEVD-CHO) they were then treated with various concentrations of curaumin for 24 h before caspase-3 activity was analyzed by flow cytometric methods. The results demonstrated that curcumin promoted the activation of caspase-3 (Fig. 5A) and those effects are dose-dependent manner. The inhibitor of caspase-3 (Ac-DEVD-CHO) decreased the levels of MMP (Fig. 5B) and percentage of apoptosis (Fig. 5C).
cells.
After A-549 cells were treated with or without 30 µM curcumin for various time periods, ROS productions were analyzed and quantitated by flow cytometric methods. The results demonstrated that curcumin promoted the levels of ROS and those effects are time-dependent manner (Fig 6).
3.7 Curcumin suppressed the levels of mitochondria membrane potential (∆Ψm)
inA-549 cells.
After A-549 cells were treated with various concentrations of curcumin for 12 h, the levels of ∆Ψm were analyzed and quantitied by flow cytometric methods. The
results demonstrated that curcumin increasedin the loss of mitochondrial ∆Ψm and
those effects are dose-dependent manner (Fig. 7).
3.8. Curcumin induced the Ca2+ production in A-549 cells.
After A-549 cells were treated with or without 30 µM curcumin for 6, 12, 24 and 48 h, Ca2+ productions were analyzed by flow cytometric methods. The results demonstrated that curcumin promoted the Ca2+ productions and those effects are
time-dependent manner (Fig. 8).
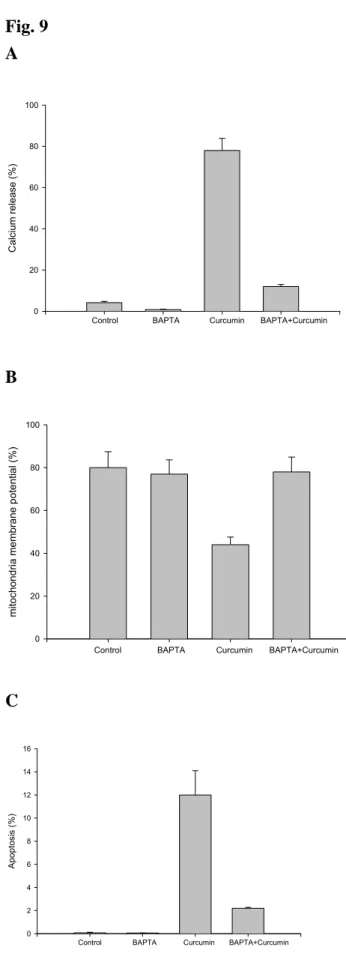
3.9. Effects of calcium antagonist BAPTA (Ca2+ chelator) on curcumin affect the levels of Ca2+, MMP and apoptosis in A-549 cells
treated with 30 µM curcumin for 24 h, while Ca2+ productions, levels of MMP and apoptosis were analyzed by flow cytometric methods. The results demonstrated that BAPTA decreased curcumin-induced Ca2+ productions (Fig. 9A), decreased the levels
of MMP (Fig. 9B) and also decreased the percentage of apoptosis (Fig. 9C).
3.10. Curcumin affected the expressions of Wee1, CHK1, Cdc25c, p53, p21, Cdc2, Cyclin B1,cspase-3 and -9, Bcl-2, COX-1 and -2, Naf-kB, catalase, SOD(Zn and Cu), MMP-2 and -9 in A-549 cells.
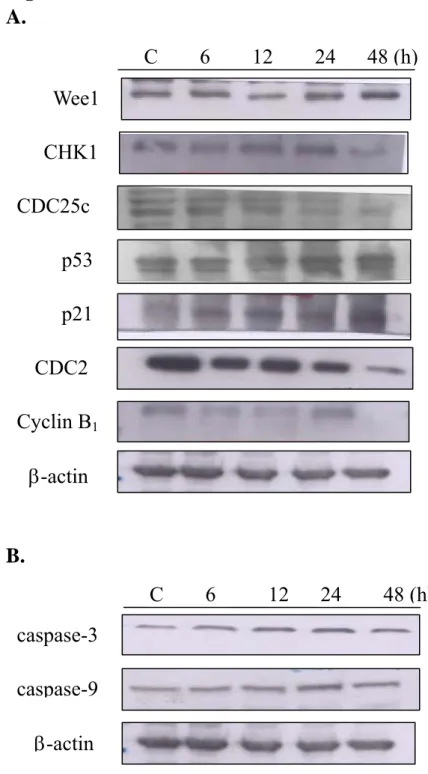
In order to characterize the molecular mechanism of curcumin-inducedcell cycle arrest and apoptosis in A-549 cells, we examinedthe expressions of cell cycle- and apoptosis-associated proteins during the treatment with curcumin by Western blotting. The protein levels of Wee1, CHK1, Cdc25c, p53, p21, Cdc2, Cyclin B1 (Fig. 10A), caspase-3 and-9 (Fig. 10B), Bcl-2, COX-1 and -2, Naf-kB, catalase, SOD(Zn and Cu) (Fig. 10C), MMP-2 and-9 (Fig. 10D) from Western blot indicate that the levels of CHK1,CDC25c, CDC2, cyclin B1, Bcl-2, NF-kaapB, catalase, MMP-2 and-9 that were decreased and the levels of Wee1, p53, p21, caspase-3 and-9, COX-2 that were increased by curcumin may have led to G2/M arrests and apoptosis.
3.11. Curcumin inhibited Gadd153mRNA expression in A-549 cells.
The results from PCR indicated that curcumin treated A-549 cells led to the levels of mRNA Gadd153 increased (Fig 11A and B). These effects are time-dependent manner.
The results from pharse-contrast microscope showed that curcumin inhibited the migration of A-549 cells and these effects are time-dependent (Fig. 12).
4. Discussion
Although many studies had demonstrated that curcumin induced cell cycle arrest and apoptosis in human cancer cell lines, the exact role of Gadd153 on the induction of apoptosis by curcumin is still unclear. Therefore, in the present study we focused on the role of Gadd153 associated apoptosis which from human lung cancer A-549 cells after exposed to curcumin. Overall our results showed that curcumin induced G2/M arrest and apoptosis in A-549 cells and this is in agreement with other reports that demonstrated that curcumin induces G2/M phase arrest and apoptosis in examined human melanoma cells [21], bladder cancer T24 cells [22] and gastric and colon cancer cells [23]. Our data also showed that curcumin induced apoptosis through decreasing the levels of ∆Ψm, cytochrome c release and promoted caspase-3
activation. This is in agreement with other reports which showed that curcumin induced apoptosis through the activation of caspase-3 [23].
It is well known that normal cells maintain the cell in cell cycle and the cell cycle checkpoints are important control mechanisms that ensure the proper execution of cell cycle events. CDKs are the major molecular players with cyclins in cell cycle progression [24]. If agent inhibits the CDK activity or affects the levels of CDK, it could cause cell cycle arrest that had been demonstrated to be the important strategy for the potential of novel anticancer drugs [24]. CDK inhibitors (flavopiridol, UCN-01, olomoucine, rocosvitine, butyrolactone I, indirubin-5-sulfonate, and indirubin-3′-monoxime) may inhibit the growth of cancer cells by causing cell cycle
arrest and apoptosis [24]. Those agents can induce DNA damage and if the CDKs are inhibited, both could lead to the cell undergoing cell-cycle arrest that may provide the cell the opportunity to repair its own damaged DNA before it resumes cell proliferation. Furthermore, if the cells continue to cycle with its damaged DNA intact, it may trigger the apoptotic machinery and the cell will undergo apoptosis. Our results also demonstrated that curcumin induced DNA damage based on the Comet assay as showed in Figure 4. Promote apoptosis of DNA damaged tumor cells could be performed either during cell cycle arrest or following premature cell cycle checkpoint exit that tumor cells re-enter the cell cycle before DNA repair is complete [24].
Our data also showed that curcumin induced ROS production, Ca2+ release and decreased the levels of ∆Ψm and promoted the cytochrome c release from
mitochondria then followed by activation of caspase-3 before apoptosis occur. High levels of ROS not only induced cell death [25] but also cause DNA damage and genomic instability [26, 27] that may cause tumorigenesis. The reason is that in certain tumor cells, there exists higher levels of ROS [28] but low levels of ROS enhance cell proliferation [29]. We also found that curcumin induced the levels of Gadd153 in earlier treated time (Fig. 11). So it may be that curcumin induced ROS production followed by ER stress and DNA damage which caused Ca2+ released then also promoted p53 levels leading to Bax increasing and decreasing the levels of Bcl-2 followed by decreased levels of ∆Ψm in mitochondria (Fig. 13). Much evidence had
showed that mitochondrion plays an important role in the regulation of apoptosis [30, 31]. Mitochondrial dysfunctions including the loss of mitochondrial membrane potential (∆Ψm), permeability transition, and release of cytochrome c from the
mitochondrion into the cytosol are associated with apoptosis [32]. It is also reported that agents induced apoptosis also promoted Gadd153 levels in earlier time [33].
We also found out that A-549 cells pretreated with BAPTA (a chelator of Ca2+) then followed by treatment with curcumin could decrease the percentage of apoptosis which was induced by curcumin. The result also showed BAPTA (a celator of Ca2+) could decrease the levels of ∆Ψm in examined cells. Apparently Ca2+ was involved
in curcumin induced apoptosis pathway and it is mitochondria-dependent.
In mammalian cells, apoptosis pathway can be divided into caspases- dependent and -independent pathway and caspases plays an important role in the initiation of apoptosis [36]. Caspases involving apoptosis can be grouped into "apoptotic initiator," such as caspase-8, and "apoptotic effector," such as caspase-3 based on the substrate specificities and target proteins of caspases [30]. In this study, curcumin induced apoptosis was observed and via ER stress which led to mitochondria dysfunction due to the curcumin increasing the levels of Bax (pro-apoptotic protein) and decreasing the levels of Bcl-2 which is a anti-apoptotic protein, and those effects are followed by the activation of caspase-3. Caspase-3 activity was analyzed by flow cytometric analysis (Fig. 5). Curcumin induced DNA damage in examined A-549 cells (Fig. 4) was based on the Comet assay and this damage caused increased levels of P53 and p21 caused the effects of Cdc2 and cyclin B and curcumin promoted the Wee1 CHK2 and cdc25c then led to G2/M arrest. Our results from Western blot also showed that curcumin induced the increase of p53 levels in A-549 cells. It is known that p53 protein can activate the transcription of a variety of apoptosis-associated genes, and to program cell death in response to genotoxic stresses [34, 35]. Therefore, understanding of the cell cycle and its relationship to p53 are critical for the successful clinical development of these agents for the treatment of patients with lung cancers.
These actions (curcumin promoted caspase-3 activation) showed a time-subsequent and a dose-corresponding manner. We also showed that the added
caspase-3 inhibitor, Ac-DEVD-CHO, and inhibited curcumin-induced caspase-3 activity led to decrease the apoptosis in these examined cells (Fig. 5). In conclusion, these results demonstrated that curcumin-induced G2/M arrest through DNA damage led to increase the levels of P53 and p21 which affect CDC2 and cyclin B and promoted the levels of Wee1 and CHK2 which affect Cdc25c before leading to G2/M arrest (Fig. 13). Curcumin induced apoptosis in A-549 cells through the mitochondria dysfunction leading to the release of cytochrome c and involves a caspase-3-mediated mechanism (Fig. 13).
Acknowledgement
This work was supported by grant NSC 94-2745-B-039-002-URD from the National Science Council of Taiwan, Taiwan, R.O.C.
References
[1] Hassan MA, Braam SR, Kruyt FA. Paclitaxel and vincristine potentiate adenoviral oncolysis that is associated with cell cycle and apoptosis modulation, whereas they differentially affect the viral life cycle in non-small-cell lung cancer cells. Cancer Gene Ther 2006; Jul 14 [Epub ahead of print]
[2] Paschka AG, Butler R, Young CY: Induction of apoptosis in prostate cancer cell lines by the green tea component, (-)-epigallocatechin-3-gallate, Cancer Lett 1998;130:1-7.
[3] Smets LA: Programmed cell death (apoptosis) and response to anti-cancer drugs. Anticancer Drugs 1994;5:3-9.
[4] Huang MT, Lou YR, Xie JG, Ma W, Lu YP, Yen P, Zhu BT, Newmark H, Ho CT: Effect of dietary curcumin and dibenzoylmethane on formation of 7,12-dimethylbenz[a]anthracene- induced mammary tumors and lymphomas/leukemias in Sencar mice. Carcinogenesis 1998;19:1697–1700.
[5] Guatam SC, Xu YX, Pindolia KR, Janakiraman N, Chapman RA: Nonselective inhibition of proliferation of transformed and transformed cells by anticancer agent curcumin (diferuloylmethane). Biochem Pharmacol 1998;55:1333–1337. [6] Dorai T, Cao YC, Dorai B, Buttyan R, Katz AE: Therapeutic potential of
curcumin in human prostate cancer. III. Curcumin inhibits proliferation, induces apoptosis, and inhibits angiogenesis of LNCaP prostate cancer cells in vivo. Prostate 2001;47:293–303.
[7] Pillai GS, Srivastava AS, Hassanein TI, Chauhan DP, Carrier E: Induction of apoptosis in human lung cancer cells by curcumin. Cancer Lett 2004;208: 163–170.
[8] Rao CV, Rivenson A, Simi B, Reddy BS: Chemoprevention of colon carcinogenesis by dietary curcumin, a naturally occurring plant phenolic compound. Cancer Res 1995;55:259–266.
[9] Kawamori T, Lubet R, Steele VE, Kelloff GJ, Kaskey RB, Rao CV, Reddy BS: Chemopreventive effect of curcumin, anaturally occurring anti-inflammatory agent, during the promotion/progression stages of colon cancer. Cancer Res 1999;59:597–601.
[10] Verma SP, Salamone E, Goldin B: Curcumin and genistein, plant natural products, show synergistic inhibitory effects on the growth of human breast cancer MCF-7 cells induced by estrogenic pesticides. Biochem Biophys Res Commun 1997; 233:692–696.
[11] Sharma RA, Euden SA, Platton SL, Cooke DN, Shafayat A, Hewitt HR, Marczylo TH, Morgan B, Hemingway D, Plummer SM, Pirmohamed M, Gescher AJ, Steward WP: Phase I clinical trial of oral curcumin: biomarkers of
systemic activity and compliance. Clin Cancer Res 2004;10:6847–6854.
[12] Shishodia S, Potdar P, Gairola C, Aggarwal BB: Curcumin (diferuloylmethane) down-regulates cigarette smoke-induced NFjB activation through inhibition of IjBa kinase in human lung epithelial cells: correlation with suppression of COX-2, MMP-9 and cyclin D1. Carcinogenesis 2003;24:1269–1279.
[13] Khar A, Ali AM, Pardhasaradhi, BV, Begum Z, Anjum R: Antitumor activity of curcumin is mediated through the ininduction of apoptosis in AK-5 tumor cells. FEBS Lett 1999;445:165–168
[14]Yang JH, Hsia TC, Kuo HM, Chao PD, Chou CC, Wei YH, Chung JG.: Inhibition of lung cancer cell growth by quercetin glucuronides via G2/M arrest and induction of apoptosis. Drug Metab Dispos 2006;34:296-304.
[15] Wu CC, Lin JP, Yang JS, Chou ST, Chen SC, Lin YT, Lin HL, Chung JG. Capsaicin induced cell cycle arrest and apoptosis in human esophagus epidermoid carcinoma CE 81T/VGH cells through the elevation of intracellular reactive oxygen species and Ca(2+) productions and caspase-3 activation. Mutat Res 2006; Aug 28[Epub ahead of print]
[16] Lin CC, Lin SY, Chung JG, Lin JP, Chen GW, Kao ST: Down-regulation of cyclin B1 and up-regulation of Wee1 by berberine promotes entry of leukemia cells into the G2/M-phase of the cell cycle. Anticancer Res 2006;26:1097-1104 [17] Tsou MF, Lu HF, Chen SC, Wu LT, Chen YS, Kuo HM, Lin SS, Chung JG.
Involvement of Bax, Bcl-2, Ca2+ and caspase-3 in capsaicin-induced apoptosis of human leukemia HL-60 cells. Anticancer Res 2006;26:1965-1971.
[18] Chung JG, Yeh KT, Wu SL, Hsu NY, Chen GW, Yeh YW, Ho HC: Novel transmembrane GTPase of non-small cell lung cancer identified by mRNA differential display. Cancer Res 2001;61:8873-8879.
[19] Tang NY, Yang JS, Chang YH, Lu HF, Hsia TC, Lin WC, Chung JG: Effects of wogonin on the levels of cytokines and functions of leukocytes associated with NF-kappa B expression in Sprague-Dawley rats. In Vivo 2006;20:527-32. [20] Yu CS, Yu FS, Chan JK, Li TM, Lin SS, Chen SC, Hsia TC, Chang YH, Chung
JG: Aloe-emodin affects the levels of cytokines and functions of leukocytes from Sprague-Dawley rats. In Vivo 2006;20:505-510.
[21] Zheng M, Ekmekcioglu S, Walch ET, Tang CH, Grimm EA: Inhibition of nuclear factor-kappaB and nitric oxide by curcumin induces G2/M cell cycle arrest and apoptosis in human melanoma cells. Melanoma Res 2004;14:165-171.
[22] Park C, Kim GY, Kim GD, Choi BT, Park YM, Choi YH: Induction of G2/M arrest and inhibition of cyclooxygenase-2 activity by curcumin in human bladder cancer T24 cells. Oncol Rep 2006;15:1225-1231.
cycle and apoptosis in gastric and colon cancer cells. Anticancer Res 2001 ;21:873-878.
[24] Buolamwini JK: Cell cycle molecular targets in novel anticancer drug discovery. Curr Pharm Des 2000;6:379-392.
[25] Droge W: Free radicals in the physiological control of cell function. Physiol Rev 2002;82:47-95.
[26] Cerutti PA: Oxy-radicals and cancer. Lancet 1994;344:862-863.
[27] Wiseman H, Halliwell B: Damage to DNA by reactive oxygen and nitrogen species: role in inflammatory disease and progression to cancer. Biochem J 1996;313:17-29.
[28] Szatrowski TP, Nathan CF: Production of large amounts of hydrogen peroxide by human tumor cells. Cancer Res 1991;51:794-798.
[29] Benhar M, Engelberg D, Levitzki A: ROS, stress-activated kinases and stress signaling in cancer. EMBO Rep 2002;3:420-425.
[30] Desagher S, Martinou JC: Mitochondria as the central control point of apoptosis. Trends Cell Biol 2000;10:369-377.
[31] Gottlieb RA, Mitochondria: execution central. FEBS Lett 2000;482:6-12. [32] Xia Z, Lundgren B, Bergstrand A, DePierre JW, Nassberger L, Changes in the
generation of reactive oxygen species and in mitochondrial membrane potential during apoptosis induced by the antidepressants imipramine, clomipramine, and citalopram and the effects on these changes by Bcl-2 and Bcl-X(L). Biochem Pharmacol 1999;57:1199-1208.
[33] Kim SH, Hwang CI, Park WY, Lee JH, Song YS: GADD153 mediates celecoxib-induced apoptosis in cervical cancer cells. Carcinogenesis. 2006; [Epub ahead of print]
[34] Clarke AR, Gledhill S, Hooper ML, Bird CC, Wyllie AH: p53 dependence of early apoptotic and proliferative responses within the mouse intestinal epithelium following gamma-irradiation. Oncogene 1994;9:1767-1773.
[35] Miyashita T, Krajewski S, Krajewska M, Wang HG, Lin HK, Liebermann DA, Hoffman B, Reed JC: Tumor suppressor p53 is a regulator of bcl-2 and bax gene expression in vitro and in vivo. Oncogene 1994;9:1799-1805.
[36] Chang J, Xie M, Shah VR, Schneider MD, Entman ML, Wei L, Schwartz RJ. Activation of Rho-associated coiled-coil protein kinase 1 (ROCK-1) by caspase-3 cleavage plays an essential role in cardiac myocyte apoptosis. Proc Natl Acad Sci USA 2006; Sep 18; [Epub ahead of print]
FIGURE LEGENDS
Figure 1. The morphological changes and percentages of the viable A-549 cells after
treated with curcumin. The A-549 cells (2x105 cells/well; 12-well plates) were plated in 90% Ham’s F12K medium with 2 mM L-glutamine adjusted to contain 1.5 g/L sodium bicarbonate + 10% FBS with 30 µM curcumin for 6, 12, 24 and 48 h. The cells were photographed (panel A) by pharse-contrast microscope and collected by centrifugation and the viable cells were determined by trypan blue exclusion and flow cytometry (panel B) as described in Materials and Methods. Each point is mean ± S.D. of three experiments. *P<0.05
Figure 2. The cell cycle arrest and sub-G1 group of A-549 cells after treated with
curcumin. The A-549 cells were exposed to 0, 5, 10, 20 and 30 µM curcumin for 48 h and the cells were harvested and analyzed for cell cycle (panel A: the percent of cells in phase). The A-549 cells were treated with 30 µM curcumin for various time periods then were analyzed for the percent of cells in different cell pharse and apoptosis (panel B) as described in Materials and Methods. Data represents mean ± S.D. of three experiments. *P<0.05
Figure 3: Apoptosis of A-549 cells treated with curcumin were examined by using
DAPI staining. The A-549 cells were incubated with 30 µM curcumin for 6, 12, 24 and 48 h and apoptosis were determined by using DAPI staining then photographed by fluorescence microscope as described in Materials and Methods
Figure 4. Curcumin induced DNA damage in human lung carcinoma A-549 cells was
incubated with various concentrations of curcumin for 24 hours and DNA damage was determinated by Comet assay as described in Materials and Methods.
Figure 5. Effects of caspase-3 inhibitor (Ac-DEVD-CHO) on curcumin induced
caspase-3 activity, MMP and apoptosis in human lung cancer A-549 cells. The A-549 cells were treated with various concentrations of curcumin or were pre-treated with caspase-3 inhibitor (Ac-DEVD-CHO) for 3 hours then treated with 30 µM curcumin before cells were harvested for caspase-3 activity and the levels of MMP and apoptosis determinations as described in Materials and Methods (Panel A: caspase-3 activity; panel B: levels of MMP; panel C: apoptosis). Data represents mean ± S.D. of three experiments. *P<0.05.
Figure 6. Flow cytometric analysis of reactive oxygen species (ROS) in A-549 cells
with curcumin for various time periods. The A-549 cells (5x105 cells/ml) were treated with 30 µM curcumin for 0, 15, 30, 45, 60, 90 and 120 minutes to detect the changes of ROS. The zero concentration was defined as control. The percentage of cells that were stained by DCFH-DA dye, and the stained cells were determined by flow cytometry as described in the Materials and Methods section. *differs between curcumin and control. *P<0.05
Figure 7. Flow cytometric analysis of mitochondrial membrane potential (∆Ψm) in
A-549 cells with curcumin. The A-549 cells (5x105 cells/ml) were treated with 0, 5, 10, 20, 30 and 50 µM curcumin for 24 hours before cells were harvested for ∆Ψm
assay. The zero concentration was defined as control. The percentage of cells that were stained by DiOL6 dye, and the stained cells were determined by flow cytometry
as described in the Materials and Methods section. *differs between curcumin and control. *P<0.05
Figure 8. Flow cytometric analysis of Ca2+ concentration in A-549 cells with with 30
µM curcumin for various time periods. The A-549 cells (5x105 cells/ml) were treated with 30 µM curcumin for 0, 6, 12, 24 and 48 h to detect the changes of Ca2+
concentration. The zero concentration was defined as control. The percentage of cells that were stained by Indo-1/AM dye, and the stained cells were determined by flow cytometry as described in the Materials and Methods section. *differs between curcumin and control. *P<0.05
Figure 9. Effects of calcium antagonist BAPTA (Ca2+ chelator) on curcumin affect
the levels of Ca2+, MMP and apoptosis in A-549 cells. Flow cytometric analysis on the
effects of BAPTA on the levels of Ca2+, ∆Ψ
m and apoptosis in A-549 cells after treated
with curcumin. The A-549 cells were pre-treated with BAPTA for 3 h then treated with 30 µM curcumin before cells were harvested for Ca2+ (Panel A), ∆Ψ
m (Panel B)
and apoptosis (Panel C) determination as described in Materials and Methods. Data represents mean ± S.D. of three experiments. *P<0.05.
Figure 10. Representative Western blot showing changes on the levels of Wee1,
CHK1, Cdc25c, p53, p21, Cdc2, Cyclin B1,cspase-3 and -9, Bcl-2, COX-1 and -2, NF-kB, catalase, SOD(Zn and Cu), MMP-2 and -9 in A-549 cells after treated with curcumin. The A-549 cells (5x106/ml) were treated with 30 µM curcumin for 24 h before cytosolic fraction and total protein were prepared and determined as described in Materials and Methods. Then followed by the evaluation of the levels of associated protein levels (Panel A: Wee1, CHK1, Cdc25c, p53, p21, Cdc2, Cyclin B1;
panel B: caspase-3 and-9; panel C: Bcl-2, COX-1 and -2, NF-kB; panel D: catalase, SOD(Zn and Cu); panel E: MMP-2 and -9) and expressions were estimated by Western blotting as described in Materials and Methods.
Figure 11. Effects of curcumin on the levels of Gadd153 mRNA in A-549 cells. The
A-549 cells (5x105 cells/ml) were treated with 30 µM curcumin for 0, 1, 3, 6 and 12 h to extract total RNA then PCR were performed as showed in Materials and Methods. The levels of mRNA Gadd153 (Panel A) and ratio of mRNA/GAPDH (panel B) Data represents mean ± S.D. of three experiments. *P<0.05
Figure 12. Effects of curcumin on the cells migration of A-549. The A-549 cells
(2x104 cells/ml) were treated with or without 30 µM curcumin for 12 h then were photographed as showed in Materials and Methods.
Figure 13. Proposed model of curcumin mechanism of action for G2/M arrest and
apoptosis in A-549 cells. Curcumin induce p53, p21 expressions before leading to decrease Cdc2, cyclin B1 and promoted Wee1 and CHK2 leading to G2/M arrest. Curcumin increased the production of ROS led to ER stress and Ca2+ production and
decreased ∆Ψm levels that led to caspase-3 activity before causing apoptosis in A-549
Fig. 1 A.
Control 6 h
12 h 24 h 48 h B. * Viability (% ) 0 20 40 60 80 * * * *
Fig.2 A. B. G0/G1: 59.32% G2/M: 18.18% S: 22.5% Apop.: 0.01% Control G0/G1: 60.64% G2/M: 17.92% S: 21.44% Apop.: 0.17% 5 µM G0/G1: 55.9% G2/M: 23.41% S: 20.69% Apop.: 0.06% 10 µM G0/G1: 54.86% G2/M: 26.33% S: 18.82% Apop.: 0.6% 20 µM G0/G1: 35.42% G2/M: 37.52% S: 27.33% Apop.: 12.99% 30 µM G0/G1: 47.90% G2/M: 22.93% S: 36.55% Apop.: 11.09% 40 µM T i m e ( h ) C 6 1 2 2 4 4 8 Rat e (% ) 0 2 0 4 0 6 0 8 0 G0/G1 G2/M S - p h a s e A p o p to s is
Fig. 3
Control 6
h
12 h 24 h 48 h
Fig. 4
C 5 µM 10 µM 20 µM 30 µM 50 µM 30% H2O2Fig. 5 A Curcumin concentrations caspase-3 acti vit y ( % ) 0 10 20 30 40 50 60 70 0 10 30 40 ( µΜ ) * * * B M ito ch ond ri al m e m b ran e p o te nt ial (% ) 0 10 20 30 40 Curcumin - + - + Ac-DEVD-CHo - - + + C A popt ot ic cel ls ( % of to tal c ell s ) 0 2 4 6 8 10 12 14 16 18 curcumin - + - + Ac-DEVD-CHo - - + +
Fig. 6 Fig. 7 Fig. 8 Time (min) C 15 30 45 60 90 120 ROS (% ) 0 10 20 30 40 50 60 * * * * * * MMP 24h dose dependent Concentrations of Curcumin (µM) C 5 10 20 30 50 MMP (%) 0 20 40 60 80 100 * * * *
Calcium ion release time dependent
Time (h) C 6 12 24 48 Ra te (% ) 0 20 40 60 80 100 * * * *
Fig. 9 A C alc ium r eleas e ( % ) 0 20 40 60 80 100
Control BAPTA Curcumin BAPTA+Curcumin
B mitoc hond ria membra ne p ote ntia l (%) 0 20 40 60 80 100
Control BAPTA Curcumin BAPTA+Curcumin
C Ap opt os is (% ) 0 2 4 6 8 10 12 14 16
Fig. 10
A.
B.
caspase-3
caspase-9
β-actin
C 6 12 24 48 (h)
C 6 12 24 48 (h)
Wee1
CHK1
CDC25c
p53
p21
CDC2
Cyclin B
1β-actin
C 6 12 24 48 (h)
Bcl-2
COX-1
COX-2
β-actin
NF-kappaB
(p50)
C.
D.
E.
C 30 60 90 120 180 (min)
Catalase
SOD
(Zn/Cu)
C 6 12 24 48 (h)
MMP-9
MMP-2
β-actin
Fig. 11 A 0 1 3 6 12 (h) Gadd153 GAPDH B 0 1 2 R atio of Gad d15 3/ β-a ct in 0 10 50 100 200 Capsaicin concentrations (µΜ)
Fig. 12
Cell migration assay